Introduction
Esophageal squamous cell carcinoma (ESCC) and
esophageal adenocarcinoma represent two main histopathological
subtypes of esophageal cancer, which is the eighth most common
cancer and the sixth leading cause of cancer-related mortality
worldwide, affecting men more than women (1). ESCC is characterized by its poor
prognosis, with a 5-year survival rate less than 15% (2). Particularly, in Western countries, the
incidence and the mortality rates of ESCC have steadily increased
during the past few decades (3).
Despite advancement in radical esophagectomy and systemic
chemoradiotherapy, the clinical outcome of ESCC patients is still
very poor due to the high prevalence of cell proliferation and
metastasis (4). Moreover, most ESCC
patients are often diagnosed at an advanced stage due to the
absence of apparent symptoms and lack of early detection methods
(5). Therefore, it is of great
clinical significance to discover and identify tumor-specific
molecular biomarkers for the early detection and effective
treatment, and subsequently for the better understanding of the
underlying biological mechanisms of ESCC.
MicroRNAs (miRNAs), a conserved group of endogenous,
non-coding and single-stranded small RNAs 18–29 nucleotides in
length, can negatively regulate gene expression by binding to the
3′-UTR of target messenger RNAs (mRNAs) post-transcriptionally,
leading to translation repression or promotion of RNA degradation
(6). miRNAs play crucial roles in
the regulation of a variety of biological and pathological
processes, including developmental timing, cellular growth,
proliferation, differentiation, death, apoptosis and carcinogenesis
(7,8). Abnormal expression of miRNAs has been
observed in multiple types of human cancers, implying that miRNAs
may function as either oncogenes or tumor suppressors by the
sequence-specific regulation of the corresponding target gene
expression (9). miR-30d, a member
of the miR-30 family, is localized in human chromosome 8q24.22
(10). Accumulating evidence has
revealed that the aberrant expression of miR-30d contributes to
cancer development and progression. For example, miR-30d
downregulation was found to contribute to the development of
anaplastic thyroid carcinoma through the regulation of the polycomb
protein enhancer of zeste homolog 2 (EZH2) (11). miR-30d expression was found to be
increased in hepatocellular carcinoma tissues and its
overexpression promoted tumor invasion and metastasis by targeting
Galphai2 (12). miR-30d suppressed
prostate cancer cell proliferation partially by targeting Bmi-1
(13). miR-30d was found to inhibit
renal carcinoma cell proliferation via the regulation of cyclin E2
expression at the post-transcriptional level (14). However, the role of miR-30d in human
ESCC has not been documented. Therefore, the present study aimed to
investigate the expression pattern, clinical significance and
biological functions of miR-30d in esophageal carcinogenesis.
In the present study, quantitative real-time PCR was
performed to detect the expression levels of miR-30d in both ESCC
tissues and cell lines. Then, the associations between miR-30d
expression and various clinicopathological features of patients
with ESCC were statistically evaluated. In addition, the target
genes of miR-30d were predicted by bioinformatic miRNA target
prediction tool, and validated by western blot analysis and
luciferase reporter assay. Moreover, the functions of miR-30d in
migration and invasion were determined using two human ESCC cell
lines following transfection with miRNA mimics or co-transfected
with miRNA mimics and the expression vector of its target gene.
Materials and methods
Statement of ethics
The present study was approved by the Ethics
Committee of Huai'an First People's Hospital. Prior informed
consent was signed by all the patients enrolled in the present
study according to guidelines of Huai'an First People's Hospital.
All tissue specimens were handled and made anonymous based on
ethical and legal standards.
Ethical approval
All procedures performed in the present study
involving human participants were in accordance with the ethical
standards of the institutional and/or national research committee
and with the 1964 Helsinki Declaration and its later amendments or
comparable ethical standards.
Patients and tissue samples
A total of 60 pairs of matched primary ESCC and
adjacent non-cancerous esophageal mucosal tissue specimens were
obtained from 60 patients who underwent esophagectomy at the
Department of Gastroenterology, Huai'an First People's Hospital
between Janurary 2012 and December 2015. None of the patients
received radiotherapy or chemotherapy before surgery. The diagnosis
was confirmed by clinical examination and histopathological
analysis of the tissue specimens. The clinicopathological
characteristics of all 60 ESCC patients, including age, gender,
tumor location, lymph node metastasis, tumor-node-metastasis (TNM)
stage (based on the 7th edition of the AJCC/UICC TNM staging
system) and pathological grade are summarized in Table I. All of the samples were collected
and immediately snap frozen in liquid nitrogen and stored at −80°C
until further use.
 | Table I.Associations between miR-30d
expression and various clinicopathological characteristics of the
60 patients with ESCC. |
Table I.
Associations between miR-30d
expression and various clinicopathological characteristics of the
60 patients with ESCC.
| Clinical
variables | No. of patients
(%) | Low miR-30d
expression (n, %) | High miR-30d
expression (n, %) | P-value |
|---|
| Age (years) |
| ≤60 | 25 (41.67) | 13 (52.00) | 12 (48.00) | NS |
|
>60 | 35 (58.33) | 19 (54.29) | 16 (45.71) |
| Gender |
| Male | 40 (66.67) | 22 (55.00) | 18 (45.00) | NS |
|
Female | 20 (33.33) | 10 (50.00) | 10 (50.00) |
| Tumor location |
| Upper
1/3-middle1/3 | 41 (68.33) | 23 (56.10) | 18 (43.90) | NS |
| Lower
1/3 | 19 (31.67) | 9 (47.37) | 10 (52.63) |
| Lymph node
metastasis |
|
Negative | 24 (40.00) | 6 (25.00) | 18 (75.00) | 0.006 |
|
Positive | 36 (60.00) | 26 (72.22) | 10 (27.78) |
| TNM stage |
|
Early | 20 (33.33) | 2 (10.00) | 18 (90.00) | <0.001 |
|
Advanced | 40 (66.67) | 30 (75.00) | 10 (25.00) |
| Histological
differentiation |
| Well | 18 (33.33) | 6 (33.33) | 12 (66.67) | 0.02 |
|
Moderate/poor | 42 (66.67) | 26 (61.90) | 16 (38.10) |
Cell lines and culture
A human normal esophageal cell line (HEEC) and two
human ESCC cell lines (ECA109 and KYSE410) were purchased from the
Shanghai Institute of Cell Biology, Chinese Academy of Sciences
(Shanghai, China). All cell lines were cultured in Dulbecco's
modified Eagles medium (DMEM; Gibco, Carlsbad, CA, USA)
supplemented with 10% fetal bovine serum (FBS). Cells were cultured
at 37°C with 5% CO2 for further use.
Cell transfection
The transfection of human ESCC cells with
miR-30d/control mimic (miR-30d/NC_mimics) and the pcDNA3.1-EZH2
expression vector (en-EZH2) (both from GenePharma Biotech,
Shanghai, China) was carried out using Lipofectamine 2000
(Invitrogen, Carlsbad, CA, USA) according to the manufacturers
instructions. At 48 h following transfection, the ESCC cells were
harvested for western blot or real-time quantitative PCR
analyses.
Western blot analysis
Proteins in fresh clinical tissue specimens and
cells were extracted using cell lysis buffer (50 mM Tris-HCl, pH
8.0, 2 mM EDTA, 1 mM DTT, 10 mM NaCl, 5 mg/ml leupeptin, 1% NP-40,
2 mg/ml pepstatin, 2 mg/ml aprotinin, 0.1% SDS and 1 mM
phenylmethylsulfonyl fluoride). Equal amounts of protein (50 µg)
were separated by 10% SDS-PAGE, and then transferred onto
polyvinylidene difluoride membranes (Qiagen China Co., Ltd.,
Shanghai, China). Then, the membranes were incubated with the
primary antibodies: anti-EZH2 (dilution 1:200; Zymed Laboratories
Inc., South San Francisco, CA, USA) and anti-GAPDH (dilution 1:500;
Abcam Inc., Cambridge, MA, USA), after blocking with 8% milk in
phosphate-buffered saline (PBS; pH 7.5). After that, the membranes
were incubated with the appropriate horseradish
peroxidase-conjugated secondary antibodies (dilution 1:1,000; Abcam
Inc.) after incubation at 4°C overnight. Finally, the proteins were
visualized using enhanced chemiluminescence reagent (Santa Cruz
Biotechnology, Santa Cruz, CA, USA). The expression level of EZH2
protein was normalized to that of GAPDH protein. Each sample was
examined in triplicate.
RNA extraction and real-time
quantitative RT-PCR
Total RNA in fresh clinical tissue specimens and
cells was extracted using the RNeasy RNA Mini kit (Qiagen, Hilden,
Germany) according to the manufacturer's instructions. First-strand
cDNA was synthesized using PowerScript reverse transcriptase
(Clontech, San Jose, CA, USA) according to the manufacturer's
instructions. Following cDNA synthesis, real-time PCR was performed
using a Fast Start Master SYBR-Green kit on a LightCycler (both
from Roche Molecular Systems, Indianapolis, IN, USA) according to
the manufacturer's instructions. The sequence-specific primer pairs
used for quantitative PCR are listed as following: miR-30d forward,
5′-UGUAAACAUCCCCGACUGGAAG-3′ and reverse,
5′-TGTAAACATCCCCGACTGGAAGA-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′; EZH2 forward,
5′-TTACTTGTGGAGCCGCTGAC-3′ and reverse, 5′-TCAGATGGTGCCAGCAATAG-3′;
GAPDH forward, 5′-GCTGAGTATGTCGTGGAGTC-3′ and reverse,
5′-AGTTGGTGGTGCAGGATGC-3′. Relative expression levels of miRNA and
mRNA expression in fresh tissues and cells were determined using
the 2−ΔΔCt method. Each sample was examined in
triplicate.
Cell invasion and migration
assays
Cell migration and invasion abilities of human ESCC
cell lines following transfection with miR-30d/NC-mimics or the
co-transfection of miR-30d/en-EZH2 were evaluated using a Millicell
Transwell chamber (Millipore, Billerica, MA, USA), with or without
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). For the
invasion and migration assays, 48 h following the transfection,
Transwell chambers were placed into 24-well plates which were
respectively precoated with or without a 5 ml mixture of BD
Matrigel and DMEM (1:1, v/v). Following incubation at 37°C in a
humidified incubator with 5% CO2 for 40 min, for cell
migration and invasion assays, 1×105 tumor cells in 0.1
µl of media without FBS were plated in the upper chamber. In the
lower chamber, 0.6 µl of medium with 10% FBS was added. Forty-eight
hours after incubation, cells on the upper surface of the Millicell
chambers, non-invasive or non-migrated cells, were scraped off
using a cotton swab. Tumor cells on the bottom surface of the
membrane were fixed with 4% paraformaldehyde at room temperature
for 30 min and stained with 0.1% crystal violet for 15 min. The
numbers of migrated or invasive cells were counted in five randomly
selected fields under an inverted microscope (Olympus CKX41;
Olympus, Tokyo, Japan). Each sample was examined in triplicate.
miRNA target prediction
Validated targets for miR-30d-5p were collected from
miRTarBase (release 6.1; http://mirtarbase.mbc.nctu.edu.tw/), an experimentally
validated miRNA-target interaction (MTI) database, which has
accumulated more than 300 and 60,000 MTIs validated experimentally
by reporter assay, western blotting, qPCR, microarray and
next-generation sequencing experiments (15). While containing the largest numbers
of validated MTIs, the miRTarBase provides the most updated
collection by comparing with other similar, previously developed
databases. In the present study, we only collected the MTIs which
were validated experimentally by reporter assay, western blotting
and qPCR.
Luciferase reporter assay
To determine the binding efficiency of miR-30d on
the 3′UTR of EZH2 mRNA, two human ESCC cell lines were
co-transfected with miR-30d/NC-mimics and pGL3-EZH2-3′-UTR-WT/MUT.
To construct the EZH2 3′-UTR luciferase reporter
(pGL3-EZH2-3′-UTR), the 263-bp 3′-UTR of EZH2 mRNA was amplifed by
PCR (forward, 5′-GTATGTCGGCATCGAAAGAG-3′ and reverse,
5′-CCTGAAAGCAGTTATTGACA-3′). The amplified fragment was cloned into
pGL3-control firefly luciferase reporter vector (Promega, Madison,
WI, USA). Deletions of the seed region in the EZH2 3′-UTR
constructs were introduced by site-directed mutagenesis (Eurofins
MWG Operon, Ebersberg, Germany). At 48 h after transfection, the
cells were collected and detected with the Dual-Luciferase reporter
assay system (Promega, San Luis Obispo, CA, USA). Luciferase
activity was measured using a Lumat LB 9507 apparatus (Berthold
Technologies, Bad Wildbad, Germany). Each sample was examined in
triplicate. The results are shown as relative luciferase activity,
which was normalized to β-galactosidase activity.
Statistical analysis
SPSS software (version 11.0; SPSS, Inc., Chicago,
IL, USA) was used to perform all statistical analyses. Data are
shown as mean ± standard deviation (SD). The differences between
groups were analyzed using the Student's t-test when there were
only two groups, or assessed by one-way analysis of variance
(ANOVA) when more than two groups were analyzed. The correlation
between miR-30d and EZH2 mRNA expression in ESCC tissues was
determined by Spearman correlation analysis. The associations
between miR-30d expression and various clinicopathological
characteristics of the patients with ESCC were assessed using the
χ2 test for categorical variables. A P-value <0.05
was considered to indicate a statistically significant result.
Results
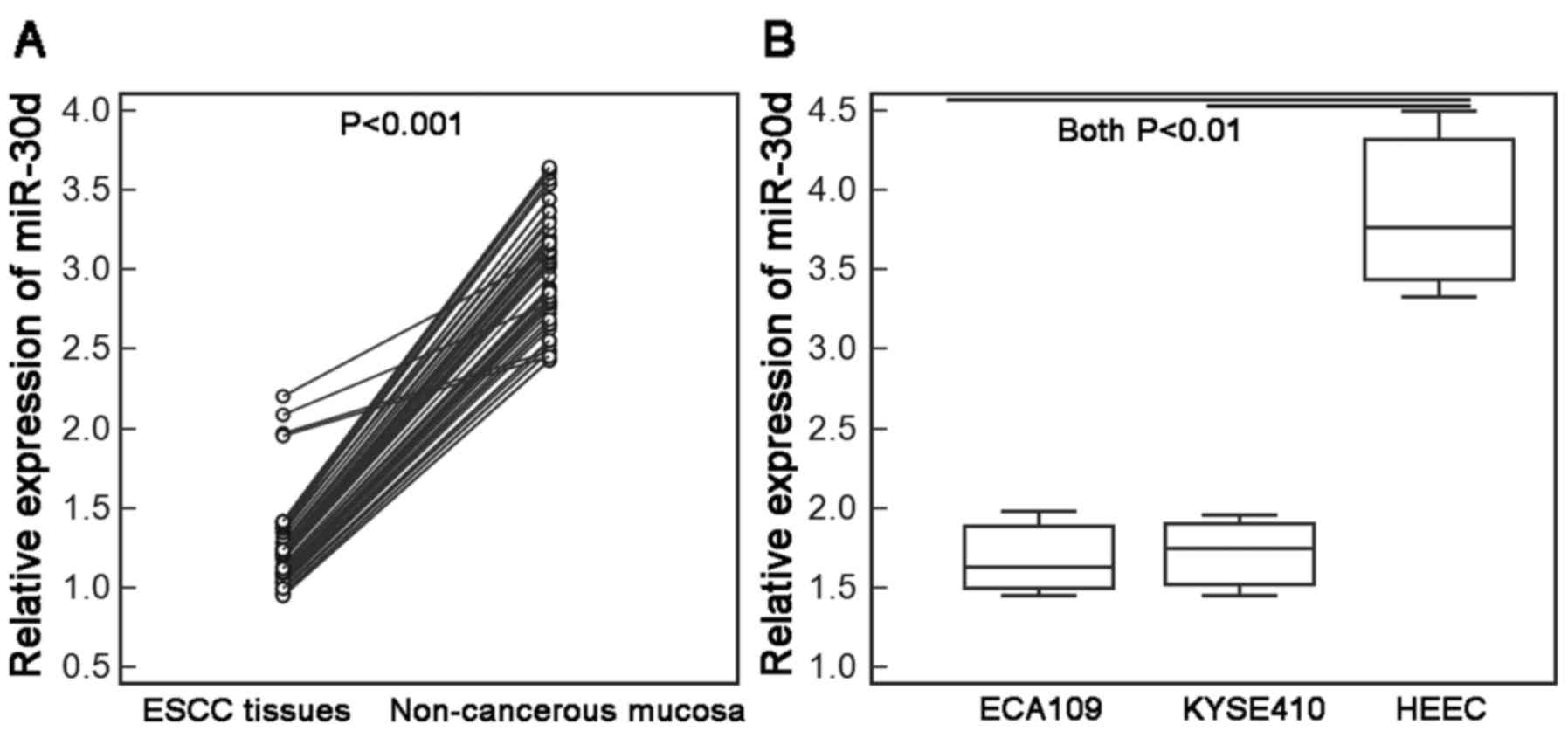
miR-30d expression is downregulated in
ESCC tissues and cells
Compared with the adjacent non-cancerous esophageal
mucosal tissues and human normal esophageal cell line (HEEC),
respectively, the expression levels of miR-30d in ESCC tissues, and
two human ESCC cell lines ECA109 and KYSE410 were all markedly
decreased [ESCC vs. non-cancerous tissues: 1.23±0.26 vs. 3.02±0.38;
P<0.001 (Fig. 1A); ECA109 vs.
HEEC cells: 1.69±0.27 vs. 3.87±0.59; P<0.01; KYSE410 vs. HEEC
cells: 1.72±0.26 vs. 3.87±0.59; P<0.01 (Fig. 1B)].
miR-30d downregulation is associated
with aggressive tumor progression in the ESCC cases
The median value of miR-30d (1.21) expression levels
in 60 ESCC tissues was used as a cut-off point to divide all 60
patients with ESCC into low miR-30d expression (n=32) and high
miR-30d expression (n=28) groups. Table
I summarizes the associations between miR-30d expression and
various clinical characteristics of the patients with ESCC. Reduced
expression of miR-30d occurred more frequently in ESCC patients
with positive lymph node metastasis (P=0.006; Table I), moderate-poor differentiation
(P=0.02; Table I) and advanced TNM
stage (P<0.001; Table I) than
those with negative lymph node metastasis, well differentiation and
early TNM stage. There were no significant associations between
miR-30d expression and patient age, gender or tumor location (all
P>0.05; Table I).
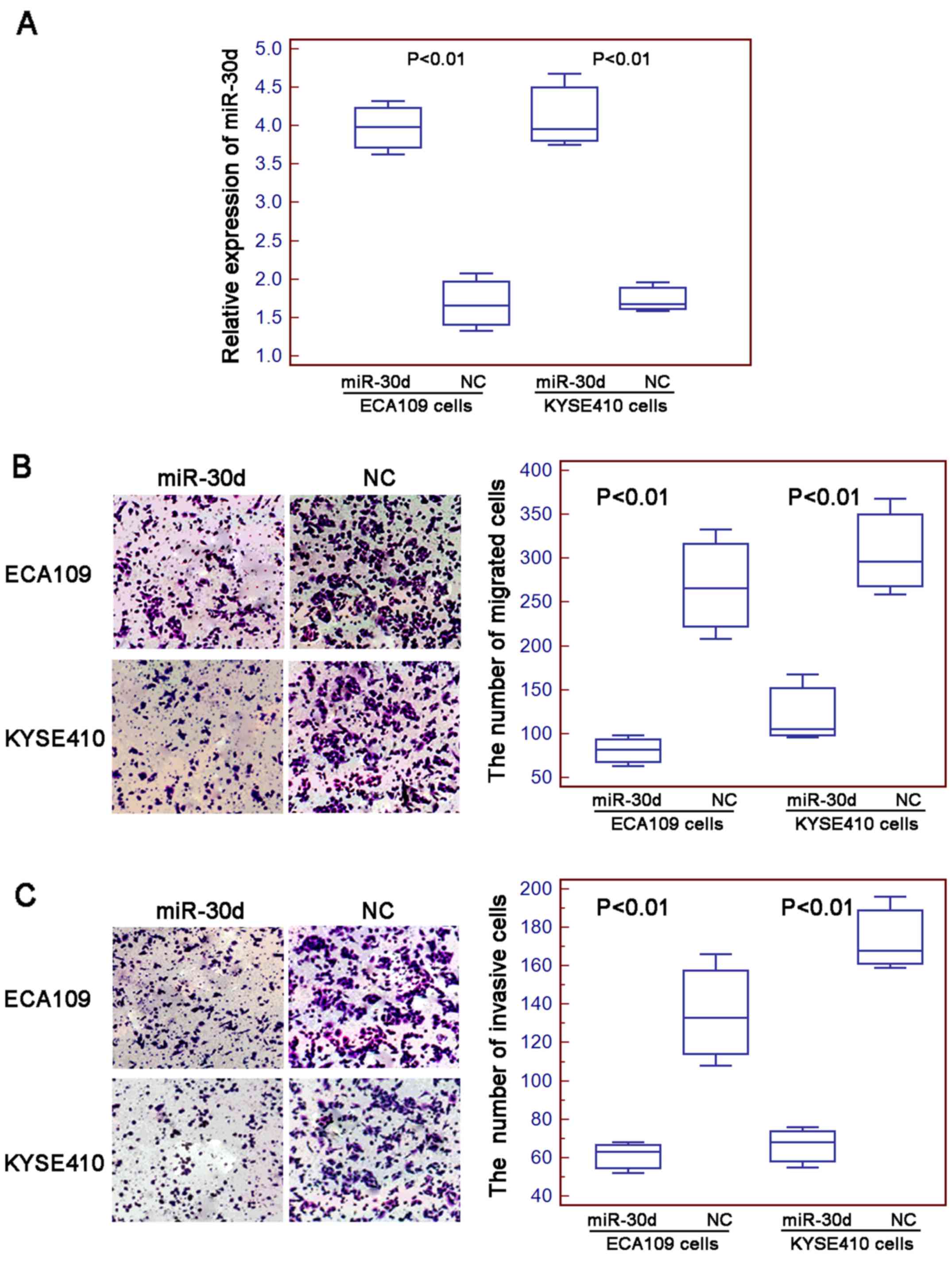
Enforced expression of miR-30d
inhibits cell migration and invasion of human ESCC cells
To determine the role of miR-30d in ESCC cell
motility, the migration and invasion abilities of two human ESCC
cell lines ECA109 and KYSE410 transfected with the miR-30d mimics
or negative control mimics were assessed. Quantitative real-time
PCR showed that the expression levels of miR-30d in the ECA109 and
KYSE410 cells transfected with the miR-30d mimics were markedly
higher than levels in the cells tranfected with the negative
control mimics (both P<0.01; Fig.
2A). As shown in Fig. 2B, the
numbers of cells that migrated through the micropore membrane were
significantly decreased in the cells transfected with the miR-30d
mimics compared to the numbers of cells transfected with the
negative control mimics (for both ECA109 and KYSE410 cell lines;
both P<0.01). Similarly, the results of the Matrigel-coated
Transwell assay revealed that the numbers of miR-30d-overexpressing
ESCC cells that invaded through the Matrigel were markedly lower
than those in the control groups (for both ECA109 and KYSE410 cell
lines; both P<0.01; Fig.
2C).
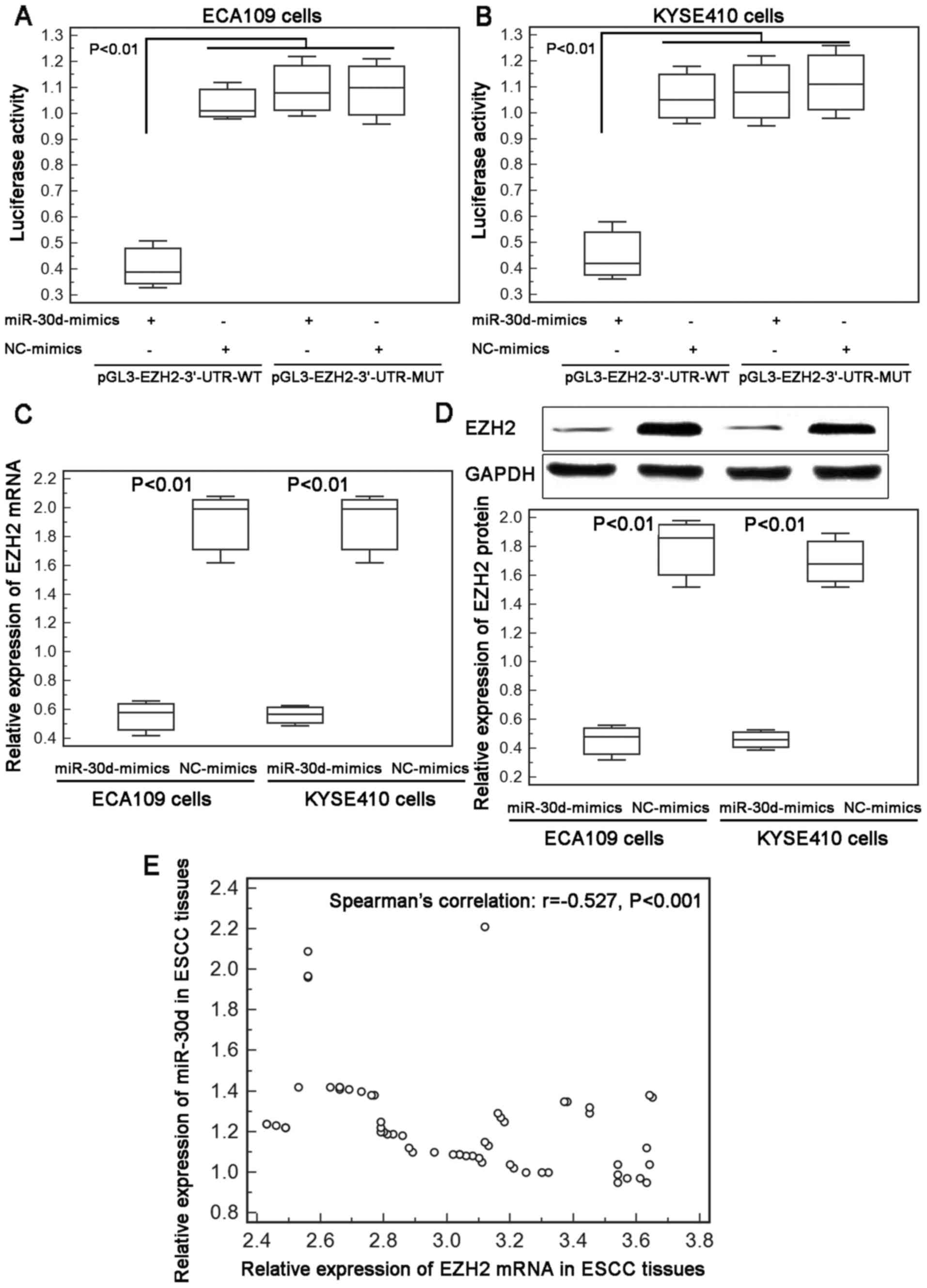
miR-30d reduces EZH2 expression by
targeting the 3′-UTR of EZH2 mRNA
The above findings indicated that miR-30d
downregulation was significantly associated with aggressive
progression of patients with ESCC and enforced expression of this
miRNA efficiently suppressed the motility of the ESCC cells. Next,
we aimed to identify the downstream target genes which are
negatively regulated by miR-30d in ESCC cells. As a result, a total
of 17 candidate target genes (including ATG12, ATG2B, ATG5, BCL9,
BECN1, BNIP3L, CASP3, EZH2, GNAI2, GPR78, KPNB1, NOTCH1, RUNX2,
SMAD1, SNAI1, SOCS1 and TP53) for which the interaction between
miR-30d was validated experimentally by reporter assay, western
blotting and qPCR, were collected from miRTarBase for miR-30d.
Among these genes, a previous study of Esposito et al
(10) found that miR-30d directly
targeted the 3′-UTR of EZH2 in anaplastic thyroid carcinoma.
Growing evidence also showed that EZH2 overexpression was
associated with tumor aggressiveness and poor prognosis in patients
with ESCC (16–18). Based on this finding, we chose EZH2
as a potential target gene of miR-30d in ESCC cells.
The luciferase activities in ECA109 and KYSE410
cells co-transfected with miR-30d/NC-mimics and
pGL3-EZH2-3′-UTR-WT/MUT were measured by luciferase report assay.
As shown in Fig. 3A and B, ECA109
and KYSE410 cells co-transfected with miR-30d-mimics and
pGL3-EZH2-3-UTR-WT both displayed a significant decrease in
reporter activity in comparison with the activity of cells
co-transfected with the NC-mimics and the pGL3-EZH2-3′-UTR-WT
plasmid. However, there were no differences with statistical
significance in the reporter activity between ECA109 and KYSE410
cells co-transfected with the miR-30d-mimics and the
pGL3-EZH2-3′-UTR-MUT plasmid and cells co-transfected with the
NC-mimics and the pGL3-EZH2-3′-UTR-MUT plasmid (Fig. 3A and B).
In addition, enforced expression of miR-30d markedly
inhibited the endogenous expression of EZH2 mRNA and protein in the
ECA109 and KYSE410 cells (Fig. 3C and
D). Moreover, our Spearman's correlation analysis showed a
negative correlation between miR-30d and EZH2 mRNA expression in
ESCC tissues (Spearman's correlation: r=−0.527, P<0.001;
Fig. 3E).
These findings provide evidence that EZH2 may be a
direct target of miR-30d in human ESCC cells.
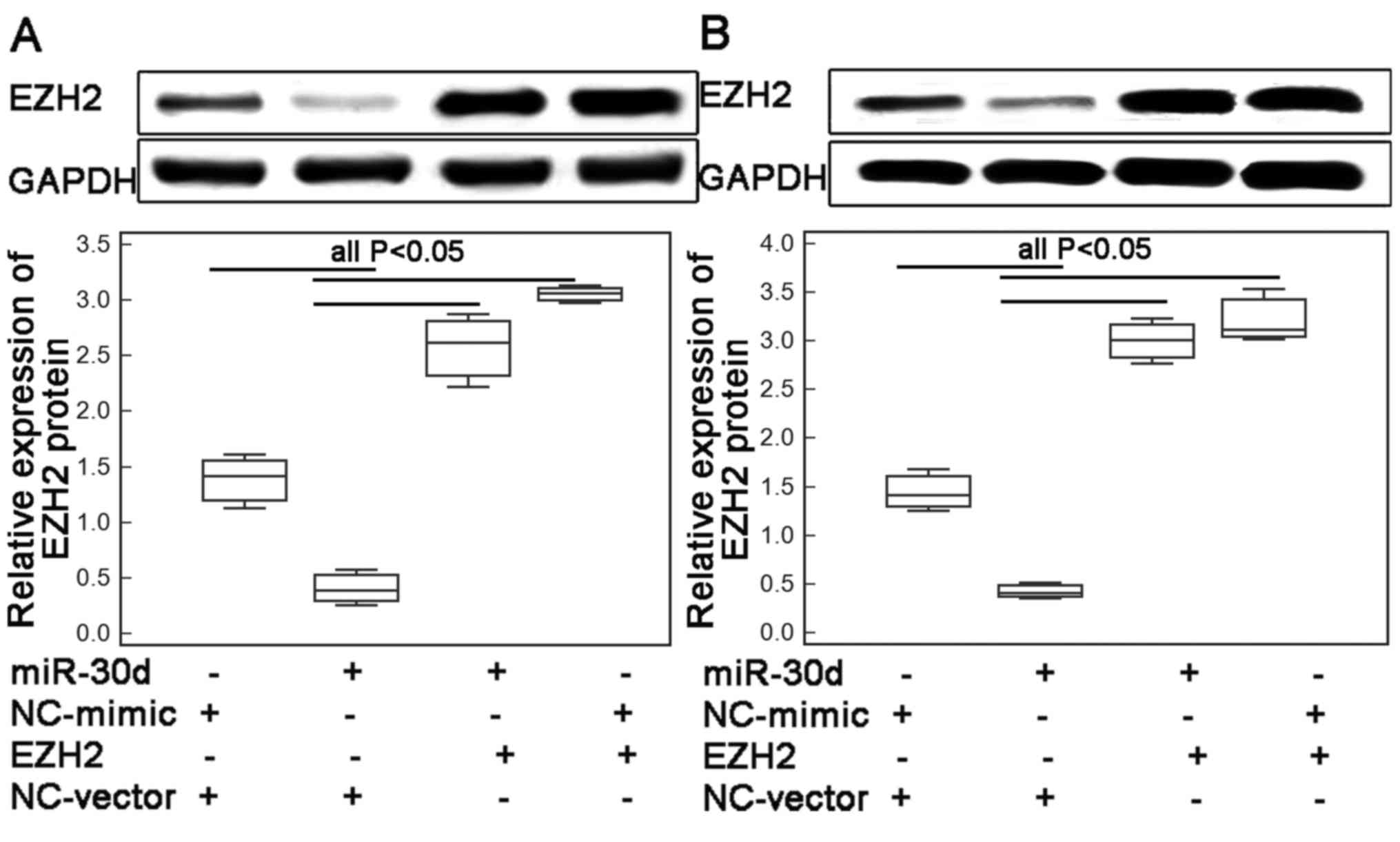
Suppression of migration and invasion
of ESCC cells by miR-30d is reversed by the overexpression of
EZH2
To determine whether miR-30d suppressed cell
migration and invasion of human ESCC cells via regulating its
target gene EZH2, the motility capacities of the ECA109 and KYSE410
cells co-transfected with the miR-30d-mimics and the pcDNA3.1-EZH2
expression vector were evaluated. As a result, western blot
analysis showed that the endogenous expression levels of EZH2
protein in the ECA109 and KYSE410 cells transfected with the
miR-30d mimic in the presence of the EZH2 expression vector were
both significantly higher than the levels in the cells
co-transfected with the miR-30d mimic and vector control (both
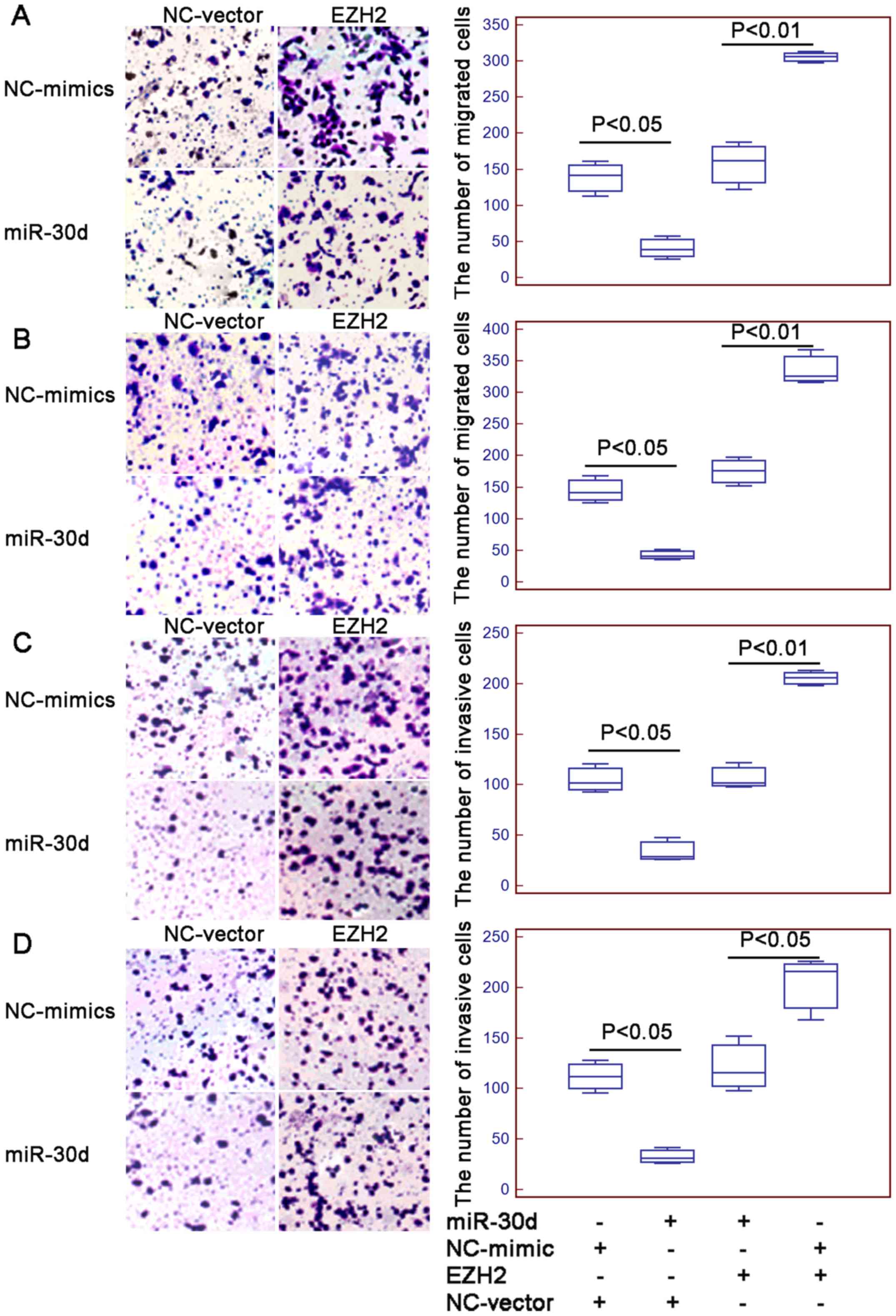
P<0.01; Fig. 4A and B,
respectively). Functionally, a markedly higher number of ESCC cells
co-transfected with the miR-30d mimics and pcDNA-EZH2 expression
vector migrated through the micropore or invaded through the
Matrigel (all P<0.05; Fig. 5),
compared with the numbers of cells co-transfected with the miR-30d
mimics and vector control.
Discussion
Accumulating evidence has revealed that miRNA-based
molecular alterations may be implicated in the initiation and
progression of human ESCC by activating or suppressing multiple
malignant processes (19,20). However, the underlying mechanisms of
esophageal carcinogenesis have not been fully elucidated. In the
present study, we demonstrated that the expression level of miR-30d
in ESCC tissues was markedly lower than the level in adjacent
non-cancerous esophageal mucosal tissues, which was similar with
the observations observed using two human ESCC cell lines and one
human normal esophageal cell line. Then, we also found that the
reduced expression of miR-30d was significantly associated with the
status of lymph node metastasis and the histological
differentiation of ESCC tissues, as well as with the TNM stage.
Morover, decreased expression of miR-30d was negatively correlated
with increased expression of EZH2 in ESCC tissues. Moreover, our
data showed that increased miR-30d expression efficiently inhibited
cell migration and invasion by reducing the expression of EZH2.
These findings imply a crucial role for the aberrant expression of
miR-30d in the carcinogenesis and metastasis of ESCC cells.
The differential expression patterns of miR-30d in
normal and cancerous cells suggest the roles of this miRNA in
sustaining normal physiological conditions as well as its
implications in cancers. It functions as either an oncogene or a
tumor suppressor by regulating the expression of its target gene
(11–14). Our data in the present study
identified a tumor-suppressive role of miR-30d in human ESCC and
also identified EZH2 as a direct target gene of this miRNA. As a
member of polycomb repressive complex (PRC)2, EZH2 functions as a
histone methyl transferase (21).
Under a normal condition, EZH2 was found to control cell growth,
cell proliferation and cell cycle (22). During carcinogenesis, EZH2
overexpression was originally observed in hematologic malignancies,
and then its amplification was confirmed in various human types of
cancers, such as oral cancer, ESCC, breast, lung and gastric
cancer, hepatocellular carcinoma, colon, prostate, bladder and
endometral cancer (16–18,23–28). Aberrant expression of EZH2 was
reported to be associated with tumor aggressive progression and
poor prognosis in patients with many types of human cancers.
Particularly, in ESCC, He et al (16) indicated that overexpression of EZH2
was found in >50% of ESCC patients, and was closely correlated
with increased cell proliferation, high histopathological grade,
regional and distant lymph node metastasis and lack of clinical
complete response to chemoradiotherapy, as well as adverse patient
outcome of ESCC patients treated with definitive chemotherapy; Ha
et al (17) also identified
the co-expression of EZH2 and another member of PRC1-Bmi1 as an
independent poor prognostic factor in ESCC. In the present study,
EZH2 was confirmed to be a direct target of miR-30d based on a
luciferase reporter system, the western blot assay and the spearman
correlation analysis of the expression levels of miR-30d and EZH2
mRNA in ESCC tissues. Rescue experiments also demonstrated that the
inhibitory effects of miR-30d in cell migration and invasion of
ESCC cells were reversed by the enforced expression of EZH2, which
indicate a role of the miR-30d-EZH2 axis in esophageal malignant
progression.
In conclusion, these findings provide convincing
evidence that the decreased expression of miR-30d may be implicated
in esophageal carcinogenesis and progression. We also confirmed
miR-30d as a tumor suppressor which may inhibit cancer cell
motility by targeting EZH2, implying a potential therapeutic target
for ESCC.
References
|
1
|
Jemal A, Bray F, Centerm M, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tan C, Qian X, Guan Z, Yang B, Ge Y, Wang
F and Cai J: Potential biomarkers for esophageal cancer.
Springerplus. 5:4672016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ohashi S, Miyamoto S, Kikuchi O, Goto T,
Amanuma Y and Muto M: Recent advances from basic and clinical
studies of esophageal squamous cell carcinoma. Gastroenterology.
149:1700–1715. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kang X, Chen K, Li Y, Li J, D'Amico TA and
Chen X: Personalized targeted therapy for esophageal squamous cell
carcinoma. World J Gastroenterol. 21:7648–7658. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yan X, Xu H and Yan Z: Functional
perspective and implications of gene expression by noncoding RNAs.
Cancer Transl Med. 1:137–152. 2015. View Article : Google Scholar
|
|
7
|
Svoronos AA, Engelman DM and Slack FJ:
OncomiR or tumor suppressor? The duplicity of microRNAs in cancer.
Cancer Res. 76:3666–3670. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Otero-Albiol D and Felipe-Abrio B:
MicroRNA regulating metabolic reprogramming in tumor cells: New
tumor markers. Cancer Transl Med. 2:175–181. 2016. View Article : Google Scholar
|
|
9
|
Rupaimoole R, Calin GA, Lopez-Berestein G
and Sood AK: miRNA deregulation in cancer cells and the tumor
microenvironment. Cancer Discov. 6:235–246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang XJ, Si RH, Liang YH, Ma BQ, Jiang ZB,
Wang B and Gao P: Mir-30d increases intracellular survival of
Helicobacter pylori through inhibition of autophagy pathway. World
J Gastroenterol. 22:3978–3991. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Esposito F, Tornincasa M, Pallante P,
Federico A, Borbone E, Pierantoni GM and Fusco A: Down-regulation
of the miR-25 and miR-30d contributes to the development of
anaplastic thyroid carcinoma targeting the polycomb protein EZH2. J
Clin Endocrinol Metab. 97:E710–E718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yao J, Liang L, Huang S, Ding J, Tan N,
Zhao Y, Yan M, Ge C, Zhang Z, Chen T, et al: MicroRNA-30d promotes
tumor invasion and metastasis by targeting Galphai2 in
hepatocellular carcinoma. Hepatology. 51:846–856. 2010.PubMed/NCBI
|
|
13
|
Kobayashi N, Uemura H, Nagahama K, Okudela
K, Furuya M, Ino Y, Ito Y, Hirano H, Inayama Y, Aoki I, et al:
Identification of miR-30d as a novel prognostic marker of prostate
cancer. Oncotarget. 3:1455–1471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu H, Lin X, Wang F, Zhang B, Wang W, Shi
H, Zou B and Zhao J: Proliferation inhibition and the underlying
molecular mechanisms of microRNA-30d in renal carcinoma cells.
Oncol Lett. 7:799–804. 2014.PubMed/NCBI
|
|
15
|
Chou CH, Chang NW, Shrestha S, Hsu SD, Lin
YL, Lee WH, Yang CD, Hong HC, Wei TY, Tu SJ, et al: miRTarBase
2016: Updates to the experimentally validated miRNA-target
interactions database. Nucleic Acids Res. 44:D239–D247. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He LR, Liu MZ, Li BK, Jia WH, Zhang Y,
Liao YJ, Chen YC, Zhang LJ, Guan XY, Zeng YX, et al: High
expression of EZH2 is associated with tumor aggressiveness and poor
prognosis in patients with esophageal squamous cell carcinoma
treated with definitive chemoradiotherapy. Int J Cancer.
127:138–147. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ha SY and Kim SH: Co-expression of Bmi1
and EZH2 as an independent poor prognostic factor in esophageal
squamous cell carcinoma. Pathol Res Pract. 208:462–469. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang SD, Yuan Y, Zhuang CW, Li BL, Gong
DJ, Wang SG, Zeng ZY and Cheng HZ: MicroRNA-98 and microRNA-214
post-transcriptionally regulate enhancer of zeste homolog 2 and
inhibit migration and invasion in human esophageal squamous cell
carcinoma. Mol Cancer. 11:512012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sakai NS, Samia-Aly E, Barbera M and
Fitzgerald RC: A review of the current understanding and clinical
utility of miRNAs in esophageal cancer. Semin Cancer Biol.
23:512–521. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mayne GC, Hussey DJ and Watson DI:
MicroRNAs and esophageal cancer - implications for pathogenesis and
therapy. Curr Pharm Des. 19:1211–1226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Trošelj K Gall, Novak Kujundzic R and
Ugarkovic D: Polycomb repressive complex's evolutionary conserved
function: The role of EZH2 status and cellular background. Clin
Epigenetics. 8:552016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Italiano A: Role of the EZH2 histone
methyltransferase as a therapeutic target in cancer. Pharmacol
Ther. 165:26–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Herviou L, Cavalli G, Cartron G, Klein B
and Moreaux J: EZH2 in normal hematopoiesis and hematological
malignancies. Oncotarget. 7:2284–2296. 2016.PubMed/NCBI
|
|
24
|
Kim KH and Roberts CW: Targeting EZH2 in
cancer. Nat Med. 22:128–134. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Martínez-Fernández M, Rubio C, Segovia C,
López-Calderón FF, Dueñas M and Paramio JM: EZH2 in bladder cancer,
a promising therapeutic target. Int J Mol Sci. 16:27107–27132.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin Y, Qiu S and Peng Y: Functional roles
of enhancer of zeste homolog 2 in gliomas. Gene. 576:189–194. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu B, Konze KD, Jin J and Wang GG:
Targeting EZH2 and PRC2 dependence as novel anticancer therapy. Exp
Hematol. 43:698–712. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Han Li C and Chen Y: Targeting EZH2 for
cancer therapy: Progress and perspective. Curr Protein Pept Sci.
16:559–570. 2015. View Article : Google Scholar : PubMed/NCBI
|