Introduction
Breast cancer is the most frequently diagnosed
cancer and the leading cause of cancer death among females
worldwide, with an estimated 1.7 million cases and 521,900 deaths
in 2012 (1,2). One third of novel cancer diagnoses in
females are breast cancer and one in eight women will be diagnosed
with breast cancer, with a lifetime risk of mortality due to breast
cancer of 3.4% (3). Systematic
chemotherapy plays a critical role in treatment of breast cancer.
Doxorubicin (Dox) treatment is one of the established clinically
effective strategies for the treatment of breast cancer, and
primarily functions by inhibiting topoisomerases and intercalating
into the DNA double helix to interfere with DNA uncoiling, which
induces cell death (4). However,
the efficacy of Dox is limited by its drug resistance as well as
side-effects. Increasing research indicates that multidrug
resistance (MDR) contributes to the failure of chemotherapeutic
drugs as treatment (5). There are
at least two molecular pumps in tumor cell membranes, multi-drug
resistance-associated protein (MRP) and P-glycoprotein (P-gp), to
expel the antitumor drugs out of the cancer cells and attenuate the
drug effect. Some P-gp inhibitors used to reverse MDR were able to
enhance chemosensitivity in resistant cells but caused cytotoxicity
and a number of complications (6).
Therefore, finding key target molecules and novel therapeutic
strategies to overcome resistance and diminish the side-effects of
chemotherapeutic agents are the main goals of any ideal cancer
treatment protocol.
The protein phosphatase 2A (PP2A) is a key tumor
suppressor that regulates signaling pathways with a high relevance
in human cancer (7). Consistent
with its role as a tumor suppressor, PP2A plays a critical role in
the regulation of survival, cell cycle progression, and
differentiation by negatively regulating the PI3K/Akt pathway and
dephosphorylating and inactivating ERK and MEK1 family kinases
(8). Aberrant expression,
mutations, and somatic alterations of the PP2A scaffold and
regulatory subunits are frequently found in human breast, lung,
colon and skin cancers (9).
Therefore, reactivation of PP2A activity based on its tumor
suppressor properties is considered to be an attractive therapeutic
strategy for human cancer treatment (10,11).
Cancerous inhibitor of protein phosphatase 2A
(CIP2A), a human oncoprotein that stabilizes c-Myc by inhibiting
PP2A-mediated dephosphorylation of Myc at serine 62 (8). CIP2A promotes the proliferation and
aggressiveness of several cancer types including head and neck
squamous cell carcinoma, oral squamous cell carcinoma, esophageal
squamous cell carcinoma, colon, gastric, breast, prostate, tongue,
lung, cervical cancer and acute myeloid leukemia (8,12–15).
In 2011, Choi et al (16)
reported that CIP2A expression is associated with Dox resistance.
CIP2A has been found to involve in regulating MDR of cervical
adenocarcinoma upon chemotherapy by enhancing MDR gene encoded P-gp
expression through E2F1 (17).
Thus, effective and discerning CIP2A inhibitors would be beneficial
for adjuvant therapy to reduce the development of cancer resistance
to Dox and reactivation of PP2A activity in breast cancer.
Cucurbitacins are tetracyclic triterpene natural
products that are mainly found in the members of family
Cucurbitaceae. It has been used as a medicinal herb because it
exhibits different biological activities such as anti-diabetic,
anti-inflammatory, and anticancer activities against different
cancer cell lines (18).
Cucurbitacin B (CuB) is one of the most abundant forms of
cucurbitacins which is shown to inhibit the growth of numerous
human cancer cell lines such as breast, colon, leukemia, hepatic,
pancreatic and glioblastoma, and xenografts (19–21).
The effect of CuB on Dox-resistant breast cancer cells has not been
previously evaluated. The aim of the present study was to
investigate antitumor effects and possible mechanisms of CuB on Dox
resistant human breast cancer cells.
Materials and methods
Reagents
Cucurbitacin B (CuB) with a purity of up to 98% was
purchased from Shanghai Yuanye Bio-Technology Co., Ltd. (Shanghai,
China). CuB was dissolved in dimethyl sulf-oxide (DMSO;
Sigma-Aldrich, St. Louis, MO, USA) at a stock solution of 40 mM and
stored at −20°C. Doxorubicin (Dox) was purchased from
Sigma-Aldrich.
Cell culture
The human breast cancer cell line MCF-7 was obtained
from the American Type Culture Collection (ATCC; Manassas, VA,
USA). Human Dox-resistant breast cancer cell line MCF-7/Adriamycin
(MCF-7/Adr) was purchased from the Cell Bank of Chinese Academy of
Sciences (Shanghai, China). MCF-7 cells were maintained in
Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (FBS; HyClone
Laboratories, Inc., Logan, UT, USA) and antibiotics. MCF-7/Adr
cells were cultured in RPMI-1640 medium supplemented with 10% fetal
bovine serum. Additionally, 2 µg/ml Adriamycin was added
into MCF-7/Adr medium. All the cells were cultured in a humidified
atmosphere with 5% CO2 at 37°C.
Cytotoxic assay and cell viability
Cells were seeded into a 96-well plate and
pre-cultured for 24 h, then treated with CuB for 24 h. Cell
cytotoxicity was determined by MTT assay. The absorbance was
measured at 490 nm by automated micro-plated reader (BioTek
Instruments, Inc., Winooski, VT, USA), and the inhibition rate was
calculated as followed: Inhibition rate (%) = (average
A490 of the control group − average A490 of
the experimental group)/(average A490 of the control
group − average A490 of the blank group) × 100%. Cell
viability was estimated by trypan blue dye exclusion (22).
Soft-agar colony formation assay
Cells were suspended in 1 ml of RPMI-1640 containing
0.3% low-melting-point agarose (Amresco, Cleveland, OH, USA) and
10% FBS, and plated on a bottom layer containing 0.6% agarose and
10% FBS in 6-well plate in triplicate. After 2 weeks, plates were
stained with 0.2% gentian violet and the colonies were counted
under a light microscope (23).
Apoptosis determination by DAPI
staining
Approximately 2×105 cells/well of cells
in a 12-well plate was treated with CuB for 24 h. Then cells in
each treatment and control were stained by DAPI and examined and
photographed by fluorescence microscopy as described (24).
Western blot analysis
Cell pellets were lysed in RIPA buffer containing 50
mM Tris pH 8.0, 150 mM NaCl, 0.1% SDS, 0.5% deoxycholate, 1% NP-40,
1 mM DTT, 1 mM NaF, 1 mM sodium vanadate, 1 mM PMSF (Sigma-Aldrich)
and 1% protease inhibitors cocktail (Merck). Protein extracts were
quantitated and loaded on 8–12% sodium dodecyl sulfate
poly-acrylamide gel, electrophoresed and transferred to a PVDF
membrane (Millipore, Kenilworth, NJ, USA). The membrane was
incubated with primary antibody, washed, and incubated with
horseradish peroxidase (HRP)-conjugated secondary antibody
(Pierce). Detection was performed by using a chemiluminescent
western detection kit (Cell Signaling Technology, Danvers, MA, USA)
(25). The antibodies used were
anti-MRP1, anti-CIP2A, anti-Akt, anti-pAkt (Ser473) (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), anti-casp-9, anti-casp-3,
anti-PARP, anti-PP2A, anti-ERK1/2, anti-pERK1/2 (Thr202/Tyr204),
(Cell Signaling Technology), anti-P-gp (Abcam), and anti-GAPDH
(AB10016; Sangon Biotech, Co., Ltd., Shanghai, China).
PP2A activity assay
PP2A immunoprecipitation phosphatase assay kit
(Upstate-Millipore, Temecula, CA, USA) was used to measure
phosphate release as an index of phosphatase activity according to
the manufacturer's instructions. Briefly, 100 µg protein
isolated from cells was incubated with 4 µg anti-PP2A
monoclonal antibody overnight. A total of 40 µl of protein A
agarose beads were added and the mixture was incubated at 4°C for 2
h. Subsequently, the beads were collected and washed three times
with 700 µl of ice-cold TBS and one time with 500 µl
Ser/Thr assay buffer. The beads were further incubated with 750 mM
phosphopeptide in assay buffer for 10 min at 30°C with constant
agitation. A total of 100 µl of Malachite Green Phosphate
detection solution was added and the absorbance at 650 nm was
measured on a microplate reader (26).
Transfection of siRNA
Two siRNAs targeting CIP2A were designed and
synthesized by Shanghai GenePharma Co., Ltd., (Shanghai, China)
referred to as siRNA1 and siRNA2. The siRNA sequences were as
follows: 5′-CUGUGGUUGUGUU UGCACUTT-3′ (CIP2A siRNA1),
5′-ACCAUUGAUAUCCUUAGAATT-3′ (CIP2A siRNA2),
5′-UUCUCCGAACGUGUCACGUTT-3′ [negative control (NC) siRNA].
Using Lipofectamine 2000 (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer's protocol, MCF-7/Adr cells were
transfected with 100 nM siRNA. In addition, 48 h after
transfection, the cells were then harvested for western blot
analysis and cell viability.
Statistical analysis
All experiments were repeated at least three times
and the data are presented as the mean ± SD unless noted otherwise.
Differences between data groups were evaluated for significance
using Student's t-test of unpaired data or one way analysis of
variance and Bonferroni post-test. P-values <0.05 indicate
statistical significance.
Results
Chemical structure of CuB and
characterization of MCF-7 and MCF-7/Adr cells
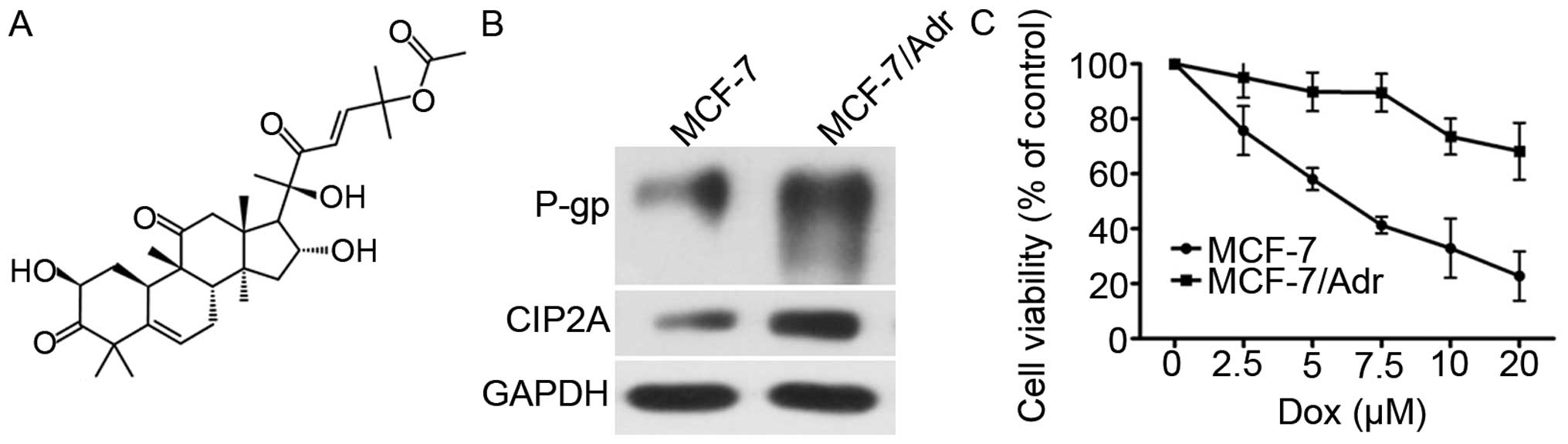
The chemical structure of CuB is shown in Fig. 1A. First, the P-gp and CIP2A
expression level was compared between MCF-7/Adr and MCF-7 cells by
western blot analysis, which confirmed P-gp and CIP2A
overexpression in MCF-7/Adr cells (Fig.
1B). MCF-7 and MCF-7/Adr cells were exposed to various
concentrations of Dox (2.5–80 µM) for 24 h. The half-maximal
inhibitory concentration (IC50) of Dox against MCF-7
cells is 6.2 µM, while IC50 of Dox against
MCF-7/Adr cells is 37.78 µM. As shown in Fig. 1C, the Dox cytotoxicity was higher in
MCF-7 cells than in MCF-7/Adr cells.
Effects of CuB on MCF-7 and MCF-7/Adr
breast cancer cells
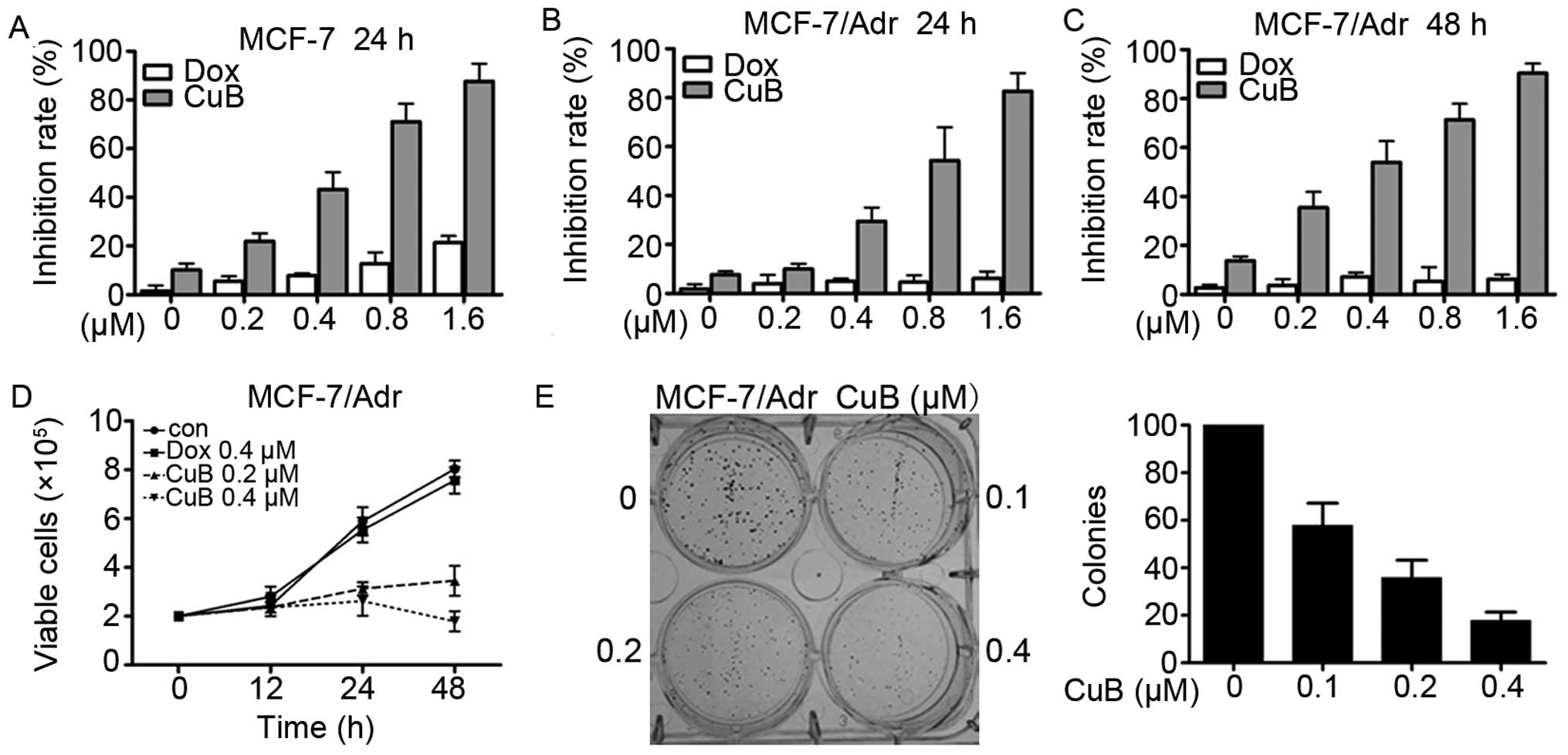
MCF-7 and MCF-7/Adr cells were seeded in 96-well
plates for 24 and 48 h and then treated with different
concentrations of CuB and Dox (Fig.
2A–C). After 24 or 48 h, the cell viability was evaluated by
the MTT assay according to the manual. Absorbance at 490 nm was
measured on an automated micro-plate reader. We found that CuB had
moderate cytotoxicity to MCF-7 and MCF-7/Adr cells with an
IC50 of 0.45 and 0.66 µM (Table I). By trypan blue exclusion assay,
we found that CuB rapidly reduced viable MCF-7/Adr (Fig. 2D) in a dose- and time-dependent
manner. We investigated CuB's effect on cell colony formation
activity, and the results showed that CuB significantly inhibited
the clonogenic ability of MCF-7/Adr (Fig. 2E). These results suggested that CuB
inhibited the anchorage-dependent (cell proliferation) and
anchorage-independent (colony formation) growth of MCF-7/Adr
cells.
 | Table IIC50s of CuB on breast
cancer cell lines. |
Table I
IC50s of CuB on breast
cancer cell lines.
| Cell lines | MCF-7 | MCF-7/Adr |
|---|
| IC50
(µM) | 0.45±0.03 | 0.66±0.05 |
CuB reverses the resistance of MCF-7/Adr
cells to Dox
MTT assay revealed a significant difference between
the growth-inhibiting effect of Dox on normal MCF-7 cells and on
Dox resistant MCF-7/Adr cells (Fig.
1C). The IC50 of MCF-7 cells was 6.2 µM, vs.
an IC50 of 37.78 µM for the MCF-7/Adr cells.
However, the Dox IC50 of MCF-7/Adr cell was 18.09
µM (0.05 µM CuB treatment) and 12.94 µM (0.1
µM CuB treatment) (Table
II). The resistance index (RI) of MCF-7/Adr parent group was
6.09, the MCF-7/Adr CuB 0.05 µM group 2.91 and the MCF-7/Adr
CuB 0.1 µM group 2.09. The RI calculation formula was used
to find the IC50 of resistant cells/IC50 of
sensitive cells. Following the treatment with CuB (0.05 and 0.1
µM), the IC50 of Dox to the MCF-7/Adr cells was
reduced from 37.78 to 18.09 and 12.94 µM by reversion fold
(RF) 2.09- and 2.92-fold. The RF calculation formula was used to
find the IC50 of Dox on MCF-7/Adr cells/IC50
of Dox (with 0.05 or 0.10 µM CuB) on MCF-7/Adr cells. Thus,
low doses of CuB (0.05 and 0.1 µM) can reverse Dox
resistance.
| Table IIReversing effect of CuB on MCF-7/Adr
cells. |
Table II
Reversing effect of CuB on MCF-7/Adr
cells.
| Groups, CuB
(µM)+Dox (µM) | Inhibition rate
(%) | IC50
(µM) | Resistance
index | Reversion fold |
|---|
| 0+5 | 9.51±0.53 | 37.78 | 6.09 | 1 |
| 0+10 | 14.84±2.94 | | | |
| 0+20 | 23.75±3.61 | | | |
| 0+30 | 52.03±5.36 | | | |
| 0+50 | 71.64±8.34 | | | |
| 0+80 | 83.35±3.89 | | | |
| 0.05+5 | 15.35±2.74 | 18.09 | 2.91 | 2.09 |
| 0.05+10 | 25.62±3.88 | | | |
| 0.05+20 | 36.48±4.92 | | | |
| 0.05+30 | 63.65±5.12 | | | |
| 0.05+50 | 86.46±3.71 | | | |
| 0.05+80 | 89.61±7.15 | | | |
| 0.1+5 | 20.17±3.58 | 12.94 | 2.09 | 2.92 |
| 0.1+10 | 35.72±5.46 | | | |
| 0.1+20 | 48.69±5.77 | | | |
| 0.1+30 | 75.47±4.83 | | | |
| 0.1+50 | 89.43±6.72 | | | |
| 0.1+80 | 98.51±6.02 | | | |
CuB induces apoptosis and influences the
expression of P-gp, MRP1 in MCF-7/Adr cells
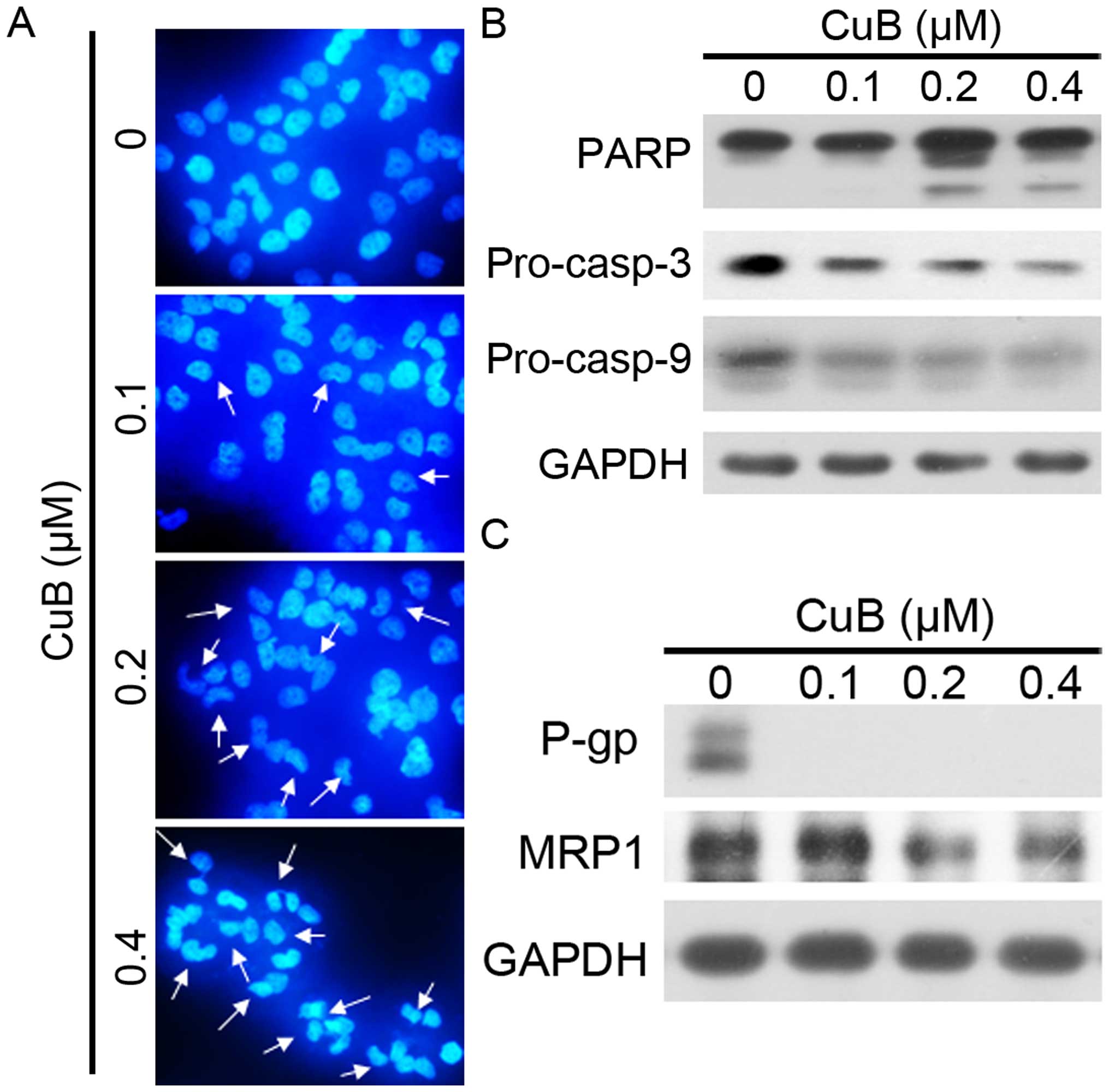
We next tested whether or not CuB induces apoptosis
of MCF-7/Adr cells. By an optical light microscope, we found some
dead MCF-7/Adr cells floating in the medium treated with CuB. The
cell death is reminiscent of the phenomena induced by apoptosis. We
investigated the nucleus morphological changes by DAPI staining. As
shown in Fig. 3A, we observed the
nuclear condensation and fragmentation with CuB treatment which are
typical changes in cell apoptosis. Furthermore, a western blot
analysis was used to detect the activation of the caspase-9
(casp-9) initiator caspase, caspase-3 (casp-3) effector caspase and
its substrate, poly(ADP-ribose) polymerase (PARP) (Fig. 3B). CuB was demonstrated to induce a
significant dose-dependent decrease in pro-casp-9, pro-casp-3 and
the cleavage of PARP, in MCF-7/Adr cells, indicating that CuB
induced caspase-dependent apoptosis. We detected expression levels
of MDR related factors P-gp and MRP1 by western blot analysis
(Fig. 3C). The results indicated
that P-gp, and MRP1 expression of MCF-7/Adr cells were
downregulated by treatment with increasing concentration of CuB. In
addition, the decreased expression of P-gp and MRP1 in MCF-7/Adr
cells may in part contribute to the reversal of MDR.
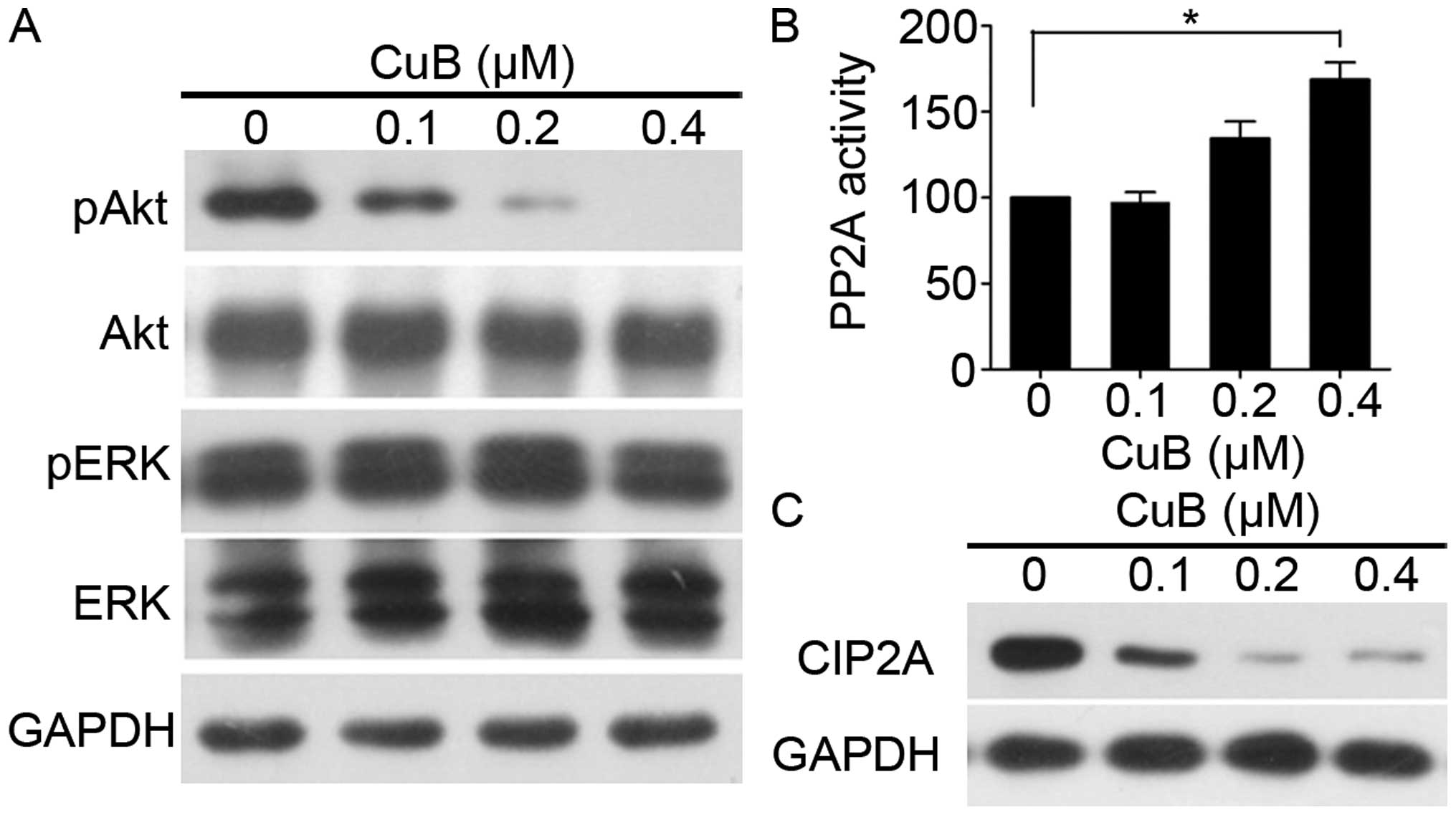
CuB targets CIP2A to reactivate protein
phosphatase 2A
Phosphorylated Akt (pAkt) is a cancer MDR locus
(27). We next investigated the
role of Akt in CuB-induced apoptosis in MCF-7/Adr cells. As shown
in Fig. 4A, CuB decreased Akt
phosphorylation in a dose-dependent manner. PP2A, one of the main
serine-threonine phosphatases, plays a critical role in the
regulation of cell cycle progression, survival, and differentiation
by negatively regulating the PI3K/Akt pathway and dephosphorylating
and inactivating MEK1 and ERK family kinases (26). We tested effects of CuB on PP2A
activity, and found that CuB upregulated PP2A activity (Fig. 4B). CIP2A is an oncogenic PP2A
inhibitor protein that is highly expressed in malignant cancers
(28). Furthermore, we tested
effects of CuB on CIP2A expression and found that the protein level
of CIP2A decreased (Fig. 4C),
indicating that CuB targeted CIP2A, at least in part, to reactivate
PP2A. Taken together, these data indicate that the CIP2A/PP2A/Akt
pathway may mediate the sensitizing effect of CuB.
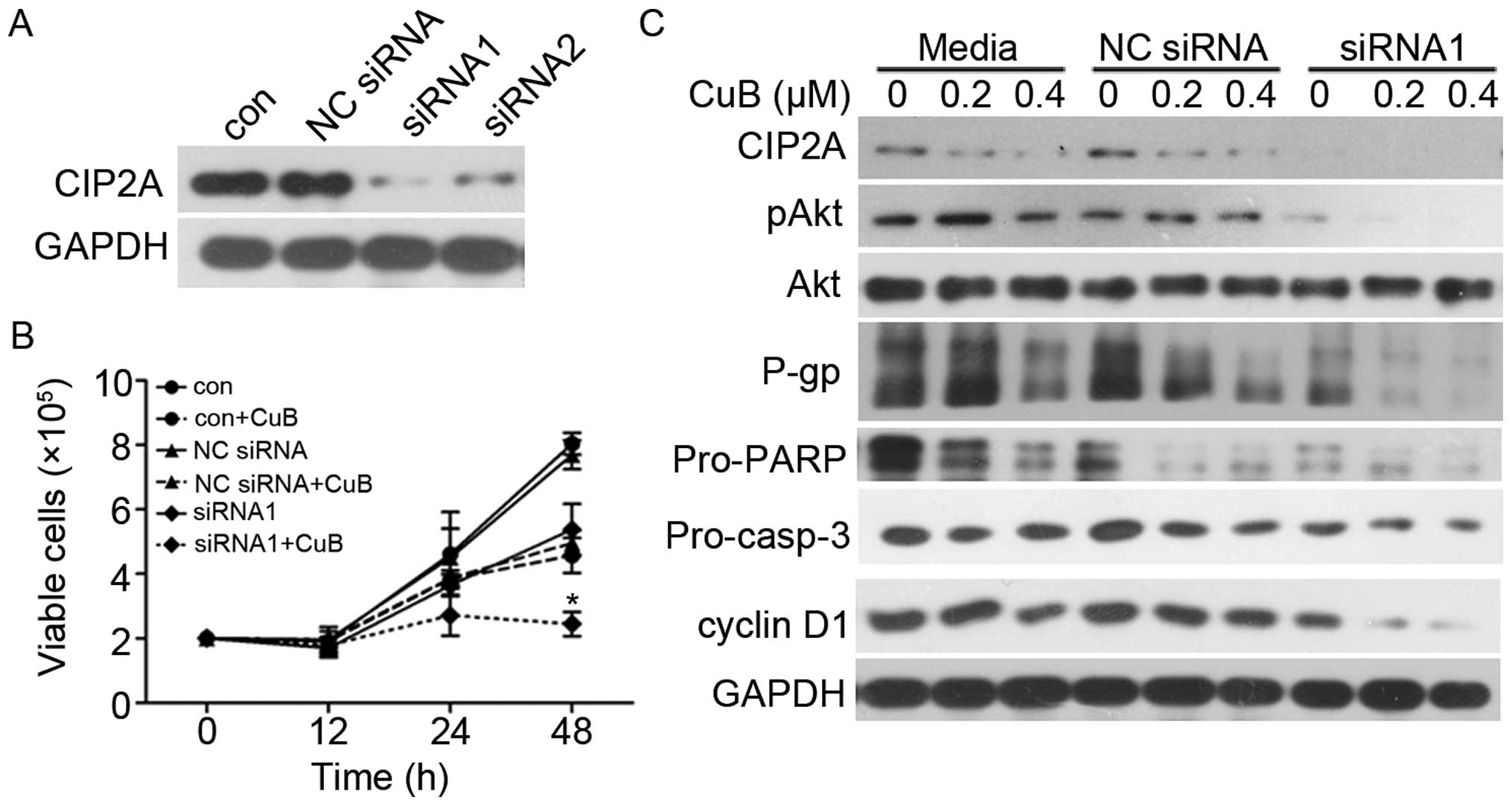
Silencing CIP2A enhances CuB-induced
growth inhibition and apoptosis in MCF-7/Adr
We next examined whether CIP2A knockdown would alter
cellular sensitivity to CuB. Two siRNAs targeting CIP2A were
synthesized and used. As shown in Fig.
5A, knockdown by both siRNA1 and siRNA2 markedly decreased
CIP2A protein in MCF-7/Adr cells. These data showed that CIP2A
siRNA1 and 2 were specific and efficient in reducing CIP2A
expression. To evaluate the role of CIP2A in CuB-induced
proliferation inhibition, MCF-7/Adr cells were transfected with
siRNA1 or siRNA2 targeting CIP2A, followed by CuB treatment. Cell
viability and western blot analysis were used to detect cell growth
and protein expression. CIP2A silencing enhanced CuB-induced growth
inhibition (Fig. 5B) and promoted
CuB-induced apoptotic effect (Fig.
5C). Notably, CIP2A depletion also enhanced CuB-induced MDR
inhibition. These data demonstrate that CIP2A plays a critical role
in CuB-reversed MCF-7/Adr multidrug resistance.
Discussion
Tumorigenesis and chemoresistance is tightly related
to malfunction of the cellular signaling pathways that control cell
proliferation, survival, or death. The malfunction contains
increased expression of ATP-dependent drug efflux pumps and
decreased influx, increased drug metabolism enzymes, impairment of
cell death pathways, enhancement of cell survival pathways,
alternation of drug metabolism, mutations in cell cycle pathways
and superior DNA repair (29). The
aims of the present study were to identify an effective MDR
reversing agent with fewer side-effects and to gain insight
regarding its molecular mechanism. Compounds from natural source
constitute an indispensable candidate drug library for
pharmacotherapy. CuB is a representative therapeutic agent for
anticancer activities. Recently, CuB was reported to enhance the
anticancer effects of cisplatin, gemcitabine, methotrexate,
docetaxel, and gemcitabine in laryngeal squamous, pancreatic, and
breast cancers and osteosarcoma (30). However, the relationship between CuB
and Dox-resistance in MCF-7/Adr cells has yet to be firmly
established. Inhibition of cell proliferation is an efficient
strategy in cancer therapy. In the present study, we first showed
that CuB inhibited MCF-7/Adr cell proliferation (Fig. 2B and C), cell viability (Fig. 2D) and soft-agar colony formation
(Fig. 2E).
We investigated the effect of the combination of CuB
and Dox on the MCF-7/Adr cells and identified that CuB in
combination with Dox had an improved effect compared with Dox or
CuB alone. Following the treatment with CuB (0.05 and 0.1
µM), the IC50 of Dox to the MCF-7/Adr cells was
significantly reduced from 37.78 to 18.09 and 12.94 µM by
2.09- and 2.92-fold (Table II).
The RI of of MCF-7/Adr parent group was 6.09, the MCF-7/Adr CuB
0.05 µM group 2.91 and the MCF-7/Adr CuB 0.1 µM group
2.09, respectively. Furthermore, the present study explored the
mechanisms of CuB in reversing Dox resistance.
Evading apoptosis is one of the hallmarks of drug
resistance, and targeting apoptosis has become a cancer therapeutic
strategy (31). Apoptosis is
accompanied by various morphological changes, including nuclear
condensation, apoptotic bodies, DNA fragmentation and cell surface
changes. Nuclear morphology in MCF-7/Adr cells was analyzed using
DAPI staining; we found changes in nucleus condensation, which are
typical characteristics of apoptosis (Fig. 3A). Therefore, CuB might have the
ability of induction of cell apoptosis. The mechanisms of apoptosis
involve two signaling pathways: the mitochondrial pathway
(intrinsic apoptotic pathway) and the cell death receptor pathway
(extrinsic apoptotic pathway) (32). The extrinsic and intrinsic apoptotic
pathways that ultimately lead to activation of effector caspases
(casp-3, -2 and -7) have been characterized (33,34).
The decrease of pro-casp-9, pro-casp-3, as well as the proteolysis
of PARP (Fig. 3B), indicate that
casp-3 is activated. Thus, CuB may trigger apoptosis by activating
the intrinsic apoptosis pathway which results in activation of
effector casp-9. Drug resistance is largely mediated through
overexpression of MDR, MRP, drug resistance protein, and proteasome
subunits, increases in antioxidant defenses, and TOP2 activity;
these results have been widely verified (35–37).
The present study identified that the treatment of CuB was able to
reverse the MDR of the MCF-7/Adr cells via the downregulation of
P-gp and MRP1 (Fig. 3C).
PP2A is a serine/threonine phosphatase that has a
critical role in regulating various cellular processes, including
signaling transduction, protein synthesis, cell cycle
determination, metabolism, apoptosis and stress response (38). Because loss of PP2A function has
been identified in various malignant diseases such as cancer of the
colon, liver, lung and breast, it has been suggested that PP2A
functions as a tumor suppressor and enhancing PP2A activity could
be an effective approach for anticancer treatment (39). In this study, we showed that by
inhibiting CIP2A, activity of PP2A was significantly enhanced and
expression of pAkt was downregulated in MCF-7/Adr cells (Fig. 4). Several cellular inhibitors of
PP2A have been identified, including SET (26) and CIP2A. CIP2A, originally named
KIAA1524 or P90, has been cloned from patients with HCC (8). CIP2A is associated with clinical
aggressiveness in human breast cancer and promotes the malignant
growth and metastasis of breast cancer cells. Induction of CIP2A is
often associated with chemoresistance in cancer cells, and the
inhibition of CIP2A in combination with chemotherapy may enhance
the efficacy of cancer treatment (40). We knocked down CIP2A expression in
MCF-7/Adr cells and found that CIP2A depletion significantly
promoted CuB induced apoptosis and reversed MDR (Fig. 5). Our data validated the mechanism
by which CuB-reversed MDR and induced cancer cell apoptosis in
MCF-7/Adr cells, that is, reversed MDR and induction of cancer cell
apoptosis by inhibiting CIP2A to reactivate PP2A and enhance
PP2A-dependent pAkt downregulation.
In conclusion, our results suggest that further
studies investigating the detailed molecular modification of the
PP2A/CIP2A signaling pathway by CuB and exploring its possible
application in other malignant diseases are warranted.
Acknowledgments
The present study was supported by grants from the
National Natural Sciences Foundation of China (grant no. 81400157);
the Natural Science Foundation of Hubei Provincial Department of
Education (grant no. Q20152106); the Faculty Development Grants
from Hubei University of Medicine (grant nos. 2014QDJZR08,
2015QDJZR16); the Foundation for Innovative Research Team of Hubei
University of Medicine (grant no. 2014CXX05); the Key Discipline
Project of Hubei University of Medicine and the National Training
Program of Innovation and Entrepreneurship for Undergraduates
(grant no. 201610929001).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhu XF, Li W, Ma JY, Shao N, Zhang YJ, Liu
RM, Wu WB, Lin Y and Wang SM: Knockdown of heme oxygenase-1
promotes apoptosis and autophagy and enhances the cytotoxicity of
doxorubicin in breast cancer cells. Oncol Lett. 10:2974–2980.
2015.
|
|
4
|
Zheng Y, Lv X, Wang X, Wang B, Shao X,
Huang Y, Shi L, Chen Z, Huang J and Huang P: MiR-181b promotes
chemoresistance in breast cancer by regulating Bim expression.
Oncol Rep. 35:683–690. 2016.
|
|
5
|
Faneyte IF, Kristel PM, Maliepaard M,
Scheffer GL, Scheper RJ, Schellens JH and van de Vijver MJ:
Expression of the breast cancer resistance protein in breast
cancer. Clin Cancer Res. 8:1068–1074. 2002.PubMed/NCBI
|
|
6
|
Jeon YJ, Kim JH, Shin JI, Jeong M, Cho J
and Lee K: Salubrinal-mediated upregulation of eIF2α
phosphorylation increases doxorubicin sensitivity in MCF-7/ADR
cells. Mol Cells. 39:129–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Perrotti D and Neviani P: Protein
phosphatase 2A: A target for anticancer therapy. Lancet Oncol.
14:e229–e238. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Junttila MR, Puustinen P, Niemelä M, Ahola
R, Arnold H, Böttzauw T, Alaaho R, Nielsen C, Ivaska J, Taya Y, et
al: CIP2A inhibits PP2A in human malignancies. Cell. 130:51–62.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seshacharyulu P, Pandey P, Datta K and
Batra SK: Phosphatase: PP2A structural importance, regulation and
its aberrant expression in cancer. Cancer Lett. 335:9–18. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schönthal AH: Role of serine/threonine
protein phosphatase 2A in cancer. Cancer Lett. 170:1–13. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv P, Wang Y, Ma J, Wang Z, Li JL, Hong
CS, Zhuang Z and Zeng YX: Inhibition of protein phosphatase 2A with
a small molecule LB100 radiosensitizes nasopharyngeal carcinoma
xenografts by inducing mitotic catastrophe and blocking DNA damage
repair. Oncotarget. 5:7512–7524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li W, Ge Z, Liu C, Liu Z, Björkholm M, Jia
J and Xu D: CIP2A is overexpressed in gastric cancer and its
depletion leads to impaired clonogenicity, senescence, or
differentiation of tumor cells. Clin Cancer Res. 14:3722–3728.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Z, Ma L, Wen ZS, Hu Z, Wu FQ, Li W,
Liu J and Zhou GB: Cancerous inhibitor of PP2A is targeted by
natural compound celastrol for degradation in non-small-cell lung
cancer. Carcinogenesis. 35:905–914. 2014. View Article : Google Scholar
|
|
14
|
Ren J, Li W, Yan L, Jiao W, Tian S, Li D,
Tang Y, Gu G, Liu H and Xu Z: Expression of CIP2A in renal cell
carcinomas correlates with tumour invasion, metastasis and
patients' survival. Br J Cancer. 105:1905–1911. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu CY, Shiau CW, Kuo HY, Huang HP, Chen
MH, Tzeng CH and Chen KF: Cancerous inhibitor of protein
phosphatase 2A determines bortezomib-induced apoptosis in leukemia
cells. Haematologica. 98:729–738. 2013. View Article : Google Scholar :
|
|
16
|
Choi YA, Park JS, Park MY, Oh KS, Lee MS,
Lim JS, Kim KI, Kim KY, Kwon J, Yoon Y, et al: Increase in CIP2A
expression is associated with doxorubicin resistance. FEBS Lett.
585:755–760. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu J, Wang M, Zhang X, Wang Q, Qi M, Hu
J, Zhou Z, Zhang C, Zhang W, Zhao W, et al: CIP2A is associated
with multidrug resistance in cervical adenocarcinoma by a
P-glycoprotein pathway. Tumour Biol. 7:2673–2682. 2015.
|
|
18
|
Lai GM, Ozols RF, Young RC and Hamilton
TC: Effect of glutathione on DNA repair in cisplatin-resistant
human ovarian cancer cell lines. J Natl Cancer Inst. 81:535–539.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
El-Senduny FF, Badria FA, El-Waseef AM,
Chauhan SC and Halaweish F: Approach for chemosensitization of
cisplatin-resistant ovarian cancer by cucurbitacin B. Tumour Biol.
37:685–698. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chan KT, Meng FY, Li Q, Ho CY, Lam TS, To
Y, Lee WH, Li M, Chu KH and Toh M: Cucurbitacin B induces apoptosis
and S phase cell cycle arrest in BEL-7402 human hepatocellular
carcinoma cells and is effective via oral administration. Cancer
Lett. 294:118–124. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chan KT, Li K, Liu SL, Chu KH, Toh M and
Xie WD: Cucurbitacin B inhibits STAT3 and the Raf/MEK/ERK pathway
in leukemia cell line K562. Cancer Lett. 289:46–52. 2010.
View Article : Google Scholar
|
|
22
|
Liu Y, Cao W, Zhang B, Liu YQ, Wang ZY, Wu
YP, Yu XJ, Zhang XD, Ming PH, Zhou GB, et al: The natural compound
magnolol inhibits invasion and exhibits potential in human breast
cancer therapy. Sci Rep. 3:30982013.PubMed/NCBI
|
|
23
|
Cao W, Liu Y, Zhang R, Zhang B, Wang T,
Zhu X, Mei L, Chen H, Zhang H, Ming P, et al: Homoharringtonine
induces apoptosis and inhibits STAT3 via IL-6/JAK1/STAT3 signal
pathway in Gefitinib-resistant lung cancer cells. Sci Rep.
5:84772015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chou CC, Yang JS, Lu HF, Ip SW, Lo C, Wu
CC, Lin JP, Tang NY, Chung JG, Chou MJ, et al: Quercetin-mediated
cell cycle arrest and apoptosis involving activation of a caspase
cascade through the mitochondrial pathway in human breast cancer
MCF-7 cells. Arch Pharm Res. 33:1181–1191. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Y, Dong Y, Zhang B and Cheng YX: Small
compound 6-O-angeloylplenolin induces caspase-dependent apoptosis
in human multiple myeloma cells. Oncol Lett. 6:556–558.
2013.PubMed/NCBI
|
|
26
|
Liu H, Gu Y, Wang H, Yin J, Zheng G, Zhang
Z, Lu M, Wang C and He Z: Overexpression of PP2A inhibitor SET
oncoprotein is associated with tumor progression and poor prognosis
in human non-small cell lung cancer. Oncotarget. 6:14913–14925.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Radisavljevic Z: AKT as locus of cancer
multidrug resistance and fragility. J Cell Physiol. 228:671–674.
2013. View Article : Google Scholar
|
|
28
|
Rincón R, Cristóbal I, Zazo S, Arpí O,
Menéndez S, Manso R, Lluch A, Eroles P, Rovira A, Albanell J, et
al: PP2A inhibition determines poor outcome and doxorubicin
resistance in early breast cancer and its activation shows
promising therapeutic effects. Oncotarget. 6:4299–4314. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Holohan C, Van Schaeybroeck S, Longley DB
and Johnston PG: Cancer drug resistance: An evolving paradigm. Nat
Rev Cancer. 13:714–726. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo J, Zhao W, Hao W, Ren G, Lu J and Chen
X: Cucurbitacin B induces DNA damage, G2/M phase arrest, and
apoptosis mediated by reactive oxygen species (ROS) in leukemia
K562 cells. Anticancer Agents Med Chem. 14:1146–1153. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Marsden VS, O'Connor L, O'Reilly LA, Silke
J, Metcalf D, Ekert PG, Huang DC, Cecconi F, Kuida K, Tomaselli KJ,
et al: Apoptosis initiated by Bcl-2-regulated caspase activation
independently of the cytochrome c/Apaf-1/caspase-9 apoptosome.
Nature. 419:634–637. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nicholson DW: Caspase structure,
proteolytic substrates, and function during apoptotic cell death.
Cell Death Differ. 6:1028–1042. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Johnstone RW, Ruefli AA and Lowe SW:
Apoptosis: A link between cancer genetics and chemotherapy. Cell.
108:153–164. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Alcantara LM, Kim J, Moraes CB, Franco CH,
Franzoi KD, Lee S, Freitas-Junior LH and Ayong LS:
Chemosensitization potential of P-glycoprotein inhibitors in
malaria parasites. Exp Parasitol. 134:235–243. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luo L, Sun YJ, Yang L, Huang S and Wu YJ:
Avermectin induces P-glycoprotein expression in S2 cells via the
calcium/calmodulin/NF-κB pathway. Chem Biol Interact. 203:430–439.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ying L, Zu-An Z, Qing-Hua L, Qing-Yan K,
Lei L, Tao C and Yong-Ping W: RAD001 can reverse drug resistance of
SGC7901/DDP cells. Tumour Biol. 35:9171–9177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cristóbal I, Rincón R, Manso R, Caramés C,
Zazo S, Madoz-Gúrpide J, Rojo F and García-Foncillas J:
Deregulation of the PP2A inhibitor SET shows promising therapeutic
implications and determines poor clinical outcome in patients with
metastatic colorectal cancer. Clin Cancer Res. 21:347–356. 2015.
View Article : Google Scholar
|
|
39
|
Yu HC, Hung MH, Chen YL, Chu PY, Wang CY,
Chao TT, Liu CY, Shiau CW and Chen KF: Erlotinib derivative
inhibits hepatocellular carcinoma by targeting CIP2A to reactivate
protein phosphatase 2A. Cell Death Dis. 5:e13592014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Côme C, Laine A, Chanrion M, Edgren H,
Mattila E, Liu X, Jonkers J, Ivaska J, Isola J, Darbon JM, et al:
CIP2A is associated with human breast cancer aggressivity. Clin
Cancer Res. 15:5092–5100. 2009. View Article : Google Scholar : PubMed/NCBI
|