Introduction
Gastric cancer (GC) is the fourth most commonly
diagnosed cancer in males and the fifth in females worldwide, with
over 751,600 new cases and 723,100 deaths estimated to have
occurred in 2012 (1). The highest
incidence rates are found in Eastern Asia (particularly in Korea,
Mongolia, Japan and China), Central and Eastern Europe, and South
America, and the lowest incidences are noted in Northern America
and most parts of Africa. The incidence is twice higher in males
than in females (1).
The tumor microenvironment plays a pivotal role in
tumourigenesis and subsequent dissemination of cancer cells by
coordinating morphological transformation of cancer cells and their
proliferation, survival and invasion (2,3).
Recent reviews have highlighted a profound role played by a group
of proteins in the tumour microenvironment which belong to the CCN
family. CCN is an acronym for cysteine-rich protein 61 (CYR61),
connective tissue growth factor (CTGF) and nephroblastoma
overexpressed (NOV) which comprises CYR61, CTGF, NOV and another
three members i.e. Wnt1-inducible signalling pathway protein
(WISP)-1, -2 and -3 (4–6).
CCNs are matricellular proteins and can be secreted
into the extracellular matrix. CCN3 can be detected in the culture
medium, extracellular matrix (ECM) and at the cell membrane
(7–9). The CCN family proteins have been
preserved during evolution which can be found in vertebrates
including zebra fish, chickens, mice, rats and humans. CCN proteins
generally have an N-terminal secretory signal peptide and four
functional domains: an insulin-like growth factor binding protein
domain (IGFBP); a Von Mayebrand factor type C (VWC) domain; a
thrombospondin type-1 repeat module (TSP-1) and a cysteine knot
domain (CT). The two N-terminal domains are separated from the two
C-domains by a linker with a variable sequence of amino acids.
However, CCN5 does not have the CT domain (10).
An elevated expression of CCN1 has been demonstrated
in a variety of malignancies including ovarian, prostate, breast
and colorectal cancers (11–14).
Similarly, an overexpression of CTGF has been noted in breast and
colorectal cancer, oesophageal squamous cell carcinoma and
pancreatic cancer (15–18), while increased expression of NOV has
been shown in prostate and cervical cancers (19,20).
In contrast to these findings, reduced expression of these three
CCNs has also been observed in certain solid tumours. For example,
CYR61 expression is reduced in endometrial cancer and lung cancer
(21,22). A reduced expression of CYR61 has
also been seen in advanced GC (23). CTGF can act as an inhibitor in lung
cancer by suppressing proliferation of non-small cell lung cancer
cells (24). A similar inhibitory
effect on the proliferation of cancer cells has also been evident
for the NOV protein in gliobastoma and malignant adrenocortical
tumour cells (25,26). This suggests that the expression and
function of CCNs in malignancies can be organ- or
tissue-specific.
Early studies have demonstrated a profound role
played by CCNs in fibrosis which has been reviewed most recently by
Riser et al (6). Their
involvement in fibrotic disorder stimulated research interest
concerning their implication in a specific histologic type of
gastric carcinoma, i.e. scirrhous carcinoma which is well known for
a vast fibrous stroma, rapid and invasive growth and poor
prognosis. Tanaka et al identified a novel variant of WISP1
that was highly expressed in scirrhous carcinomas. This variant
lacks the von Mayebrand type C module and is named WISP1v. WISP1v
can induce transformation and promote proliferation and invasion of
GC cells through both autocrine and paracrine pathways (27). Tanaka et al also reported a
loss of function mutation of WISP3 with a frequency of 10–20% in
microsatellite unstable gastric carcinoma. This mutation resulted
in a truncated variant of WISP3 that was lacking the TSP-1 and CT
domains and was unable to suppress the invasiveness of GC cells
(28). Although reduced expression
of CYR61 was reported in advanced GC which was inversely correlated
with the expression of MMP-7 (23),
most studies demonstrated a positive role played by CYR61 in GC by
promoting invasion, metastasis and also tumour-associated
angiogenesis (29,30). CYR61 promotes GC cell invasion
through hypoxia-inducing factor-1α (HIF-1α)-dependent upregulation
of plasminogen activator inhibitor-1 (PAI-1) (31).
Elevated expression of CYR61 and CTGF has been
observed in GC which is associated with lymph node metastasis,
however the expression of NOV in GC is yet to be revealed. The
present study aimed to determine the expression of these three CCNs
in a cohort of GC tumours, in particular, by dissecting the role
played by NOV in GC.
Materials and methods
Cell lines and culture conditions
Human GC cell lines AGS and HGC27 were purchased
from the European Collection of Cell Cultures (ECACC; Salisbury,
UK) and incubated at 37°C, with 5% CO2 and 95% humidity.
The wild-type cells were maintained in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% fetal calf serum (PAA
Laboratories Ltd., Somerset, UK), penicillin, streptomycin and
amphotericin B.
Human gastric tissues
Gastric adenocarcinoma and Siewert type III
gastrooesophageal junction adenocarcinoma tissues (n=245) with
matched adjacent background tissues (n=158) were immediately
collected after surgical resection at the Beijing Cancer Hospital
with informed consent from the patients. All patients underwent
surgery without any prior treatment. The tissue samples were stored
at −80°C at the Tissue Bank of Peking University Oncology School
with a record of the relevant clinical and histopathological data.
All protocols were reviewed and approved by the Beijing Cancer
Hospital Research Ethics Committee (MTA10062009).
RNA isolation, reverse
transcription-polymerase chain reaction (RT-PCR) and quantitative
real-time PCR (qPCR)
RNA was extracted from confluent cells in a 25
cm2 flask using total RNA isolation (TRI) reagent
(Sigma-Aldrich, Dorset, UK). Fresh frozen tissues were also first
homogenised in the TRI reagent. First strand of cDNA was
synthesised from 1 µg RNA using a first-strand DNA synthesis
kit (Bio-Rad, Hemel Hempstead, UK). Quantitative analysis of NOV
mRNA expression in GC tissues was performed using Amplifluor™-based
real-time PCR, in which a 6-carboxy-fluorescine-tagged Uniprimer™
(Biosearch Technologies, Inc.) was used as a probe along with a
pair of specific primers with an addition of a Z-sequence
(actgaacctgaccgtaca) to the 5′-end of the reverse primer. The
quality of cDNA samples was verified using
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a housekeeping
gene. All the primer sequences are listed in Table I.
 | Table IPrimers used for PCR and qPCR. |
Table I
Primers used for PCR and qPCR.
| Primer | Forward primer | Reverse primer |
|---|
| NOV (PCR) |
CTCCAAGAAAAGTTGAGGTG |
CTGGCTTCTTGACTATTTGC |
| NOV (qPCR) |
CTGTGAACAAGAGCCAGAG |
ACTGAACCTGACCGTACACTTGAACTGCAGGTGGAT |
| GAPDH (PCR) |
GGCTGCTTTTAACTCTGGTA |
GACTGTGGTCATGAGTCCTT |
| GAPDH (qPCR) |
CTGAGTACGTCGTGGAGTC |
ACTGAACCTGACCGTACACAGAGATGACCCTTTTG |
| CYR61 (qPCR) |
GGGCTGGAATGCAACTTC |
ACTGAACCTGACCGTACACGTTTTGGTAGATTCTGGAG |
| CTGF (qPCR) |
GAGTGGGTGTGTGACGAG |
ACTGAACCTGACCGTACAGGCAGTTGGCTCTAATCATA |
Quantitative analyses of transcript
expression of CYR61, CTGF and NOV in human GC
Following the real-time PCR quantification of each
gene transcripts, the number of samples with valid data for each
individual genes were: 322 samples for NOV, 252 samples for
CYR61 and 320 samples for CTGF. These cohorts were
composed of 230 men (71.4%) and 92 women (28.6%) in the NOV
cohort, 180 men (71.4%) and 72 women (28.6%) in the CYR61
cohort, and 228 men (71.3%) and 92 women (28.7%) in the CTGF
cohort. Data are shown in Tables
II–IV.
| Table IIExpression of NOV in gastric
cancer. |
Table II
Expression of NOV in gastric
cancer.
| Category | No. | Mean ± SEM
(copies) | P-value |
|---|
| Tissue | | | |
| Tumour | 322 |
8,893±1,303 | |
| Normal | 183 |
3,262±1,058 |
0.0009 |
| Gender | | | |
| Male | 230 |
9,583±1,645 | |
| Female | 92 |
7,170±1,971 |
0.35 |
| Location | | | |
| Cardia | 66 |
8,384±2,795 | |
| Fundus | 21 |
8,679±4,797 |
0.86 |
| Corpus | 61 |
7,678±2,795 |
0.59 |
| Pylorus | 131 |
8,326±1,786 |
0.84 |
|
Differentiation | | | |
| Diff-H | 1 | 33,009 | |
| Diff-HM | 6 | 172.2±56.6 | |
| Diff-M | 62 |
8,677±2,779 |
0.0033 |
| Diff-ML | 82 |
9,130±2,435 |
0.0004 |
| Diff-L | 136 |
8,610±1,848 | <0.001 |
| T stage | | | |
| T1 | 16 |
2,754±2,450 | |
| T2 | 26 |
4,456±2,325 |
0.62 |
| T3 | 41 |
9,478±5,181 |
0.25 |
| T4 | 231 |
9,503±1,500 |
0.026 |
| T1+T2 | 42 |
3,808±1,701 | |
| T3+T4 | 272 |
9,499±1,490 |
0.013 |
| N stage | | | |
| N0 | 70 |
8,735±2,479 | |
| N1 | 48 |
9,123±3,036 |
0.92 |
| N2 | 65 |
5,845±3,214 |
0.48 |
| N3 | 133 | 10,311±2,106 |
0.63 |
| N1+N2+N3 | 246 |
8,899±1,538 |
0.96 |
| M stage | | | |
| M0 | 280 |
8,644±1,281 | |
| M1 | 41 | 10,813±5,350 |
0.7 |
| TNM stage | | | |
| I | 25 |
1,847±1,569 | |
| II | 60 |
9,689±2,758 |
0.016 |
| III | 219 |
9,261±1,672 |
0.0017 |
| IV | 9 |
9,756±9,560 |
0.44 |
| II+III+IV | 228 |
9,366±1,421 |
0.0007 |
| Vascular
invasion | | | |
| No invasion | 151 |
6,865±1,526 | |
| Invasion | 156 |
9,257±1,803 |
0.63 |
| Clinical
outcome | | | |
| Disease-free | 119 |
7,464±1,832 | |
| Metastases | 15 | 12,921±6,376 |
0.42 |
| Death | 185 |
9,626±1,869 |
0.41 |
| Table IVExpression of CTGF in gastric
cancer. |
Table IV
Expression of CTGF in gastric
cancer.
| Category | No. | Mean ± SEM
(copies) | P-value |
|---|
| Tissue | | | |
| Tumour | 320 |
224.3±27.9 | |
| Normal | 183 |
247.7±38.7 | 0.62 |
| Gender | | | |
| Male | 228 | 209.5±22.92 | |
| Female | 92 |
261.4±79.1 | 0.53 |
| Location | | | |
| Cardia | 65 |
251.4±54.2 | |
| Fundus | 21 |
234.6±66.3 | 0.85 |
| Corpus | 61 |
158.2±44.0 | 0.18 |
| Pylorus | 130 | 251±52.8 | 1 |
|
Differentiation | | | |
| Diff-H | 1 | 16.77 | |
| Diff-HM | 6 |
142.2±61.4 | |
| Diff-M | 62 |
259.6±53.0 | 0.17 |
| Diff-ML | 81 |
266.6±60.9 | 0.17 |
| Diff-L | 135 |
197.6±48.1 | 0.49 |
| T stage | | | |
| T1 | 16 |
49±12.2 | |
| T2 | 25 | 218.8±93.55 | 0.084 |
| T3 | 41 |
252.3±90.0 | 0.031 |
| T4 | 230 |
231.6±33.6 | 0 |
| T1+T2 | 41 | 154.1±59.08 | |
| T3+T4 | 271 |
234.8±31.5 | 0.23 |
| N stage | | | |
| N0 | 70 |
250.7±78.5 | |
| N1 | 48 |
120±35.1 | 0.13 |
| N2 | 64 |
292.2±57.6 | 0.67 |
| N3 | 132 |
218.7±43.3 | 0.72 |
| N1+N2+N3 | 244 |
218.9±28.9 | 0.7 |
| M stage | | | |
| M0 | 278 |
199.4±26.6 | |
| M1 | 41 | 400±119 | 0.11 |
| TNM stage | | | |
| I | 25 | 225.8±97.01 | |
| II | 59 |
197.2±83.2 | 0.82 |
| III | 218 |
224.5±29 | 0.99 |
| IV | 9 | 407±349 | 0.63 |
| II+III+IV | 286 |
224.5±29.9 | 0.99 |
| Vascular
invasion | | | |
| No invasion | 150 |
246.6±45.5 | |
| Invasion | 155 |
206.7±36.3 | 0.49 |
| Clinical
outcome | | | |
| Disease-free | 119 |
237.3±48.9 | |
| Metastases | 15 |
119.2±47.6 | 0.09 |
| Death | 183 |
227.1±36.7 | 0.87 |
Construction of the ribozyme transgene
targeting human NOV and the establishment of corresponding stable
transfectants
Anti-human NOV hammerhead ribozymes were designed
using the Zuker RNA mFold program (Zuker 2003). The ribozymes and
full-length coding sequence of human NOV were cloned into a
pEF6/V5-His TOPO vector (Invitrogen, Paisley, UK). The verified
ribozyme transgenes, NOV expression vectors and empty plasmids were
transfected into AGS and HGC27 cells, respectively, using an
Easyjet Plus electroporator (EquiBio, Kent, UK). After one week of
selection with 5 µg/ml blasticidin, the selected cells were
maintained in DMEM with 0.5 µg/ml blasticidin.
Western blot analysis
The protein concentrations in the cell lysates were
determined using the DC protein assay kit (Bio-Rad) and the ELx800
spectrophotometer (Bio-Tek, Winooski, VT, USA). Proteins were
probed with the anti-NOV antibody (1:2,000; Abcam, Ltd., Cambridge,
UK) and anti-GAPDH antibody (1:2,000; Santa Cruz Biotechnology,
Inc.) as housekeeping gene control, followed by a
peroxidase-conjugated secondary antibody (1:2,000; Sigma). Protein
bands were visualised using a chemiluminescence detection kit
(Luminata, Millipore Ltd., Watford, UK) and photographed using
Syngene imager (Syngene International Ltd., Bangalore, India).
In vitro cell growth assay
Cells (3,000) were seeded into 96-well plates in
normal culture medium. The cells were incubated over a period of up
to 4 days, and were then fixed with 4% formaldehyde followed by
staining with 0.5% crystal violet. The crystal violet was dissolved
in 10% acetic acid prior to a colorimetric detection of cell
density at a wavelength of 540 nm using the ELx800
spectrophotometer.
In vitro invasion assay
The in vitro invasion assay was previously
described (32). All the culture
plate inserts containing 8-µm pores were pre-coated with 50
µg of Matrigel (BD Bioscience, Oxford, UK) and air-dried.
Cells (30,000) were seeded to each well after a 40-min rehydration
of the Matrigel. After an incubation of 72 h, cells that had
migrated through the matrix to the other side of the insert were
fixed and stained. The cell number was determined.
Cell-matrix adhesion assay
The cell-matrix adhesion assay was conducted as
previously described (32). The
96-well culture plate was pre-coated with 5 µg of Matrigel
and air-dried. Following the rehydration, 30,000 cells were seeded
to each well. After an incubation of 40 min, non-adherent cells
were washed-off using phosphate-buffered saline (PBS). The adherent
cells were counted after a fixation and staining using crystal
violet.
Wound healing assay
The assay was performed following a previously
described procedure (32). In
brief, the monolayer of cells was scraped with a 10 µl
pipette tip. The migration of cells was photographed using a
time-lapse image system (EVOS, Life Technologies Ltd., Paisley,
UK).
Statistical analysis
Statistical analyses were performed using SPSS
(version 11; SPSS, Inc., Chicago, IL, USA). Mann-Whitney U test and
t-test were used for non-parametric and normally distributed data,
respectively, including research data from the clinical cohort and
cell-based experiments. Kaplan-Meier survival analysis was also
performed using SPSS statistical software. Differences were
considered to be statistically significant at p<0.05.
Results
Increased expression of NOV in human
GC
Transcript levels of CYR61, CTGF and
NOV were determined in the GC cohort using real-time PCR,
respectively. The results showed that NOV expression was
significantly upregulated in gastric tumours compared to normal
tissue (p=0.009) (Table II). An
increased expression of NOV in GC was associated with local
invasion. The transcript levels of NOV were higher in the tumours
with more advanced local invasion. According to the
tumor-node-metastasis (TNM) staging, T4 tumours which invaded the
serosa or adjacent structures expressed higher levels of NOV
transcripts, p=0.0026 vs. T1. Tumours classified as T3 and T4 which
invaded beyond subserosal connective tissues expressed higher
levels of NOV transcripts in comparison with tumours (T1 and T2)
with less local invasion (p=0.0013). According to the overall TNM
staging, stage I GCs exhibited lower expression levels of NOV,
p=0.016 vs. stage II, p=0.0017 vs. stage III, and p=0.0007 vs.
tumours of stage II-IV. Notably, we found that high-moderately
differentiated tumours exhibited significantly lower levels of NOV
expression in comparison with moderately and/or poorly
differentiated tumours. However, no association was observed for
lymph node and distant metastases.
CYR61 is upregulated in GCs
CYR61 mRNA levels were significantly elevated
in the GC tissues compared to levels in the non-cancerous tissues
(p=0.0019), particularly in paired tissues (p=0.0013) (Table III). However, the expression of
CYR61 transcripts appeared to be lower in more advanced
tumours (stage III and IV) according to the TNM staging, although
it did not reach a statistically significant level. According to
clinical outcomes, tumours with distant metastases had lower
expression levels of CYR61 (p=0.055) compared with that of
patients who remained disease-free. No association was observed
between CYR61 expression and differentiation and local
invasion.
| Table IIIExpression of CYR61 in gastric
cancer. |
Table III
Expression of CYR61 in gastric
cancer.
| Category | No. | Mean ± SEM
(copies) | P-value |
|---|
| Tissue | | | |
| Tumour | 252 |
2,225±668 | |
| Normal | 175 |
120.1±50.5 | 0.0019 |
| Gender | | | |
| Male | 180 |
2,737±913 | |
| Female | 72 | 947±475 | 0.083 |
| Location | | | |
| Cardia | 50 | 3,558±2,288 | |
| Fundus | 12 | 322±172 | 0.86 |
| Corpus | 52 | 2,824±1,622 | 0.59 |
| Pylorus | 102 |
1,717±780 | 0.84 |
|
Differentiation | | | |
| Diff-H | 1 | 4400.7 | |
| Diff-HM | 5 | 1,898±1,897 | |
| Diff-M | 52 | 1,890±1,426 | 1 |
| Diff-ML | 64 | 2,537±1,747 | 0.81 |
| Diff-L | 106 |
1,255±558 | 0.76 |
| T stage | | | |
| T1 | 13 | 8,723±6,131 | |
| T2 | 21 | 8.96±8.53 | 0.18 |
| T3 | 27 | 2,821±1,860 | 0.37 |
| T4 | 185 |
1,756±715 | 0.28 |
| T1+T2 | 34 | 3,341±2,402 | |
| T3+T4 | 212 |
1,892±666 | 0.56 |
| N stage | | | |
| N0 | 59 | 3,178±1,572 | |
| N1 | 42 | 1,050±1,035 | 0.26 |
| N2 | 50 | 1,612±1,206 | 0.43 |
| N3 | 96 | 2,108±1,166 | 0.59 |
| N1+N2+N3 | 188 |
1,740±712 | 0.41 |
| M stage | | | |
| M0 | 220 |
2,439±758 | |
| M1 | 32 | 755±663 | 0.097 |
| TNM stage | | | |
| I | 21 | 5,400±3,855 | |
| II | 50 |
1,452±923 | 0.33 |
| III | 169 |
1,928±791 | 0.39 |
| IV | 6 | 450±445 | 0.22 |
| II+III+IV | 225 |
1,783±628 | 0.36 |
| Vascular
invasion | | | |
| No invasion | 124 |
1,739±678 | |
| Invasion | 117 |
2,523±1193 | 0.57 |
| Clinical
outcome | | | |
| Disease-free | 98 | 2,744±1,131 | |
| Metastases | 10 | 418±399 | 0.055 |
| Death | 142 |
2,026±893 | 0.62 |
Higher expression of CTGF in GC and the
involvement in local invasion
Although no statistically significant differences
were noted for CTGF mRNA levels in GCs compared with
adjacent normal gastric tissues, a higher transcript level of
CTGF in GCs was positively associated with local invasion
(T4 vs. T1, p<0.001; T3 vs. T1, p=0.031; Table IV). However, there were no other
significant correlations between CTGF mRNA levels and other
clinical parameters.
Expression of CYR61, CTGF and NOV and
survival of patients with GC
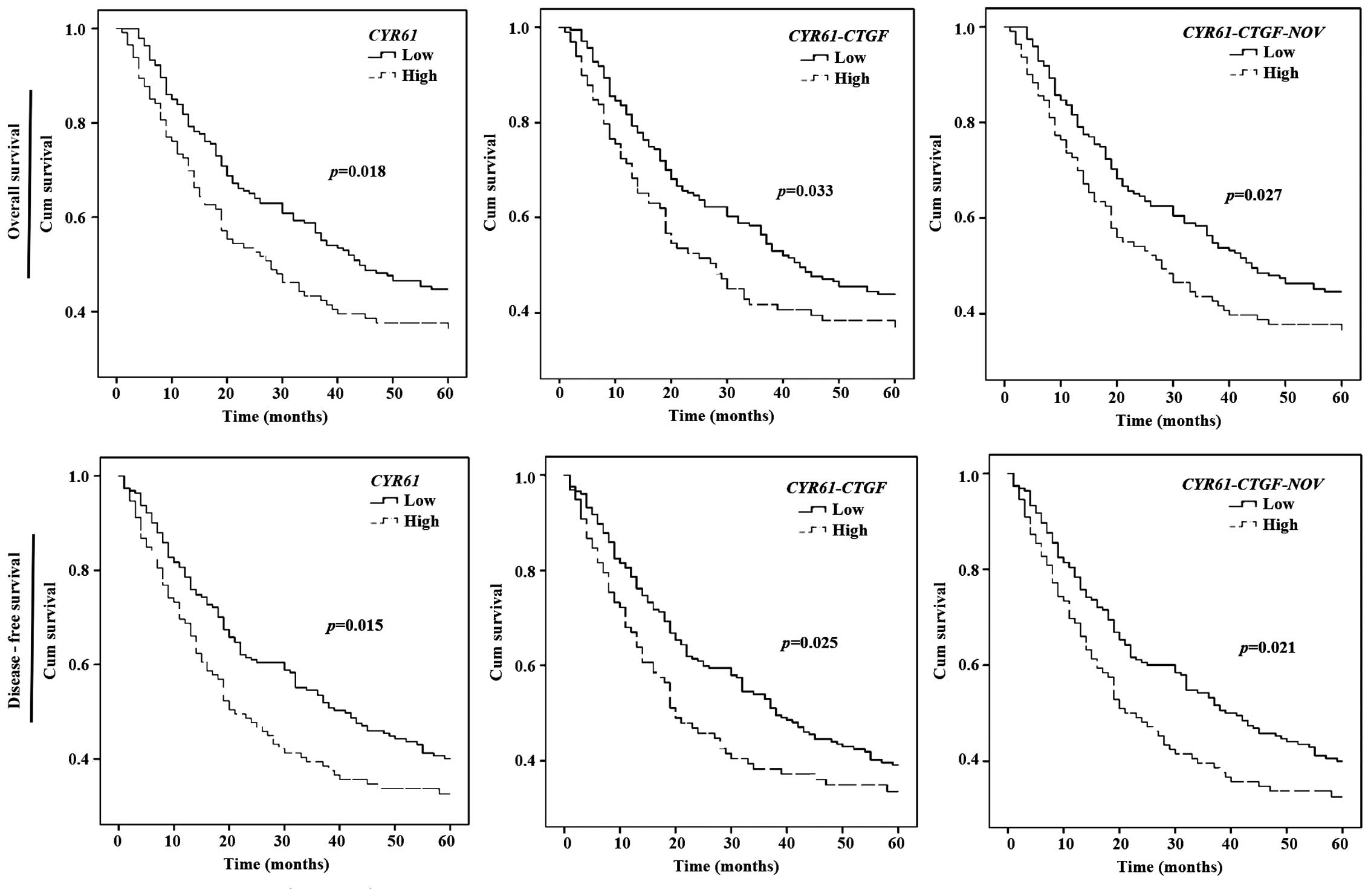
Kaplan-Meier survival curves revealed that GC
patients with a low CYR61 transcript level had longer
overall survival (OS) (p=0.018) and disease-free survival (DFS)
(p=0.015) than those with a higher CYR61 transcript level
(Fig. 1). Transcript levels of
CTGF and NOV exhibited no correlation with either OS
or DFS when individually analysed; however, analysis of the
combination of CYR61 and CTGF showed that patients
with lower transcript levels of these two genes had longer OS
(p=0.033) and DFS (p=0.025). Similarly, analysis of combined
CYR61, CTGF and NOV showed that patients with
higher transcript levels of all three genes had a poorer OS
(p=0.027) and DFS (p=0.021) compared to patients with lower
expression of all these genes.
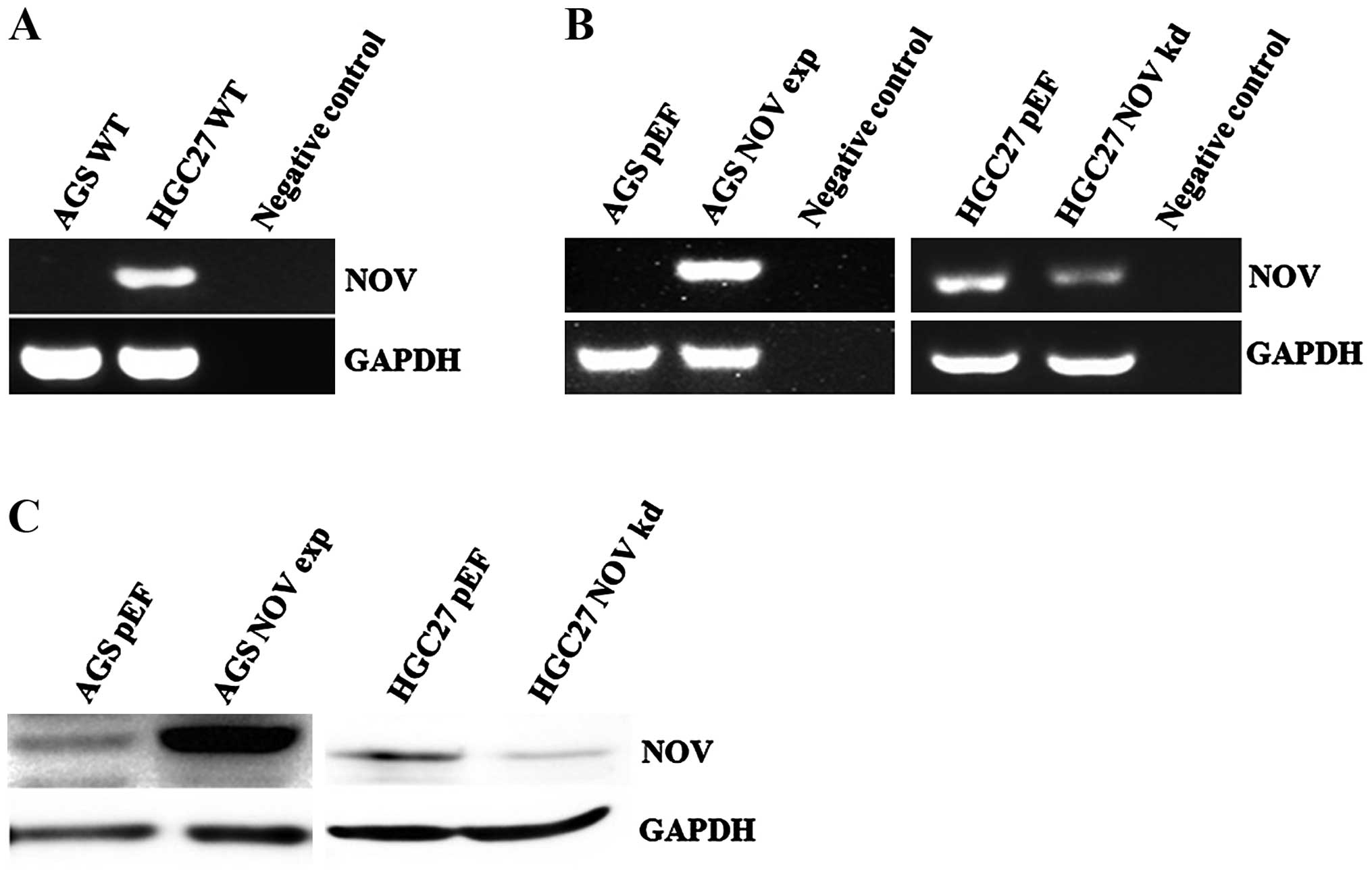
Knockdown and overexpression of NOV in GC
cells
The expression profile of NOV in AGS and HGC27 cell
lines was assessed using RT-PCR (Fig.
2A). For assessing the effect of NOV on cellular functions,
overexpression of NOV was performed in the AGS cells which had an
almost undetectable level of NOV as determined using PCR, while
knockdown of NOV was carried out in the HGC27 cells which highly
expressed NOV. The overexpression and knockdown of NOV in AGS and
HGC27 transfectants were confirmed using RT-PCR (Fig. 2B) and western blotting (Fig. 2C).
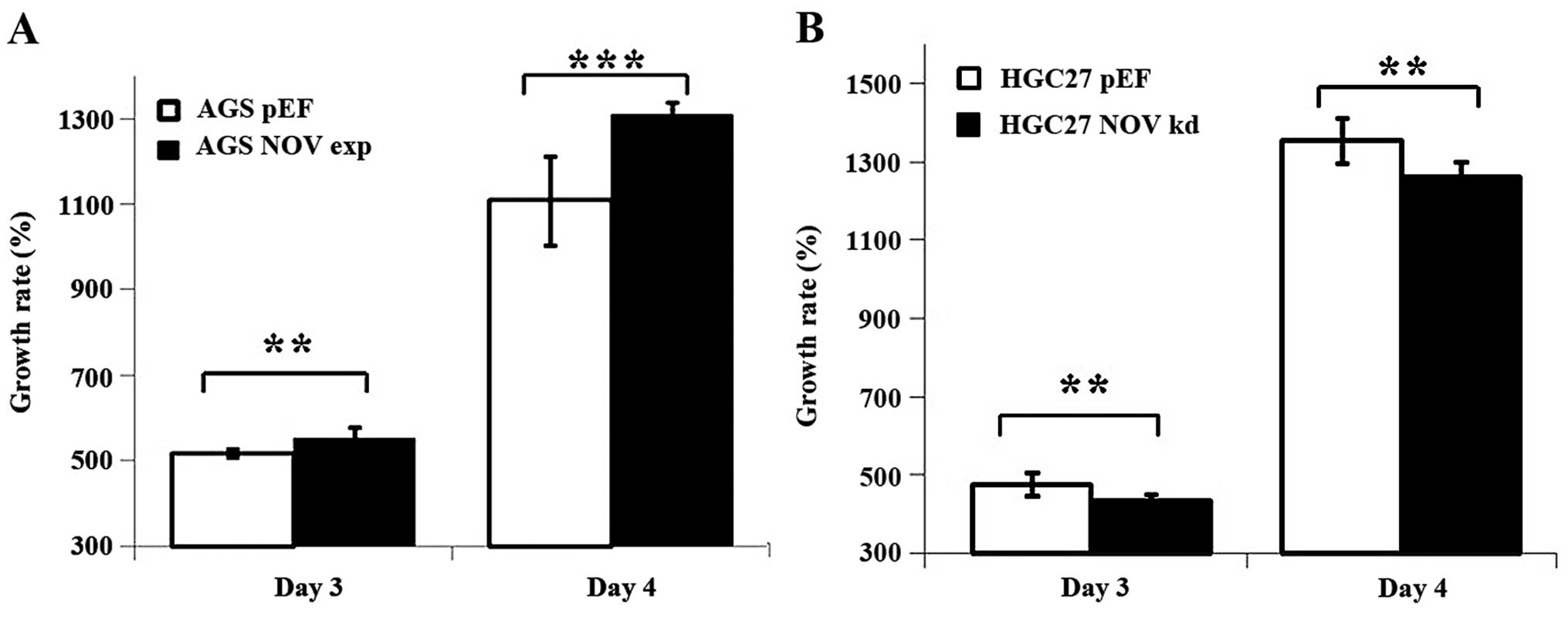
Effect of NOV knockdown and
overexpression on cell growth in vitro
Overexpression of NOV increased the growth of AGS
cells over the periods of 3 (p<0.01) and 4 days (p<0.001)
compared with the control cells. An opposite effect was observed in
the HGC27 NOV-knockdown cells over the periods of 3 (p<0.01) and
4 days (p<0.01) (Fig. 3).
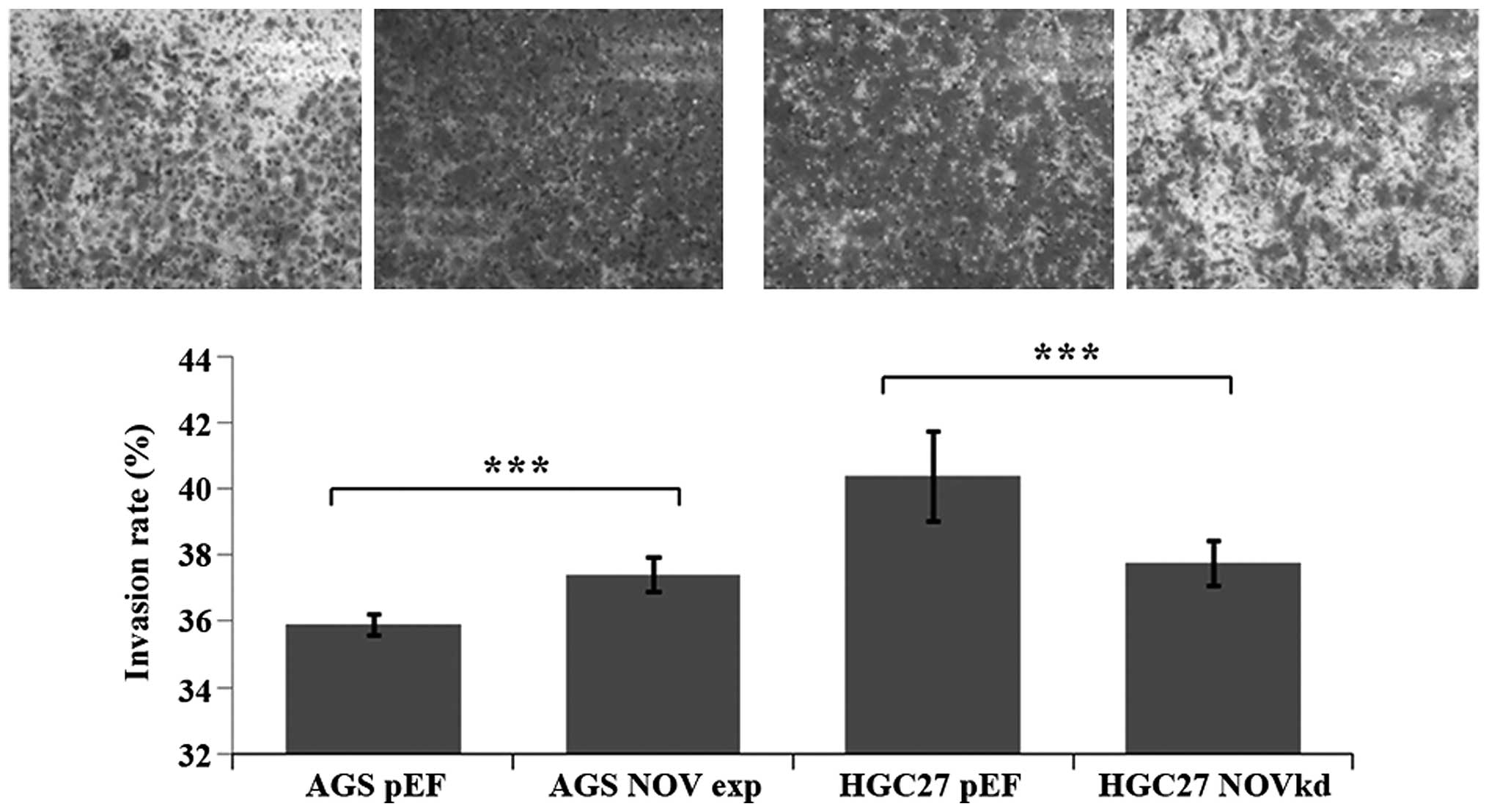
Effect of NOV knockdown and
overexpression on the invasion, adhesion and migration of GC
cells
Overexpression of NOV resulted in increased invasion
in the AGS cells, while NOV knockdown exhibited reduced
invasiveness in the HGC27 cells (p<0.001) (Fig. 4). Knockdown and overexpression of
NOV did not enhance or reduce the adhesion of the GC cells to
Matrigel in comparison with the control cells. A wound healing
assay was employed to determine the influence of NOV on cell
migration. There was no obvious effect observed in our experiments
(data not shown).
Discussion
This is the first study to assess the role played by
NOV (CCN3) in gastric cancer (GC). In the present study, we
determined the expression of NOV, along with CYR61 and CTGF
transcripts in a cohort of human GCs and paired adjacent normal
gastric tissues. Increased expression of NOV was observed in the GC
tissues compared with the expression noted in the adjacent normal
gastric tissues. Increased expression was found to be associated
with local invasion and poorer differentiation. Elevated expression
of NOV in GC is consistent with observations in other cancers. For
example, overexpression of NOV is evident in prostate cancer and
cervical cancer and is involved in tumourigenesis and disease
progression (19,20). Overexpression of NOV in cervical
cancers was found to be associated with lymph node metastases and
poorer prognosis of cervical cancer patients (20). This suggests that NOV is positively
involved in tumourigenesis and disease progression by regulating
morphological transformation and the invasiveness of GC cells. Our
previous study showed reduced expression of NOV in breast cancer
which was associated with poor prognosis and mortality of the
disease (15). Differential
expression patterns and roles can be played by NOV in different
malignancies which can be organ- or tissue-specific.
In the present study, we also determined the
transcript levels of CYR61 and CTGF in GC samples. Similarly,
overexpression of CYR61 transcripts was noted in GC although no
difference was observed for the expression of CTGF. No association
was clearly evident for elevated expression of CYR61 and disease
progression in the present cohort of GC cases. Overexpression of
CYR61 has been observed in many different cancers, including
prostate, breast, ovarian, endometrial and colorectal carcinomas
(11,12,33–35).
In contrast to the elevated expression of CYR61 in these
malignancies, reduced expression has also been noted in various
types of cancers, in particular GC which is associated with local
invasion (23). In the present
study, a trend of reduced expression of CYR61 was also noted in the
more advanced GCs appearing to be in line with observations made by
Maeta et al (23). A
controversy may still exist for its role in GC as experimental
evidence using GC cell lines indicates that CYR61 can promote
invasion, metastasis and angiogenesis in GC (29–31).
The present study also showed a link between higher CYR61
expression and poorer survival outcomes of patients with GC. This
was also reflected in the survival analysis of the combined
expression of CYR61 with CTGF, or CTGF and NOV (Fig. 1). CTGF and NOV alone or combined did
not exhibit any correlation with survival (data not shown). This
suggests that a more profound role is played by CYR61 in GC which
is also supported by studies of this molecule in GC (30,36).
In a comparison with NOV expression and corresponding association
between elevated NOV expression and local invasion and poorer
differentiation of GCs, the increased expression CYR61 exhibited
little implication with these clinicopathological features of the
disease apart from its correlation with poor prognosis. A larger
cohort of GC tissue samples may help to clarify this. Better
understanding of the molecular mechanisms of GC may shed light on
this issue in the near future.
Although the expression of CTGF transcripts in the
present cohort of GCs was not different from its expression in the
paired adjacent normal gastric tissues, its expression was
increased in more invasive tumours. This tends to concur with
observations from other studies focusing on this molecule and its
role in GC. Higher expression of CTGF in GC exhibits involvement in
local lymph node metastasis and also peritoneal metastasis
(37,38). Suppression of CTGF inhibits the
growth and invasion of GC cells, and also their peritoneal
dissemination (39). In addition to
its role in invasion and metastases, CTGF also promotes
angiogenesis to facilitate tumour growth (40).
In addition to the evaluation of NOV, CYR61 and CTGF
transcripts in the GC samples, we further examined the impact of
NOV on cellular functions in GC cell lines. The two GC cell lines
examined in the present study exhibited differential expression of
NOV, where NOV was highly expressed in the HGC27 cells and almost
absent in the AGS cells. This allowed us to establish contrasting
models, i.e. overexpression of NOV in AGS cells and knockdown of
NOV expression in HGC27 cells, for examining the consequent effect
on the cellular functions. NOV overexpression promoted the in
vitro proliferation of AGS cells while the knockdown resulted
in reduced proliferation of the HGC27 cells. A similar effect was
observed in regards to the invasion of these two GC cell lines.
These results indicate a positive role played by NOV in promoting
proliferation and invasion of GC which is consistent with its
increased expression in the GC tumour samples. Increased expression
of NOV has also been observed in other malignancies, such as
cervical and prostate cancer (19,20).
Certainly, an inhibitory effect on cellular functions has also been
noted for NOV in a variety of cancer cells. For example, NOV had an
anti-proliferative effect on glioblastoma cells by interfering with
S/G2 transition of the cell cycle leading to an accumulation at the
S phase (25). Gap junction protein
connexin 43 was also found to be involved in its inhibitory effect
on the proliferation of glioma cells (41). NOV has also been reported to
decrease the transcription and activation of matrix
metalloproteinases and suppress the invasion of melanoma cells
(42). However, our in vitro
experimental data indicate that NOV promotes both proliferation and
invasion of GC cells. Further investigation may shed light on the
underlying molecular mechanisms for such a differential impact.
In summary, the expression of NOV and CYR61 was
increased in GC. The elevated expression of CYR61 was associated
with poorer survival, and NOV promoted the proliferation and
invasion of GC cells.
Acknowledgments
The authors thank Cancer Research Wales for
supporting the present study. Dr Li and Dr Gao are recipients of
the Chinese Medical Research Scholarship of Cardiff University.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brock A, Krause S and Ingber DE: Control
of cancer formation by intrinsic genetic noise and
microenvironmental cues. Nat Rev Cancer. 15:499–509. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jia Q, Dong Q and Qin L: CCN: Core
regulatory proteins in the microenvironment that affect the
metastasis of hepatocellular carcinoma? Oncotarget. 7:1203–1214.
2016.
|
|
5
|
Chang CC, Lin BR, Wu TS, Jeng YM and Kuo
ML: Input of microenvironmental regulation on colorectal cancer:
Role of the CCN family. World J Gastroenterol. 20:6826–6831. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Riser BL, Barnes JL and Varani J: Balanced
regulation of the CCN family of matricellular proteins: A novel
approach to the prevention and treatment of fibrosis and cancer. J
Cell Commun Signal. 9:327–339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Perbal B: CCN proteins: A centralized
communication network. J Cell Commun Signal. 7:169–177. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Joliot A, Triller A, Volovitch M and
Prochiantz A: Are embryonic forms of NCAM homeobox receptors? C R
Acad Sci III. 314(Suppl 9): S59–S63. 1992.In French.
|
|
9
|
Kyurkchiev S, Yeger H, Bleau AM and Perbal
B: Potential cellular conformations of the CCN3 (NOV) protein. Cell
Commun Signal. 2:92004. View Article : Google Scholar
|
|
10
|
Desnoyers L: Structural basis and
therapeutic implication of the interaction of CCN proteins with
glycoconjugates. Curr Pharm Des. 10:3913–3928. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gery S, Xie D, Yin D, Gabra H, Miller C,
Wang H, Scott D, Yi WS, Popoviciu ML, Said JW, et al: Ovarian
carcinomas: CCN genes are aberrantly expressed and CCN1 promotes
proliferation of these cells. Clin Cancer Res. 11:7243–7254. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lv H, Fan E, Sun S, Ma X, Zhang X, Han DM
and Cong YS: Cyr61 is up-regulated in prostate cancer and
associated with the p53 gene status. J Cell Biochem. 106:738–744.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsai MS, Hornby AE, Lakins J and Lupu R:
Expression and function of CYR61, an angiogenic factor, in breast
cancer cell lines and tumor biopsies. Cancer Res. 60:5603–5607.
2000.PubMed/NCBI
|
|
14
|
Ladwa R, Pringle H, Kumar R and West K:
Expression of CTGF and Cyr61 in colorectal cancer. J Clin Pathol.
64:58–64. 2011. View Article : Google Scholar
|
|
15
|
Jiang WG, Watkins G, Fodstad O,
Douglas-Jones A, Mokbel K and Mansel RE: Differential expression of
the CCN family members Cyr61, CTGF and Nov in human breast cancer.
Endocr Relat Cancer. 11:781–791. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin BR, Chang CC, Chen RJ, Jeng YM, Liang
JT, Lee PH, Chang KJ and Kuo ML: Connective tissue growth factor
acts as a therapeutic agent and predictor for peritoneal
carcinomatosis of colorectal cancer. Clin Cancer Res. 17:3077–3088.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng YZ, Chen PP, Wang Y, Yin D, Koeffler
HP, Li B, Tong XJ and Xie D: Connective tissue growth factor is
overexpressed in esophageal squamous cell carcinoma and promotes
tumorigenicity through beta-catenin-T-cell factor/Lef signaling. J
Biol Chem. 282:36571–36581. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bennewith KL, Huang X, Ham CM, Graves EE,
Erler JT, Kambham N, Feazell J, Yang GP, Koong A and Giaccia AJ:
The role of tumor cell-derived connective tissue growth factor
(CTGF/CCN2) in pancreatic tumor growth. Cancer Res. 69:775–784.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maillard M, Cadot B and Ball RY, Maillard
M, Cadot B and Ball RY: Differential expression of the ccn3 (nov)
proto-oncogene in human prostate cell lines and tissues. Mol
Pathol. 54:275–280. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang T, Zhao C, Luo L, Xiang J, Sun Q,
Cheng J and Chen D: The clinical and prognostic significance of
CCN3 expression in patients with cervical cancer. Adv Clin Exp Med.
22:839–845. 2013.
|
|
21
|
Chen PP, Li WJ, Wang Y, Zhao S, Li DY,
Feng LY, Shi XL, Koeffler HP, Tong XJ and Xie D: Expression of
Cyr61, CTGF, and WISP-1 correlates with clinical features of lung
cancer. PLoS One. 2:e5342007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chien W, Kumagai T, Miller CW, Desmond JC,
Frank JM, Said JW and Koeffler HP: Cyr61 suppresses growth of human
endometrial cancer cells. J Biol Chem. 279:53087–53096. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maeta N, Osaki M, Shomori K, Inaba A,
Kidani K, Ikeguchi M and Ito H: CYR61 downregulation correlates
with tumor progression by promoting MMP-7 expression in human
gastric carcinoma. Oncology. 73:118–126. 2007. View Article : Google Scholar
|
|
24
|
Chien W, Yin D, Gui D, Mori A, Frank JM,
Said J, Kusuanco D, Marchevsky A, McKenna R and Koeffler HP:
Suppression of cell proliferation and signaling transduction by
connective tissue growth factor in non-small cell lung cancer
cells. Mol Cancer Res. 4:591–598. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bleau AM, Planque N, Lazar N, Zambelli D,
Ori A, Quan T, Fisher G, Scotlandi K and Perbal B:
Antiproliferative activity of CCN3: Involvement of the C-terminal
module and post-translational regulation. J Cell Biochem.
101:1475–1491. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thibout H, Martinerie C, Créminon C,
Godeau F, Boudou P, Le Bouc Y and Laurent M: Characterization of
human NOV in biological fluids: An enzyme immunoassay for the
quantification of human NOV in sera from patients with diseases of
the adrenal gland and of the nervous system. J Clin Endocrinol
Metab. 88:327–336. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tanaka S and Sugimachi K, Saeki H,
Kinoshita J, Ohga T, Shimada M, Maehara Y and Sugimachi K: A novel
variant of WISP1 lacking a Von Mayebrand type C module
overexpressed in scirrhous gastric carcinoma. Oncogene.
20:5525–5532. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tanaka S, Sugimachi K, Maehara S, Shimada
M and Maehara Y: A loss of function mutation in WISP3 derived from
microsatellite unstable gastric carcinoma. Gastroenterology.
125:1563–1564. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Babic AM, Kireeva ML, Kolesnikova TV and
Lau LF: CYR61, a product of a growth factor-inducible immediate
early gene, promotes angiogenesis and tumor growth. Proc Natl Acad
Sci USA. 95:6355–6360. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin MT, Zuon CY, Chang CC, Chen ST, Chen
CP, Lin BR, Wang MY, Jeng YM, Chang KJ, Lee PH, et al: Cyr61
induces gastric cancer cell motility/invasion via activation of the
integrin/nuclear factor-kappaB/cyclooxygenase-2 signaling pathway.
Clin Cancer Res. 11:5809–5820. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin MT, Kuo IH, Chang CC, Chu CY, Chen HY,
Lin BR, Sureshbabu M, Shih HJ and Kuo ML: Involvement of
hypoxia-inducing factor-1alpha-dependent plasminogen activator
inhibitor-1 up-regulation in Cyr61/CCN1-induced gastric cancer cell
invasion. J Biol Chem. 283:15807–15815. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang WG, Hiscox S, Hallett MB, Horrobin
DF, Mansel RE and Puntis MC: Regulation of the expression of
E-cadherin on human cancer cells by gamma-linolenic acid (GLA).
Cancer Res. 55:5043–5048. 1995.PubMed/NCBI
|
|
33
|
Zuo GW, Kohls CD, He BC, Chen L, Zhang W,
Shi Q, Zhang BQ, Kang Q, Luo J, Luo X, et al: The CCN proteins:
Important signaling mediators in stem cell differentiation and
tumorigenesis. Histol Histopathol. 25:795–806. 2010.PubMed/NCBI
|
|
34
|
Watari H, Xiong Y, Hassan MK and Sakuragi
N: Cyr61, a member of ccn (connective tissue growth
factor/cysteine-rich 61/nephroblastoma overexpressed) family,
predicts survival of patients with endometrial cancer of
endometrioid subtype. Gynecol Oncol. 112:229–234. 2009. View Article : Google Scholar
|
|
35
|
Monnier Y, Farmer P, Bieler G, Imaizumi N,
Sengstag T, Alghisi GC, Stehle JC, Ciarloni L, Andrejevic-Blant S,
Moeckli R, et al: CYR61 and alphaVbeta5 integrin cooperate to
promote invasion and metastasis of tumors growing in preirradiated
stroma. Cancer Res. 68:7323–7331. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei J, Yu G, Shao G, Sun A, Chen M, Yang W
and Lin Q: CYR61 (CCN1) is a metastatic biomarker of gastric cardia
adenocarcinoma. Oncotarget. Apr 20–2016.Epub ahead of print.
View Article : Google Scholar
|
|
37
|
Liu L, Li Z, Feng G, You W and Li J:
Expression of connective tissue growth factor is in agreement with
the expression of VEGF, VEGF-C, -D and associated with shorter
survival in gastric cancer. Pathol Int. 57:712–718. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu LY, Han YC, Wu SH and Lv ZH:
Expression of connective tissue growth factor in tumor tissues is
an independent predictor of poor prognosis in patients with gastric
cancer. World J Gastroenterol. 14:2110–2114. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jiang CG, Lv L, Liu FR, Wang ZN, Liu FN,
Li YS, Wang CY, Zhang HY, Sun Z and Xu HM: Downregulation of
connective tissue growth factor inhibits the growth and invasion of
gastric cancer cells and attenuates peritoneal dissemination. Mol
Cancer. 10:1222011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Inoki I, Shiomi T, Hashimoto G, Enomoto H,
Nakamura H, Makino K, Ikeda E, Takata S, Kobayashi K and Okada Y:
Connective tissue growth factor binds vascular endothelial growth
factor (VEGF) and inhibits VEGF-induced angiogenesis. FASEB J.
16:219–221. 2002.
|
|
41
|
Sin WC, Bechberger JF, Rushlow WJ and Naus
CC: Dose-dependent differential upregulation of CCN1/Cyr61 and
CCN3/NOV by the gap junction protein Connexin43 in glioma cells. J
Cell Biochem. 103:1772–1782. 2008. View Article : Google Scholar
|
|
42
|
Fukunaga-Kalabis M, Martinez G, Telson SM,
Liu ZJ, Balint K, Juhasz I, Elder DE, Perbal B and Herlyn M:
Downregulation of CCN3 expression as a potential mechanism for
melanoma progression. Oncogene. 27:2552–2560. 2008. View Article : Google Scholar
|