Introduction
Lung cancer is the leading cause of cancer
associated death all over the world (1). Tumor metastasis as the major cause for
high mortality of lung cancer has attracted a great deal of
research interest. During the past decades, many metastasis related
processes had been identified, such as genetic mutation (2), angiogenesis (3), drug resistance (4), inflammation (5), cancer stem cells (6) and epithelial-mesenchymal transition
(EMT) (7). In addition,
understanding the molecular mechanism of metastatic lung cancer has
greatly improved patient survival.
ITGB3 is a receptor of various proteins such as
fibronectin, laminin, matrix metalloproteinase-2, osteomodulin and
vitronectin (8). Highly elevated
expression of ITGB3 has been observed in various kinds of malignant
carcinoma. In leukemia, ITGB3 plays a crucial role in
leukemogenesis, making ITGB3 be a potential therapeutic target in
AML (9). Another study confirmed
that ITGB3 was an important regulator in reactive oxygene species
induced migration and invasion of colorectal cancer cells (10). In breast cancer, mRNA profile array
revealed that several angiogenesis related proteins including ITGB3
were significantly upregulated in metastatic tumor cells (11). Moreover, a recent study proved that
let-7c, which is downregulated in lung cancer, inhibited the
migration and invasion of lung cancer cells by targeting ITGB3
(12).
MicroRNAs (miRNAs) are non-coding RNA,
20–22-nucleotides in length and can regulate gene expression by
repressing gene translation or promoting mRNA degradation (13). Disordered expression of miRNAs play
an important role in tumor initiation, progression and recurrence.
Some miRNAs such as miR-92b (14),
miR-9 (15), miR-224 (16) and miR-183 (17) act as oncogenes and drive tumor
metastasis in lung cancer. While, miR-101 (18), miR-133a (19) and miR-141 (20) which are downregulated in lung cancer
could significantly suppresses tumor metastasis. Besides, some
exosomal miRNAs play a key role in lung carcinogenesis, making
miRNAs new tumor biomarkers (21).
miR-338 was firstly identified as an intronic miRNA
of its host gene AATK and was also functionally antagonistic to
AATK (22). Recently, miR-338 was
proved downregulated in hepatocellular carcinoma (23), oral carcinoma and esophageal
squamous cell carcinoma (24). In
gastric cancer, overexpressing miR-338 inhibited cell proliferation
and promoted apoptosis (25).
Restoring miR-338 level in hepatocarcinoma sensitized cells to
sorafenib (26). Although one study
had proved that miR-338 was able to inhibit colorectal cancer cell
invasion and migration by targeting Smoothened (27). The mechanism of miR-338 in tumor
metastasis is still unclear. In the present study, we assessed the
expression of miR-338 in 115 pairs of lung cancer by real-time PCR
assay. We also attempted to clarify the function and molecular
mechanism of miR-338 in lung cancer metastasis.
Materials and methods
Cell culture and patients samples
Five lung tumor cell lines A549, NCI-H292, NCI-H460,
NCI-H446, NCI-H1299 and one human lung fibroblast cell MRC-5 were
purchased from the Cell Bank of Shanghai. All cells were maintained
in Dulbecco's modified Eagle's media (DMEM) or RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS; Life Technologies,
Carlsbad, CA, USA), 100 U/ml penicillin and 100 U/ml streptomycin.
A549 or NCI-H292 miR-338 overexpressing cell lines were constructed
by lentivirus assay (Shanghai Genechem, Co., Ltd., Shanghai,
China). Human lung cancer and adjacent normal tissues were obtained
from 115 patients in the First Affiliated Hospital of Zhengzhou
University. All samples were obtained with written patient informed
consent and the study was approved by the Medical Ethics and Human
Clinical Trial Committee.
RNA isolation and real-time quantitative
PCR
Total RNA from 115 pairs of lung cancer tissues or
cell lines were extracted with TRIzol (Invitrogen, Carlsbad, CA
USA) and the small non-coding miRNAs were isolated by mirVana miRNA
Isolation kit (Ambion, Austin, TX, USA) according to the
manufacturer's instruction. A total of 1 µg miRNAs were
reverse transcripted with Mir-X™ miRNA First-Strand Synthesis kit
(Takara Bio, Beijing, China) and the expression of mR-338 was
detected with Mir-X™ miRNA qRT-PCR SYBR kit (Takara Bio). Primers
for miR-338 and U6 snRNA were also purchased from Takara Bio.
CCK-8 proliferation assay
In order to examine the effect of miR-338 on
cellular proliferation, miR-338 overexpressing or miR-control cells
were seeded on 96-well plates (2,000/well). Then, 10 µl Cell
Counting kit-8 (CCK-8) was added for 1 h at 37°C. The absorbance of
OD450 was measured by micro-plate reader (Bio-Rad Laboratories,
Hercules, CA, USA), with OD630 as a reference wavelength. All the
experiments were repeated three time in triplicate.
Cell adhesion assay
Cell adhesion assay was performed to assess the
effect of miR-338 on cellular adhesion. Firstly, the 24-well plates
(Corning, Inc., Corning, NY, USA) were coated with matrix gel (1:50
dilution; BD Biosciences, San Jose, CA, USA) overnight at 4°C.
Then, 1×105 miR-338/miR-control stable infected cells
were seeded on the plates, and incubated for 30 min at 37°C. The
suspended cells were washed out with PBS twice, and adhesion cells
were fixed with 4% paraformaldehyde. Finally, cells were stained
with 0.4% typan blue, photographed and counted under a
microscope.
Transwell migration and invasion
assay
Transwell migration assays were performed with
24-well Transwell plates (Corning). Lung cancer cells
(5×104) suspended in 100 µl serum-free medium
were added into the upper chamber of plates, and 500 µl
medium with 10% FBS was added into the lower chamber. After 12 h,
the upper chamber was fixed, cells on the inner layer were removed
with a cotton swabs, stained with 0.4% typan blue and counted at
×100 magnification. For invasion assay, 1×105 cells were
seeded on matrix gel (BD Biosciences) pre-coated Transwell chamber,
then following the procedure of migration assay.
Xenograf tumor model
Six pairs of 5-weeks old femal BALB/c nude mice
(Vital River Laboratory Animal Technology, Co., Ltd., Beijing,
China) were implanted with 1×106 NCI-H292 miR-control or
miR-338 cells in 100 µl PBS by lateral tail vein injection.
Two months later, all animals were sacrificed and the lung were
dissected and fixed with 4% paraformaldehyde. Then the tissues were
sectioned and stained with hematoxylin and eosin (H&E).
Furthermore, the number of tumor clones in the lungs were
counted.
Dual-luciferase activity assay
Lung cancer cells (1×104) seeded in
24-well plates were co-transfected with 50 ng ITGB3 wild-type 3′UTR
(ITGB3-3′UTR-WT)/ITGB3 mutant 3′UTR (ITGB3-3′UTR-MUT) (GeneCopoeia,
Guangzhou, China), 100 nM miRNA control/miR-338 mimics (Shanghai
GenePharma, Co., Ltd., Shanghai, China), and 10 ng Renilla
luciferase vector (Promega, Madison, WI, USA) with Lipofectamine
3000 (Life Technologies). Cells were harvested for luciferase
activity assay at 48 h after transfection using Dual-luciferase
reporter assay kit (Promega). All experiments were performed
independently in triplicate.
Western blot analysis
Cells were lysed in RIPA lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China) supplemented with
protease inhibitor mix (Roche Diagnostics, Indianapolis, IN, USA).
Equal amount of protein was separated by 8% SDS-PAGE and then
transferred to PVDF membranes (Millipore, Billerica, MA, USA).
Membranes were blocked by 5% non-fat milk for 1 h, incubated with
ITGB3 antibody (Cell Signaling Technology, Danvers, MA, USA) or
β-actin antibody (Zhongshan Bio, Co., Ltd., Zhongshan, China)
overnight, then incubated with HRP-conjugated secondary antibody
and detected by Pierce™ ECL Plus western blotting substrate kit
(Thermo Fisher Scientific, Waltham, MA, USA).
Statistical analysis
Data were analyzed and presented as the mean ±
standard deviation (mean ± SD) using SPSS 13.0. Difference between
two group were estimated with Paired-samples t-test. The
association between miR-338 expression and clinicopathological
factors were analyzed with one-way ANOVA and survival curves were
plotted according to the Kaplan-Meier method using log-rank test.
Difference between groups were considered statistically significant
at P<0.05.
Results
miR-338 was downregulated in lung cancer
cell lines
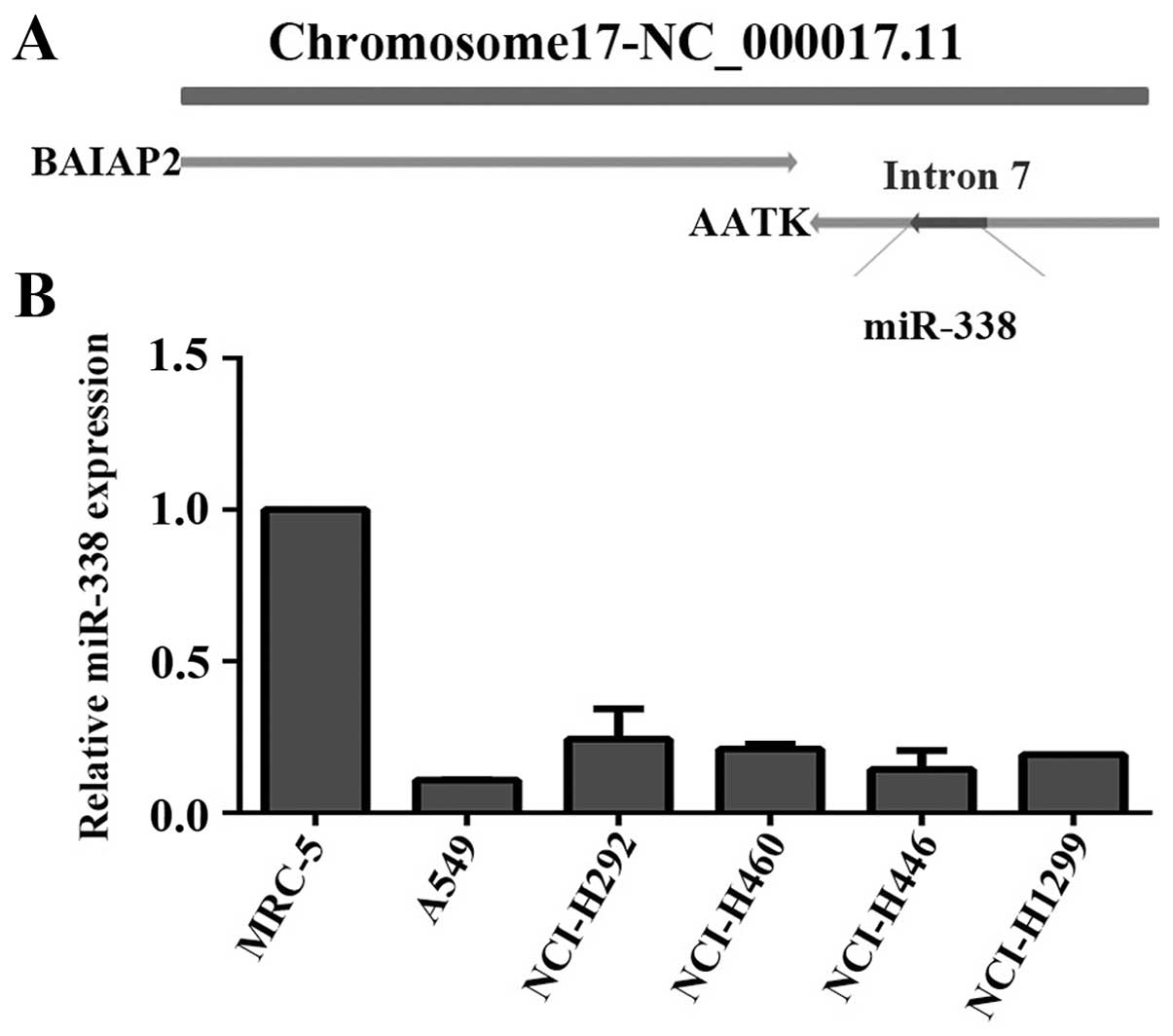
We firstly analyzed the gene location of miR-338 in
Gene Database. As shown in Fig. 1A,
miR-338 was located in the seventh intron of AATK gene which was an
adjacent gene of Brain specific angiogenesis inhibitor-associated
protein 2 (BAIAP2). Studies had demonstrated BAIAP2 was an
important adaptor protein that links Rho-family small GTPases, and
was involved in cell motility and the reorganization of actin
cytoskeleton (28–30). Considering adjacent genes often has
similar or reverse function (31,32),
we inferred that miR-338 may participate in the progress of
angiogenesis or cell motility.
Then, we analyzed miR-338 expression in normal human
lung fibroblast MRC-5 and five lung cancer cell lines. From the
results we found that miR-338 was evidently repressed in all lung
cancer cell lines compared with MRC cells (Fig. 1B).
The expression and clinical significance
of miR-338 in lung cancer tissues
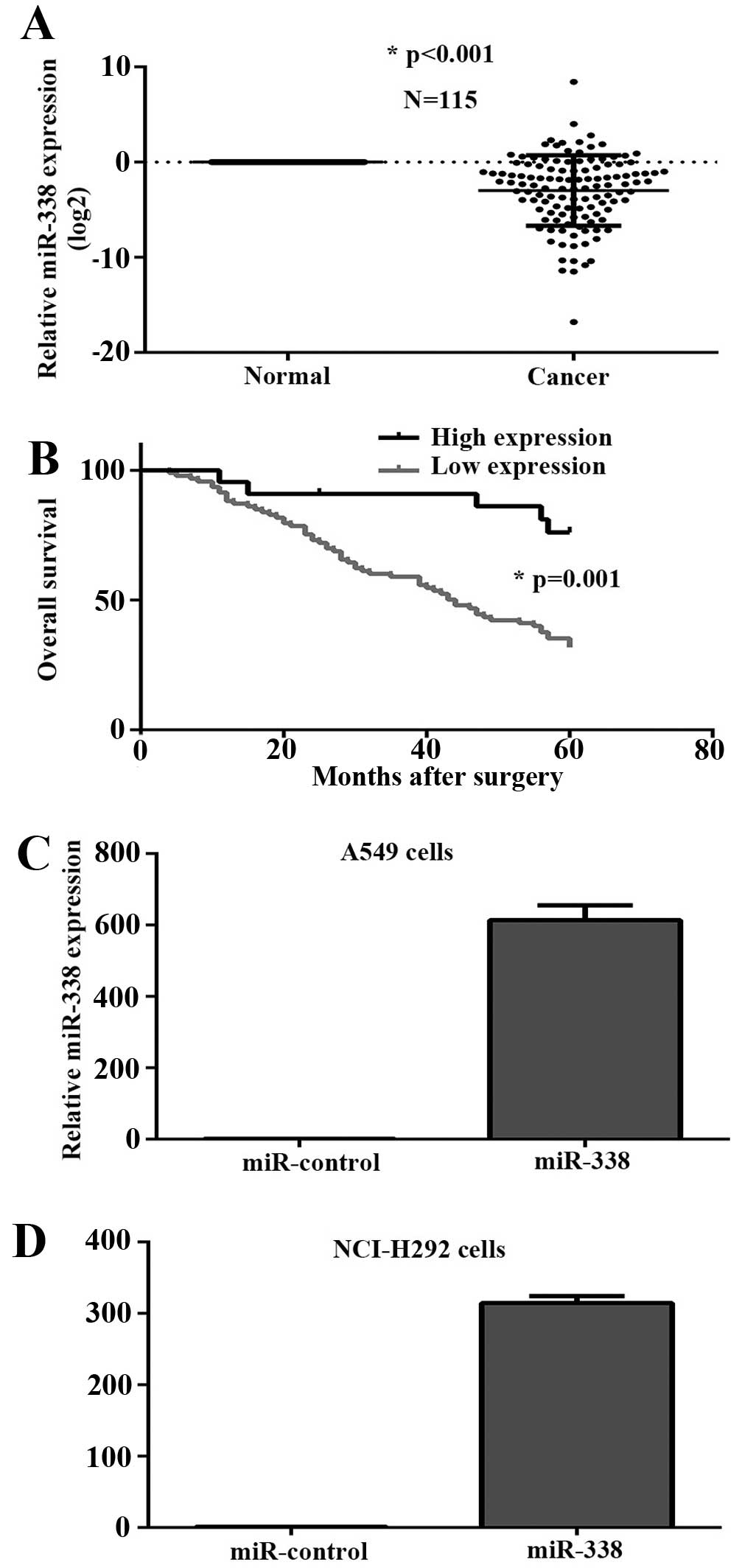
In order to further confirm the level of miR-338 in
lung cancer, we assessed the expression of miR-338 in 115 pairs of
lung cancer using real-time quantitative PCR. Results demonstrated
that miR-338 was significantly downregulated in tumor tissues
(Fig. 2A). Moreover, patients with
tumor emboli and recurrence had a lower expression of miR-338.
Decreased miR-338 level was also associated with TNM stage, while
there were no obviously difference about gender, age, smoking
history, tumor size and lymph node metastasis (Table I). As shown in Fig. 2B, the 5-year overall survival rate
of low miR-338 expressing group was significantly lower than that
of the high miR-338 expressing group (P=0.001).
 | Table IThe expression of miR-338 and
clinocopathological factors in lung cancer. |
Table I
The expression of miR-338 and
clinocopathological factors in lung cancer.
| Factors | N | Relative
expression | P-valuea |
|---|
| Gender |
| Male | 87 | −2.72±3.58 | 0.236 |
| Female | 28 | −3.68±3.99 | |
| Age (years) |
| <55 | 45 | −2.74±3.30 | 0.618 |
| ≥55 | 70 | −3.09±3.93 | |
| Smoking |
| Yes | 93 | −2.84±3.72 | 0.507 |
| No | 22 | −3.43±3.58 | |
| Tumor size
(cm) |
| ≤3 | 60 | −2.45±3.69 | 0.129 |
| >3 | 55 | −3.50±3.64 | |
| Tumor emboli |
| Yes | 26 | −4.82±4.35 | 0.005b |
| No | 89 | −2.44±3.44 | |
| Lymph node
metastasis |
| Yes | 39 | −3.28±3.52 | 0.496 |
| No | 76 | −2.78±3.78 | |
| Tumor
recurrence |
| Yes | 27 | −4.72±4.23 | 0.004b |
| No | 88 | −2.41±3.35 | |
| TNM |
| I–II | 65 | −2.11±3.16 | 0.025b |
| III–IV | 50 | −4.01±4.03 | |
miR-338 inhibits the proliferation and
adhesion of lung cancer cells
To investigate the function of miR-338 in the
progress of lung cancer miR-338 overexpressing stable cell lines
were constructed, and miR-338 RNA level was confirmed by real-time
PCR assay (Fig. 2C and D). CCK-8
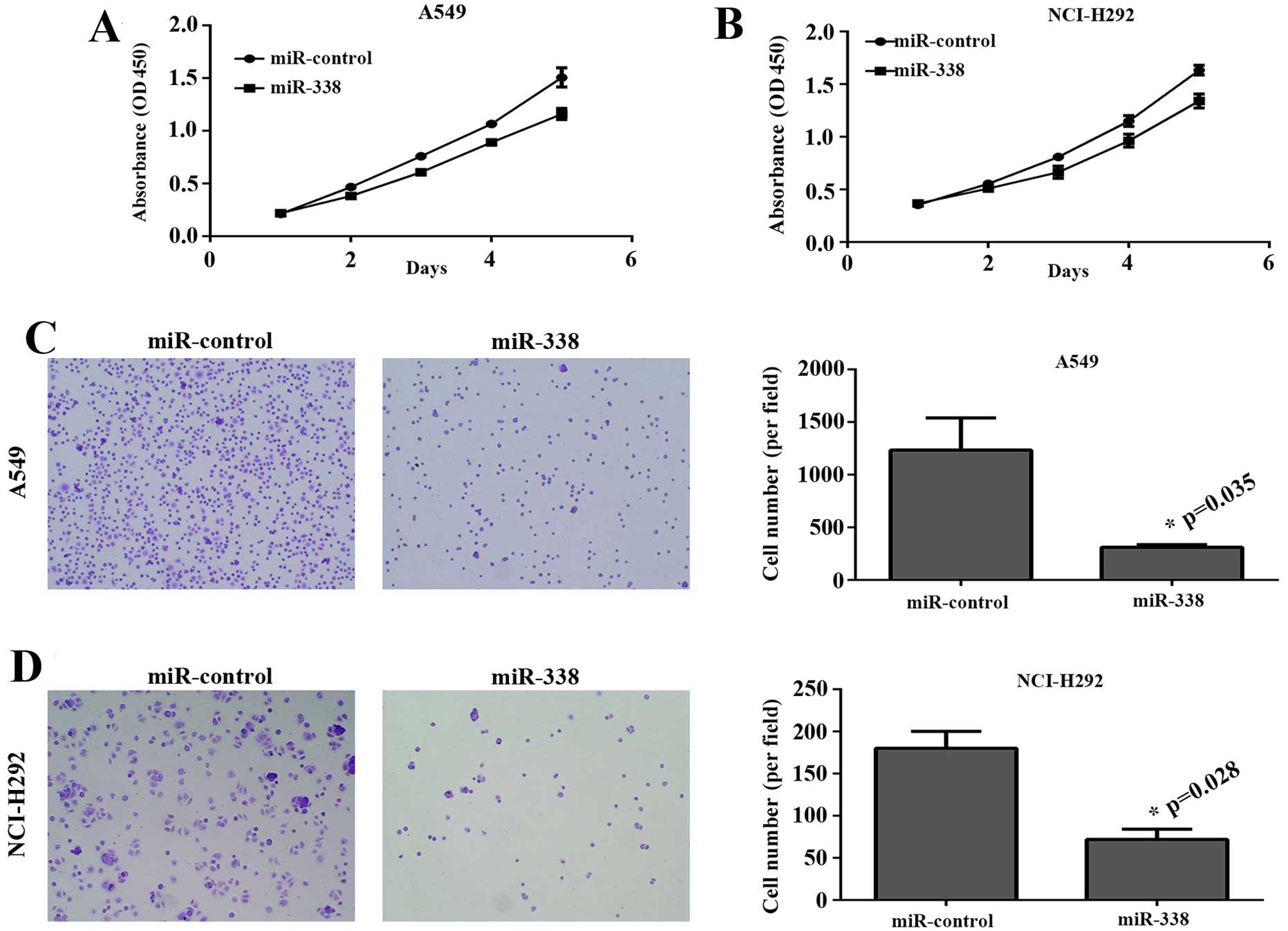
proliferation assay was performed to assess the effect of miR-338
on tumor growth. As shown in Fig. 3A
and B, upregulation of miR-338 inhibited the proliferation
rate. Furthermore, we decided to check whether miR-338 could affect
cellular adhesion. Adhesion assay results revealed that
overexpression of miR-338 in lung cancer cells obviously impaired
the ability of adhesion (Fig. 3C and
D).
miR-338 suppresses tumor migration and
invasion
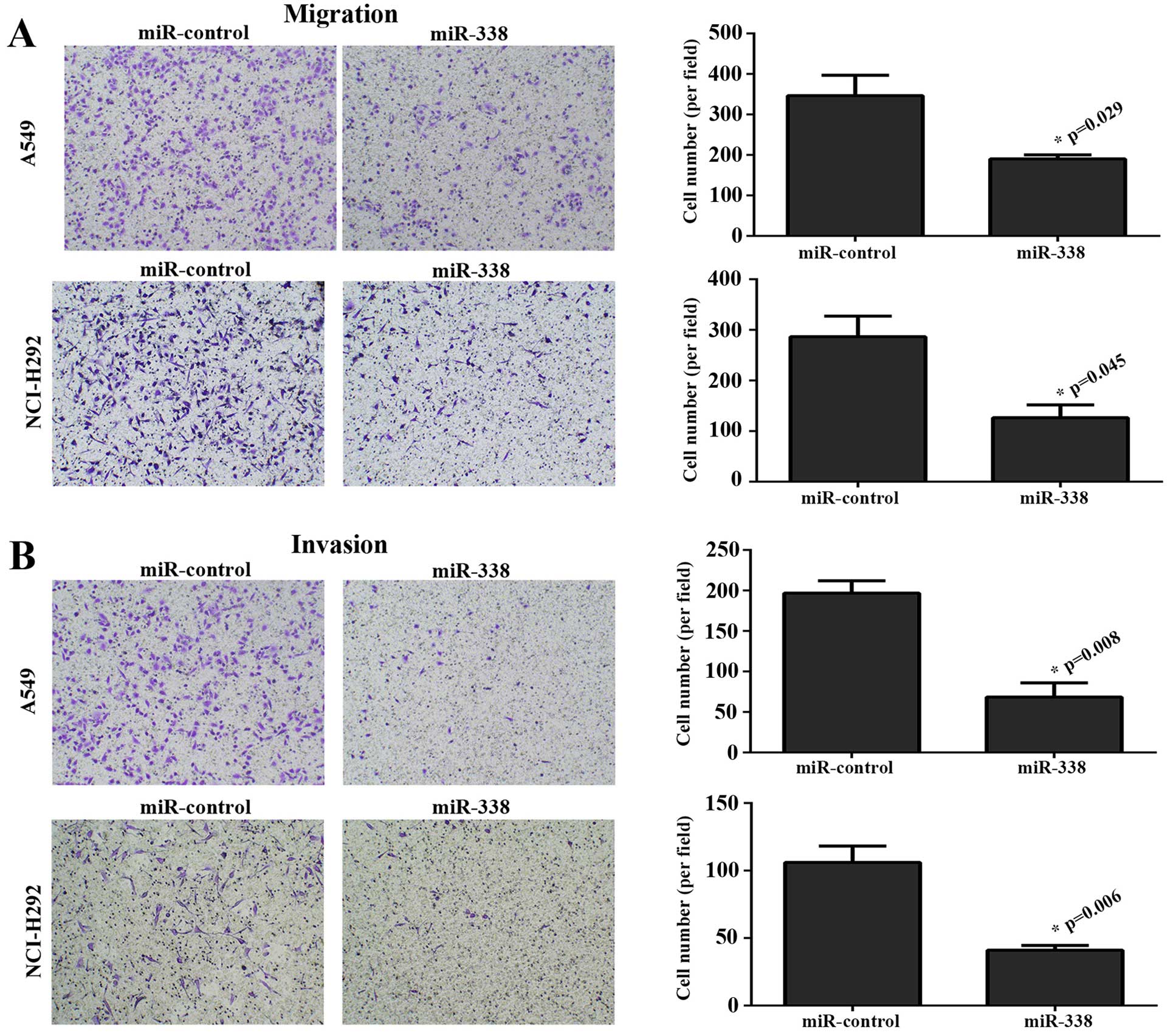
To explore the effect of miR-338 on cellular
migration and invasion, Transwell assays were performed. Fig. 4A shows that elevating the expression
of miR-338 in A549 and NCI-H292 cell lines markedly weakened the
migration ability. This effect was also observed in the invasion
assay (Fig. 4B).
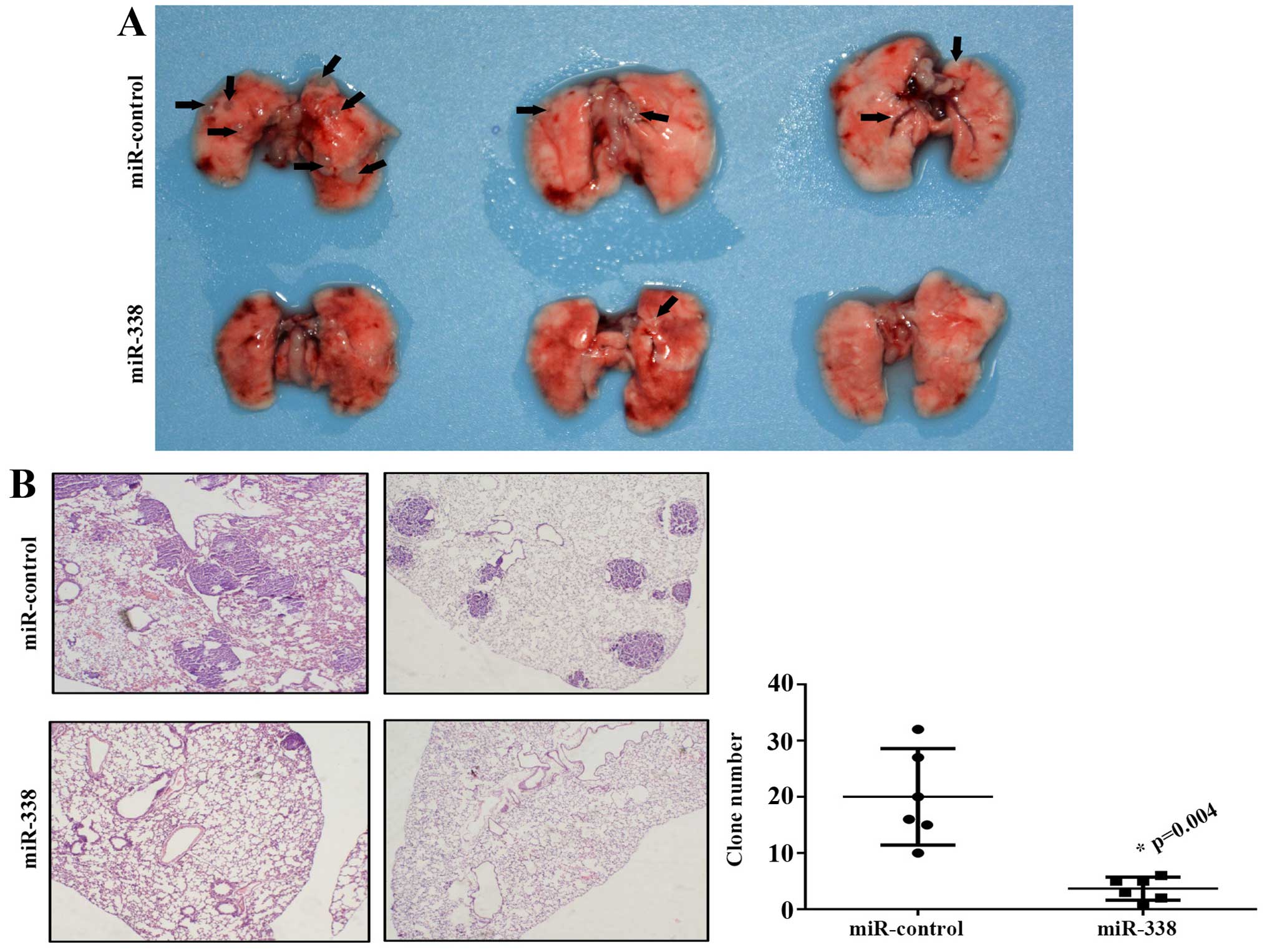
miR-338 abated lung cancer metastasis in
vivo
To further investigate whether miR-338 is sufficient
to lighten tumor metastasis in vivo, miR-338 overexpressing
and miR-control NCI-H292 cell lines were injected into the tail
vein of BALB/c nude mice. Results shown that the miR-338 group had
fewer tumor metastatic lesions compared with the control group
(Fig. 5A). The tumor foci in lung
tissues were confirmed by pathological section, and the tumor clone
number was counted by microscope (Fig.
5B).
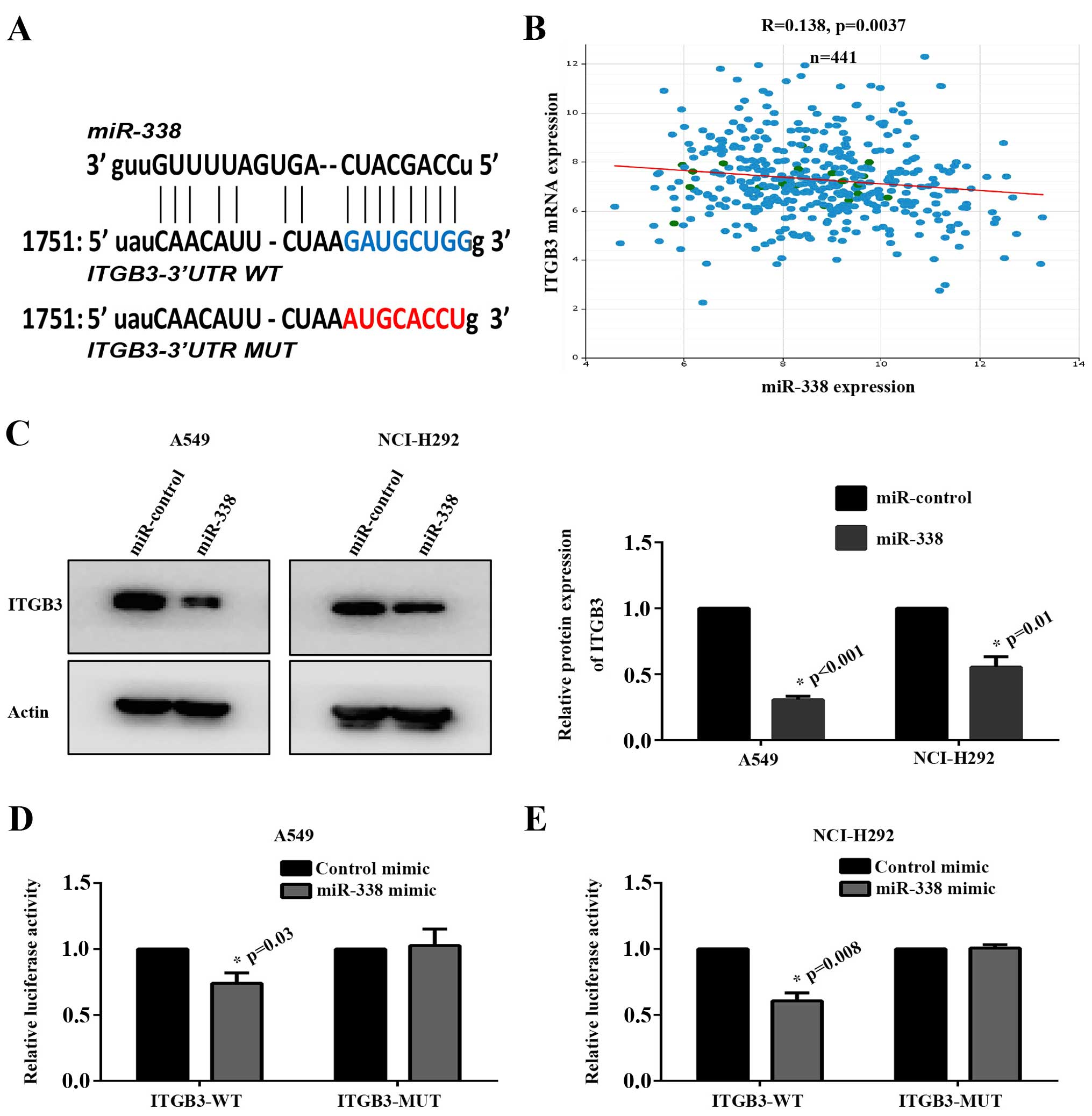
ITGB3 was a novel target gene of
miR-338
To uncover the molecular mechanism by which miR-338
performed a suppressor role in cancer metastasis. The target genes
were analyzed by software including StarBase, TargetScan and
microRNA.org. Finally, we identified ITGB3 which was
involved in tumor angiogenesis, cellular adhesion and cytoskeleton
rearrangement as a potential target gene of miR-338 (Fig. 6A). Furthermore, we analyzed the
correlation between miR-338 expression and ITGB3 mRNA expression in
441 lung cancer tissues from TCGA database. As shown in Fig. 6B, ITGB3 was negatively associated
with miR-338 in lung cancer (P=0.0037).
To confirm this hypothesis, protein expression of
ITGB3 in A549 and NCI-H292 miR-338 overexpressing stable cell lines
were detected by western blot assay. The results revealed that
exogenous transfection with miR-338 markedly suppressed ITGB3
expression (Fig. 6C). In order to
examine whether ITGB3 was a direct target of miR-338,
ITGB3-3′UTR-WT and ITGB3-3′UTR-MUT were constructed.
Dual-luciferase activity assay revealed that the luciferase
activity was significantly reduced in miR-338 mimics overexpressing
group compared with the control group. However, this change was
abrogated when the binding seed region was mutant (Fig. 6D and E).
Discussion
The role of miRNAs in the regulation of tumor
metastasis has been widely recognized in the recent years. The
progress on systemic delivery as well as applications for miRNAs as
therapeutic agents have witnessed the advancement of miR-34a in
liver cancer or metastatic cancer with liver involvement (33). Studies have proved that recovering
some tumor suppressor miRNAs such as miR-204, miR-145, miR-21
significantly abrogated the metastasis of human lung cancer in
vitro and in vivo (34–36).
Moreover, some plasma miRNAs including miR-486, miR-150, miR-152
and let-7c have been identified as biomarkers for lung cancer
(37,38). In the present study, we found that
miR-338 was downregulated in lung cancer cell lines. We also
validated that miR-338 was obviously impaired in lung cancer
tissues. Lower expression of miR-338 was associated with tumor
emboli and recurrence, indicating miR-338 may be a vital negative
regulator of metastatic lung cancer.
AATK is a brain apoptosis-associated tyrosine
kinase, and is a necessary pre-requisite for the induction of
growth arrest, apoptosis and neuronal differentiation. Similar to
host gene, miR-338 also displays its ability to regulate neurocyte
differentiation or apoptosis (39–41).
In carcinoma, miR-338 suppresses the proliferation of liver tumor
cells by targeting foxp4, and induces gastric cancer apoptosis by
targeting SSX2IP. The anti-proliferation effect of miR-338 was also
confirmed in the present study. BAIAP2 as an adjacent gene of AATK
is involved in neuronal growth-cone guidance, formation of stress
fibers and cytokinesis. As studies have validated that adjacent
genes usually share similar or related function (42) besides AATK has also been identified
as a regulator of neurite outgrowth (43). These findings hint that miR-338 may
participate in cellular migration or invasion. Our further results
validated that restoring the expression of miR-338 markedly impeded
lung cancer cells metastasis and adhesion.
ITGB3 as a receptor of various extracellular matrix
protein, and plays an important role in number of physiological and
pathological progress such as bone resorption, angiogenesis,
adhesion, tumor invasion and metastasis (44). Elevated expression of ITGB3 is
closely correlated with the metastatic potential of colorectal
cancer (10). In lung cancer, high
expression of ITGB3 promotes tumor metastasis and vascular invasion
(45). Studies have proven that
some miRNAs including let-7a, let-7c and miR-320 inhibit tumor
metastasis by targeting ITGB3 (12,46,47).
In this study, we identified that ITGB3 was a potential target gene
of miR-338 through bioinformatics analysis. This was further
confirmed by western blot assay and Dual-luciferase activity assay.
We also found that ITGB3 was negatively correlated with miR-338 in
441 lung cancer tissues by analysing data from TCGA database.
In conclusion, our results show that miR-338 was
downregulated in lung cancer and associated with tumor metastasis
and recurrence. We also identified ITGB3, a tumor
metastasis-related gene, as a novel target gene of miR-338,
suggesting that miR-338 is a potential target for lung cancer
therapy.
References
|
1
|
Siegel R, Ward E, Brawley O and Jemal A:
Cancer statistics, 2011: The impact of eliminating socioeconomic
and racial disparities on premature cancer deaths. CA Cancer J
Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang X, Xiao D, Wang Z, Zou Y, Huang L,
Lin W, Deng Q, Pan H, Zhou J, Liang C, et al: MicroRNA-26a/b
regulate DNA replication licensing, tumorigenesis, and prognosis by
targeting CDC6 in lung cancer. Mol Cancer Res. 12:1535–1546. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hall RD, Le TM, Haggstrom DE and Gentzler
RD: Angiogenesis inhibition as a therapeutic strategy in non-small
cell lung cancer (NSCLC). Transl Lung Cancer Res. 4:515–523.
2015.PubMed/NCBI
|
|
4
|
Liu MX, Zhou KC and Cao Y: MCRS1
overexpression, which is specifically inhibited by miR-129*,
promotes the epithelial-mesenchymal transition and metastasis in
non-small cell lung cancer. Mol Cancer. 13:2452014. View Article : Google Scholar
|
|
5
|
Manraj K and Saurabh M: Chronic pain grade
questionnaire. J Physiother. 59:602013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luanpitpong S, Li J, Manke A, Brundage K,
Ellis E, McLaughlin SL, Angsutararux P, Chanthra N, Voronkova M,
Chen YC, et al: SLUG is required for SOX9 stabilization and
functions to promote cancer stem cells and metastasis in human lung
carcinoma. Oncogene. Sep 21–2015.Epub ahead of print. PubMed/NCBI
|
|
7
|
Jin Y, Li F, Zheng C, Wang Y, Fang Z, Guo
C, Wang X, Liu H, Deng L, Li C, Wang H, et al: NEDD9 promotes lung
cancer metastasis through epithelial-mesenchymal transition. Int J
Cancer. 134:2294–2304. 2014. View Article : Google Scholar
|
|
8
|
Nurden AT, Fiore M, Nurden P and Pillois
X: Glanzmann thrombasthenia: A review of ITGA2B and ITGB3 defects
with emphasis on variants, phenotypic variability, and mouse
models. Blood. 118:5996–6005. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Miller PG, Al-Shahrour F, Hartwell KA, Chu
LP, Järås M, Puram RV, Puissant A, Callahan KP, Ashton J, McConkey
ME, et al: In Vivo RNAi screening identifies a leukemia-specific
dependence on integrin beta 3 signaling. Cancer Cell. 24:45–58.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lei Y, Huang K, Gao C, Lau QC, Pan H, Xie
K, Li J, Liu R, Zhang T, Xie N, et al: Proteomics identification of
ITGB3 as a key regulator in reactive oxygen species-induced
migration and invasion of colorectal cancer cells. Mol Cell
Proteomics: MCP. 10:M110 0053972011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Al-Sahaf O, Wang JH, Browne TJ, Cotter TG
and Redmond HP: Surgical injury enhances the expression of genes
that mediate breast cancer metastasis to the lung. Ann Surg.
252:1037–1043. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao B, Han H, Chen J, Zhang Z, Li S, Fang
F, Zheng Q, Ma Y, Zhang J, Wu N, et al: MicroRNA let-7c inhibits
migration and invasion of human non-small cell lung cancer by
targeting ITGB3 and MAP4K3. Cancer Lett. 342:43–51. 2014.
View Article : Google Scholar
|
|
13
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lei L, Huang Y and Gong W: Inhibition of
miR-92b suppresses nonsmall cell lung cancer cells growth and
motility by targeting RECK. Mol Cell Biochem. 387:171–176. 2014.
View Article : Google Scholar
|
|
15
|
Xu T, Liu X, Han L, Shen H, Liu L and Shu
Y: Upregulation of miR-9 expression as a poor prognostic biomarker
in patients with non-small cell lung cancer. Clin Transl Oncol.
16:469–475. 2014. View Article : Google Scholar
|
|
16
|
Cui R, Meng W, Sun HL, Kim T, Ye Z, Fassan
M, Jeon YJ, Li B, Vicentini C, Peng Y, et al: MicroRNA-224 promotes
tumor progression in nonsmall cell lung cancer. Proc Natl Acad Sci
USA. 112:E4288–E4297. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu F, Zhang H, Su Y, Kong J, Yu H and Qian
B: Up-regulation of microRNA-183-3p is a potent prognostic marker
for lung adenocarcinoma of female non-smokers. Clin Transl Oncol.
16:980–985. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yan F, Shen N, Pang J, Xie D, Deng B,
Molina JR, Yang P and Liu S: Restoration of miR-101 suppresses lung
tumorigenesis through inhibition of DNMT3a-dependent DNA
methylation. Cell Death Dis. 5:e14132014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fujiwara T, Katsuda T, Hagiwara K, Kosaka
N, Yoshioka Y, Takahashi RU, Takeshita F, Kubota D, Kondo T,
Ichikawa H, et al: Clinical relevance and therapeutic significance
of microRNA-133a expression profiles and functions in malignant
osteosarcoma-initiating cells. Stem Cells. 32:959–973. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lv M, Xu Y, Tang R, Ren J, Shen S, Chen Y,
Liu B, Hou Y and Wang T: miR141-CXCL1-CXCR2 signaling-induced Treg
recruitment regulates metastases and survival of non-small cell
lung cancer. Mol Cancer Ther. 13:3152–3162. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun T, Kalionis B, Lv G, Xia S and Gao W:
Role of exosomal noncoding RNAs in lung carcinogenesis. BioMed Res
Int. 2015:1258072015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barik S: An intronic microRNA silences
genes that are functionally antagonistic to its host gene. Nucleic
Acids Res. 36:5232–5241. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Recht M, Nemes L, Matysiak M,
Manco-Johnson M, Lusher J, Smith M, Mannucci P, Hay C, Abshire T,
O'Brien A, et al: Clinical evaluation of moroctocog alfa (AF-CC), a
new generation of B-domain deleted recombinant factor VIII
(BDDrFVIII) for treatment of haemophilia A: demonstration of
safety, efficacy, and pharmacokinetic equivalence to full-length
recombinant factor VIII. Haemophilia. 15:869–880. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang M, Liu R, Sheng J, Liao J, Wang Y,
Pan E, Guo W, Pu Y and Yin L: Differential expression profiles of
microRNAs as potential biomarkers for the early diagnosis of
esophageal squamous cell carcinoma. Oncol Rep. 29:169–176.
2013.
|
|
25
|
Peng Y, Liu YM, Li LC, Wang LL and Wu XL:
MicroRNA-338 inhibits growth, invasion and metastasis of gastric
cancer by targeting NRP1 expression. PLoS One. 9:e944222014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu H, Zhao L, Fang Q, Sun J, Zhang S, Zhan
C, Liu S and Zhang Y: MiR-338-3p inhibits hepatocarcinoma cells and
sensitizes these cells to sorafenib by targeting hypoxia-induced
factor 1α. PLoS One. 9:e1155652014. View Article : Google Scholar
|
|
27
|
Xue Q, Sun K, Deng HJ, Lei ST, Dong JQ and
Li GX: MicroRNA-338-3p inhibits colorectal carcinoma cell invasion
and migration by targeting smoothened. Jpn J Clin Oncol. 44:13–21.
2014. View Article : Google Scholar
|
|
28
|
Funato Y, Terabayashi T, Suenaga N, Seiki
M, Takenawa T and Miki H: IRSp53/Eps8 complex is important for
positive regulation of Rac and cancer cell motility/invasiveness.
Cancer Res. 64:5237–5244. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu PS, Jong TH, Maa MC and Leu TH: The
interplay between Eps8 and IRSp53 contributes to Src-mediated
transformation. Oncogene. 29:3977–3989. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oh SY, Knelson EH, Blobe GC and Mythreye
K: The type III TGFβ receptor regulates filopodia formation via a
Cdc42-mediated IRSp53-N-WASP interaction in epithelial cells.
Biochem J. 454:79–89. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Wit E and van Steensel B: Chromatin
domains in higher eukaryotes: Insights from genome-wide mapping
studies. Chromosoma. 118:25–36. 2009. View Article : Google Scholar
|
|
32
|
Moreno-Hagelsieb G: The power of operon
rearrangements for predicting functional associations. Comput
Struct Biotechnol J. 13:402–406. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sampson VB, Yoo S, Kumar A, Vetter NS and
Kolb EA: MicroRNAs and potential targets in osteosarcoma: Review.
Front Pediatr. 3:692015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Stephens NA and Sparks LM: Resistance to
the beneficial effects of exercise in type 2 diabetes: Are some
individuals programmed to fail? J Clin Endocrinol Metab. 100:43–52.
2015. View Article : Google Scholar
|
|
35
|
Ling DJ, Chen ZS, Zhang YD, Liao QD, Feng
JX, Zhang XY and Shi TS: MicroRNA-145 inhibits lung cancer cell
metastasis. Mol Med Rep. 11:3108–3114. 2015.
|
|
36
|
Yan S, Liu G, Pei C, Chen W, Li P, Wang Q,
Jin X, Zhu J, Wang M and Liu X: Inhibition of NADPH oxidase
protects against metastasis of human lung cancer by decreasing
microRNA-21. Anticancer Drugs. 26:388–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dou H, Wang Y, Su G and Zhao S: Decreased
plasma let-7c and miR-152 as noninvasive biomarker for
non-small-cell lung cancer. Int J Clin Exp Med. 8:9291–9298.
2015.PubMed/NCBI
|
|
38
|
Li W, Wang Y, Zhang Q, Tang L, Liu X, Dai
Y, Xiao L, Huang S, Chen L, Guo Z, et al: MicroRNA-486 as a
biomarker for early diagnosis and recurrence of non-small cell lung
cancer. PLoS One. 10:e01342202015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao X, He X, Han X, Yu Y, Ye F, Chen Y,
Hoang T, Xu X, Mi QS, Xin M, et al: MicroRNA-mediated control of
oligodendrocyte differentiation. Neuron. 65:612–626. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Saba R, Goodman CD, Huzarewich RL,
Robertson C and Booth SA: A miRNA signature of prion induced
neurodegeneration. PLoS One. 3:e36522008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ebrahimi-Barough S, Massumi M,
Kouchesfahani HM and Ai J: Derivation of pre-oligodendrocytes from
human endometrial stromal cells by using overexpression of microRNA
338. J Mol Neurosci. 51:337–343. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ribeiro DM and Sonati MF: Regulation of
human alpha-globin gene expression and alpha-thalassemia. Genet Mol
Res. 7:1045–1053. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tomomura M, Hasegawa Y, Hashikawa T,
Tomomura A, Yuzaki M, Furuichi T and Yano R: Differential
expression and function of apoptosis-associated tyrosine kinase
(AATYK) in the developing mouse brain. Brain Res Mol Brain Res.
112:103–112. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vellon L, Menendez JA and Lupu R:
AlphaVbeta3 integrin regulates heregulin (HRG)-induced cell
proliferation and survival in breast cancer. Oncogene.
24:3759–3773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xia ZJ, Hu W, Wang YB, Zhou K and Sun GJ:
Expression heterogeneity research of ITGB3 and BCL-2 in lung
adenocarcinoma tissue and adenocarcinoma cell line. Asian Pac J
Trop Med. 7:473–477. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Müller DW and Bosserhoff AK: Integrin beta
3 expression is regulated by let-7a miRNA in malignant melanoma.
Oncogene. 27:6698–6706. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sun L, Liu B, Lin Z, Yao Y, Chen Y, Li Y,
Chen J, Yu D, Tang Z, Wang B, et al: MiR-320a acts as a prognostic
factor and inhibits metastasis of salivary adenoid cystic carcinoma
by targeting ITGB3. Mol Cancer. 14:962015. View Article : Google Scholar : PubMed/NCBI
|