Introduction
With an estimated 1.7 million cases of breast cancer
diagnosed worldwide in 2012, this type of cancer remains the most
common in women. Among the patients with breast cancer,
triple-negative breast cancer (TNBC) accounts for 10–20% of the
invasive breast cancer which is defined as estrogen receptor
(ER)-negative, progesterone receptor (PR)-negative and human
epidermal growth factor receptor 2 (HER2)-negative (1). Due to the lack of markers, it carries
poor prognosis and presents an emerging need to understand the
biology of this subtype of breast cancer and develop alternative
therapeutic options.
Retinoids are structurally related to the hormone of
vitamin A with all-trans-retinoic acid (ATRA) being the
active metabolite. Despite the toxicity associated with retinoids,
it is in general considered to be well tolerated pharmacological
agent. Based on the notion that retinoic acid (RA) promotes cell
differentiation, regulates proliferation and apoptosis, it has been
combined with anthracycline-based chemotherapy to successfully
treat acute promyelocytic leukemia with a success rate of 80%
(2). Depending on its interacting
partner, RA has distinct biological functions. The ligand binding
domain of the retinoic acid receptor (RAR) which includes RARα,
RARβ and RARγ and retinoic X receptors RXRα, RXRβ and RXRγ can
interact with RA (3,4) and activate genes that are involved in
anti-proliferation, apoptosis, differentiation and cell cycle
arrest (5–9). However, binding of RA to the
alternative nuclear receptor, peroxisome proliferator-activated
receptor β/δ (PPARβ/δ) transduces signals which facilitate cell
growth, promote cell survival and protect cells against apoptosis
(10–13). Delivery of RA to RARs is facilitated
by cellular RA-binding protein II (CRABPII), which sequesters RA,
translocates to the nucleus, channels RA to RAR and enhances the
transcriptional activity of RAR target genes (6,8,14). On
the other hand, fatty acid-binding protein 5 (FABP5) can transport
RA to its cognate receptor, PPARβ/δ which targets genes that are
involved in proliferation and cell growth (10–13).
Due to the dual and opposing function of RA, the growth inhibitory
effects of RA are determined by the expression of the regulatory
factors, CRABPII, RARs, FABP5 and PPARβ/δ. The differential effects
of retinoids on anti-proliferation, cell cycle arrest and apoptosis
is dependent on the concentration of RA and the time frame of RA
treatment (15,16). Retinoids repress genes involved in
cell division and cell proliferation, which is followed by
differentiation, without affecting cell viability. While decrease
in proliferation and cell cycle arrest of cancer cells occurs at an
earlier time frame of within 2 days, the apoptotic effects of
retinoids begin on day 4 of treatment, with increased apoptosis
after day 5 or 6 of treatment (15,16).
Recent study has demonstrated the importance of
CRABPII in mammary carcinoma tumor growth suppression through
RA-dependent and RA-independent mechanism (17). The clinical significance of CRABPII
has been highlighted in several types of cancer, including
non-small lung cancer (18),
non-myeloma skin cancer (19) and
pancreatic cancer cells (20),
pinpointing that restoring the CRABPII signaling pathway may serve
as a therapeutic intervention to ameliorate the efficacy of RA or
sensitize cancer cells to this hormone. While CRABPII acts to
deliver RA to its RAR receptors, each of the different isoforms of
RAR exhibits specific function with each receptor regulating a
subset of distinct genes (21–26).
RARβ has been implicated in inflammation and tumor suppression
(21–23), while the loss of RARγ has been
demonstrated in the progression of malignant squamous cell
carcinoma (24,25). Selective activation of RARα by
retinoids induces autophagy in ER-positive breast cancer cells
(26). Though there may be
alternative RA-resistant mechanisms in cancer, the loss of
sensitivity to RA in cancers has been related to the lack of
CRABPII and RAR expression with increased expression of FABP5 and
PPARβ/δ (11,19,27,28).
Phytochemicals have been extensively studied for the
treatment of diseases such as cancer. Curcumin, an active
ingredient in the dietary spice turmeric (Curcuma Longa),
possesses antioxidant, anti-inflammatory and anticancer properties
(29,30). The apoptotic effects of curcumin
have been observed in several cancers including breast (31), pancreatic (32), prostate (33) and lung (34), while having no cytotoxic effects on
healthy cells (35). Despite its
low solubility and bioavailability, the combination of curcumin
with conventional chemotherapeutic agents has been demonstrated to
be effective in cancer regression (36,37).
Synthesis of nanocarriers has not only increased the solubility in
aqueous solution but has also improved the bioavailability of
curcumin towards cancer cells (38–40).
Several studies have demonstrated the ability of curcumin to
increase the sensitivity of cancer cells to chemotherapeutic drugs
(41,42). Through multiple mechanisms, curcumin
and its analogs sensitize cancer cells to chemotherapeutic agents,
thus overcoming drug resistance and improving susceptibility to
growth suppression by conventional drug treatments (43–47).
Previously, we reported that 30 µM curcumin
sensitizes TNBC cells to RA-mediated growth suppression by altering
the expression level of FABP5/PPARβ/δ pathway and targeting PPARβ/δ
target genes (45). However, we did
not explore the role of curcumin on the second arm of the retinoid
pathway, namely CRABPII/RAR, and in this study we evaluated the
hypothesis that curcumin mediated upregulation of the CRABPII/RAR
pathway in TNBC cells promotes sensitivity to RA-mediated
apoptosis. The overall aim of this study is to investigate the
effect of curcumin on the CRABPII/RAR pathway and assess the
contribution of this pathway in sensitizing TNBC to RA-mediated
growth suppression by triggering cell death. In order to achieve
this aim, we examine whether regulation of CRABPII/RAR by curcumin
is dose-dependent and the functional consequence of this pathway in
increasing sensitivity to RA. Furthermore, we explored the impact
of silencing CRABPII on apoptosis facilitated by curcumin and/or
RA. In the present study, we demonstrate that while 30 µM curcumin
induces CRABPII, it does not affect RARs in TNBC cells. However,
lower doses of curcumin (5 and 10 µM) upregulate CRABPII, RARβ and
RARγ expression in these cells. We also provide evidence that
induction of the CRABPII/RAR pathway by curcumin sensitizes
RA-resistant TNBC cells to RA-mediated apoptosis, and knockdown of
CRABPII in TNBC cell lines reverses the sensitization of the
apoptotic effects of RA by curcumin.
Materials and methods
Reagents
Antibodies for RARα, RARβ, RARγ and glyceraldehyde
3-phosphate dehydrogenase (GAPDH) were obtained from Abcam
(Cambridge, MA, USA). Anti-CRABPII was purchased from R&D
Systems (Minneapolis, MN, USA). Caspase-9 and poly(ADP-ribose)
polymerase (PARP) antibodies were obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Anti-mouse and anti-rabbit
immunoglobulin horseradish peroxidase-conjugated antibodies were
from Bio-Rad Laboratories, Inc. (Hercules, CA, USA) and anti-goat
immunoglobulin was from Santa Cruz Biotechnology, Inc. (Santa Cruz,
CA, USA). Curcumin (C-1386) and ATRA (R-2625) were purchased from
Sigma-Aldrich (St. Louis, MO, USA).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
reagent was purchased from Sigma-Aldrich. CRABPII and control siRNA
were obtained from Santa Cruz Biotechnology, Inc.
Cell lines
MDA-MB-231 and MDA-MB-468 cells were maintained in
Dulbeccos modified Eagles medium (DMEM) supplemented with 10% fetal
bovine serum (FBS) with antibiotics. MDA-MB-231 cells were a kind
gift from Dr Ming Tan (Mitchell Cancer Institute, Mobile, AL, USA).
MDA-MB-468 was purchased from American Type Culture Collection
(ATCC; Manassas, VA, USA).
Western blot analyses
Cells were cultured in 100-mm plates and treated
with curcumin and/or ATRA in media containing 10% charcoal treated
FBS for the indicated time. Cells were lysed in a buffer containing
150 mM NaCl, 10 mM Tris, pH 7.2, 0.1% SDS, 1% Triton X-100, 1%
deoxycholate, 5 mM EDTA and 1 mM PMSF for 1 h. The concentration of
the whole cell protein was determined using the Bradford assay.
Cell lysate was resolved by SDS-PAGE and transferred onto a
nitrocellulose membrane. The membrane was blocked in 10% bovine
serum albumin (BSA) in Tris-buffered saline containing 0.05%
Tween-20 for 1 h at room temperature. The membrane was then probed
using the appropriate antibody, CRABPII, RARα, RARβ, RARγ, PARP or
caspase-9 at a dilution of 1:1,000 overnight at 4°C. After a 30 min
wash, the appropriate secondary antibody was added for 1 h at room
temperature prior to exposure. Anti-GAPDH (1:2,000) was used as a
loading control and was incubated with the membrane for 1 h at 4°C.
The antigen-antibody complex was visualized with SuperSignal West
Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Inc.,
Hanover Park, IL, USA).
Quantitative real-time polymerase
chain reaction (qRT-PCR)
Cells were treated with curcumin for 4 h, and RNA
was extracted using TRIzol (Life Technologies, Grand Island, NY,
USA). As outlined in the protocol for the high-capacity RNA to cDNA
kit from Applied Biosystems (Foster City, CA, USA), 2 µg total RNA
was reverse transcribed into cDNA. To determine expression of
CRABPII, RARα, RARβ and RARγ, qRT-PCR was carried out by using
commercially available TaqMan chemistry and assay on demand probes
(Applied Biosystems). GAPDH was used for normalization. Detection
and data analysis were carried out on the ABI StepOnePlus Real-Time
PCR system. Relative quantity of gene expression was performed
using 2−ΔΔCt method (48).
Cell viability assay
MDA-MB-231 and MDA-MB-468 cells (5,000 cells/well)
were cultured in media containing 10% charcoal treated FBS and
plated in a 96-well plate overnight. Cells were then treated with
curcumin in the presence or absence of ATRA for 72 h. Controls used
were DMSO and/or ethanol depending on the drug treatment. After 72
h, 5 µg/ml of MTT reagent was added directly to the cells for 3 h
and allowed to incubate at 37°C. The media was removed from the
plate and the intact cells were resuspended in 150 µl of 0.04 M HCl
in isopropanol. The cells were placed in the incubator for 15 min
to solubilize the cells, and were then mixed completely. Absorbance
was read at 570 nm to determine cell proliferation.
Cell transfection
MDA-MB-231 and MDA-MB-468 cells were transfected
with CRABPII siRNA according to the protocol from Invitrogen Life
Technologies (Grand Island, NY, USA). Briefly, 30 pmol of control
or CRABPII siRNA was mixed with 500 µl Gibco Opti-MEM® I
medium without serum by Life Technologies in a 6-well plate.
Lipofectamine™ RNAiMAX (5 µl) was added to the diluted RNAi
molecules for 20 min at room temperature. MDA-MB-231 and MDA-MB-468
(250,000 cells) were diluted in 2.5 ml of antibiotics-free media
and added to the plates containing the RNAi duplex/Lipofectamine™
mixture labeled with control or CRABPII siRNA. After 24 h, the
cells were treated with 10 µM curcumin in the presence or absence
of 1 µM ATRA for 96 h. ATRA was added every 2 days. Cells were then
lysed, protein was extracted, quantitated and protein extract was
loaded onto a gel for SDS-PAGE. The blot was probed with the
appropriate antibodies.
Statistical analysis
Statistical significance of differences between
treatments was determined using two-tailed Student's t-test and
P-values were noted. Differences between groups were considered
statistically significant at p<0.05.
Results
Concentration-dependent impact of
curcumin on CRABPII and RAR expression in TNBC cell lines
Knowing that TNBC cells are resistant to RA due to
the expression patterns of CRABPII and FABP5 (11,20,49),
our previous study demonstrated that 30 µM curcumin reduced the
expression of FABP5 and its cognitive receptor, PPARβ/δ (45). In this study, we sought to examine
the effect of curcumin on the CRABPII/RAR pathway in TNBC cells. As
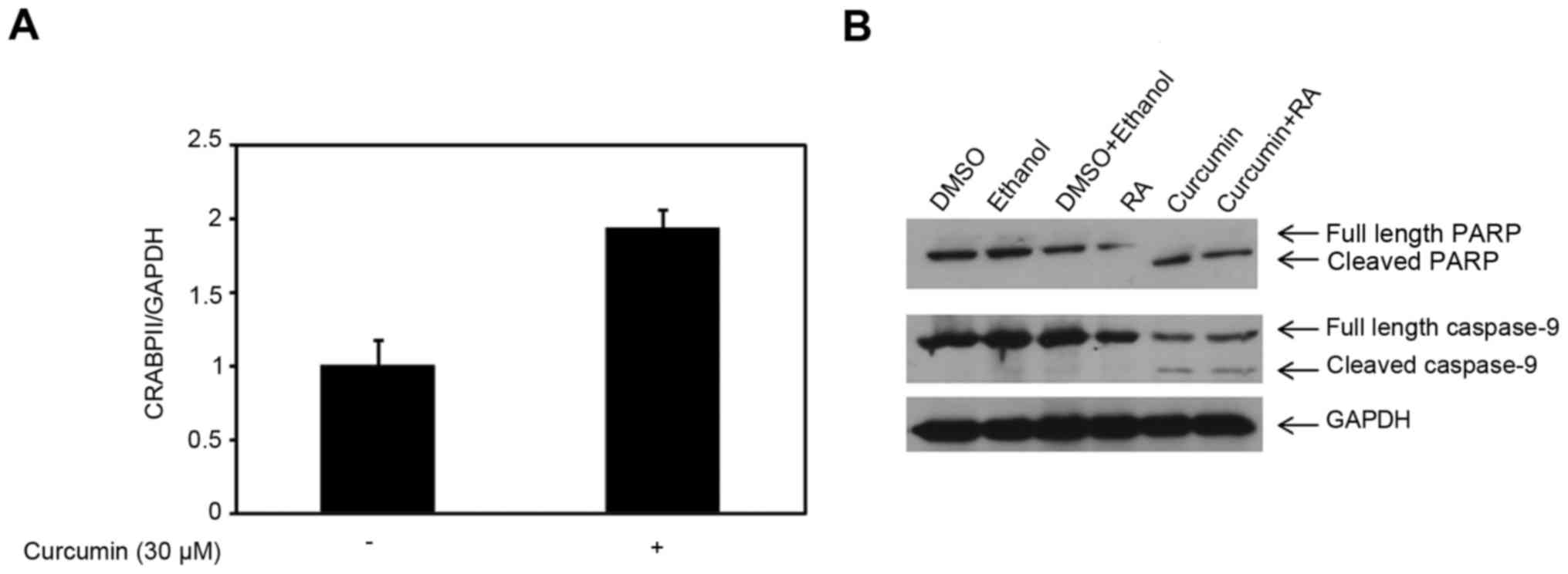
shown in Fig. 1A, 30 µM curcumin
enhanced the CRABPII mRNA expression level compared to control
(p=0.016) in MDA-MB-231 cells, however, had no effect on the RAR
isoforms (data not shown). Since CRABPII delivers RA to RAR
isoforms, we tested whether the induction of CRABPII was sufficient
to sensitize RA-resistant MDA-MB-231 cells to RA-mediated
apoptosis. To investigate cell death, we tested PARP, an indicator
of apoptosis and RAR target, caspase-9. As shown in Fig. 1B, treatment of MDA-MB-231 cells with
30 µM curcumin for 48 h completely converted full length PARP to
cleaved PARP, and thus there was no effect with the combination of
curcumin and RA. Examination of the caspase-9 showed there was no
difference in the expression of cleaved caspase-9 in cells treated
with curcumin compared to the co-treatment with curcumin and ATRA
(Fig. 1B). Despite upregulation of
CRABPII by 30 µM curcumin, this dose of curcumin induced apoptosis
in 48 h, independent of the CRABPII pathway.
Because 30 µM curcumin is a high-dose which alone
induces apoptosis within 48 h, we examined the expression level of
CRABPII and RAR isoforms at lower doses of curcumin (5 and 10 µM).
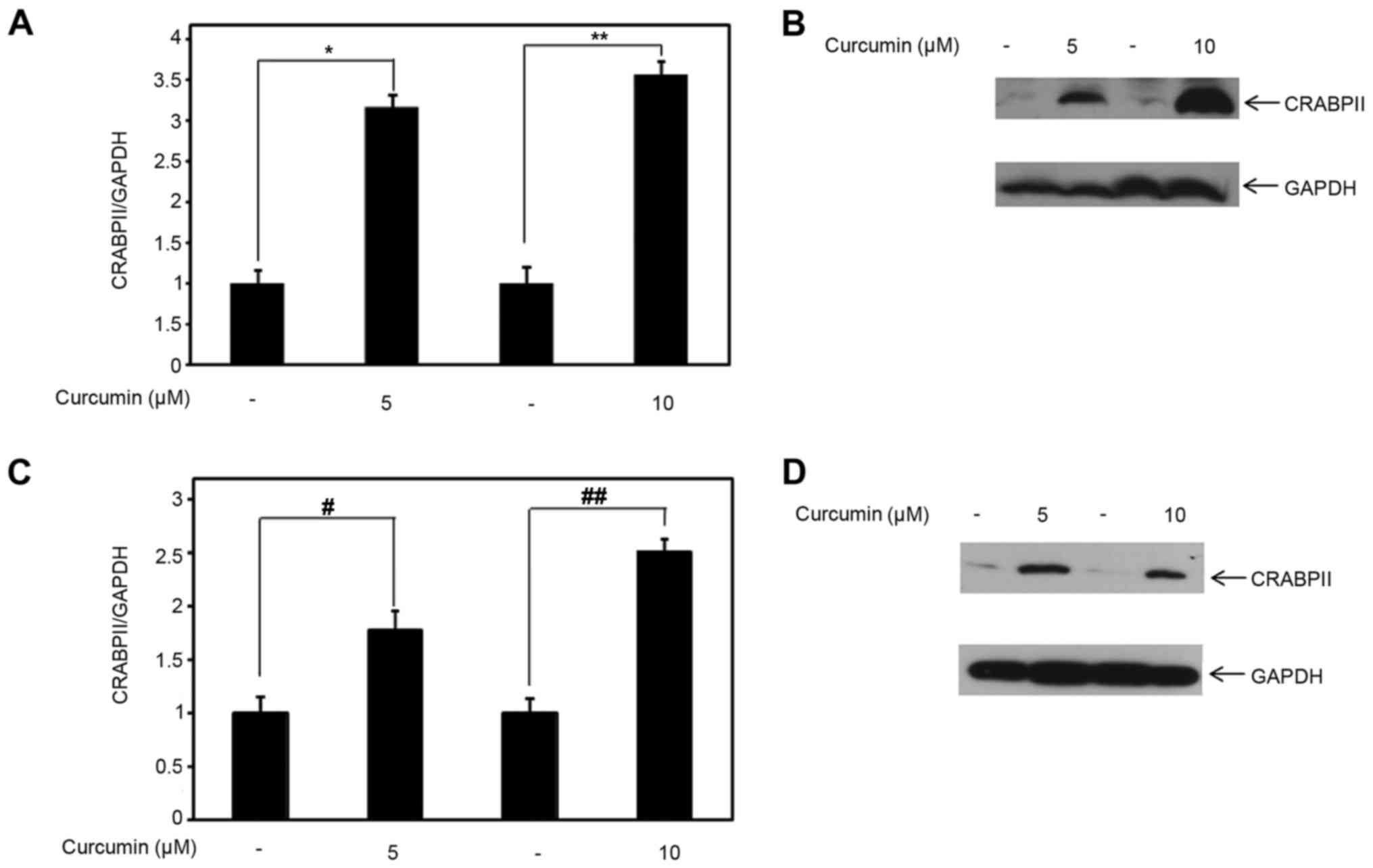
Compared to control, 5 and 10 µM curcumin induced CRABPII mRNA
expression, with statistical significance (p<0.05), in
MDA-MB-231 cells by ~3-3.5-fold (Fig.
2A). Concomitantly, we examined the effect of lower doses of
curcumin on CRABPII protein expression. Consistent with the
upregulation of CRABPII mRNA expression by 5 and 10 µM curcumin, we
also observed that at both of these doses, curcumin induced CRABPII
protein expression in MDA-MB-231 cells (Fig. 2B). Because curcumin affects CRABPII
expression in RA-resistant MDA-MB-231 cells, we further tested
whether curcumin regulates CRABPII mRNA and protein expression in
the TNBC cell line, MDA-MB-468. As shown in Fig. 2C, both of the lower doses of
curcumin, in comparison to control, upregulated the expression of
the CRABPII mRNA (p<0.05). Correspondingly, curcumin induced
CRABPII protein expression level in MDA-MB-468 (Fig. 2D).
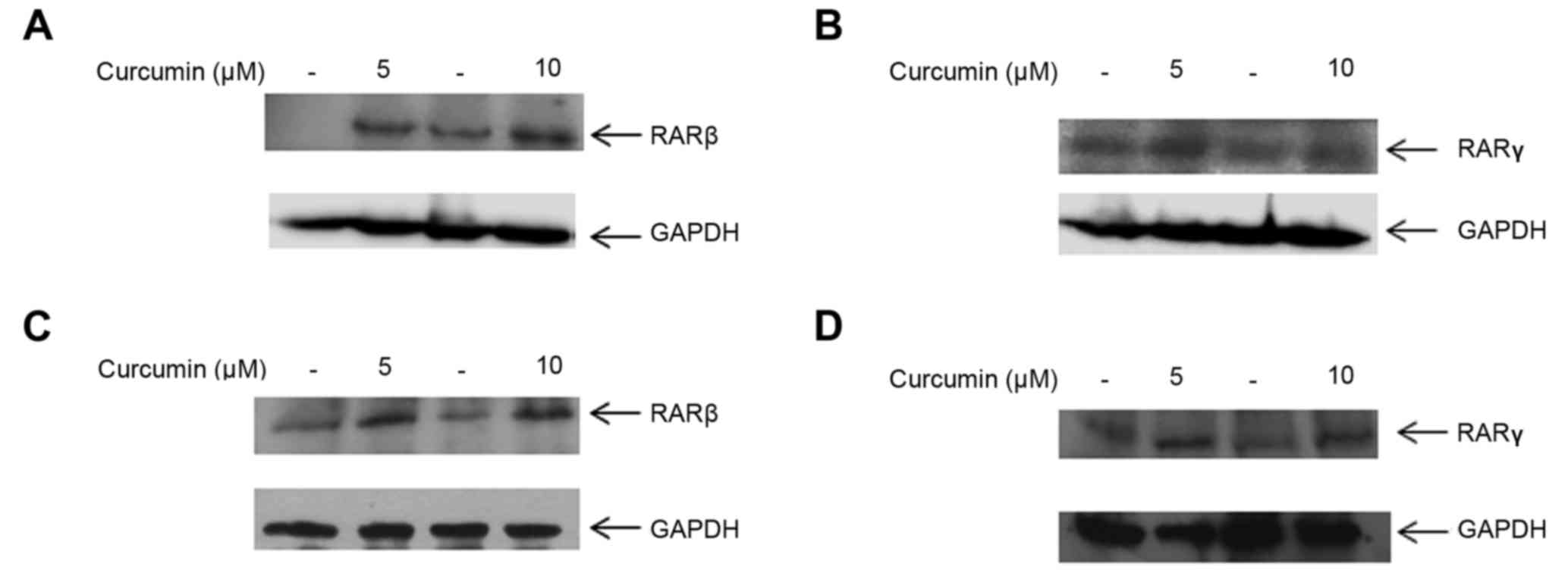
We next investigated whether 5 and 10 µM curcumin
had an effect on the cognate receptors of CRABPII, RAR isoforms α,
β and γ. At both of these doses, RARβ and RARγ protein expression
were upregulated in MDA-MB-231 cells compared to their respective
controls (Fig. 3A and B). To
determine whether regulation of RARβ and RARγ by curcumin is a
global effect among TNBC cells, treatment of MDA-MB-468 cells with
5 and 10 µM curcumin induced both RARβ and RARγ protein expression
(Fig. 3C and D). Interestingly, we
observed that curcumin did not affect the expression of RARα in
either of the TNBC cell lines, MDA-MB-231 and MDA-MB-468 (data not
shown). These results suggest that lower doses of 5 and 10 µM
curcumin enhance RARβ and RARγ protein expression in TNBC
cells.
Lower doses of curcumin sensitize TNBC
cells to RA-mediated apoptosis
Suppression of cell growth by RA is followed by
RA-induced apoptosis. Previously, we showed that treatment of TNBC
cells with 30 µM curcumin for 48 h sensitized these cells to
RA-mediated growth suppression by reducing the expression level of
the FABP5/PPARβ/δ pathway (45). In
this study, we observed that lower doses of curcumin (5 and 10 µM)
induced the expression of CRABPII, RARβ and RARγ (Figs. 2 and 3). To determine the effect of lower doses
of curcumin on sensitizing TNBC cells to RA-mediated apoptosis, we
treated MDA-MB-231 and MDA-MB-468 cells with 5 and 10 µM curcumin
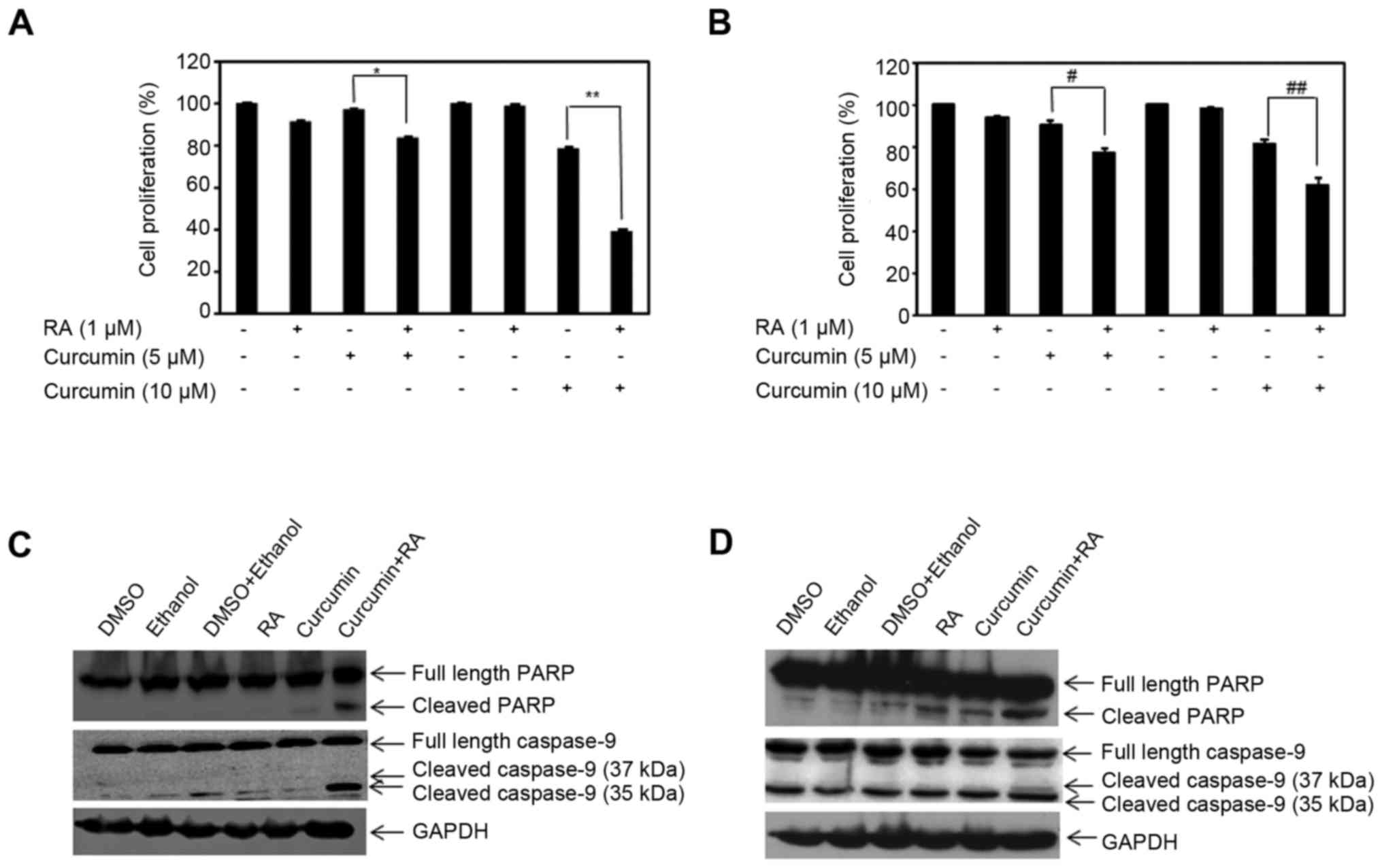
in the presence or absence of 1 µM ATRA for 72 h. As expected, RA
did not inhibit cell growth in MDA-MB-231 cells (Fig. 4A). While the lower concentration of
curcumin (5 µM) did not suppress growth of MDA-MB-231 cells after
72 h, 10 µM curcumin did reduce growth compared to control, with
statistical significance (p=0.03) by ~20% (Fig. 4A). Combining either 5 or 10 µM
curcumin with ATRA sensitized MDA-MB-231 cells to RA-mediated
growth suppression, with a robust effect with 10 µM curcumin and
ATRA (Fig. 4A). Similar findings
were concluded in MDA-MB-468 cells. RA treatment had marginal
growth inhibitory effects on MDA-MB-468 cells (p<0.05). In
comparison to control treated cells, lower doses of curcumin (5 and
10 µM) statistically reduced growth of MDA-MB-468 cells (p<0.05)
but the combination of curcumin with RA sensitized MDA-MB-468 cells
to RA-induced growth suppression, with a more pronounced effect
with 10 µM curcumin (Fig. 4B).
Knowing that 10 µM curcumin had a greater impact on
sensitization of TNBC cells to RA-induced growth suppression at 72
h (Fig. 4A and B), and the fact
that 10 µM curcumin induced CRABPII, RARβ and RARγ expression in
these cells (Figs. 2 and 3), we tested whether this dose of curcumin
could sensitize TNBC cells to RA-mediated apoptosis. Because the
apoptotic effects of RA occur subsequent to growth suppression, we
examined cell death by curcumin and RA at a later time-point. Cells
were treated with 10 µM curcumin in the presence or absence of 1 µM
ATRA for 96 h and PARP expression was examined. In MDA-MB-231
cells, RA did not activate PARP cleavage, while 10 µM curcumin
potentiated cleaved PARP (Fig. 4C).
Co-treatment of curcumin and RA further enhanced active, cleaved
PARP compared to curcumin treatment alone. Activation of RAR
induces the transcriptional activity of several downstream targets,
such as apoptotic protein caspase-9. Concurrently, we examined the
effect of curcumin and RA on caspase-9 expression in MDA-MB-231
cells. As shown in Fig. 4C,
curcumin alone marginally induced cleaved caspase-9, while the
combination of curcumin with RA enhanced the expression of
cleaved/active caspase-9. Similarly, the effects of ATRA and/or
curcumin on the activation of PARP and caspase-9 was examined in
MDA-MB-468 cells. The combination of curcumin and RA further
enhanced apoptosis as evidenced by increased cleaved PARP in
comparison to curcumin or RA alone (Fig. 4D). In addition, the treatment of
MDA-MB-468 cells with curcumin and RA induced cleavage of caspase-9
(37 kDa) and reduced inactive caspase-9 expression compared to
either treatment (Fig. 4D). Results
from the data demonstrate the ability of curcumin to potentiate the
apoptotic effects of RA and reverse RA-resistance in TNBC by
curcumin-mediated induction of CRABPII and RARs.
Sensitization to RA-mediated apoptosis
by curcumin is regulated by CRABPII
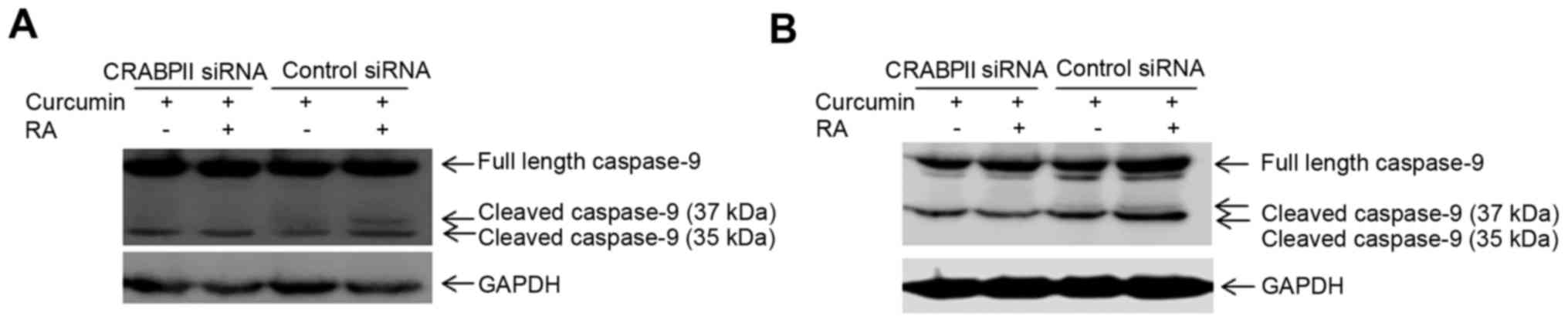
Since CRABPII/RAR is involved in RA-mediated cell
death, we sought to investigate whether upregulation of CRABPII by
curcumin (Fig. 2) regulates
RA-induced apoptosis. To test this hypothesis, we silenced CRABPII
expression in MDA-MB-231 cells, treated the cells with 10 µM
curcumin in the presence or absence of 1 µM ATRA for 96 h and
examined for the protein expression of caspase-9. MDA-MB-231
transfected with control siRNA was also treated with curcumin and
as expected, curcumin induced apoptosis by activation of caspase-9
(37 kDa), which was further enhanced in the presence of RA
(Fig. 5A). However, when the cells
were transfected with CRABPII siRNA, the combination of curcumin
and RA abolished the active 37 kDa form of caspase-9 (Fig. 5A). These results suggest that
curcumin mediated upregulation of CRABPII sensitizes TNBC cells to
RA-mediated apoptosis by the induction of caspase-9, one of the RAR
target genes. We also silenced CRABPII in MDA-MB-468 cells and
combinatorial treatment of the cells with curcumin and RA enhanced
expression of cleaved/active caspase-9 (37 and 35 kDa), in
comparison to curcumin alone (Fig.
5B). However, knockdown of CRABPII reduced active caspase-9 (35
kDa) and abolished the expression of the cleaved/active 37 kDa
caspase-9 (Fig. 5B).
Discussion
Sensitivity of cancer cells to RA is determined by
two distinct pathway, CRABPII/RAR and FABP5/PPARβ/δ (10,11,14,
20,49). While the delivery of RA to the
FABP5/PPARβ/δ pathway results in increased proliferation,
transporting RA by CRABPII to RAR inhibits proliferation and
promotes apoptosis (10,11,14).
Reducing the ratio of FABP5:CRABPII can overcome the resistance of
cancer cells to RA by shifting the delivery of RA to RAR by CRABPII
instead of activating the FABP5/PPARβ/δ pathway (10,11,14).
Phytochemicals such as curcumin exhibit
chemosensitizing properties which may circumvent toxicity issues
faced with traditional chemotherapeutic agents (50). While they are less toxic and have no
side effects, combinatorial treatment of phytochemicals with
chemotherapeutic agents can be an alternative method to reduce the
dosage of traditional chemotherapeutic regimens and lower cardiac
toxicity associated with them (50). Owing to drug resistance in cancer,
curcumin has been shown to be an effective adjuvant in reversing
chemoresistance and sensitizing cancer cells to chemotherapeutic
drugs (41–47,51–54).
Our previous study reported that the potential
therapeutic activity of curcumin to sensitize RA-resistant TNBC
cells to RA-mediated growth suppression was exhibited through the
inhibition of the FABP5/PPARβ/δ pathway (45). The present study focused on the
effect of curcumin on the CRABPII/RAR pathway to reverse resistance
of TNBC cells to RA by activation of apoptosis. Results of the
present study demonstrate that low concentrations of 5 and 10 µM
curcumin increase the expression level of CRABPII, while
concomitantly induces expression of its cognate receptor, RARβ and
RARγ. Similarly, a recent study demonstrated that curcumin
upregulated the expression of RARβ in several cancer cells,
including TNBC MDA-MB-231 cells and activated RARβ via epigenetic
regulation (22). Though the RAR
subtypes have different functions and regulate distinct RARE
targeted genes, RAR signaling plays a critical role in cancer
progression. While RARβ is epigenetically silenced in tumor cells
and its loss has been associated with lymph node metastasis
(21,55,56),
RARγ has been implicated as a tumor suppressor and restoration of
this retinoid receptor reverses the tumorigenic potential of mouse
keratinocytes (57,58). While curcumin induces the expression
of several regulatory genes involved in apoptosis (59), we present evidence in this study
that curcumin upregulates genes involved in the retinoid signaling
pathway, namely CRABPII, RARβ and RARγ in mammary carcinoma
MDA-MB-231 and MDA-MB-468 cells.
Although 30 µM of curcumin induces the mRNA
expression level of CRABPII, it did not affect the expression of
RARs. However, lower doses of curcumin (5 and 10 µM) induces the
protein expression level of not only CRABPII but also RARβ and RARγ
in two TNBC cell lines suggesting that this is a global effect
among TNBC cells. Alterations in gene expression associated with
different doses of curcumin are not unusual and have been well
documented with other genes (60).
The differential effects of curcumin have been observed with
several apoptotic genes that are upregulated or downregulated by
only higher dose of curcumin, while the reverse has also been
observed for some genes that are upregulated only at lower doses of
curcumin (60). As noted in this
study, the differences in concentration of curcumin to induce the
CRABPII/RAR pathway may be an important determinant in identifying
the required optimum dosage to sensitize TNBC cells to RA for
future in vivo studies.
The concentration-dependent regulation of CRABPII
and RARs by curcumin determined the outcome on the activation of
apoptotic proteins, PARP and caspase-9. Although 30 µM curcumin
upregulates the mRNA level of CRABPII, this dose of curcumin
completely activates PARP and induces activation of caspase-9 in 48
h. However, the fact that 30 µM curcumin does not regulate the RARs
indicates that this concentration of curcumin induces apoptosis
independent of the CRABPII/RAR and 30 µM curcumin does not
sensitize MDA-MB-231 cells to RA-induced apoptosis.
Curcumin has a differential effect on gene
regulation and cell death initiated by this agent dose- and
time-dependently (60,61). The present data demonstrates that
curcumin can re-activate the CRABPII/RAR pathway in TNBC cells and
cause RA to initiate apoptosis by activation of PARP and caspase-9.
Such doses of curcumin (5 and 10 µM) upregulates RARβ and RARγ, as
well as CRABPII in TNBC cells. Combination of 10 µM curcumin with
RA for 96 h sensitizes TNBC cells to apoptosis mediated by RA as
evidenced by increased PARP cleavage. Because 10 µM curcumin
induces RARβ and RARγ in TNBC cells, curcumin at this dose
sensitizes the cells to RA-mediated apoptosis through RAR-dependent
activation of caspase-9. Initiation of cell death by RAR itself is
not sufficient to regulate apoptosis by RA, and hence shuttling of
RA from the cytosol to the nucleus by CRABPII facilitates binding
of RA to RARs and enhances the transcriptional activation of genes
such as caspase-9 involved in the retinoid signaling pathway. To
extend these studies and gain a mechanistic understanding on the
role of curcumin on the CRABPII/RAR pathway, our results also
provide evidence that silencing CRABPII prevents curcumin from
sensitizing TNBC cells to RA-induced activation of caspase-9. Taken
together, our data suggest that in order to activate cell death by
RA in RA-resistant TNBC cells, CRABPII and RAR pathway have to be
upregulated by lower concentrations of curcumin and these two
proteins work in concert to sensitize cells to RA-mediated
apoptosis.
In conclusion, the present study revealed that
reversing the resistance of TNBC to RA-induced apoptosis is
dependent on the dose of curcumin and length of treatment.
Accordingly, lower concentrations of curcumin induce CRABPII, RARβ
and RARγ, and thus upregulation of CRABPII/RAR pathway contributes
to the sensitization of TNBC cells to apoptosis by RA. As such this
study highlights a novel mechanism by which RA-resistant mammary
carcinoma cells can be resensitized to RA-mediated apoptosis by
curcumin. The effectiveness in the combination of curcumin with RA
warrants further consideration for its use in RA-resistant TNBC
cells. Overall, this study provides mechanistic insights on the
role of curcumin to reverse RA resistance in breast cancer cells
through the regulation of the CRABPII/RAR pathway, and highlights
the potential of using curcumin as a therapeutic adjuvant in RA
resistant cancers.
Acknowledgements
This study was supported by the Research and
Scholarship Development Grant Program, University of South Alabama,
Office of Research and Economic Development and the start-up funds
from the College of Allied Health Professions at University of
South Alabama. We thank the Department of Pharmacology, University
of South Alabama for use of their film developer.
Glossary
Abbreviations
Abbreviations:
|
RA
|
retinoic acid
|
|
CRABPII
|
cellular retinoic acid- binding
protein II
|
|
RAR
|
retinoic acid receptor
|
|
TNBC
|
triple-negative breast cancer
|
|
PARP
|
poly(ADP-ribose) polymerase
|
|
ER
|
estrogen receptor
|
|
PR
|
progesterone receptor
|
|
HER2
|
human epidermal growth factor receptor
2
|
|
ATRA
|
all-trans-retinoic acid
|
|
PPARβ/δ
|
peroxisome proliferator-activated
receptor β/δ
|
|
FABP5
|
fatty acid-binding protein 5
|
|
GAPDH
|
glyceraldehyde 3-phosphate
dehydrogenase
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
DMEM
|
Dulbeccos modified Eagles medium
|
|
FBS
|
fetal bovine serum
|
|
qRT-PCR
|
quantitative real-time polymerase
chain reaction
|
|
DMSO
|
dimethyl sulfoxide
|
References
|
1
|
Boyle P: Triple-negative breast cancer:
epidemiological considerations and recommendations. Ann Oncol.
23:(Suppl 6). vi7–vi12. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lo-Coco F, Avvisati G, Vignetti M, Thiede
C, Orlando SM, Iacobelli S, Ferrara F, Fazi P, Cicconi L, Di Bona
E, et al: Gruppo Italiano Malattie Ematologiche dellAdulto;
German-Austrian Acute Myeloid Leukemia Study Group; Study Alliance
Leukemia: Retinoic acid and arsenic trioxide for acute
promyelocytic leukemia. N Engl J Med. 369:111–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chambon P: A decade of molecular biology
of retinoic acid receptors. FASEB J. 10:940–954. 1996.PubMed/NCBI
|
|
4
|
Germain P, Chambon P, Eichele G, Evans RM,
Lazar MA, Leid M, De Lera AR, Lotan R, Mangelsdorf DJ and
Gronemeyer H: International Union of Pharmacology. LXIII. Retinoid
X receptors. Pharmacol Rev. 58:760–772. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koeffler HP: Is there a role for
differentiating therapy in non-APL AML? Best Pract Res Clin
Haematol. 23:503–508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Donato LJ, Suh JH and Noy N: Suppression
of mammary carcinoma cell growth by retinoic acid: the cell cycle
control gene Btg2 is a direct target for retinoic acid receptor
signaling. Cancer Res. 67:609–615. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Altucci L, Rossin A, Raffelsberger W,
Reitmair A, Chomienne C and Gronemeyer H: Retinoic acid-induced
apoptosis in leukemia cells is mediated by paracrine action of
tumor-selective death ligand TRAIL. Nat Med. 7:680–686. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Donato LJ and Noy N: Suppression of
mammary carcinoma growth by retinoic acid: proapoptotic genes are
targets for retinoic acid receptor and cellular retinoic
acid-binding protein II signaling. Cancer Res. 65:8193–8199. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kitareewan S, Pitha-Rowe I, Sekula D,
Lowrey CH, Nemeth MJ, Golub TR, Freemantle SJ and Dmitrovsky E:
UBE1L is a retinoid target that triggers PML/RARalpha degradation
and apoptosis in acute promyelocytic leukemia. Proc Natl Acad Sci
USA. 99:3806–3811. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schug TT, Berry DC, Shaw NS, Travis SN and
Noy N: Opposing effects of retinoic acid on cell growth result from
alternate activation of two different nuclear receptors. Cell.
129:723–733. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schug TT, Berry DC, Toshkov IA, Cheng L,
Nikitin AY and Noy N: Overcoming retinoic acid-resistance of
mammary carcinomas by diverting retinoic acid from PPARbeta/delta
to RAR. Proc Natl Acad Sci USA. 105:7546–7551. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tan NS, Shaw NS, Vinckenbosch N, Liu P,
Yasmin R, Desvergne B, Wahli W and Noy N: Selective cooperation
between fatty acid binding proteins and peroxisome
proliferator-activated receptors in regulating transcription. Mol
Cell Biol. 22:5114–5127. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Di-Poï N, Tan NS, Michalik L, Wahli W and
Desvergne B: Antiapoptotic role of PPARbeta in keratinocytes via
transcriptional control of the Akt1 signaling pathway. Mol Cell.
10:721–733. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wolf G: Cellular retinoic acid-binding
protein II: a coactivator of the transactivation by the retinoic
acid receptor complex RAR. RXR. Nutr Rev. 58:151–153. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
El-Metwally TH, Hussein MR, Pour PM,
Kuszynski CA and Adrian TE: Natural retinoids inhibit proliferation
and induce apoptosis in pancreatic cancer cells previously reported
to be retinoid resistant. Cancer Biol Ther. 4:474–483. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
El-Metwally TH, Hussein MR, Pour PM,
Kuszynski CA and Adrian TE: High concentrations of retinoids induce
differentiation and late apoptosis in pancreatic cancer cells in
vitro. Cancer Biol Ther. 4:602–611. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vreeland AC, Levi L, Zhang W, Berry DC and
Noy N: Cellular retinoic acid-binding protein 2 inhibits tumor
growth by two distinct mechanisms. J Biol Chem. 289:34065–34073.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Favorskaya I, Kainov Y, Chemeris G,
Komelkov A, Zborovskaya I and Tchevkina E: Expression and clinical
significance of CRABP1 and CRABP2 in non-small cell lung cancer.
Tumour Biol. 35:10295–10300. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Passeri D, Doldo E, Tarquini C, Costanza
G, Mazzaglia D, Agostinelli S, Campione E, Di Stefani A, Giunta A,
Bianchi L, et al: Loss of CRABP-II characterizes human skin poorly
differentiated squamous cell carcinomas and favors DMBA/TPA-induced
carcinogenesis. J Invest Dermatol. 136:1255–1266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gupta S, Pramanik D, Mukherjee R, Campbell
NR, Elumalai S, de Wilde RF, Hong SM, Goggins MG, De Jesus-Acosta
A, Laheru D, et al: Molecular determinants of retinoic acid
sensitivity in pancreatic cancer. Clin Cancer Res. 18:280–289.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Albino-Sanchez ME, Vazquez-Hernandez J,
Ocadiz-Delgado R, Serafin-Higuera N, León-Galicia I, Garcia-Villa
E, Hernandez-Pando R and Gariglio P: Decreased RARβ expression
induces abundant inflammation and cervical precancerous lesions.
Exp Cell Res. 346:40–52. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang A, Wang X, Shan X, Li Y, Wang P,
Jiang P and Feng Q: Curcumin reactivates silenced tumor suppressor
gene RARβ by reducing DNA methylation. Phytother Res. 29:1237–1245.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu XC: Tumor-suppressive activity of
retinoic acid receptor-beta in cancer. Cancer Lett. 253:14–24.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Darwiche N, Celli G, Tennenbaum T, Glick
AB, Yuspa SH and De Luca LM: Mouse skin tumor progression results
in differential expression of retinoic acid and retinoid X
receptors. Cancer Res. 55:2774–2782. 1995.PubMed/NCBI
|
|
25
|
Xu XC, Wong WY, Goldberg L, Baer SC, Wolf
JE, Ramsdell WM, Alberts DS, Lippman SM and Lotan R: Progressive
decreases in nuclear retinoid receptors during skin squamous
carcinogenesis. Cancer Res. 61:4306–4310. 2001.PubMed/NCBI
|
|
26
|
Brigger D, Schläfli AM, Garattini E and
Tschan MP: Activation of RARα induces autophagy in SKBR3 breast
cancer cells and depletion of key autophagy genes enhances ATRA
toxicity. Cell Death Dis. 6:e18612015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tomita A, Kiyoi H and Naoe T: Mechanisms
of action and resistance to all-trans retinoic acid (ATRA)
and arsenic trioxide (As2O3) in acute
promyelocytic leukemia. Int J Hematol. 97:717–725. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Applegate CC and Lane MA: Role of
retinoids in the prevention and treatment of colorectal cancer.
World J Gastrointest Oncol. 7:184–203. 2015.PubMed/NCBI
|
|
29
|
Ak T and Gülçin I: Antioxidant and radical
scavenging properties of curcumin. Chem Biol Interact. 174:27–37.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Joe B, Vijaykumar M and Lokesh BR:
Biological properties of curcumin-cellular and molecular mechanisms
of action. Crit Rev Food Sci Nutr. 44:97–111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Yu J, Cui R, Lin J and Ding X:
Curcumin in treating breast cancer (Review). J Lab Autom.
21:723–731. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bimonte S, Barbieri A, Leongito M,
Piccirillo M, Giudice A, Pivonello C, de Angelis C, Granata V,
Palaia R and Izzo F: Curcumin anticancer studies in pancreatic
cancer. Nutrients. 8:82016. View Article : Google Scholar :
|
|
33
|
Jordan BC, Mock CD, Thilagavathi R and
Selvam C: Molecular mechanisms of curcumin and its semisynthetic
analogues in prostate cancer prevention and treatment. Life Sci.
152:135–144. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Howells LM, Mahale J, Sale S, McVeigh L,
Steward WP, Thomas A and Brown K: Translating curcumin to the
clinic for lung cancer prevention: evaluation of the preclinical
evidence for its utility in primary, secondary, and tertiary
prevention strategies. J Pharmacol Exp Ther. 350:483–494. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chainani-Wu N: Safety and
anti-inflammatory activity of curcumin: a component of tumeric
(Curcuma longa). J Altern Complement Med. 9:161–168. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cheah YH, Nordin FJ, Sarip R, Tee TT,
Azimahtol HL, Sirat HM, Rashid BA, Abdullah NR and Ismail Z:
Combined xanthorrhizol-curcumin exhibits synergistic growth
inhibitory activity via apoptosis induction in human breast cancer
cells MDA-MB-231. Cancer Cell Int. 9:12009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Javvadi P, Segan AT, Tuttle SW and
Koumenis C: The chemopreventive agent curcumin is a potent
radiosensitizer of human cervical tumor cells via increased
reactive oxygen species production and overactivation of the
mitogen-activated protein kinase pathway. Mol Pharmacol.
73:1491–1501. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Anitha A, Maya S, Deepa N, Chennazhi KP,
Nair SV and Jayakumar R: Curcumin-loaded N,O-carboxymethyl chitosan
nanoparticles for cancer drug delivery. J Biomater Sci Polym Ed.
23:1381–1400. 2012.PubMed/NCBI
|
|
39
|
Mukerjee A and Vishwanatha JK:
Formulation, characterization and evaluation of curcumin-loaded
PLGA nanospheres for cancer therapy. Anticancer Res. 29:3867–3875.
2009.PubMed/NCBI
|
|
40
|
Ma Z, Haddadi A, Molavi O, Lavasanifar A,
Lai R and Samuel J: Micelles of poly(ethylene
oxide)-b-poly(epsilon-caprolactone) as vehicles for the
solubilization, stabilization, and controlled delivery of curcumin.
J Biomed Mater Res A. 86:300–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bordoloi D, Roy NK, Monisha J, Padmavathi
G and Kunnumakkara AB: Multi-targeted agents in cancer cell
chemosensitization: what we learnt from curcumin thus far. Recent
Patents Anticancer Drug Discov. 11:67–97. 2016. View Article : Google Scholar
|
|
42
|
Saha S, Adhikary A, Bhattacharyya P, Das T
and Sa G: Death by design: where curcumin sensitizes drug-resistant
tumours. Anticancer Res. 32:2567–2584. 2012.PubMed/NCBI
|
|
43
|
Chen P, Li J, Jiang HG, Lan T and Chen YC:
Curcumin reverses cisplatin resistance in cisplatin-resistant lung
caner cells by inhibiting FA/BRCA pathway. Tumour Biol.
36:3591–3599. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Toden S, Okugawa Y, Jascur T, Wodarz D,
Komarova NL, Buhrmann C, Shakibaei M, Boland CR and Goel A:
Curcumin mediates chemosensitization to 5-fluorouracil through
miRNA-induced suppression of epithelial-to-mesenchymal transition
in chemoresistant colorectal cancer. Carcinogenesis. 36:355–367.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Thulasiraman P, McAndrews DJ and
Mohiudddin IQ: Curcumin restores sensitivity to retinoic acid in
triple negative breast cancer cells. BMC Cancer. 14:7242014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou B, Huang J, Zuo Y, Li B, Guo Q, Cui
B, Shao W, Du J and Bu X: 2a, a novel curcumin analog, sensitizes
cisplatin-resistant A549 cells to cisplatin by inhibiting
thioredoxin reductase concomitant oxidative stress damage. Eur J
Pharmacol. 707:130–139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jiang M, Huang O, Zhang X, Xie Z, Shen A,
Liu H, Geng M and Shen K: Curcumin induces cell death and restores
tamoxifen sensitivity in the antiestrogen-resistant breast cancer
cell lines MCF-7/LCC2 and MCF-7/LCC9. Molecules. 18:701–720. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2[-Delta Delta C(T)] method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liu RZ, Graham K, Glubrecht DD, Germain
DR, Mackey JR and Godbout R: Association of FABP5 expression with
poor survival in triple-negative breast cancer: implication for
retinoic acid therapy. Am J Pathol. 178:997–1008. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Prasad NR, Muthusamy G, Shanmugam M and
Ambudkar SV: South Asian medicinal compounds as modulators of
resistance to chemotherapy and radiotherapy. Cancers (Basel).
8:82016. View Article : Google Scholar :
|
|
51
|
Montgomery A, Adeyeni T, San K, Heuertz RM
and Ezekiel UR: Curcumin sensitizes silymarin to exert synergistic
anticancer activity in colon cancer cells. J Cancer. 7:1250–1257.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
de Ruiz Porras V, Bystrup S,
Martínez-Cardús A, Pluvinet R, Sumoy L, Howells L, James MI, Iwuji
C, Manzano JL, Layos L, et al: Curcumin mediates
oxaliplatin-acquired resistance reversion in colorectal cancer cell
lines through modulation of CXC-chemokine/NF-κB signalling pathway.
Sci Rep. 6:246752016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Selvendiran K, Ahmed S, Dayton A,
Kuppusamy ML, Rivera BK, Kálai T, Hideg K and Kuppusamy P: HO-3867,
a curcumin analog, sensitizes cisplatin-resistant ovarian
carcinoma, leading to therapeutic synergy through STAT3 inhibition.
Cancer Biol Ther. 12:837–845. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chanvorachote P, Pongrakhananon V,
Wannachaiyasit S, Luanpitpong S, Rojanasakul Y and Nimmannit U:
Curcumin sensitizes lung cancer cells to cisplatin-induced
apoptosis through superoxide anion-mediated Bcl-2 degradation.
Cancer Invest. 27:624–635. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
di Masi A, Leboffe L, De Marinis E, Pagano
F, Cicconi L, Rochette-Egly C, Lo-Coco F, Ascenzi P and Nervi C:
Retinoic acid receptors: from molecular mechanisms to cancer
therapy. Mol Aspects Med. 41:1–115. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Flamini MI, Gauna GV, Sottile ML, Nadin
BS, Sanchez AM and Vargas-Roig LM: Retinoic acid reduces migration
of human breast cancer cells: role of retinoic acid receptor beta.
J Cell Mol Med. 18:1113–1123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen CF, Goyette P and Lohnes D: RARgamma
acts as a tumor suppressor in mouse keratinocytes. Oncogene.
23:5350–5359. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hatoum A, El-Sabban ME, Khoury J, Yuspa SH
and Darwiche N: Overexpression of retinoic acid receptors alpha and
gamma into neoplastic epidermal cells causes retinoic acid-induced
growth arrest and apoptosis. Carcinogenesis. 22:1955–1963. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shehzad A, Wahid F and Lee YS: Curcumin in
cancer chemoprevention: molecular targets, pharmacokinetics,
bioavailability, and clinical trials. Arch Pharm (Weinheim).
343:489–499. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ramachandran C, Rodriguez S, Ramachandran
R, Nair Raveendran PK, Fonseca H, Khatib Z, Escalon E and Melnick
SJ: Expression profiles of apoptotic genes induced by curcumin in
human breast cancer and mammary epithelial cell lines. Anticancer
Res. 25:3293–3302. 2005.PubMed/NCBI
|
|
61
|
Van Erk MJ, Teuling E, Staal YC, Huybers
S, Van Bladeren PJ, Aarts JM and Van Ommen B: Time- and
dose-dependent effects of curcumin on gene expression in human
colon cancer cells. J Carcinog. 3:82004. View Article : Google Scholar : PubMed/NCBI
|