Introduction
Chronic myeloid leukemia (CML) is a clonal disease
of hematopoietic stem cells characterized by a reciprocal
translocation between chromosome 9 and 22 [t(9;22) (q34;q11)], an
event that leads to the formation of the BCR-ABL oncogene which
encodes the BCR-ABL oncogenic fusion protein with constitutive
tyrosine kinase activity (1,2). The
incidence of CML is approximately one individual/100,000
population/year with a slight male predominance and rising
incidence with age, accounting for ~15% of newly diagnosed cases of
leukemia in adults (1). Since
characterized by the constitutively active BCR-ABL, an important
understanding of the molecular pathogenesis of CML has afforded the
potential for target-oriented therapy. As expected, the advent of
tyrosine kinase inhibitors (TKIs) has drastically revolutionized
the treatment prognosis of CML patients from a potentially fatal
disorder to a disorder amenable simply with lifelong oral
medication and compatible with a normal lifespan (1,2).
However, ~10–15% of patients with CML remain resistant to TKIs and
more than 62.5% of the patients may experience multidrug resistance
(MDR) at risk of disease progression (1,3). The
acquisition of MDR has become a major clinical problem and
continues to be a critical obstacle for chemotherapy regimen
treatment in CML. Therefore, elucidation of the molecular
mechanisms underlying cancer MDR has been performed extensively in
order to identify putative targets in human cancers.
MDR is considered to possess a multifactorial
phenotype and occurs as a result of two most crucial causes: the
overexpression of the inhibitors of apoptosis proteins (IAPs) and
cell membrane-bound adenosine triphosphate (ATP)-binding cassette
(ABC) transporters (4–8). It has been recognized that resistance
to apoptosis promotes the genesis and progression of cancer and
helps cancer cells escape from chemotherapy-induced cytotoxicity,
thus, causing cancer cell chemoresistance (6,9,10).
Apollon/BIR-containing protein 6 (BIRC6; also known as BRUCE) the
largest and the only membrane-bound member of the IAP family, which
is located on the outer membrane of the Golgi apparatus, plays an
important role in cancer genesis and progression and is always
abnormally overexpressed in a variety of human cancers (9,11,12).
ABC transporters which work as drug efflux pumps, play critical
roles in human cancer cell resistance to chemotherapy drugs. Among
these ABC transporters, multidrug resistance 1 [MDR1; also known as
P-glycoprotein (P-gp) or ABCB1] is the most representative and
relevant drug transporter and plays a critical role in human CML
cell resistance to chemotherapy drugs (4,8,13).
Although IAPs and P-gp are always overexpressed and
coexist in multidrug resistant cancer cell lines (5,14–16),
the interrelationship between IAPs and ABC transporters is far from
being completely understood. Clones of adriamycin (ADM) resistant
CML K562/ADM cells were established by continuous exposure to ADM.
Our previous study, demonstrated that the ADM-selected K562/ADM
subline of our laboratory revealed a 31.78-fold higher resistance
to ADM when compared with the parental K562 cells (17). In the present study, we investigated
the effect of short hairpin RNA (shRNA) knockdown of the Apollon or
MDR1 gene combined with ADM on the chemosensitivity of the human
CML cell line K562/ADM, and evaluated the possible relationship
between Apollon and MDR1 expression and the targeted gene therapy
response.
Materials and methods
Chemicals and reagents
ADM purchased from Melone Pharmaceutical Co., Ltd.
(Dalian, China) was dissolved in ddH2O at a
concentration of 2 g/l and stored at −20°C. Anti-Apollon/BIRC6,
anti-MDR1/P-gp and anti-GADPH rabbit polyclonal antibodies (pAb)
were respectively obtained from Abcam Inc. (Cambridge, UK), Bioss
Bio-Technology Co., Ltd. (Beijing, China) and Goodhere
Biotechnology Co., Ltd. (Hangzhou, China).
Cell lines and cell culture
The human CML K562 cells were obtained from the Key
Laboratory of Tumour Molecular Biology of Binzhou Medical
University (Binzhou, China) and its MDR subline K562/ADM was
obtained from the Department of Pharmacology at The Institute of
Hematology of the Chinese Academy of Medical Sciences (Tianjin,
China). The cells were maintained in RPMI-1640 medium supplemented
with 10% fetal bovine serum (FBS; both from HyClone, Logan, UT,
USA), 100 U/ml of penicillin and 0.1 mg/ml streptomycin at 37°C in
a humidified atmosphere containing 5% CO2. K562/ADM
cells were cultured in the same medium with an additional 4 mg/l
ADM. Before the experiment, K562/ADM cells were cultured in
drug-free medium for 72 h.
Determination of gene expression by
reverse transcription-polymerase chain reaction (RT-PCR)
Approximately 3×106 cells were harvested
for RT-PCR analysis. Total RNA was isolated from the cells using
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer's instructions. Total RNA was reverse transcribed to
cDNA and stored at −20°C. Primers were designed using Primer 5
version 5.6.0 software and synthesized by Sangon Biotech Co., Ltd.
(Shanghai, China). Primers used in RT-PCR were as follows: Apollon
forward, 5′-TGGCTCAAGCTGGATTTTAT-3′ and reverse,
5′-TTCAGACCAAGGTTCATCAG-3′; MDR1 forward,
5′-GGAGCCTACTTGGTGGCACATAA-3′ and reverse,
5′-TGGCATAGTCAGGAGCAAATGAAC-3′; GAPDH forward,
5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse, 5′-GAAGATGGTGATGGGATTTC-3′.
RT-PCR was performed on an ABI Prism 7500 Real-Time PCR system
(Applied Biosystems, Foster City, CA, USA) using a SYBR-Green
Reaction kit (Takara Bio, Otsu, Shiga, Japan). The reaction system
of the PCR was: SYBR-Green reagent, forward and reverse primer,
template cDNA and nuclease-free distilled water. PCR programs were
carried out as follows: 95°C for 30 sec, followed by 45 cycles at
95°C for 5 sec and 60°C for 30 sec. GAPDH served as an internal
control. The PCR products were separated by 1% agarose gels (Takara
Biotechnology Co., Ltd., Dalian, China) and stained with ethidium
bromide for 15 min. The gels were scanned and analyzed by the Tanon
Gel Imaging system. RT-PCR for each gene of each cDNA sample was
assayed in triplicate.
Plasmid construction and stable
transfection of cells
The Apollon-targeted shRNA (termed as shApollon),
MDR1-targeted shRNA (termed as shMDR1) and the negative control
shRNA (termed as shControl) (Table
I) were synthesized by recombinant plasmids containing the
green fluorescent protein (GFP) vector, pGPH1, purchased from
Shanghai GenePharma Co., Ltd. (Shanghai, China) after being
chemically synthesized, annealed, desalted and purified. The
shApollon and shMDR1 sequences were selected according to a
previous study in our laboratory and the research was performed by
Du et al, respectively (18,19).
Both of the constructs were confirmed by sequence analysis.
 | Table I.Synthetic Apollon and MDR1 shRNA
sequence. |
Table I.
Synthetic Apollon and MDR1 shRNA
sequence.
| Genes |
|
| Forward and reverse
primers |
|---|
| Apollon | F |
5′-CUGCCUCUUUCAGGCAAUATT-3′ |
|
| R |
5′-UAUUGCCUGAAAGAGGCAGTT-3′ |
| MDR1 | F |
5′-GATCCCGAAACCAACTGTCAGTGTATCAAGAGTACACTGACAGTTGGTTTCTTTTTT-3′ |
|
| R |
5′-CTAGAAAAAAGAAACCAACTGTCAGTGTACTCTTGATACACTGACAGTTGGTTTCGG-3′ |
| Control | F |
5′-GTTCTCCGAACGTGTCACGTCAAG-3′ |
|
| R |
5′-GATTACGTGACACGTTCGGAGAATT-3′ |
Prior to transfection, K562/ADM cells were seeded
into 6-well plates and cultured for 12 h in serum-free RPMI-1640
medium and antibiotics to 60–80% confluency. Transfection was
performed at a final concentration of 1 µg of plasmid-expressing
shRNA using Lipofectamine 2000 transfection reagent (Invitrogen)
according to the manufacturer's instructions. Six hours after
transfection, RPMI-1640 medium containing 10% FBS was added, and
the cells were then incubated under the aforementioned conditions.
G418 (500–1,000 ng/ml; Sigma, St. Louis, MO, USA) was then added to
the medium after 24 h of transfection, and the cells were cultured
for 1 month to permit selection. Stable cells were assessed by
fluorescence microscopy (Olympus DP71; Olympus, Tokyo, Japan), and
analyzed using RT-PCR and western blotting. GFP expression was
considered as an indicator of efficiency of gene delivery. K562/ADM
cells were divided into 4 groups: group 1 (K562/ADM, untransfected
cells); group 2 (K562/ADM/shControl, pGPH1-GFP-Neo-NC-transfected
cells); group 3 (K562/ADM/shApollon,
pGPH1-GFP-Neo-Apollon-transfected cells); and group 4
(K562/ADM/shMDR1, pGPH1-GFP-Neo-MDR1 transfected cells).
Cytotoxicity assays
Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Shanghai, China) was used to determine the
survival rate of all the cell groups incubated with ADM. Firstly,
the cells were seeded in a 96-well plate at a density of
5×103 cells/well in RPMI-1640 medium containing 10% FBS.
Then, various concentrations of ADM at 4–64 mg/l were added to each
well, accordingly. After the cells were incubated at 37°C in 5%
CO2 for 24 h, 10 µl CCK-8 solution was added to each
well and incubation followed for 4 h. Subsequently, the absorbance
was assessed at 490 nm with a fluorescence spectrofluorometer
(F-7000; Hitachi High-Technologies Corporation, Tokyo, Japan). A
blank well containing only medium and drugs, was used as a control.
The 50% inhibition of cell growth (IC50) produced by ADM
was calculated using the untreated cells as the 100% viable
control. The reversal fold (RF) values, as potency of reversal,
were obtained from the following formula: RF = IC50 of
ADM in untransfected cells/IC50 of ADM in the
differently transfected cell groups.
Cellular uptake of ADM
All cell groups were plated in 6-well plates at a
concentration of 1×106 cells in 1 ml of growth medium.
After incubation at 37̊C for 24 h, 2 mg/l of ADM was added for
another 1 h of incubation at 37°C. Cells without ADM were used to
evaluate cell auto-fluorescence. Then, the cells were harvested by
centrifugation and washed twice with ice-cold phosphate-buffered
saline (PBS). The cell-associated mean fluorescence intensity (MFI)
of ADM was detected by flow cytometer using a FACSCalibur (Beckman
Coulter, Brea, CA, USA) with excitation/emission wavelengths at
485/580 nm.
Western blotting
The cells were harvested and washed with PBS. Lysis
buffer (100 µl) was added and the protein concentration of the
lysate was determined using a Bicinchoninic Acid Protein Assay kit
(both from Beyotime Biotechnology, Shanghai, China). The lysed
samples containing 50 µg of protein were separated by 6–8% sodium
dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE;
Beyotime Biotechnology) with a constant voltage of 80 V for 0.5 h
and 120 V for another 1.5 h. The resolved proteins were
electrophoretically transferred to polyvinylidine difluoride
membranes (EMD Millipore, Bedford, MA, USA) and blocked with 5%
skimmed milk for 2 h. Subsequently, the membranes were incubated
overnight at 4°C with specific primary rabbit polyclonal antibodies
against Apollon (1:1,000), P-gp (1:500) and GADPH (1:1,000). The
following day, the membranes were incubated in horseradish
peroxidase-labeled goat anti-rabbit immunoglobulin G (1:5,000) for
2 h at room temperature. Finally, images were captured using a
FluorChem FC2 Gel Imaging System (Alpha Innotech, San Leandro, CA,
USA). The intensity of each band was normalized to that of GADPH
for their respective lanes.
Statistical analysis
Statistical analyses were performed using SPSS 17. 0
software (IBM SPSS, Armonk, NY, USA). Data are expressed as the
means ± SD. Statistical comparisons were evaluated by one-way
ANOVA. Values of P<0.05 were considered to indicate a
statistically significant result.
Results
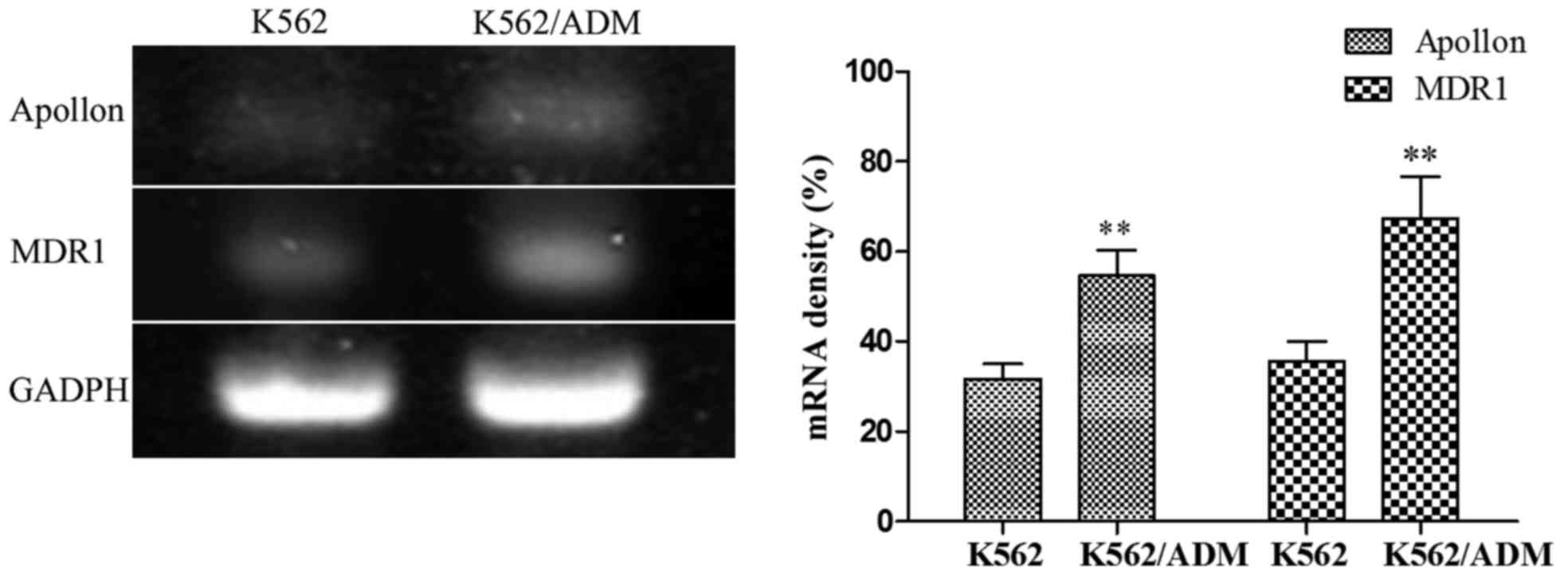
K562/ADM cells overexpress Apollon and
MDR1 in comparison with the parental sensitive cells
RT-PCR analysis was performed to determine the
expression of Apollon and MDR1 in parental K562 cells, and the
resistant K562/ADM cells. As shown in Fig. 1, our results demonstrated that both
the K562 and K562/ADM cells exhibited high expression levels of
Apollon and MDR1, but that the K562/ADM cells expressed much higher
levels of Apollon and MDR1 than the K562 cells (P<0.05).
Overall, our results revealed that in contrast to parental
sensitive K562 cells, MDR K562/ADM cells exhibited a higher level
of Apollon/MDR1 co-expression, which may give rise to MDR.
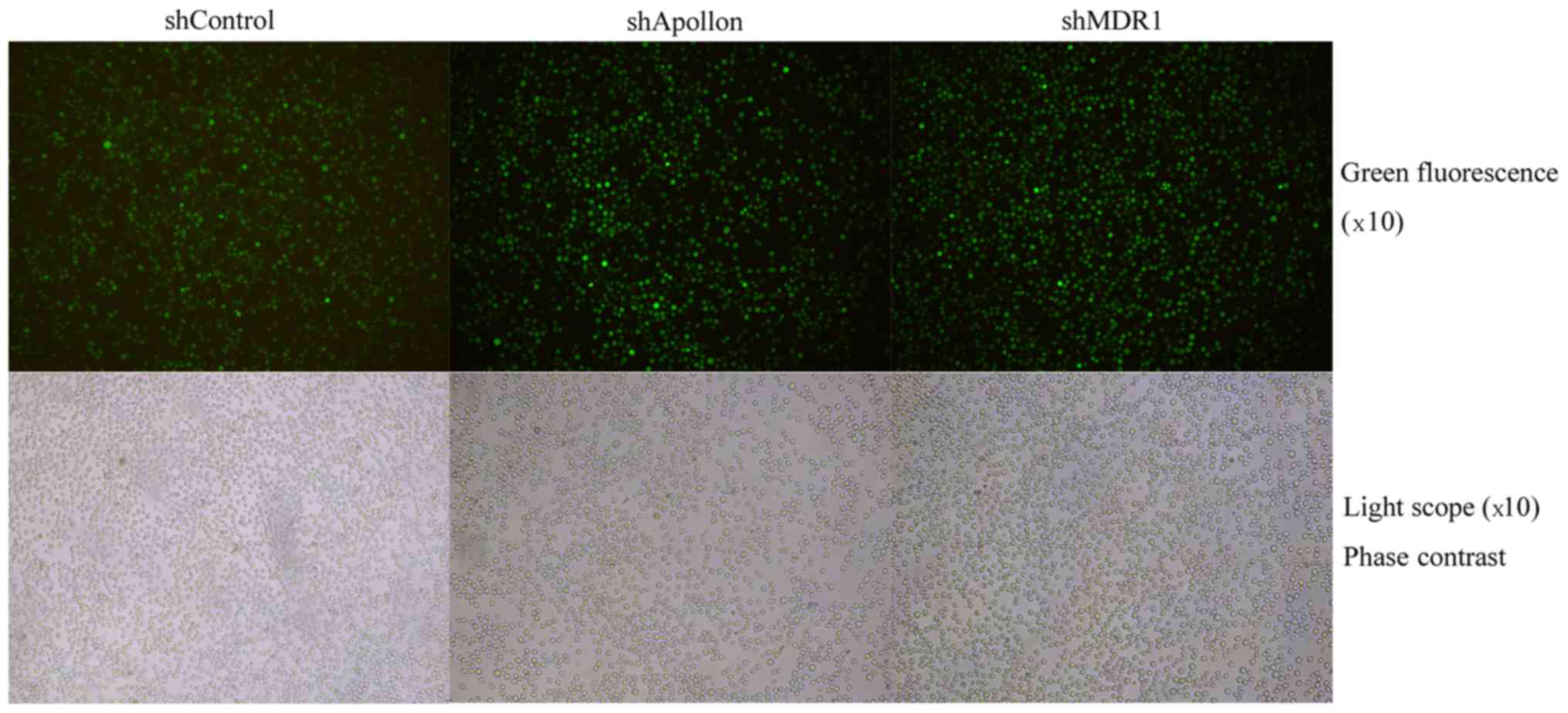
Transfection efficiency of the
recombinant plasmids in the K562/ADM cells
Fluorescence was detected using a fluorescence
microscope after G418 selection. The expression of green
fluorescent protein was detected in most of the stably-transfected
cells (>90%), indicating a very high transfection efficiency
(Fig. 2). These results indicated
that the K562/ADM cells were successfully transfected with the
plasmids.
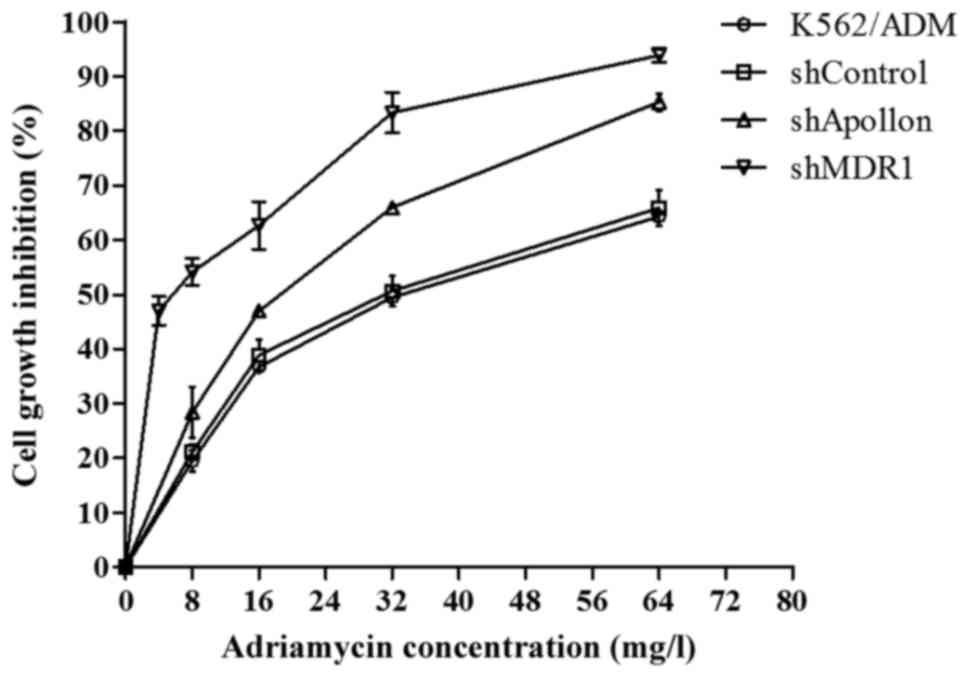
Apollon and MDR1 knockdown increases
the chemosensitivity of K562/ADM cells to ADM
We examined whether Apollon or MDR1 knockdown
sensitizes CML K562/ADM cells to ADM. The CCK-8 assay detected the
growth inhibition rate of cells after exposure to a range of
concentrations of ADM for 24 h. As shown in Fig. 3, the K562/ADM/shApollon and
K562/ADM/shMDR1 cells exhibited significantly greater growth
inhibition than the K562/ADM/shControl and the untransfected
K562/ADM cells by ADM. Then, the IC50 values of ADM were
calculated by CCK-8 assay data to determine whether downregulation
of Apollon and MDR1 contributed to drug sensitivity in the K562/ADM
cells. The IC50 values of ADM in the K562/ADM cells were
32.60±3.215, 30.18±4.03, 17.53±1.595 and 5.785±1.8895 mg/l for the
ADM alone, shControl, shApollon and shMDR1 groups, respectively.
The reversal fold-change of shApollon and shMDR1 was 1.86 and 5.64,
respectively (Table II;
P<0.05). Therefore, the downregulation of Apollon and MDR1
increased the sensitivity of K562/ADM cells to ADM.
| Table II.Effect of Apollon and MDR1 knockdown
on the sensitivity of K562/ADM cells towards ADM by CCK-8
assay. |
Table II.
Effect of Apollon and MDR1 knockdown
on the sensitivity of K562/ADM cells towards ADM by CCK-8
assay.
| Treatment | IC50
(µg/ml) | RF |
|---|
| ADM alone | 32.60±3.215 |
|
| shControl | 30.18±4.03 | 1.08 |
| shApollon |
17.53±1.595a | 1.86 |
| shMDR1 |
5.785±1.8895a | 5.64 |
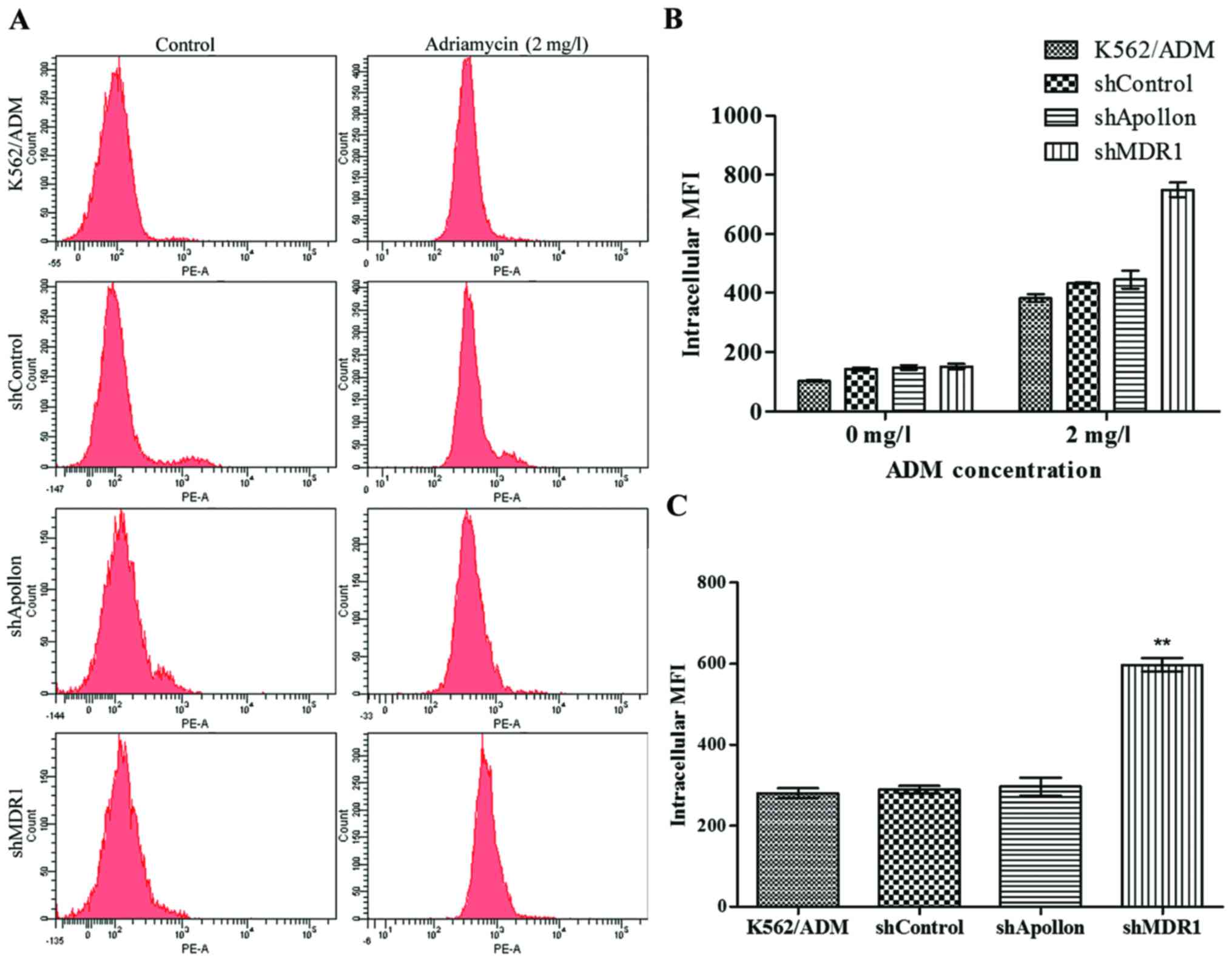
Effect of Apollon and MDR1 knockdown
on the intracellular accumulation of ADM
The intracellular accumulation of ADM was previously
found to decrease significantly in K562/ADM cells compared to
parental K562 cells (20). Thus, we
examined whether knockdown of Apollon and MDR1 increases the
intracellular accumulation of ADM in K562/ADM cells. As shown in
Fig. 4, our results demonstrated
that the K562/ADM/shMDR1 cells had an increased intracellular
accumulation of ADM (P<0.05) while the other cell groups did not
exhibit this change. Thus, our result indicated that shMDR1
significantly inhibited the expression and function of P-gp and
that the knockdown of Apollon did not affect the function of
P-gp.
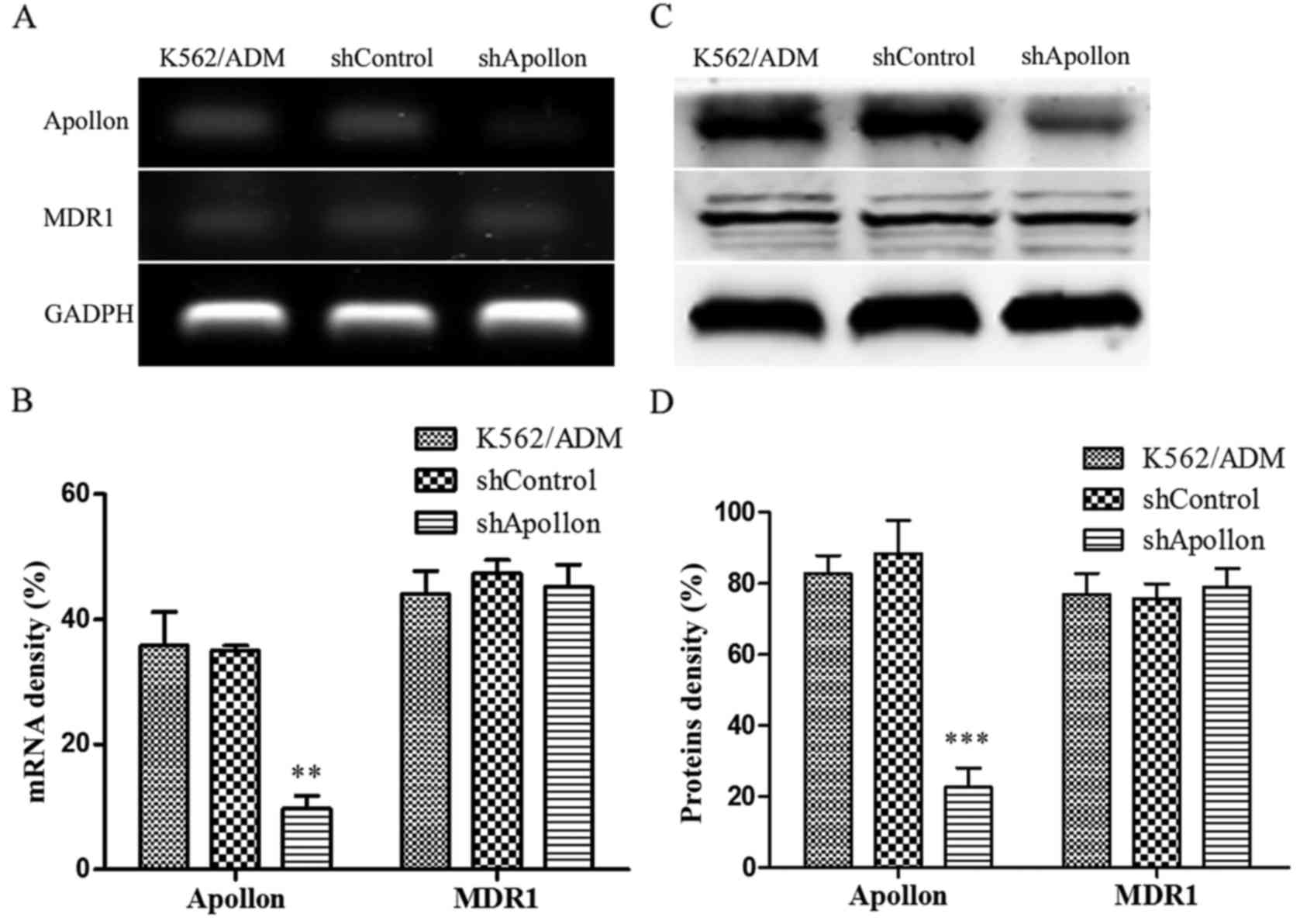
Apollon knockdown shRNA significantly
decreases the expression level of Apollon in K562/ADM/shApollon
cells, but has no impact on MDR1 expression
As Apollon and MDR1 were co-expressed and at high
levels in the CML K562/ADM cells, we explored whether knockdown of
Apollon affects MDR1 expression. After stable transfection with
plasmids shControl and shApollon, we used RT-PCR and western blot
assays to confirm the knockdown efficiency on Apollon mRNA and
protein levels and the expression change in MDR1. As shown in
Fig. 5, compared with the
untransfected K562/ADM cells and the K562/ADM/shControl cells, the
levels of Apollon mRNA and protein in the K562/ADM/shApollon cells
were significantly decreased (P<0.05). However, the levels of
MDR1 mRNA and protein in the K562/ADM/shApollon cells were not
altered.
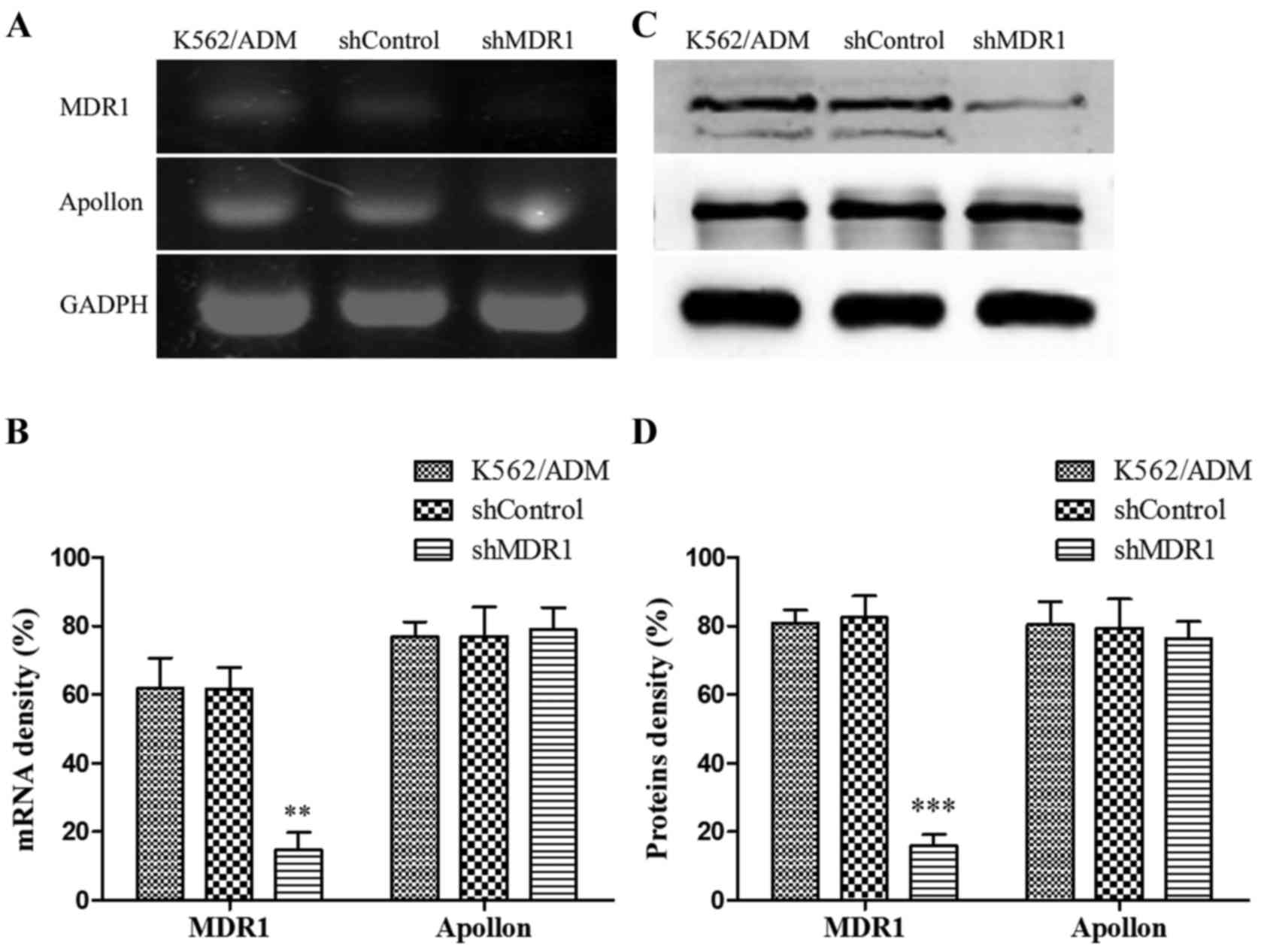
MDR1 knockdown shRNA efficiently
downregulates the expression of MDR1 in K562/ADM/shMDR1 cells but
has no affect on the expression of Apollon
Likewise, to explore whether knockdown of MDR1
affects the expression of Apollon, after stable transfection with
the plasmids shControl and shMDR1, we investigated the expression
changes in MDR1 and Apollon by RT-PCR and western blot assays. As
shown in Fig. 6, the
K562/ADM/shMDR1 cells exhibited significantly decreased mRNA and
protein levels of MDR1 and unchanged expression of Apollon.
Collectively, our experiments revealed that silencing of Apollon or
MDR1 respectively, did not affect the expression of the other, and
that the reversal effect of Apollon and MDR1 knockdown in CML
K562/ADM cells was mainly dependent on the downregulation of the
targeted gene expression.
Discussion
The constitutive tyrosine kinase (TK) activity of
the BCR-ABL oncoprotein in CML plays an important role in promoting
cell proliferation and survival, which provides an understanding of
the pathogenesis of CML and affords a target-oriented therapy.
Although targeted BCR-ABL tyrosine kinase inhibitors (TKIs)
significantly improve the prognosis of CML patients, the
development of MDR becomes the most serious clinical problem
responsible for the failure of chemotherapy. There is a growing
concern in understanding the mechanisms of MDR and finding new
target-oriented therapies to overcome drug resistance.
The cause of MDR is often multifactorial. However,
the most widely studied and important mechanism is the
overexpression of ABC transporters. MDR1, also known as
P-glycoprotein (P-gp) or ABCB1, is the first identified member of
the ABC transporter family, mainly expressed in the plasma membrane
that pumps cytotoxic drugs out of the cells through active
transportation (21). There is an
increasing amount of preclinical data supporting that MDR1 is one
of the most important and representative ABC transporters and plays
a crucial role in MDR of human CML. MDR1 is always overexpressed
and abnormally activated in cancer cells and the expression level
of MDR1 is correlated with MDR (8).
Moreover, it has been reported that the overexpression of MDR1 may
be a risk factor for the treatment and prognosis of CML (22). Furthermore, studies have shown that
the genetic polymorphisms of MDR1 predict the clinical response of
CML patients to TKIs (23–25). Numerous studies revealed that
downregulating the expression of P-gp restored drug sensitivity
(3,6,7,17,19,20).
Thus, MDR1 plays a crucial role in human CML cell resistance to
chemotherapy drugs.
Meanwhile, another important cause for the
acquisition of MDR is apoptosis inhibition. Apoptosis is the
process of cell self-destruction during physiologic or pathologic
conditions under the control of genes, which plays a central role
in the development and homeostasis of multicellular organisms. The
ability to evade apoptosis is considered to be a major cause of
tumor resistance to chemotherapy since chemotherapeutic agents
mainly act depending on the induction of apoptosis (11). The inhibitors of apoptosis proteins
(IAPs) play a strong antagonistic role to cell apoptosis and
potentially contribute to oncogenesis and resistance to antitumor
treatment (6,9). Increased IAP expression has been found
in a variety of human cancers, including CML (4–6,9–12,14–16).
Apollon is the largest and the only membrane-bound member of the
IAP family. Previous studies have reported that overexpression of
the IAP, Apollon, is associated with unfavorable clinical features
and negatively impacts relapse-free survival in childhood acute
leukemia (AL) (26,27). Our previous study demonstrated that
Apollon is constitutively expressed in CML K562 cells and knockdown
of Apollon by short hairpin RNA (shRNA) significantly enhanced K562
cell response to cytotoxic drugs (18).
Since IAPs and ABC drug transporters are the two
most important causes of MDR, the interactions between IAPs and ABC
transporters have been extensively investigated in recent years. It
has been reported that IAPs are always co-expressed with MDR1 at
high levels in a variety of human cancers, and that some members of
the IAP family have a positive correlation with MDR1. The
expression of several IAPs could even be modulated by MDR1
(4–6,15,16).
However, some researchers have reported that the overexpression of
survivin and XIAP in MDR cancer cells is not directly related to
MDR1 (14). Even so, the
relationship between Apollon and MDR1 in MDR cancer cells remains
unclear.
In the present study, we designed and screened shRNA
molecules, respectively targeting Apollon and MDR1 genes to
investigate the role of Apollon and MDR1 in chemotherapy resistance
and their mechanism of interaction in CML K562/ADM cells. Firstly,
we determined that the expression of Apollon and MDR1 in K562/ADM
cells was significantly higher than in the parental K562 cells.
Then, we respectively stably transfected the targeting shRNAs of
Apollon and MDR1 in K562/ADM cells to block gene expression. We
found that both Apollon and MDR1 knockdown significantly restored
the sensitivity of ADM in K562/ADM cells. Then, our data
demonstrated that MDR1 knockdown by shRNA significantly inhibited
the expression and function of P-gp with increased intracellular
accumulation of ADM. However, the intracellular accumulation of ADM
did not change significantly in the Apollon-knockdown K562/ADM
cells, which indicates that the downregulation of Apollon did not
affect the function of P-gp. Additionally, the Apollon-targeting
shRNA transfection significantly decreased the expression of
Apollon without affecting the expression of MDR1. Similarly, we
found that the expression of MDR1 was potently inhibited in
MDR1-targeting shRNA transfected K562/ADM cells which did not
exhibit any effect on Apollon expression. Therefore, it is
reasonable to believe that Apollon and MDR1 play an important role
in the chemotherapy resistance of CML K562/ADM cells although they
have no direct interaction.
It has been revealed that Apollon and MDR1 both
contribute to chemotherapy resistance, but they do not directly
interact with each other. The shRNAs targeting Apollon and MDR1
significantly inhibited the transcription, translation and function
of the targeted genes thereby enhancing the sensitivity of CML
K562/ADM cells to ADM. These studies provide evidence in support of
using shRNAs as a molecularly defined therapeutic approach for MDR
in the treatment of CML and further studies are warranted to
understand the mechanism of MDR in CML cells.
Acknowledgements
The present study was supported by the Shandong
Science and Technology Committee (no. 2010GSF10264), the Foundation
of Shandong Educational Committee (nos. J10LC60 and J11LC01), the
Natural Science Foundation of Shandong Province (no. ZR2014HQ079),
and the Projects of Medical and Health Technology Development
Program in Shandong Province (no. 2014WS0183).
References
|
1
|
Apperley JF: Chronic myeloid leukaemia.
Lancet. 385:1447–1459. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jabbour E: Chronic myeloid leukemia:
First-line drug of choice. Am J Hematol. 91:59–66. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang X, Dong W, Zhou H, Li H, Wang N,
Miao X and Jia L: α-2,8-Sialyltransferase is involved in the
development of multidrug resistance via PI3K/Akt pathway in human
chronic myeloid leukemia. IUBMB Life. 67:77–87. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu M, Liu Y, Deng C, Han R, Jia Y, Liu S,
Jiang Z, Cao X, He L and Zhang Q: Enhanced invasiveness in
multidrug resistant leukemic cells is associated with
overexpression of P-glycoprotein and cellular inhibitor of
apoptosis protein. Leuk Lymphoma. 52:1302–1311. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu F, Liu S, He S, Xie Z, Zu X and Jiang
Y: Survivin transcription is associated with P-glycoprotein/MDR1
overexpression in the multidrug resistance of MCF-7 breast cancer
cells. Oncol Rep. 23:1469–1475. 2010.PubMed/NCBI
|
|
6
|
Notarbartolo M, Cervello M, Dusonchet L,
Cusimano A and D'Alessandro N: Resistance to diverse apoptotic
triggers in multidrug resistant HL60 cells and its possible
relationship to the expression of P-glycoprotein, Fas and of the
novel anti-apoptosis factors IAP (inhibitory of apoptosis
proteins). Cancer Lett. 180:91–101. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kachalaki S, Baradaran B, Majidi J,
Yousefi M, Shanehbandi D, Mohammadinejad S and Mansoori B: Reversal
of chemoresistance with small interference RNA (siRNA) in etoposide
resistant acute myeloid leukemia cells (HL-60). Biomed
Pharmacother. 75:100–104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Arrigoni E, Galimberti S, Petrini M,
Danesi R and Di Paolo A: ATP-binding cassette transmembrane
transporters and their epigenetic control in cancer: An overview.
Expert Opin Drug Metab Toxicol. 12:1419–1432. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Saleem M, Qadir MI, Perveen N and Ahmad B,
Saleem U, Irshad T and Ahmad B: Inhibitors of apoptotic proteins:
New targets for anticancer therapy. Chem Biol Drug Des. 82:243–251.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Notarbartolo M, Cervello M, Poma P,
Dusonchet L, Meli M and D'Alessandro N: Expression of the IAPs in
multidrug resistant tumor cells. Oncol Rep. 11:133–136.
2004.PubMed/NCBI
|
|
11
|
Zhang S, Tang WQ, Weng SQ, Liu XJ, Rao BQ,
Gu JX, Chen S, Wang Q, Shen XZ, Xue RY and Dong L: Apollon
modulates chemosensitivity in human esophageal squamous cell
carcinoma. Oncotarget. 5:(16). 7183–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z, Naito M, Hori S, Mashima T, Yamori
T and Tsuruo T: A human IAP-family gene, Apollon, expressed
in human brain cancer cells. Biochem Biophys Res Commun.
264:847–854. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Peng XX, Tiwari AK, Wu HC and Chen ZS:
Overexpression of P-glycoprotein induces acquired resistance to
imatinib in chronic myelogenous leukemia cells. Chin J Cancer.
31:110–118. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi Z, Liang YJ, Chen ZS, Wang XH, Ding Y,
Chen LM and Fu LW: Overexpression of Survivin and XIAP in MDR
cancer cells unrelated to P-glycoprotein. Oncol Rep. 17:969–976.
2007.PubMed/NCBI
|
|
15
|
Reis FR, Vasconcelos FC, Pereira DL,
Moellman-Coelho A, Silva KL and Maia RC: Survivin and
P-glycoprotein are associated and highly expressed in late phase
chronic myeloid leukemia. Oncol Rep. 26:471–478. 2011.PubMed/NCBI
|
|
16
|
Silva KL, de Souza PS, de Nestal Moraes G,
Moellmann-Coelho A, Vasconcelos FC and Maia RC: XIAP and
P-glycoprotein co-expression is related to imatinib resistance in
chronic myeloid leukemia cells. Leuk Res. 37:1350–1358. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen JR, Jia XH, Wang H, Yi YJ, Wang JY
and Li YJ: Timosaponin A-III reverses multi-drug resistance in
human chronic myelogenous leukemia K562/ADM cells via
downregulation of MDR1 and MRP1 expression by inhibiting PI3K/Akt
signaling pathway. Int J Oncol. 48:2063–2070. 2016.PubMed/NCBI
|
|
18
|
Jia XH, Xiao FF and Li JC: Effect of
Apollon siRNA combined with tetramethylpyrazine on proliferation
and apoptosis of leukemia K562 cells. Zhongguo Dang Dai Er Ke Za
Zhi. 16:135–140. 2014.(In Chinese). PubMed/NCBI
|
|
19
|
Du WT, Liu B, Gu DS, Han ZB, Liu PX, Xu J,
Liang L, Zhao HF, Lu SH and Yang RC: Reversal of leukemia multidrug
resistance by sequence-specific short hairpin RNA. Zhongguo Shi Yan
Xue Ye Xue Za Zhi. 17:563–567. 2009.(In Chinese). PubMed/NCBI
|
|
20
|
Wang L, Meng Q, Wang C, Liu Q, Peng J, Huo
X, Sun H, Ma X and Liu K: Dioscin restores the activity of the
anticancer agent adriamycin in multidrug-resistant human leukemia
K562/adriamycin cells by down-regulating MDR1 via a mechanism
involving NF-κB signaling inhibition. J Nat Prod. 76:909–914. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sarkadi B, Homolya L, Szakács G and Váradi
A: Human multidrug resistance ABCB and ABCG transporters:
Participation in a chemoimmunity defense system. Physiol Rev.
86:1179–1236. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Agrawal M, Hanfstein B, Erben P, Wolf D,
Ernst T, Fabarius A, Saussele S, Purkayastha D, Woodman RC, Hofmann
WK, et al: MDR1 expression predicts outcome of Ph+
chronic phase CML patients on second-line nilotinib therapy after
imatinib failure. Leukemia. 28:1478–1485. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zu B, Li Y, Wang X, He D, Huang Z and Feng
W: MDR1 gene polymorphisms and imatinib response in chronic
myeloid leukemia: A meta-analysis. Pharmacogenomics. 15:667–677.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zheng Q, Wu H, Yu Q, Kim DH, Lipton JH,
Angelini S, Soverini S, Vivona D, Takahashi N and Cao J: ABCB1
polymorphisms predict imatinib response in chronic myeloid leukemia
patients: A systematic review and meta-analysis. Pharmacogenomics
J. 15:127–134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lardo M, Castro M, Moiraghi B, Rojas F,
Borda N, Rey JA and Lazarowski A: MDR1/ABCB1 gene
polymorphisms in patients with chronic myeloid leukemia. Blood Res.
50:154–159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sung KW, Choi J, Hwang YK, Lee SJ, Kim HJ,
Lee SH, Yoo KH, Jung HL and Koo HH: Overexpression of Apollon, an
antiapoptotic protein, is associated with poor prognosis in
childhood de novo acute myeloid leukemia. Clin Cancer Res.
13:5109–5114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ismail EA, Mahmoud HM, Tawfik LM, Habashy
DM, Adly AA, El-Sherif NH and Abdelwahab MA: BIRC6/Apollon gene
expression in childhood acute leukemia: Impact on therapeutic
response and prognosis. Eur J Haematol. 88:118–127. 2012.
View Article : Google Scholar : PubMed/NCBI
|