Introduction
Lung cancer is one of the fastest-growing morbidity
and mortality and the most serious threat tohuman health and life
due to malignant tumors. Lung cancer is the most important cause of
cancer-related deaths worldwide (1–3). It
has been reported that more than 160,000 patients died of lung
cancer which is more than from colon, prostate and even breast
cancer, in the US during 2013 (4).
At present, surgery, radiotherapy and chemotherapy have been used
in the treatment of lung cancer. Nevertheless, the long-term
survival rate is still very low (5). Research has also shown that most
patients with lung cancer had been smoking in the US (6), and the genetic damage of lung cancer
patients was mainly caused by smoking (7,8). The
expression levels of mRNA and protein among thousands of genes can
be used to analyze the molecular network among lung carcinogenesis
(9,10). At present, epidermal growth factor
receptor (EGFR) and echinoderm microtubule associated protein-like
4-anaplastic lymphoma kinase (EML4-ALK) fusion genes have been used
to detect lung cancer. Although many known genes and proteins have
provided a large amount of information for the treatment of lung
cancer, there are also a large number of unknown markers including
non-coding RNAs (ncRNAs) which may be crucial regulators of
cellular processes such as proliferation, gene regulation and
apoptosis and may also serve as novel biomarkers for the treatment
of lung cancer.
In recent years, the biomarker research has also
focused on non-coding RNAs (ncRNAs), particularly lncRNAs, which
are greater than 200 nt and most are transcribed by RNA polymerase
(Pol) II/Pol I, small are transcribed by RNA Pol III (11), playing an important role in the
regulation of gene expression (12–15).
In addition, there is growing evidence that lncRNA is related to
cell cycle and cell differentiation (16), apoptosis (17,18)
and chromatin remodeling (19–21).
It has also been shown that lncRNA expression are concerned with
the development of various types of cancer, such as liver (22), lung (23) and breast cancers (24). For example, maternally expressed
gene 3 (MEG3) is related to bladder cancer (25), and antisense non-coding RNA in the
INK4 locus (ANRIL) is relative to plexiform neurofibromas (26). For lung cancer, recent studies
predicted that MALAT1 was a critical regulator of lung cancer cells
(27), SCAL1 promoted lung cancer
by cigarette smoke (28), AK126698
produced drug resistance in lung cancer cells (29). Therefore, lncRNA is considered as an
important regulating factor for gene expression, disease as well as
cancers. However, the investigation into function and dysregulation
of lncRNA in cancer has only just begun; increasing research is
urgently needed to lead to a deeper understanding for the lncRNA
regulatory network. In our study, we found that lncRNA-SNHG7 was
obviously upregulated in lung cancer tissues compared to adjacent
noncancerous tissues. Furthermore, silence of lncRNA-SNHG7 by siRNA
suppressed cell proliferation, migration and invasion and
accelerated apoptosis of A594 cells in vitro.
The present study showed, lncRNA-SNHG7 markedly
inhibited apoptosis of A594 cells. Fas apoptotic inhibitory
molecule 2 (FAIM2), as a 35 kDa membrane protein, is an
anti-apoptotic protein conserved in evolution and known as the
Lifeguard (LFG) family which is a distinct gene family of
apoptotic-related genes (30). A
previous study demonstrated that FAIM2 interacted with Fas upstream
of Fas-associated death domain containing protein (FADD) and
suppressed apoptosis (31). The
high expression of FAIM2 enhanced resistance to Fas regulated
apoptosis (32). Therefore, FAIM2
was used as apoptosis detection index. The present study found that
the expression levels of FAIM2 also increased in lung cancer
tissues. In addition, lncRNA-SNHG7 was of positive relevance with
FAIM2 in human lung cancer tissues. Silence of FAIM2 by siRNA
suppressed cell proliferation, migration, and invasion and
accelerated apoptosis of A594 cells in vitro.
In the present study, we found that the expression
levels of lncRNA-SNHG7 and FAIM2 obviously increased, and
lncRNA-SNHG7 was of positive relevance with FAIM2 in lung cancer
tissues. Silence of lncRNA-SNHG7 and silence of FAIM2 by siRNA
suppressed cell proliferation, migration and invasion, and also
accelerated the apoptosis of A594 cells in vitro,
respectively. These results indicate that the lncRNA-SNHG7 network
may lead to potential therapy for lung cancer in the future.
Materials and methods
Patients and clinical specimens
In the present study, we collected lung cancer and
matched adjacent non-cancerous tissues samples (5 cm from the edge
of the cancer as assessed by a pathologist) from patients who had
undergone treatment for lung cancer between 2013 and 2015 at The
First Affiliated Hospital of Southern Medical University,
Guangzhou, China. The lung cancer histological diagnosis was
confirmed according to the World Health Organization (WHO). Written
informed consent was provided from each patient or her/his
guardian. All tissue samples were washed with sterile
phosphate-buffered saline (PBS) and was immediately saved at −80°C
until use.
Cell lines
Three human lung cancer cell lines (H125, 95D and
A594) and human bronchial epithelial cells (BEAS-2B) were obtained
from the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Cells were cultured according to the ATCC
protocols, BEAS-2B cells were cultured in bronchial epithelial
growth medium (BEGM; PromoCell GmbH, Heidelberg, Germany). H125,
95D and A594 were cultured in RPMI-1640 medium (Invitrogen,
Carlsbad, CA, USA), and all the media contained 10% fetal bovine
serum (FBS) and 100 U/ml penicillin and 1 µg/ml streptomycin
(both from Invitrogen). Then, the cells were cultured in an
incubator with 5% CO2 at 37°C.
siRNA transfection
According to the protocol, we performed the siRNA
transfection using Lipofectamine 3000 (Thermo Fisher Scientific,
Rockford, IL, USA). siRNA oligomers were synthesized by Gemma
(Shanghai, China). lncRNA-SNHG7_siRNA sequence was:
5′-GCUGGAAUAAAGAGUAACAUU-3′; and FAIM2_siRNA sequence was:
5′-CUGGCUCCAUGCAGUUUAUUU-3′). A594 cells were seeded into 6-well
plates (200,000 cells/well), and then transduced with control and
lncRNA-SNHG7_siRNA or control and FAIM2_siRNA (final concentration
50 µM) using Lipofectamine RNAiMax (Life Technologies, Inc.,
Grand Island, NY, USA). Forty-eight hours after transfection, cells
were used forthe tests.
RNA preparation and reverse
transcription
According to the manufacturer's instructions, total
RNA was extracted from cells or the lung cancer tissues samples by
the TRIzol reagent (Invitrogen). RNA (1.0 µg) was used as a
template to synthesize corresponding cDNA with random primers using
a RevertAid First Strand cDNA synthesis kit (Thermo Fisher
Scientific).
Quantitative real-time reverse
transcription PCR (qRT-PCR) assay
qRT-PCR was completed using SYBR-Green PCR Master
Mix kit (Takara, Shiga, Japan) and an ABI 7500 Real-Time PCR system
(Applied Biosystems, Warrington, UK). The primers of endogenous
housekeeping gene (GAPDH chosen as internal loading control),
lncRNA-SNHG7 and FAIM2 were designed. The sequences of GAPDH
primers were: 5′-GTCAGCCGCATCTTCTTTTG-3′ (sense) and 5′-GC
GCCCAATACGACCAAATC-3′ (antisense). The sequences of lncRNA-SNHG7
primers were: 5′-GTTGGGGTGTTGGCATTCTTGTT-3′ (sense) and
5′-TGGTCAGCCTGGTCACTCTGG-3′ (antisense). The sequences of FAIM2
primers were: 5′-GGCGTGCTCTTCGTGCTTC-3′ (sense) and
5′-TGGCGTCGGTACCCATCA-3′ (antisense). The expression level of
lncRNA-SNHG7 and FAIM2 mRNA was analyzed relative to the GAPDH mRNA
level. All results are shown as the mean ± SD of three independent
experiments.
Cell proliferation
Cell proliferation was measured using a Cell
Counting Kit-8 (CCK-8 Kit; Beyotime Institute of Biotechnology,
Jiangsu, China), transfected were seeded into 96-well plates at a
density of 2×103 cells/well with 100 µl RPMI-1640
medium (with 10% FBS) and cultured for 24, 48 and 72 h,
respectively. The 10 µl of CCK-8 solution was put into the
well at each time point and incubated for 4 h at 37°C, and then the
absorbance at 450 nm was measured using a microplate reader. Each
experimental condition was detected in quintuplicates, and the
experiments were repeated at least three times.
Flow cytometric analysis of
apoptosis
The apoptotic cell death rate was analyzed by flow
cytometry and double staining using propidium iodide (PI) and the
Annexin V-FITC staining kit (BD Biosciences, San Diego, CA, USA)
according to the manufacturer's instructions. A594 cells were
digested with trypsin, dispersed, centrifuged, collected, washed
with Dulbecco's modified Eagle's medium (DMEM) medium and cold PBS
and resuspended with 1X binding buffer at a concentration of
1×106 cells/ml. PI (5 µl) and Annexin V-FITC were
added into A594 cell buffer solution (1×105 cells) and
incubated for 15 min at RT in the dark, and then 400 µl 1X
binding buffer was put in each sample tube and analyzed by a BD
FACSCalibur cytometer (BD Biosciences). Data were analyzed using
FlowJo software version 8.8.6 (Tree Star Inc., Ashland, OR, USA).
A594 cells transfected with NC-siRNA were used as the negative
control (NC).
Migration and invasion assays
The migration and invasive capacity of the lung
cancer cells in vitro were detected in 24-well Transwell
cell culture chamber with 8-µm pores (both from Corning
Costar Corp., Cambridge, MA, USA) according to the manufacturer's
instructions. Cells were transfected with 50 µM lncRNA-SNHG7
siRNA, or control and FAIM2 siRNA for 48 h, and then 200 µl
A594 cells (2.5×105/100 µl) were seeded in
serum-free media to 8-µm pore cell culture inserts, and
complete medium with 10% FBS was added to the lower chambers as a
chemoattractant. Cells were incubated for 24 h at 37°C, and the
cells with migration ability spread through the chamber.
Paraformaldehyde (4%) was used to fix the cells and 0.1% crystal
violet solution was used to stain the lower chamber cells. The
cells above the upper surface that did not migrate were cleared
with a cotton swab, but did not touch the lower layer of the
membrane. The number of migratory cells was confirmed through
counting five areas of constant size/well under the microscope
using a 20× objective. For the invasion assays, the polycarbonate
membrane of Transwell inserts were covered with 10 µl 1:3
diluted Matrigel (BD Biosciences), and then were dried at 37°C for
30 min. All experiments were completed in triplicate.
Western blot analysis
A594 cell homogenates were obtained with lysis
buffer containing a protease inhibitor cocktail (P8340;
Sigma-Aldrich, St. Louis, MO, USA), and lung cancer and matched
adjacent non-cancerous tissue homogenates were obtained with RIPA
buffer (Pierce/Thermo Fisher Scientific, Lafayette, CO, USA)
containing a protease inhibitor cocktail. A BCA Protein Assay kit
(Thermo Fisher Scientific) was used to measure the concentrations
of total proteins. Proteins (30 µg) were separated using 8%,
12% Tris/glycine SDS/PAGE gels according to the molecular weight of
the objective proteins, and then proteins were transferred to
polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA,
USA). The PVDF membranes were blocked with 5% skim milk (BD
Biosciences) in 1X Tris-buffered saline including Tween-20 (TBST)
for 2 h at room temperature, the transferred membranes were
incubated with primary antibodies with a proper dilution at 4°C
overnight. Next day, the transferred membranes were washed with 1X
Tris-buffered saline including Tween-20 three times, incubated with
horseradish peroxidase-conjugated secondary antibodies with a
proper dilution for 1 h at room temperature, and then washed with
1X Tris-buffered saline including Tween-20 three times, again. The
experimental results were detected with the enhanced
chemiluminescence (ECL) substrate kit and the enhanced
chemiluminescence detection system (both from Amersham Biosciences,
Piscataway, NJ, USA). The anti-human FAIM2 antibody (rabbit, 1:500;
Sigma-Aldrich, Taufkirchen, Germany) which was used in the present
study is a monoclone antibody. The rabbit anti-GAPDH monoclone
antibody (1:4,000; Cell Signaling Technology, Beverly, MA, USA) was
used as an internal control.
Statistical analysis
Statistical significance was analyzed using GraphPad
(GraphPad Prism Software, La Jolla, CA, USA) and the SPSS 15.0
software [analysis of variance and Student's t-test or one-way
analysis of variance (ANOVA)]. The data are presented as the means
± standard deviation (SD). P<0.05 was considered significant.
P<0.05, P<0.01 and P<0.001.
Results
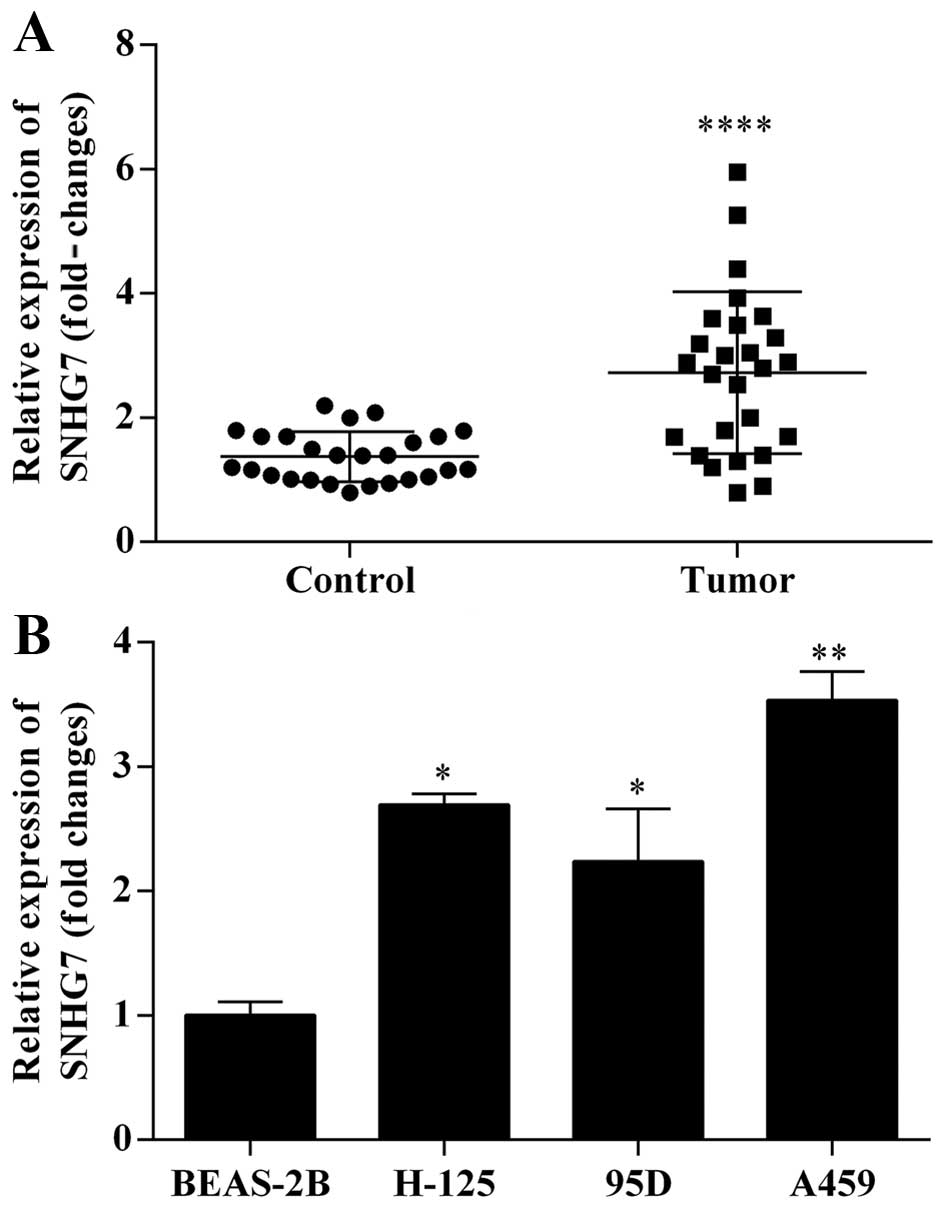
High expression of lncRNA-SNHG7 mRNA in
lung cancer tissues and lung cancer cell lines
In order to investigate the expression of
lncRNA-SNHG7 mRNA in lung cancer tissues, we adopted qRT-PCR assay
to analyze the expression level of lncRNA-SNHG7 mRNA in lung cancer
tissues relative to adjacent non-cancerous tissues. The results
indicated that lncRNA-SNHG7 mRNA was obviously upregulated in lung
cancer tissues compared to adjacent non-cancerous tissues (Fig. 1A). Furthermore, we analyzed the
effect of lncRNA-SNHG7 mRNA in human bronchial epithelial cells
(BEAS-2B) and three human lung cancer cell lines (H125, 95D and
A594). qRT-PCR was used to measure the expression level of
lncRNA-SNHG7 mRNA in the cell lines above. lncRNA-SNHG7 mRNA
markedly increased in three human lung cancer cell lines (H125, 95D
and A594) compared to human bronchial epithelial cells (BEAS-2B).
Among the three human lung cancer cell lines, the expression level
of lncRNA-SNHG7 mRNA in A594 cells was the most remarkable
(Fig. 1B). Thus, we chose the A594
cells as target cells.
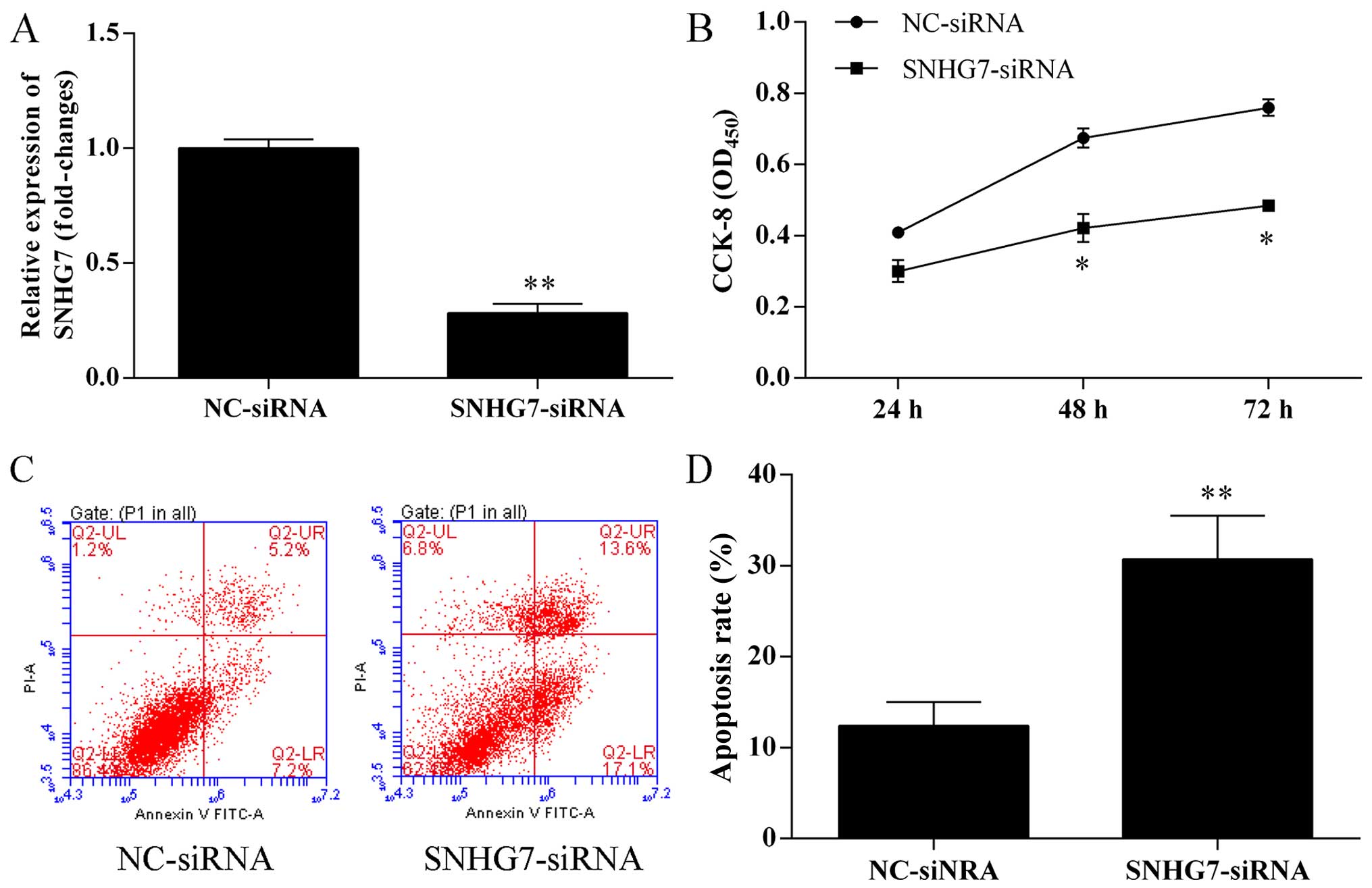
Silence of lncRNA-SNHG7 expression by
siRNA inhibits proliferation, migration and invasion, and promotes
apoptosis of A594 cells
A594 cells were transfected with negative control
(NC) siRNAs and lncRNA-SNHG7 siRNA for 48 h, respectively. The
expression effects of silence of lncRNA-SNHG7 mRNA expression by
siRNA in A594 cells were measured by qRT-PCR. As shown in Fig. 2A, the expression of lncRNA-SNHG7 was
markedly suppressed through lipofectin transfection compared to NC.
CCK-8 assay was used to detected cell proliferation ability. As
shown in Fig. 2B, the proliferation
ability of A594 cells transfected with lncRNA-SNHG7 siRNA was
significantly suppressed compared with the control group
(P<0.05). Apoptotic cell death was detected by flow cytometric
analysis with Annexin V-FITC and PI staining in A594 cells
transfected with lncRNA-SNHG7 siRNA for 48 h. Knockdown of
lncRNA-SNHG7 in A594 cells by siRNA accelerated apoptosis of A594
cells. As shown in Fig. 2C and D,
the apoptosis cells percentage of A594 cells transfected with
lncRNA-SNHG7 siRNA was 30.7%, while the control group was 12.4%.
Thus, the apoptotic cell percentage of in A594 cells with knockdown
of lncRNA-SNHG7 by siRNA increased 1.48 times. Transwell assay was
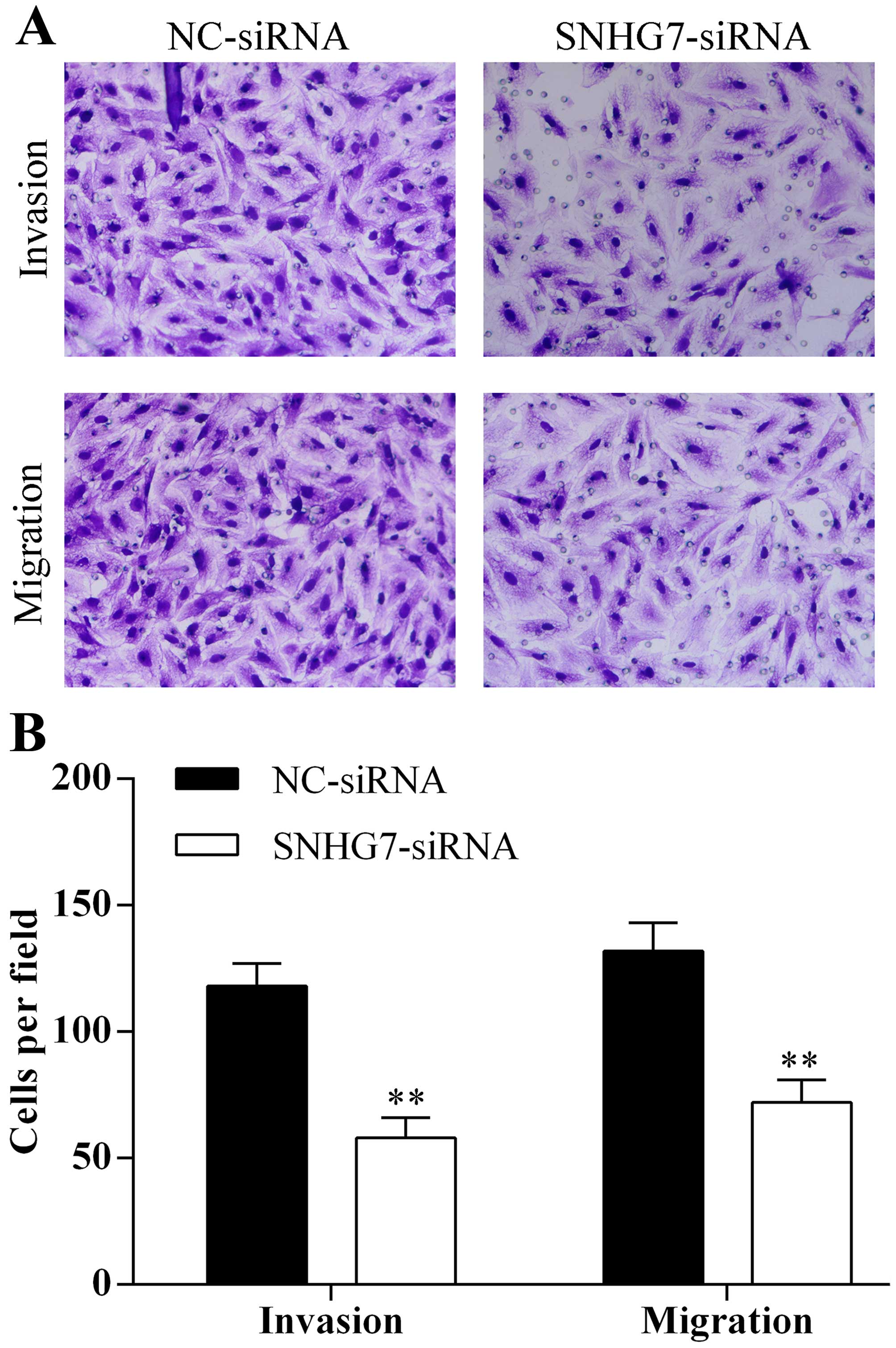
used to detect migration and invasion abilities of A594 cells
transfected with lncRNA-SNHG7 siRNA for 48 h. Knockdown of
lncRNA-SNHG7 in A594 cells by siRNA decreased the ability of cell
migration and invasion (Fig. 3A and
B). Therefore, knockdown of lncRNA-SNHG7 in A594 cells
inhibited the capacity of proliferation, migration and invasion and
promoted the capacity of apoptosis of A594 cells.
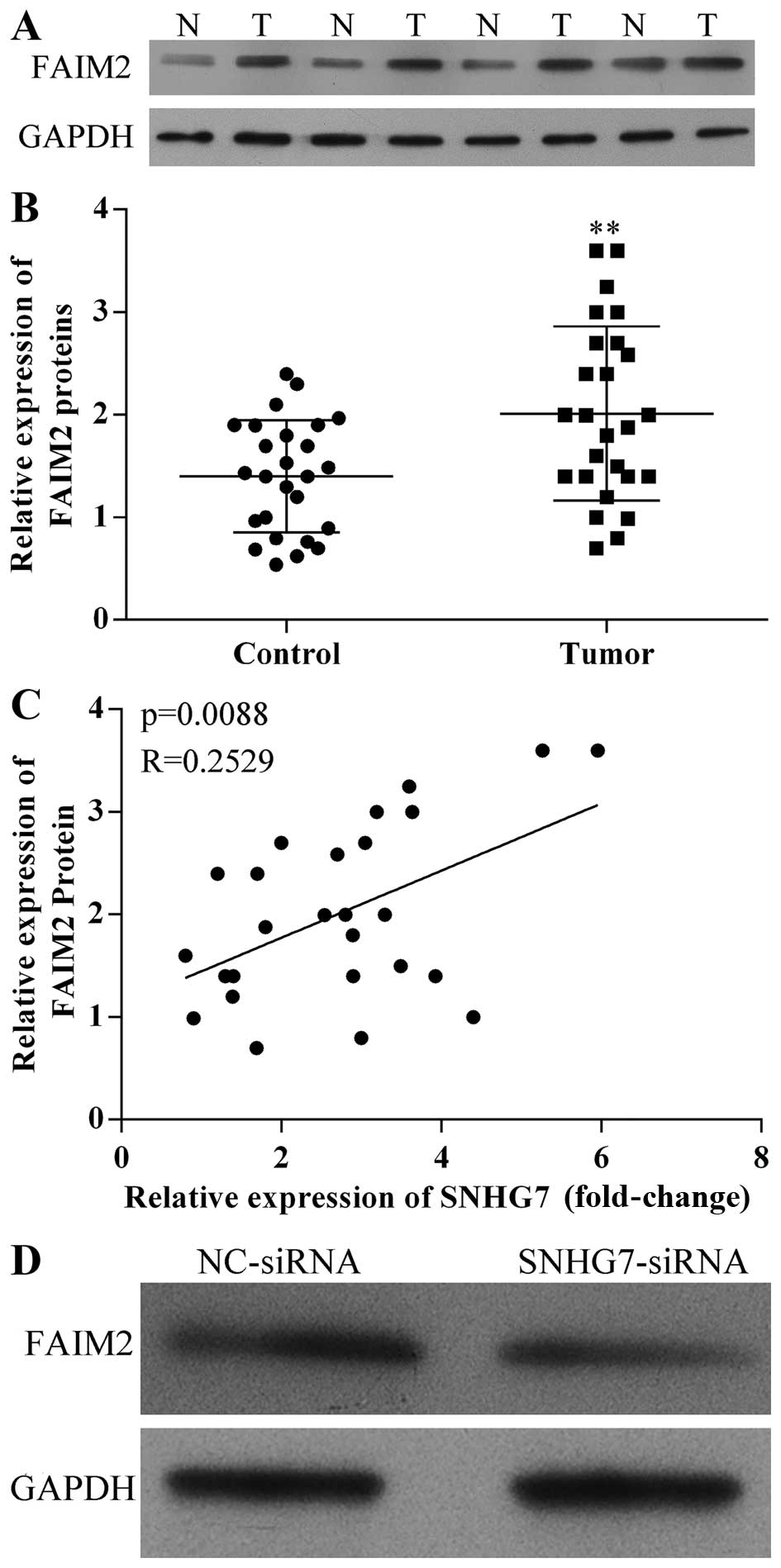
lncRNA-SNHG7 physically interacts with
FAIM2 in lung cancer clinical specimens and A594 cells
In our research, we demonstrated that lncRNA-SNHG7
inhibited the capacity of apoptosis of A594 cells. Therefore,
FAIM2, an inhibitor of the Fas signaling pathway, was studied in
lung cancer. In order to investigate the expression level of FAIM2
in lung cancer tissues, western blotting was used to detect the
expression levels of FAIM2 proteins in human lung cancer and paired
adjacent non-cancerous tissues from four random lung cancer
patients. As shown in Fig. 4A,
FAIM2 protein was higher expressed in lung cancer tissues. qRT-PCR
was used to measure the expression levels of FAIM2 mRNA in human
lung cancer tissues. FAIM2 mRNA was also higher expression in lung
cancer tissues (Fig. 4B).
Furthermore, to investigate the correlation between
lncRNA-SNHG7 and FAIM2 in human lung cancer tissues qRT-PCR results
were analyzed. As shown in Fig. 4C,
lncRNA-SNHG7 was of positive relevance with FAIM2 gene expression
in human lung cancer tissues when A594 cells were transfected with
FAIM2 siRNA for 48 h. The expression level of FAIM2 protein
declined relative to NC (Fig. 4C).
Therefore, lncRNA-SNHG7 increased the expression level of FAIM2 in
lung cancer clinical specimens and A594 cells.
Silence of FAIM2 expression by siRNA
inhibits proliferation, migration and invasion, and promotes
apoptosis of A594 cells
Based on previous results that positive correlation
between lncRNA-SNHG7 and FAIM2 gene expression was shown in human
lung cancer tissues, in addition, lncRNA-SNHG7 can promote the
expression level of FAIM2 in A594 cells. We further detected the
effects of FAIM2 on A594 cell growth, migration, invasion and
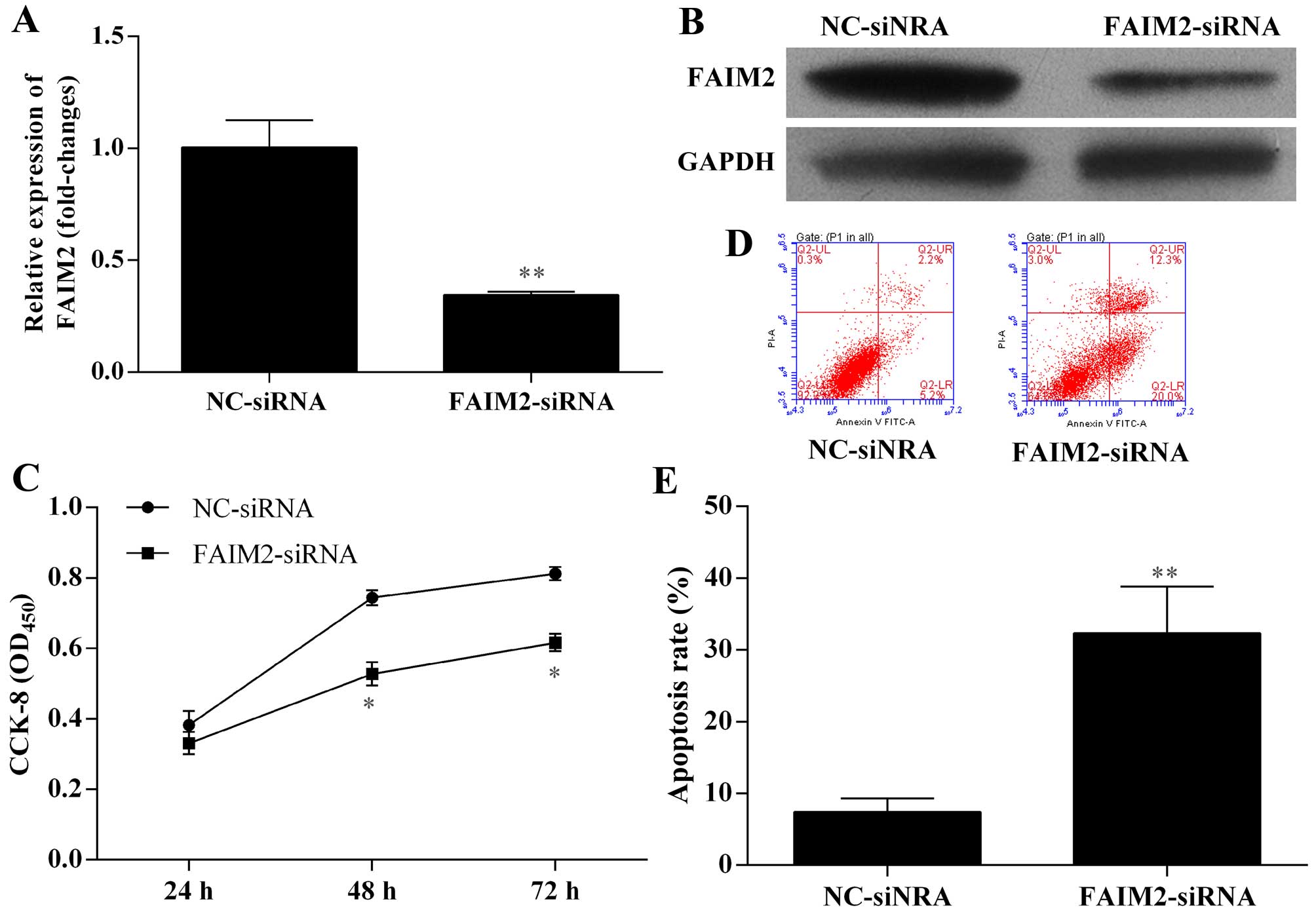
apoptosis. A594 cells were transfected with NC siRNAs and FAIM2
siRNA for 48 h, respectively. The expression effects of silence of
FAIM2 mRNA and protein expression by siRNA in A594 cells were
performed by qRT-PCR and western blotting. The expression of FAIM2
was markedly suppressed through transfecting with FAIM2 siRNA
compared to NC (P<0.01) (Fig. 5A and
B). The proliferation ability of A594 cells transfected with
FAIM2 siRNA was measured by CCK-8 assay. The proliferation ability
of A594 cells transfected with FAIM2 siRNA was significantly
decreased compared with the control group (P<0.05) (Fig. 5C). Flow cytometric analysis was used
to detect the apoptotic cell death of A594 cells transfected with
FAIM2 siRNA with Annexin V-FITC and PI staining. We found that the
apoptotic cell percentage of A594 cells transfected with FAIM2
siRNA was 32.3%, while the control group was 7.4%. Thus, the
apoptotic cell percentage of A594 cells with knockdown of FAIM2 by
siRNA increased 3.36 times (Fig. 5D and
E). Thus, we indicated that silence of FAIM2 expression by
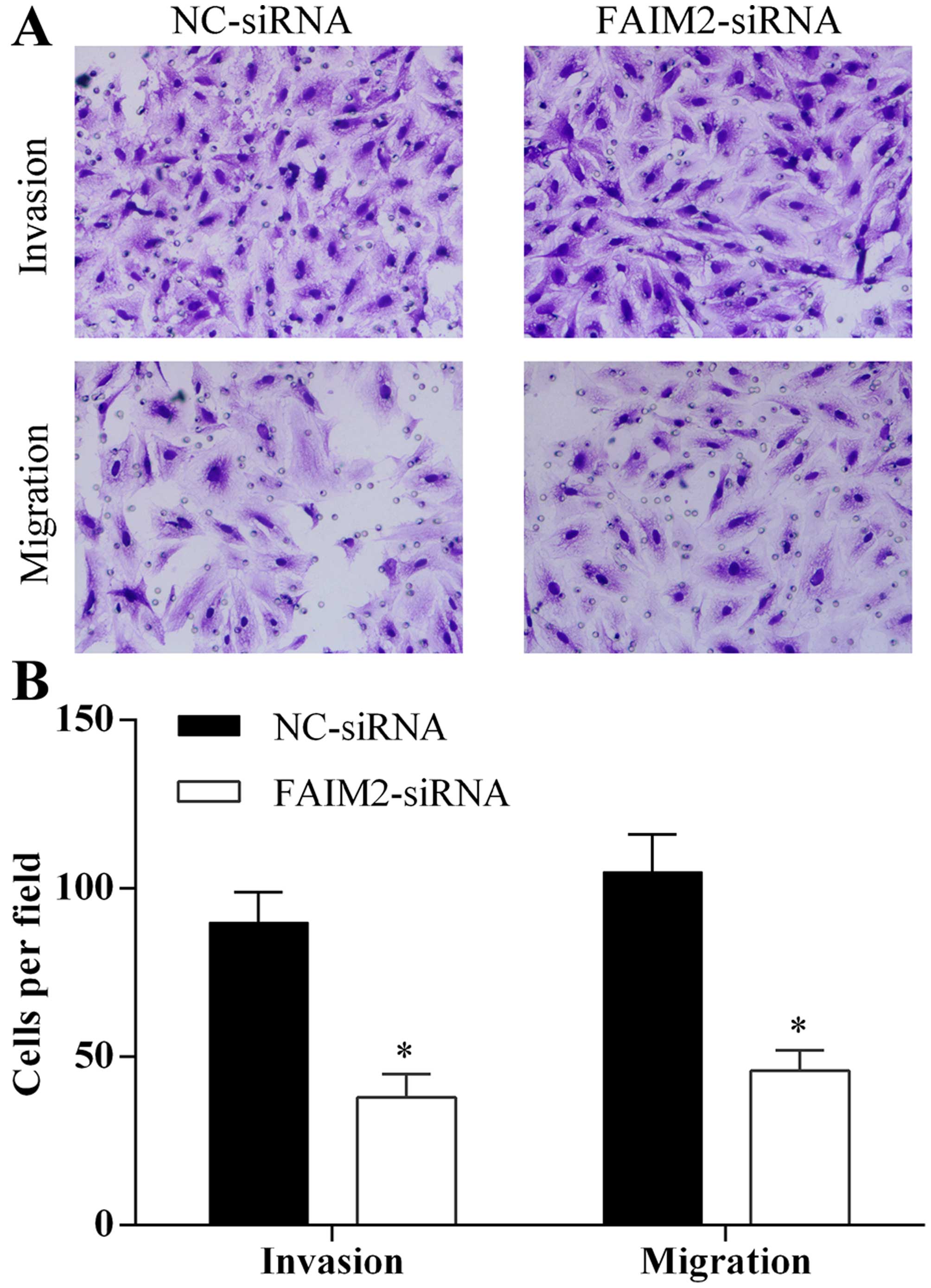
siRNA increased apoptosis of A594 cells. Furthermore, we evaluated
the ability of migration and invasion of A594 cells transfected
with FAIM2 siRNA using Transwell assay. Silence of FAIM2 expression
by siRNA in A594 cells reduced the ability of cell migration and
invasion (Fig. 6A and B).
Therefore, we indicated that knockdown of FAIM2 in A594 cells
reduced the capacity of proliferation, migration and invasion and
enhanced the capacity of apoptosis of A594 cells.
Discussion
The crucial results of the present study were that
lncRNA-SNHG7 was highly expressed in lung cancer tissues relative
to adjacent non-cancerous tissues. FAIM2 was also overexpressed in
lung cancer tissues. lncRNA-SNHG7 was associated with FAIM2 and
there was a positive correlation between them in lung cancer.
Silence of lncRNA-SNHG7 by siRNA repressed the level of FAIM2
protein in A594 cells. Silence of lncRNA-SNHG7 and silence of FAIM2
by siRNA repressed the level of FAIM2 protein and suppressed cell
proliferation, migration and invasion and accelerated apoptosis of
A594 cells in vitro, respectively. Thus, our results showed
that lncRNA-SNHG7 promotes the proliferation, migration and
invasion and inhibits apoptosis of lung cancer cells by enhancing
the FAIM2 expression and may be regarded as a potential therapeutic
target for lung cancer.
Increasing number of studies has found that lncRNAs
are connected with multiple genetic phenomena, such as
transcriptional regulations, DNA methylation and chromatin
remodeling (14,33,34).
Increasing evidence indicates that lncRNAs are interacted with
genes, proteins or chromatin remodeling to influence the expression
levels of genes (35,36). LncRNAs which are generated in
intronic, intergenic, antisense loci or overlapping regions play a
great role in diverse gene regulatory functions (37,38).
At present, numerous studies have demonstrated that lncRNAs are
involved in tumor carcinogenesis since it can accelerate cell
proliferation through controlling correlative proteins (39–41).
In the present study, we found that the expression levels of
lncRNA-SNHG7 mRNA and protein obviously increased in lung cancer
tissues compared to adjacent non-cancerous tissues. lncRNA-SNHG7
promoted the expression of FAIM2 protein and then inhibited A594
cell apoptosis. We also showed that silence of lncRNA-SNHG7 by
siRNA suppressed cell proliferation, migration, and invasion and
accelerated apoptosis of A594 cells in vitro. These results
suggest that lncRNA-SNHG7 is an important molecule for tumor
development and may become a potential biomarker for the treatment
of lung cancer.
FAIM2, as a gene of the LFG family, will have very
important effect in protecting cells against apoptosis by directly
bonding the Fas receptor (31,42).
It has been discovered that LFG was overexpressed in the mostly
neuron and hippocampus cells (31).
Recent studies have shown that expression of the FAIM2 was
interrelated to high fat (43) and
the different methylation levels of the FAIM2 promoter was markedly
related to obesity (44). FAIM2
gene is associated with obesity (45), simultaneously, obesity is partly
related to tobacco smoking which causes the development of cancer.
However, the functional mechanism of FAIM2 is not entirely clear in
lung cancer. In the present study, we have demonstrated that the
expression level of FAIM2 mRNA increased in lung cancer tissues
compared to adjacent non-cancerous tissues. Furthermore, positive
correlation between lncRNA-SNHG7 and FAIM2 gene expression in human
lung cancer tissues and silence of lncRNA-SNHG7 expression by siRNA
reduced the expression of FAIM2 proteins in A594 cells. We also
found that silence of FAIM2 expression by siRNA inhibited
proliferation, migration and invasion and promoted apoptosis of
A594 cells. These results suggest that FAIM2 also is an important
molecule for tumor development.
In brief, the present study demonstrated that
lncRNA-SNHG7 as an oncogene promoting proliferation, migration and
invasion, and inhibiting apoptosis of lung cancer cells by
enhancing the FAIM2 expression. This finding shows that
lncRNA-SNHG7 is a momentous molecule for tumor progression and
provides significant potential to develop new therapies to prevent
or treat lung cancer.
References
|
1
|
Parkin D, Whelan S, Ferlay J, Teppo L and
Thomas D: Cancer Incidence in Five Continents. VIII. IARC
Publications; pp. 1552002
|
|
2
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Malarcher AM, Schulman J, Epstein LA, Thun
MJ, Mowery P, Pierce B, Escobedo L and Giovino GA: Methodological
issues in estimating smoking-attributable mortality in the United
States. Am J Epidemiol. 152:573–584. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Organization WH: International statistical
classification of diseases and related health problems. 1. World
Health Organization; 2004
|
|
8
|
Travis WD, Brambilla E, Muller-Hermelink
HK and Harris CC: Pathology and Genetics of Tumours of the Lung,
Pleura, Thymus and Heart. IARC Publications; 2004
|
|
9
|
Granville CA and Dennis PA: An overview of
lung cancer genomics and proteomics. Am J Respir Cell Mol Biol.
32:169–176. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meyerson M and Carbone D: Genomic and
proteomic profiling of lung cancers: Lung cancer classification in
the age of targeted therapy. J Clin Oncol. 23:3219–3226. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bierhoff H, Schmitz K, Maass F, Ye J and
Grummt I: Noncoding transcripts in sense and antisense orientation
regulate the epigenetic state of ribosomal RNA genes. Cold Spring
Harb Symp Quant Biol. 75:357–364. 2010. View Article : Google Scholar
|
|
12
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Loewer S, Cabili MN, Guttman M, Loh YH,
Thomas K, Park IH, Garber M, Curran M, Onder T, Agarwal S, et al:
Large intergenic non-coding RNA-RoR modulates reprogramming of
human induced pluripotent stem cells. Nat Genet. 42:1113–1117.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu X, Li D, Zhang W, Guo M and Zhan Q:
Long non-coding RNA gadd7 interacts with TDP-43 and regulates Cdk6
mRNA decay. EMBO J. 31:4415–4427. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lakhotia SC: Long non-coding RNAs
coordinate cellular responses to stress. Wiley Interdiscip Rev RNA.
3:779–796. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Paralkar VR and Weiss MJ: A new 'Linc'
between noncoding RNAs and blood development. Genes Dev.
25:2555–2558. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saxena A and Carninci P: Long non-coding
RNA modifies chromatin: Epigenetic silencing by long non-coding
RNAs. BioEssays. 33:830–839. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kotake Y, Nakagawa T, Kitagawa K, Suzuki
S, Liu N, Kitagawa M and Xiong Y: Long non-coding RNA ANRIL is
required for the PRC2 recruitment to and silencing of
p15INK4B tumor suppressor gene. Oncogene. 30:1956–1962.
2011. View Article : Google Scholar
|
|
21
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang F, Zhang L, Huo XS, Yuan JH, Xu D,
Yuan SX, Zhu N, Zhou WP, Yang GS, Wang YZ, et al: Long noncoding
RNA high expression in hepatocellular carcinoma facilitates tumor
growth through enhancer of zeste homolog 2 in humans. Hepatology.
54:1679–1689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Enfield KS, Pikor LA, Martinez VD and Lam
WL: Mechanistic roles of noncoding RNAs in lung cancer biology and
their clinical implications. Genet Res Int.
2012:7374162012.PubMed/NCBI
|
|
24
|
Piao HL and Ma L: Non-coding RNAs as
regulators of mammary development and breast cancer. J Mammary
Gland Biol Neoplasia. 17:33–42. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ying L, Huang Y, Chen H, Wang Y, Xia L,
Chen Y, Liu Y and Qiu F: Downregulated MEG3 activates autophagy and
increases cell proliferation in bladder cancer. Mol Biosyst.
9:407–411. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dereure O: Role of non-coding RNA ANRIL in
the genesis of plexiform neurofibromas in neurofibromatosis type 1.
Ann Dermatol Venereol. 139:421–422. 2012.In French. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gutschner T, Hämmerle M, Eissmann M, Hsu
J, Kim Y, Hung G, Revenko A, Arun G, Stentrup M, Gross M, et al:
The noncoding RNA MALAT1 is a critical regulator of the metastasis
phenotype of lung cancer cells. Cancer Res. 73:1180–1189. 2013.
View Article : Google Scholar
|
|
28
|
Thai P, Statt S, Chen CH, Liang E,
Campbell C and Wu R: Characterization of a novel long noncoding
RNA, SCAL1, induced by cigarette smoke and elevated in lung cancer
cell lines. Am J Respir Cell Mol Biol. 49:204–211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang Y, Li H, Hou S, Hu B, Liu J and Wang
J: The noncoding RNA expression profile and the effect of lncRNA
AK126698 on cisplatin resistance in non-small-cell lung cancer
cell. PLoS One. 8:e653092013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu L, Smith TF and Goldberger G: LFG: A
candidate apoptosis regulatory gene family. Apoptosis.
14:1255–1265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Somia NV, Schmitt MJ, Vetter DE, Van
Antwerp D, Heinemann SF and Verma IM: LFG: An anti-apoptotic gene
that provides protection from Fas-mediated cell death. Proc Natl
Acad Sci USA. 96:12667–12672. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Beier CP, Wischhusen J, Gleichmann M,
Gerhardt E, Pekanovic A, Krueger A, Taylor V, Suter U, Krammer PH,
Endres M, et al: FasL (CD95L/APO-1L) resistance of neurons mediated
by phosphatidylinositol 3-kinase-Akt/protein kinase B-dependent
expression of lifeguard/neuronal membrane protein 35. J Neurosci.
25:6765–6774. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mercer TR and Mattick JS: Structure and
function of long noncoding RNAs in epigenetic regulation. Nat
Struct Mol Biol. 20:300–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Geisler S and Coller J: RNA in unexpected
places: Long non-coding RNA functions in diverse cellular contexts.
Nat Rev Mol Cell Biol. 14:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Khalil AM, Guttman M, Huarte M, Garber M,
Raj A, Rivea Morales D, Thomas K, Presser A, Bernstein BE, van
Oudenaarden A, et al: Many human large intergenic noncoding RNAs
associate with chromatin-modifying complexes and affect gene
expression. Proc Natl Acad Sci USA. 106:11667–11672. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mattick JS and Gagen MJ: The evolution of
controlled multi-tasked gene networks: The role of introns and
other noncoding RNAs in the development of complex organisms. Mol
Biol Evol. 18:1611–1630. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shamovsky I and Nudler E: Gene control by
large noncoding RNAs. Sci STKE. 2006:pe402006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kaikkonen MU, Lam MT and Glass CK:
Non-coding RNAs as regulators of gene expression and epigenetics.
Cardiovasc Res. 90:430–440. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cooper C, Guo J, Yan Y,
Chooniedass-Kothari S, Hube F, Hamedani MK, Murphy LC, Myal Y and
Leygue E: Increasing the relative expression of endogenous
non-coding Steroid Receptor RNA Activator (SRA) in human breast
cancer cells using modified oligonucleotides. Nucleic Acids Res.
37:4518–4531. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Okamura D, Maeda I, Taniguchi H, Tokitake
Y, Ikeda M, Ozato K, Mise N, Abe K, Noce T, Izpisua Belmonte JC, et
al: Cell cycle gene-specific control of transcription has a
critical role in proliferation of primordial germ cells. Genes Dev.
26:2477–2482. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Prensner JR, Iyer MK, Balbin OA,
Dhanasekaran SM, Cao Q, Brenner JC, Laxman B, Asangani IA, Grasso
CS, Kominsky HD, et al: Transcriptome sequencing across a prostate
cancer cohort identifies PCAT-1, an unannotated lincRNA implicated
in disease progression. Nat Biotechnol. 29:742–749. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fernández M, Segura MF, Solé C, Colino A,
Comella JX and Ceña V: Lifeguard/neuronal membrane protein 35
regulates Fas ligand-mediated apoptosis in neurons via microdomain
recruitment. J Neurochem. 103:190–203. 2007.PubMed/NCBI
|
|
43
|
Yoganathan P, Karunakaran S, Ho MM and
Clee SM: Nutritional regulation of genome-wide association obesity
genes in a tissue-dependent manner. Nutr Metab. 9:652012.
View Article : Google Scholar
|
|
44
|
Wu L, Zhao X, Shen Y, Zhang MX, Yan Y, Hou
D, Meng L, Liu J, Cheng H and Mi J: Promoter methylation of fas
apoptotic inhibitory molecule 2 gene is associated with obesity and
dyslipidaemia in Chinese children. Diab Vasc Dis Res. 12:217–220.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Boender AJ, van Rozen AJ and Adan RA:
Nutritional state affects the expression of the obesity-associated
genes Etv5, Faim2, Fto, and Negr1. Obesity. 20:2420–2425. 2012.
View Article : Google Scholar : PubMed/NCBI
|