Introduction
Head and neck cancer represents the sixth most
common human cancer and includes cancers of the lip, oral cavity
(mouth), nasal cavity (inside the nose), paranasal sinuses,
pharynx, and larynx (1).
Approximately 90% of head and neck cancers are squamous cell
carcinoma (HNSCC), which is the leading malignancy in humans
(2). An estimated 300,400 new cases
and 145,400 deaths from oral cavity cancers occurred in 2012
worldwide (3). The development of
HNSCC is a result of multiple genetic and epigenetic alterations,
including the activation of oncogenes and the loss of function of
tumor-suppressor genes (4).
Nevertheless, the precise mechanism of the tumorigenesis in HNSCC
remains largely unknown.
p12, a cyclin-dependent kinase 2 (CDK2)-associating
protein 1 (p12CDK2-AP1), was initially identified as a
cancer-related gene in a hamster oral cancer model (5). The human p12CDK2-AP1 gene
locates on chromosome 12q24 and encodes a 115-amino acid protein
(6). In human oral squamous cell
carcinoma (OSCC), p12CDK2-AP1 functions as a tumor
suppressor by negatively regulating the activity of CDK2, which
plays a crucial role in mediating cell cycle progression (7,8). In
addition to CDK2, p12CDK2-AP1 has been found to interact
with other proteins including DNA polymerase α/primase (9) and its homologous protein p14 (10). Protein interaction plays a pivotal
role in the control of various essential biological processes;
thus, it is important to identify and validate other
p12CDK2-AP1 interacting proteins to understand its role
in cell cycle regulation and tumorigenesis.
Computational approaches have been developed for the
large-scale prediction of protein-protein interaction based on
protein sequence, structure and evolutionary relationships in
complete genomes in attempts to unravel the global protein
interactome (11-13). In the present study, a multiple and
pairwise kernel support vector machine was employed to predict the
interactive proteins of p12CDK2-AP1. CD82 was identified
to be a protein with high binding affinity to
p12CDK2-AP1. We further found that the interaction
between p12CDK2-AP1 and CD82 played a synergistic role
in suppressing the tumorigenesis and development of OSCC,
suggesting that the interference of p12CDK2-AP1/CD82
interaction may provide a valuable strategy for managing OSCC.
Materials and methods
Prediction of protein-protein
interactions (PPIs)
To predict the interacting proteins of
p12CDK2-AP1, we employed a multiple and pairwise kernel
support vector machine as previously described (11,14).
With these approaches, the one- and two-dimensional structures as
well as the eigenvector of p12CDK2-AP1 were simulated. A
total of 10 proteins from the Swiss-Port database and the potential
PPIs were predicted based on the decision template.
Reagents
PCR primers were synthesized by Sangon Biotech
(Shanghai, China). The pIRES2-EGFP vector was purchased from YouBio
(Changsha, China). Dulbecco's modified Eagle's medium (DMEM) and
Opti-MEM® medium (low serum culture medium) were
obtained from Sigma-Aldrich (St. Louis, MO, USA) and Invitrogen
(Carlsbad, CA, USA), respectively. Fetal bovine serum (FBS) was
purchased from Gibco (New York, NY, USA). Six-well Transwell
polycarbonate filters (8-µm pore size) were purchased from
Millipore (Boston, MA, USA). Matrigel was obtained from BD
Biosciences (New York, NY, USA). Annexin V-FITC was purchased from
BLKW Biotechnology (Beijing, China).
Construction of the recombinant
plasmids
The full-length coding sequences of
p12CDK2-AP1 and CD82 were amplified from the human brain
library using PCR. The sequences of the PCR primers were as
follows: p12CDK2-AP1 forward,
5′-ATGTCTTACAAACCGAACTTGG-3′ and reverse,
3′-GGATCTGGCATTCCGTTCCG-5′; and CD82 forward,
5′-ATGGGCTCAGCCTGTATCAAAG-3′ and reverse,
3′-GTACTTGGGGACCTTGCTGTA-5′. The amplified products were inserted
into the pIRES2-EGFP vector to obtain
pIRES2-EGFP-p12CDK2-AP1 and pIRES2-EGFP-CD82 plasmids.
The sequences of the plasmids were confirmed via direct DNA
sequencing.
Cell culture and transfection
Both human embryonic kidney 293T cells and human
oral squamous cell carcinoma OSCC-15 cells were purchased from the
American Type Culture Collection (ATCC; Manassas, VA, USA). The
cells were cultured in DMEM supplemented with 10% FBS at 37°C in a
humidified atmosphere with 5% CO2. The cultured cells
were passaged every two days with fresh culture medium. 293T and
OSCC-15 cells were transfected with the
pIRES2-EGFP-p12CDK2-AP1, pIRES2-EGFP-CD82, or
pIRES2-EGFP negative control (NC) plasmids using Lipofectamine™
2000 transfection reagent in Opti-MEM® medium according
to the manufacturer's recommendations (Invitrogen). The protein
levels of p12CDK2-AP1 and CD82 were examined using
western blot analysis at 0, 24, 48, and 72 h after transfection.
Fluorescence was examined under a fluorescence microscope
(BX51-32FB3F01; Olympus, Japan).
Western blot analysis
Total proteins were extracted from cultured cells
using RIPA lysis buffer (1% Nonidet P-40, 0.5% sodium deoxycholate
and 0.1% SDS in 1X phosphate buffer solution) containing protease
inhibitor (2 µg/ml aprotinin, 2 µg/ml leupeptin and 1
M PMSF) for 30 min on ice. After centrifuging, the supernatant was
resuspended in buffer containing 1% SDS and 1% dithiothreitol and
heated at 100°C for 5 min. Equal amounts of proteins were
electrophoresed using 10% SDS-PAGE and then transferred onto
nitrocellulose membranes (#88018; Pierce, USA). After being blocked
with 5% non-fat dry milk in Tris-buffered saline and Tween-20 (10
mM Tris-HCl, pH 8.0, 100 mM NaCl and 0.05% Tween, TBS-T), the
membrane was incubated at 4°C overnight with the mouse polyclonal
anti-CDKA1 (1:200, ab167256), mouse monoclonal anti-CD82 (1:200,
ab140238) (both from Abcam, Cambridge, UK) or mouse polyclonal
anti-GAPDH (1:800; Santa Cruz Biotechnology, Santa Cruz, CA, USA)
primary antibodies. After incubation, the membrane was washed twice
with TBS-T for 15 min and incubated with horseradish peroxidase
(HRP)-conjugated goat anti-mouse secondary antibody (Abcam). After
TBS-T washing, the immunoreactivities were visualized using an
enhanced chemiluminescence (ECL) kit according to the
manufacturer's instructions (Abcam). The relative optical density
of the bands of interest was analyzed using Image-Pro Plus 6.0
software.
Co-immunoprecipitation (Co-IP)
OSCC-15 cells were lysed in IP cell lysis buffer
(0.6 ml 1 M Tris-HCl pH 6.8, 5 ml 50% glycerinum, 2 ml 10% SDS, 0.5
ml 2-mercaptoethanol, 1 ml 1% bromophenol blue, 0.9 ml distilled
water) (P0013; Beyotime, Shanghai, China) containing protease
inhibitors (Roche). The whole cell lysates were incubated with an
antibody together with 20 µl Protein A Plus agarose beads
(#20333; Pierce) overnight at 4°C. The immunoprecipitates were
washed three times with IP cell lysis buffer, resuspended in 20
µl of 2X SDS loading buffer, heated at 100°C for 10 min and
then loaded for SDS-PAGE electrophoresis and western blotting using
a goat anti-mouse IgG/HRP.
MTT assay
To determine the proliferation capability of the
cells, an MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide] colorimetric assay was performed. Non-transfected 293T
cells (OSCC-15) and cells transfected with NC plasmid (OSCC-15+NC),
p12CDK2-AP1 (OSCC-15+p12CDK2-AP1), CD82
(OSCC-15+CD82) or p12CDK2-AP1 plus CD82
(OSCC-15+p12CDK2-AP1+CD82) were used for the MTT assay.
The cells were seeded in 96-well plates at a density of
1×104 cells/well and maintained at 37°C in a humidified
atmosphere with 5% CO2 overnight. After incubation, 20
µl of MTT reagent (5 mg/ml) was added to each well followed
by another 4 h of incubation. The MTT solution was removed, and
dimethyl sulfoxide (DMSO) was added to each well to dissolve the
metabolic product. The absorbance at 570 nm was recorded. The
relative proliferation rate of the cells was calculated according
to the following equation: Relative proliferation rate of the cells
= (Absorbance value of the sample/Absorbance value of the control)
× 100%. Data were calculated from three independent
experiments.
Determination of the cell invasive
capability
Cell invasive ability was determined using a 6-well
Transwell system. In brief, 50 mg/l of Matrigel was diluted in
serum-free medium to a final concentration of 3.9 µg/μl.
After that, 60–80 µl of Matrigel was added onto the
polycarbonate membrane and air-dried for 24 h. The transfected
OSCC-15 cells were trypsinized, washed in PBS and suspended in 0.1%
serum-containing medium. Approximately 5×104 cells were
loaded into the upper chamber wells and incubated at 37°C in 5%
CO2 for 18 h. After incubation, the cells that had
traversed the filter were fixed with methanol, washed in PBS,
stained with hexamethyl pararosaniline and counted. The average
number of invaded cells was calculated from 3 independent
experiments.
Evaluation of cell apoptosis
OSCC-15 cells were collected at 72 h after
transfection. Non-transfected cells were used as a control. The
cells were washed twice with cold PBS, and approximately
1×105–1×106 cells were re-suspended in 100
µl solution containing 5% Annexin V-FITC and incubated at
37°C for 15 min in the dark. After PI labeling, the cells were
analyzed using flow cytometry (BD Biosciences). The experiments
were performed in triplicate. The early apoptosis rate (%) and the
overall cell apoptosis rate (%) (early plus late) were
calculated.
Tumor formation in nude mice
To understand the influences of
p12CDK2-AP1 and CD82 on in vivo tumor formation,
non-transfected cells (OSCC-15), cells transfected with the NC
plasmid (OSCC-15+NC), or the p12CDK2-AP1 plus CD82
(OSCC-15+p12CDK2-AP1+CD82) plasmids were subcutaneously
inoculated into nude mice. Sixty-three 5-week-old BALB/c nude mice
weighing 21±3 g were obtained from the Animal Laboratory of the
Fourth Military Medical University. The animals were randomly
divided into three groups with 21 mice in each group for tumor cell
inoculation. On day 1, 100 µl cell suspension
(1×106 cells) was inoculated subcutaneously into the
right rear flank of the nude mice. The maximum diameter (a, mm) and
vertical short diameter (b, mm) of the tumor mass were measured
using a precision caliper every three days initiated on day 7 and
ending on day 25. The tumor size was calculated using the following
formula: V (mm3) = a × b2 × 0.52. The animals
were sacrificed 30 days after tumor cell injection, and the tumor
masses were weighted. The tumor inhibitory rate was calculated
using the following formula: Tumor inhibition rate (%) =
[(V1 – V2)/V1] × 100%, in which
V1 represents the tumor size of the OSCC-15 group and
V2 represents the tumor size of the OSCC-15+NC or
OSCC-15+p12CDK2-AP1+CD82 group. The experiment
procedures involving animals and their care were conducted in
conformity with the NIH guidelines (NIH Pub. no. 85–23, revised
1996) and were approved by the Animal Care and Use Committee of the
Fourth Military Medical University.
Statistical analysis
The statistical analysis was conducted using
SPSS19.0 software. The expression of p12CDK2-AP1 and
CD82 in oral cancer was compared using the Chi-square test. Data
are presented as the means ± SD. P<0.05 was considered to
indicate a significant difference.
Results
Prediction of p12CDK2-AP1 and
CD82 interaction
Using a multiple and pairwise kernel support vector
machine, 10 proteins (including Ig α-1 chain C region, α-amylase 1
precursor, α-2 chain C region, CD82, fructose-bisphosphate aldolase
A, Kruppel-like factor 6, sulfatase-modifying factor 2, inhibitor
of CDK interacting with cyclin A1, insulinoma-associated protein 1
and 60S ribosomal protein L13) were predicted to have high
potential in binding with p12CDK2-AP1 (Table I). Among these proteins, tumor
suppressor gene CD82 was chosen for further analysis. The sequences
of p12CDK2-AP1 and CD82 were input into the pairwise
kernel support vector machine, and the two proteins were predicted
as an interacting protein-protein pair (Fig. 1).
 | Table IProtein interactors of
p12CDK2-AP1 identified by computational decision
templates. |
Table I
Protein interactors of
p12CDK2-AP1 identified by computational decision
templates.
| Swiss-Prot Protein
Sequence Database | Protein |
|---|
| p01876 | Ig α-1 chain C
region |
| p01877 | Ig α-2 chain C
region |
| p27701 | CD82 antigen |
| p04075 |
Fructose-bisphosphate aldolase A |
| q99612 | Krueppel-like
factor 6 |
| q9by67 | α-amylase 1
precursor |
| q8nbj7 | Sulfatase-modifying
factor 2 |
| q0vd86 | Inhibitor of CDK
interacting with cyclin A1 |
| q01101 |
Insulinoma-associated protein 1 |
| a8k4c8 | 60S ribosomal
protein L13 |
Identification of the protein-protein
binding between p12CDK2-AP1 and CD82
Co-immunoprecipitation was conducted to validate the
direct binding between p12CDK2-AP1 and CD82. For this
purpose, pIRES2-EGFP-p12CDK2-AP1 and pIRES2-EGFP-CD82
recombinant plasmids were constructed and overexpressed in the 293T
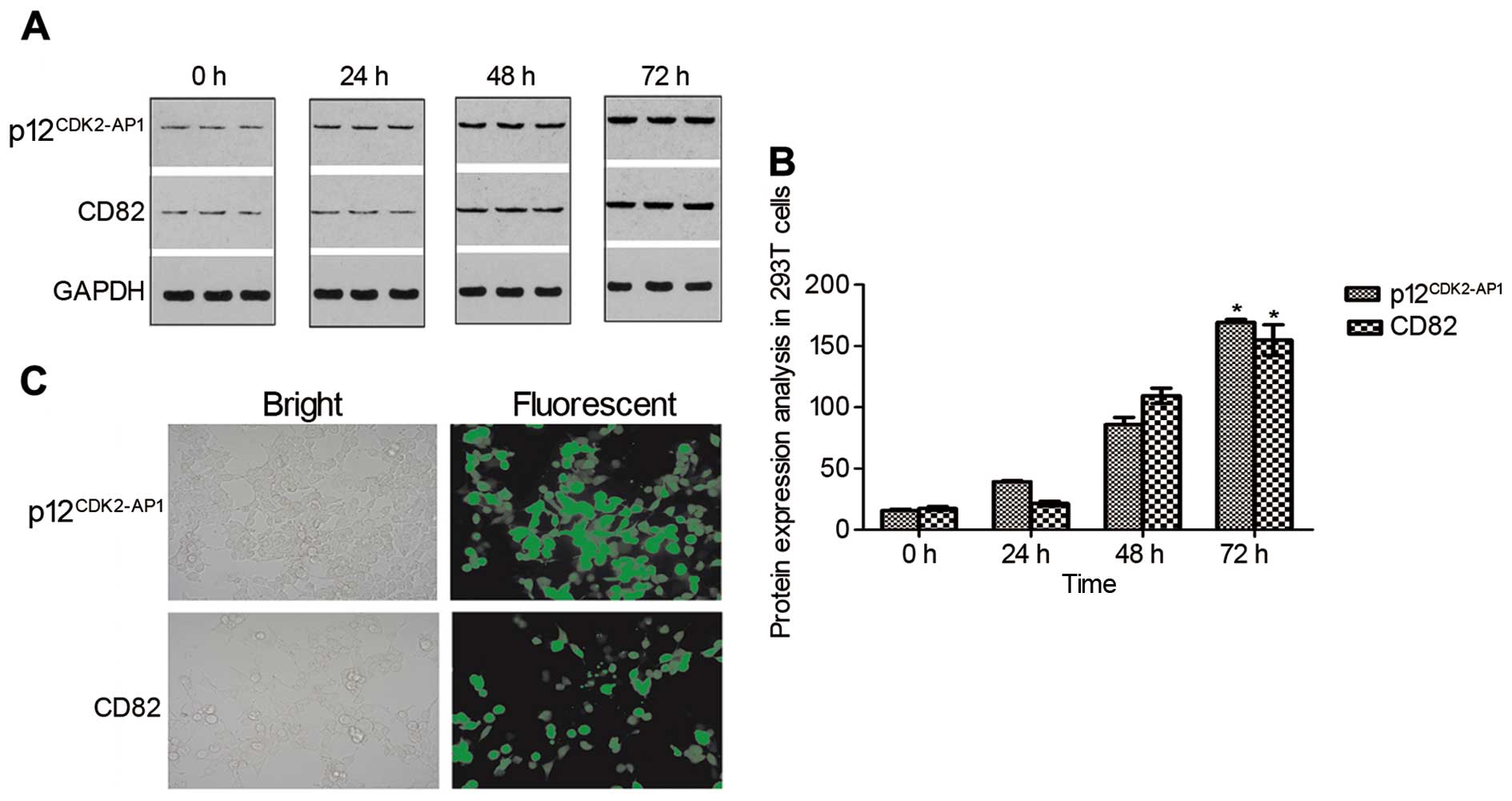
cells, respectively. Western blot analysis demonstrated that
p12CDK2-AP1 and CD82 were weakly expressed in the 293T
cells under baseline conditions but were gradually increased after
transfection in a time-dependent manner (Fig. 2A). A statistical analysis revealed a
significant upregulation of p12CDK2-AP1 and CD82 at 72 h
after transfection (P<0.05 compared with the control) (Fig. 2B). Additionally, bright GFP signals
were detected in over 90% of the 293T cells at 72 h after
transfection (Fig. 2C), indicating
the successful overexpression of p12CDK2-AP1 and CD82 in
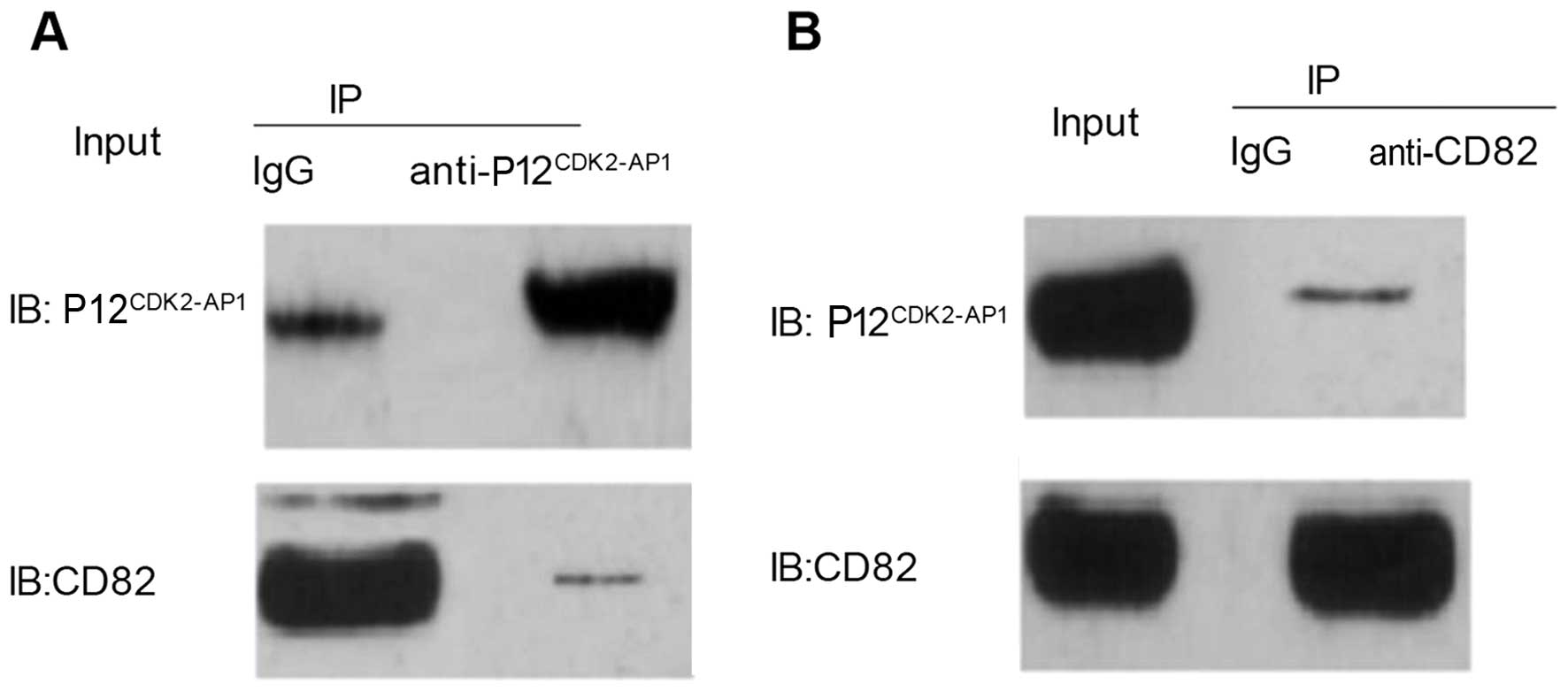
the 293T cells. A co-immunoprecipitation analysis indicated that
CD82 could be specifically pulled down by the
anti-p12CDK2-AP1 antibody (Fig. 3A); conversely,
p12CDK2-AP1 could be specifically pulled down by the
anti-CD82 antibody (Fig. 3B). These
results demonstrated that p12CDK2-AP1 and CD82 could be
reciprocally pulled down in OSCC-15 cells, indicating a direct
physical interaction between p12CDK2-AP1 and CD82.
Overexpression of p12CDK2-AP1
and CD82 suppresses the proliferation of OSCC-15 cells
To elucidate the potential influences of
p12CDK2-AP1 and CD82 on the growth, proliferation and
survival of human oral squamous cell carcinoma cells,
p12CDK2-AP1 and CD82 were overexpressed in the OSCC-15
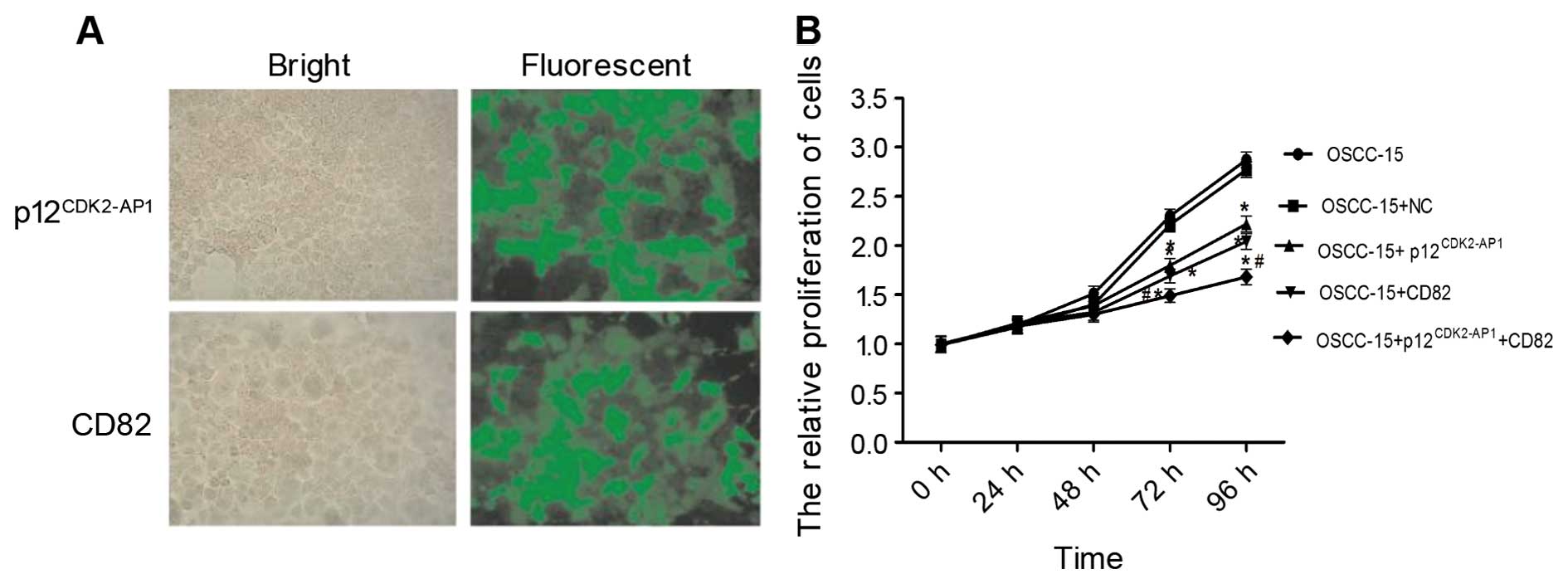
cells via plasmid transfection. At 72 h after transfection, over
90% of the OSCC-15 cells expressed GFP signals (Fig. 4A). The MTT analysis showed that
transfection with the negative control (NC) vector did not
significantly affect cell proliferation (P>0.05 compared with
the control) (Fig. 4B). The
overexpression of either p12CDK2-AP1 or CD82
significantly suppressed cell proliferation at 72 and 96 h after
transfection (P<0.05 compared with the NC). Notably, the
combined transfection of pIRES2-EGFP-p12CDK2-AP1 and
pIRES2-EGFP-CD82 yielded an enhanced growth inhibition (inhibition
rate of 1.68%) compared with single plasmid transfection (P<0.05
compared with the NC), suggesting that p12CDK2-AP1 and
CD82 may synergistically inhibit the proliferation of OSCC-15
cells. These data suggest that the overexpression of
p12CDK2-AP1 and CD82 inhibits the proliferation of
OSCC-15 cells.
Overexpression of
p12CDK2-AP1and CD82 inhibits the invasion of OSCC-15
cells
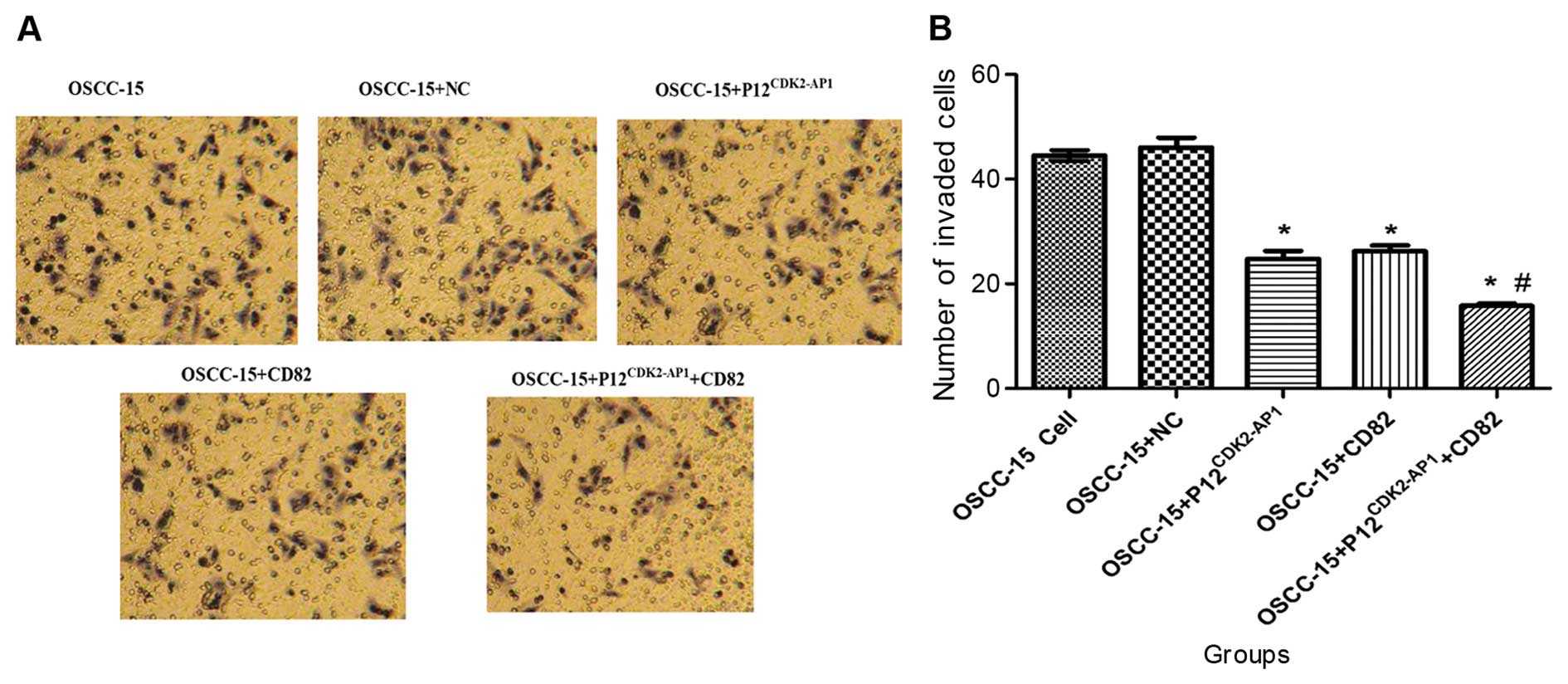
We next investigated the effects of
p12CDK2-AP1 and CD82 overexpression on the invasion
capability of OSCC-15 cells in vitro using a Transwell
system. There were no significant differences in the number of
invaded cells between the non-transfected control and the cells
transfected with the NC plasmid (P>0.05, Fig. 5). However, the overexpression of
either p12CDK2-AP1 or CD82 significantly inhibited cell
invasion (P<0.05 compared with the NC). Additionally, combined
transfection of pIRES2-EGFP-p12CDK2-AP1 and
pIRES2-EGFP-CD82 led to a more efficient inhibition of cell
invasion compared with single plasmid transfection (P<0.05;
Fig. 5), indicating a synergistic
inhibitory effect by p12CDK2-AP1 and CD82 on cell
invasion. This evidence indicates that the overexpression of
p12CDK2-AP1 and CD82 can suppress the in vitro
invasion capability of OSCC-15 cells.
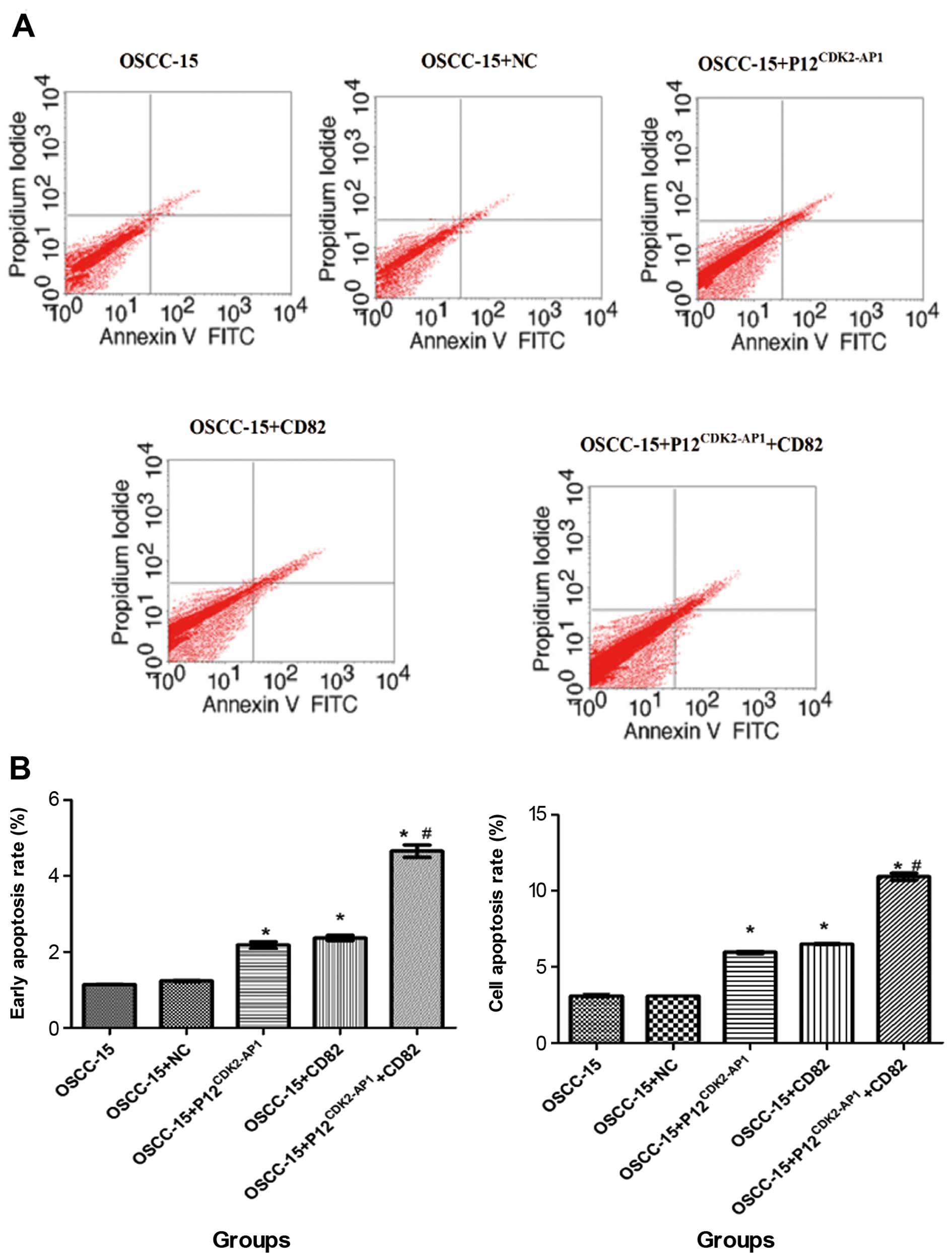
Overexpression of p12CDK2-AP1
and CD82 induces apoptosis in OSCC-15 cells
To assess the potential role of
p12CDK2-AP1 and CD82 on cell apoptosis, OSCC-15 cells
transfected with NC, pIRES2-EGFP-p12CDK2-AP1 and
pIRES2-EGFP-CD82 alone or in combination were probed with Annexin
V-FITC and PI, followed by flow cytometry analysis. The
transfection of NC did not significantly influence cell apoptosis
compared with that in the non-transfected control cells (P>0.05)
(Fig. 6). However, the
overexpression of p12CDK2-AP1 or CD82 significantly
increased the percentage of both the early and overall apoptotic
cells (P<0.05 compared with the NC). Moreover, the combined
transfection of both plasmids was significantly more efficient in
promoting cell apoptosis compared with single plasmid transfection
(P<0.01), suggesting a synergistic effect for
p12CDK2-AP1 and CD82 on cell apoptosis. These data
indicate that the overexpression of p12CDK2-AP1 and CD82
induces apoptosis in OSCC-15 cells.
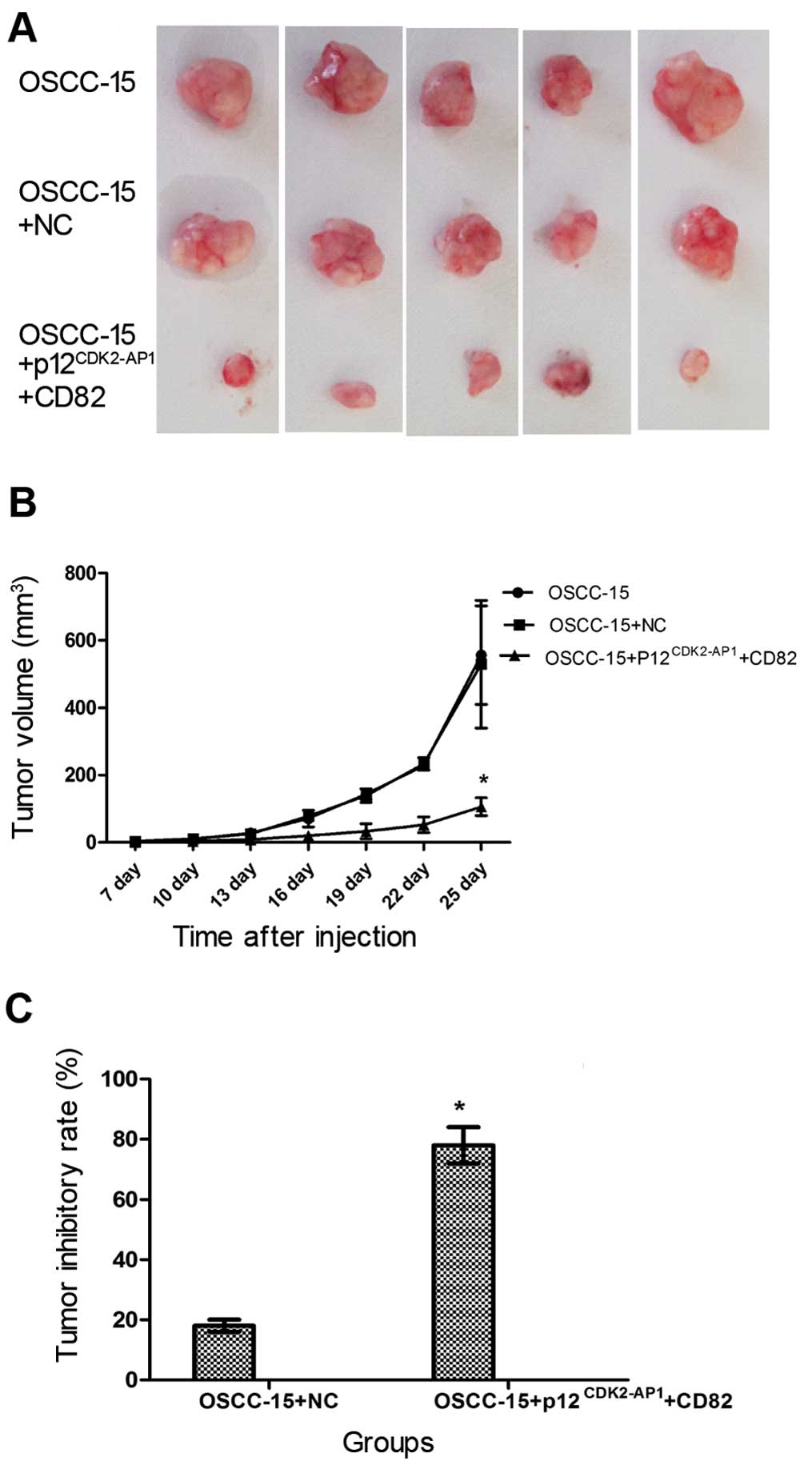
Overexpression of p12CDK2-AP1
and CD82 inhibits the in vivo growth of OSCC-15 cells in tumor
mouse xenografts
Finally, we investigated the combined overexpression
of p12CDK2-AP1 and CD82 on the growth of OSCC-15 cells
in vivo. To this end, OSCC-15 cells transfected with NC or
those transfected with pIRES2-EGFP-p12CDK2-AP1 plus
pIRES2-EGFP-CD82 were subcutaneously injected into nude mice.
Twenty-five days after inoculation, no significant differences were
detected regarding the size of the tumor mass between the
non-transfected OSCC-15 cell group and the OSCC-15 cells
transfected with NC (Fig. 7A).
However, the combined overexpression of p12CDK2-AP1 and
CD82 substantially reduced the tumor size. Additionally, the
combined overexpression of p12CDK2-AP1 and CD82
significantly reduced the tumor volume and yielded a significant
tumor inhibitory rate at 25 days after inoculation compared with
the NC group (Fig. 7B and C;
P<0.05). These findings suggest that the combined overexpression
of p12CDK2-AP1 and CD82 inhibits the growth of OSCC-15
cells in tumor mouse xenografts.
Discussion
In this study, using a multiple and pairwise kernel
support vector machine, 10 proteins were predicted to have high
binding potential with p12CDK2-AP1. Among these
proteins, CD82 was further verified to interact with
p12CDK2-AP1 via co-immunoprecipitation. Moreover, the
overexpression of either p12CDK2-AP1 or CD82
significantly inhibited proliferation and invasion but induced
apoptosis in the OSCC-15 cells. However, the combined
overexpression of p12CDK2-AP1 and CD82 was significantly
more efficient in promoting cell apoptosis and inhibiting
proliferation and invasion compared with single overexpression.
Importantly, the combined overexpression of p12CDK2-AP1
and CD82 inhibited the in vivo growth of OSCC-15 cells in
tumor mouse xenografts. Our data suggest that
p12CDK2-AP1 and CD82 may synergistically inhibit the
growth of OSCC-15 cells.
The dysregulated cell cycle control system is a
crucial process during oral carcinogenesis (15). Cyclin-dependent kinases (CDKs) are
widely accepted to control cell cycle progression.
p12CDK2-AP1 has been found to regulate cell cycle
progression and cell proliferation by negatively mediating the
kinase activities of CDK2 (16). A
growing body of evidence supports the notion that
p12CDK2-AP1 inhibits the progression of several human
cancers, such as breast cancer (17), gastric cancer (18), and esophageal squamous cell
carcinoma (19). The silencing of
p12CDK2-AP1 expression was reported to accelerate the
proliferation of human skin keratinocyte (HaCaT) cells (20), whereas the upregulation of
p12CDK2-AP1 expression led to the reduced proliferation
and invasion of HaCaT cells (21).
Consistent with these findings, in the present study, the
overexpression of p12CDK2-AP1 in OSCC-15 cells
significantly suppressed cell proliferation and invasion and
promoted cell apoptosis, indicating the inhibitory role of
p12CDK2-AP1 in OSCC.
Increasing evidence indicates that
p12CDK2-AP1 interacts with other proteins such as DNA
polymerase α/primase (9) and the
homologous protein p14 (10). Using
a yeast two-hybrid system, our previous study also identified a
novel unnamed protein product (UPP) that interacts with
p12CDK2-AP1 and inhibits the proliferation of 293T and
HeLa cells (22). Using
computational approaches, CD82 was predicted to be an interacting
protein of p12CDK2-AP1, which was further confirmed via
Co-IP analysis. CD82, also known as KAI1, was originally identified
in human prostate carcinoma and mapped to human chromosome 11p11.2
(23). CD82 has been shown to be a
metastasis suppressor in many types of human cancers, including
prostate (24), bladder (25), breast (26), colon (27), pancreas (28), and lung (29,30)
cancers. Additionally, the mutation or downregulation of CD82
protein has been detected in esophageal (31,32)
and oral (33–35) cancers. In non-small cell lung
carcinoma h1299 cells, CD82 was found to negatively regulate cell
motility and migration (36,37).
However, the regulatory role of CD82 in OSCC cells is not yet fully
understood. In this study, we found that overexpression of CD82
yielded an inhibitory effect on OSCC-15 cells similar to that of
p12CDK2-AP1, implying that CD82 may also act as a tumor
suppressor in OSCC.
More importantly, our results revealed that when
p12CDK2-AP1 and CD82 were co-expressed in OSCC-15 cells,
a synergistic tumor inhibition was detected both in vitro
and in vivo. Compared with single plasmid transfection, the
combined transfection of p12CDK2-AP1 with CD82
demonstrated more efficiency in suppressing cell growth and
invasion and promoting cell apoptosis. Moreover, the simultaneous
overexpression of p12CDK2-AP1 and CD82 significantly
inhibited the growth of OSCC-15 cells in tumor mouse xenografts.
These data indicate that p12CDK2-AP1 and CD82 serve as
tumor suppressors and may act synergistically in suppressing the
tumorigenesis of OSCC.
In summary, our results demonstrated that
p12CDK2-AP1 interacted with CD82 and negatively
regulated the growth and survival of OSCC-15 cells. Moreover, our
findings suggest that targeting the interaction between
p12CDK2-AP1 and CD82 may represent a potential strategy
for the development of treatment strategies for oral cancers.
Future studies will be needed to explore the molecular mechanisms
in modulating the p12CDK2-AP1-CD82 interaction.
Acknowledgments
This study was supported by the Project of
Scientific and Technological Research Development of Shaanxi
Province (grant no. 2014K12-16) and in part by the National Natural
Science Foundation of China (grant no. 81072230).
References
|
1
|
Marcu LG and Yeoh E: A review of risk
factors and genetic alterations in head and neck carcinogenesis and
implications for current and future approaches to treatment. J
Cancer Res Clin Oncol. 135:1303–1314. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen C, Méndez E, Houck J, Fan W,
Lohavanichbutr P, Doody D, Yueh B, Futran ND, Upton M, Farwell DG,
et al: Gene expression profiling identifies genes predictive of
oral squamous cell carcinoma. Cancer Epidemiol Biomarkers Prev.
17:2152–2162. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Todd R, McBride J, Tsuji T, Donoff RB,
Nagai M, Chou MY, Chiang T and Wong DT: Deleted in oral cancer-1
(doc-1), a novel oral tumor suppressor gene. FASEB J. 9:1362–1370.
1995.PubMed/NCBI
|
|
6
|
Tsuji T, Duh FM, Latif F, Popescu NC,
Zimonjic DB, McBride J, Matsuo K, Ohyama H, Todd R, Nagata E, et
al: Cloning, mapping, expression, function, and mutation analyses
of the human ortholog of the hamster putative tumor suppressor gene
Doc-1. J Biol Chem. 273:6704–6709. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shintani S, Mihara M, Terakado N, Nakahara
Y, Matsumura T, Kohno Y, Ohyama H, McBride J, Kent R, Todd R, et
al: Reduction of p12DOC-1 expression is a negative
prognostic indicator in patients with surgically resected oral
squamous cell carcinoma. Clin Cancer Res. 7:2776–2782.
2001.PubMed/NCBI
|
|
8
|
Kohno Y, Patel V, Kim Y, Tsuji T, Chin BR,
Sun M, Bruce Donoff R, Kent R, Wong D and Todd R: Apoptosis,
proliferation and p12(doc-1) profiles in normal, dysplastic and
malignant squamous epithelium of the Syrian hamster cheek pouch
model. Oral Oncol. 38:274–280. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matsuo K, Shintani S, Tsuji T, Nagata E,
Lerman M, McBride J, Nakahara Y, Ohyama H, Todd R and Wong DT:
p12(DOC-1), a growth suppressor, associates with DNA polymerase
alpha/primase. FASEB J. 14:1318–1324. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Buajeeb W, Zhang X, Ohyama H, Han D,
Surarit R, Kim Y and Wong DT: Interaction of the CDK2-associated
protein-1, p12(DOC-1/CDK2AP1), with its homolog, p14(DOC-1R).
Biochem Biophys Res Commun. 315:998–1003. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang SW, Hao LY and Zhang TH: Prediction
of protein-protein interaction with pairwise kernel support vector
machine. Int J Mol Sci. 15:3220–3233. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hue M, Riffle M, Vert JP and Noble WS:
Large-scale prediction of protein-protein interactions from
structures. BMC Bioinformatics. 11:1442010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Craig RA and Liao L: Improving protein
protein interaction prediction based on phylogenetic information
using a least-squares support vector machine. Ann NY Acad Sci.
1115:154–167. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen W, Zhang SW, Cheng YM and Pan Q:
Prediction of protein-protein interaction types using the decision
templates based on multiple classier fusion. Math Comput Model.
52:2075–2084. 2010. View Article : Google Scholar
|
|
15
|
Todd R, Hinds PW, Munger K, Rustgi AK,
Opitz OG, Suliman Y and Wong DT: Cell cycle dysregulation in oral
cancer. Crit Rev Oral Biol Med. 13:51–61. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shintani S, Ohyama H, Zhang X, McBride J,
Matsuo K, Tsuji T, Hu MG, Hu G, Kohno Y, Lerman M, et al:
p12(DOC-1) is a novel cyclin-dependent kinase 2-associated protein.
Mol Cell Biol. 20:6300–6307. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou W, Guan X, Wang L, Liao Y and Huang
J: p12(CDK2-AP1) inhibits breast cancer cell proliferation and in
vivo tumor growth. J Cancer Res Clin Oncol. 138:2085–2093. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi MG, Sohn TS, Park SB, Paik YH, Noh
JH, Kim KM, Park CK and Kim S: Decreased expression of p12 is
associated with more advanced tumor invasion in human gastric
cancer tissues. Eur Surg Res. 42:223–229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hiyoshi Y, Watanabe M, Hirashima K,
Karashima R, Sato N, Imamura Y, Nagai Y, Yoshida N, Toyama E,
Hayashi N, et al: p12CDK2-AP1 is associated with tumor
progression and a poor prognosis in esophageal squamous cell
carcinoma. Oncol Rep. 22:35–39. 2009.PubMed/NCBI
|
|
20
|
Sun M, Zheng J, Xue H, Jiang Y, Li C, Li
J, Jin W, Shen M, Yang X and Ni Q: Silencing P12CDK2AP1
with a lentivirus promotes HaCaT cell proliferation. Mol Med Rep.
7:471–475. 2013.
|
|
21
|
Zheng J, Xue H, Wang T, Jiang Y, Liu B, Li
J, Liu Y, Wang W, Zhang B and Sun M: miR-21 downregulates the tumor
suppressor P12 CDK2AP1 and stimulates cell proliferation and
invasion. J Cell Biochem. 112:872–880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu L, Yang X, Ni Q, Xiao Z, Zhao Y, Han
J, Sun M and Chen B: Interaction between p12CDK2AP1 and
a novel unnamed protein product inhibits cell proliferation by
regulating the cell cycle. Mol Med Rep. 9:156–162. 2014.
|
|
23
|
Dong JT, Lamb PW, Rinker-Schaeffer CW,
Vukanovic J, Ichikawa T, Isaacs JT and Barrett JC: KAI1, a
metastasis suppressor gene for prostate cancer on human chromosome
11p11.2. Science. 268:884–886. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang Y, Cheng Y, Martinka M, Ong CJ and Li
G: Prognostic significance of KAI1/CD82 in human melanoma and its
role in cell migration and invasion through the regulation of ING4.
Carcinogenesis. 35:86–95. 2014. View Article : Google Scholar
|
|
25
|
Rowe A and Jackson P: Expression of
KITENIN, a KAI1/CD82 binding protein and metastasis enhancer, in
bladder cancer cell lines: Relationship to KAI1/CD82 levels and
invasive behaviour. Oncol Rep. 16:1267–1272. 2006.PubMed/NCBI
|
|
26
|
Malik FA, Sanders AJ, Kayani MA and Jiang
WG: Effect of expressional alteration of KAI1 on breast cancer cell
growth, adhesion, migration and invasion. Cancer Genomics
Proteomics. 6:205–213. 2009.PubMed/NCBI
|
|
27
|
Pal'tseva EM, Samofalova OIu, Gorbacheva
IuV and Tsar'kov PV: Expression of some biomarkers in primary colon
adenocarcinomas and their lymph node metastases. Arkh Patol.
74:12–18. 2012.In Russian. PubMed/NCBI
|
|
28
|
Friess H, Guo XZ, Tempia-Caliera AA,
Fukuda A, Martignoni ME, Zimmermann A, Korc M and Büchler MW:
Differential expression of metastasis-associated genes in papilla
of vater and pancreatic cancer correlates with disease stage. J
Clin Oncol. 19:2422–2432. 2001.PubMed/NCBI
|
|
29
|
Choi UJ, Jee BK, Lim Y and Lee KH:
KAI1/CD82 decreases Rac1 expression and cell proliferation through
PI3K/Akt/mTOR pathway in H1299 lung carcinoma cells. Cell Biochem
Funct. 27:40–47. 2009. View Article : Google Scholar
|
|
30
|
Shiwu WU, Lan Y, Wenqing S, Lei Z and
Yisheng T: Expression and clinical significance of CD82/KAI1 and
E-cadherin in non-small cell lung cancer. Arch Iran Med.
15:707–712. 2012.PubMed/NCBI
|
|
31
|
Liu FS, Dong JT, Chen JT, Hsieh YT, Ho ES,
Hung MJ, Lu CH and Chiou LC: KAI1 metastasis suppressor protein is
down-regulated during the progression of human endometrial cancer.
Clin Cancer Res. 9:1393–1398. 2003.PubMed/NCBI
|
|
32
|
Miyazaki T, Kato H, Shitara Y, Yoshikawa
M, Tajima K, Masuda N, Shouji H, Tsukada K, Nakajima T and Kuwano
H: Mutation and expression of the metastasis suppressor gene KAI1
in esophageal squamous cell carcinoma. Cancer. 89:955–962. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Imai Y, Sasaki T, Shinagawa Y, Akimoto K
and Fujibayashi T: Expression of metastasis suppressor gene
(KAI1/CD82) in oral squamous cell carcinoma and its
clinico-pathological significance. Oral Oncol. 38:557–561. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Farhadieh RD, Smee R, Ow K, Yang JL,
Russell PJ, Crouch R, Jackson P and Jacobson IV: Down-regulation of
KAI1/CD82 protein expression in oral cancer correlates with reduced
disease free survival and overall patient survival. Cancer Lett.
213:91–98. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kawasaki G, Yoshitomi I, Yanamoto S,
Yamada S, Mizuno A and Umeda M: Expression of thymidylate synthase
and dihydropyrimidine dehydrogenase in primary oral squamous cell
carcinoma and corresponding metastases in cervical lymph nodes:
Association with the metastasis suppressor CD82. Anticancer Res.
31:3521–3526. 2011.PubMed/NCBI
|
|
36
|
Abe M, Sugiura T, Takahashi M, Ishii K,
Shimoda M and Shirasuna K: A novel function of CD82/KAI-1 on
E-cadherin-mediated homophilic cellular adhesion of cancer cells.
Cancer Lett. 266:163–170. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Takahashi M, Sugiura T, Abe M, Ishii K and
Shirasuna K: Regulation of c-Met signaling by the tetraspanin
KAI-1/CD82 affects cancer cell migration. Int J Cancer.
121:1919–1929. 2007. View Article : Google Scholar : PubMed/NCBI
|