Introduction
Ovarian cancer is the most frequent cause of
cancer-related death among all gynecological cancers. Approximately
70% of patients with ovarian cancer are diagnosed at an advanced
stage (1). The degree of peritoneal
dissemination and chemotherapy-resistant tumors is related to the
prognosis of patients with advanced-stage ovarian cancer. The
molecular mechanisms that allow ovarian cancer cells to detach from
the primary tumor, invade the peritoneal surface and regrow at this
site are not yet well understood. Thus, obtaining a better
understanding of the molecular events that contribute to tumor
invasion and metastasis is crucial for developing novel treatment
strategies for ovarian cancer.
The epithelial-mesenchymal-transition (EMT) is a
developmental process in which polarized epithelial cells undergo
multiple biochemical changes and assume a mesenchymal phenotype,
with increased migratory capacity, invasiveness, resistance to
apoptosis and the expression of extracellular matrix components
(2–6). The EMT is regulated through the
downregulation of the epithelial marker E-cadherin by
transcriptional repressors such as Snail and Zeb (5). Importantly, the EMT is involved in
cell invasion, resistance to chemotherapy, and the formation of
side populations of cancer stem-like cells (CSCs) (7). Such CSCs or tumor initiating cells
comprise a sub-population of cancer cells within each tumor that
have tumorigenic potential in vivo and which are able to
recreate the full heterogeneity of the parental tumor (8). However, the molecular mechanism
underlying the regeneration of CSCs through EMT has not yet been
clarified.
Cluster of differentiation (CD) 24 is a small,
heavily glycosylated mucin-like cell surface protein (27–30 amino
acids in length), which binds to the membrane via a
glycosyl-phosphatidylinositol anchor (9). Under physiological conditions, CD24
was initially identified as a B cell marker (10). Later, it was found to be expressed
not only in developing or regenerating tissue, but also in
granulocytes, Pre-B cells, keratinocytes and renal tubules
(11). CD24 has also been reported
to be a ligand for P-selectin, an adhesion receptor on activated
endothelial cells and platelets (12,13).
These findings suggest that the molecule functionally enhances the
metastatic potential of cancer cells. Various studies have
identified CD24 as a marker of a poor prognosis in patients with
hematological malignancies (14),
and various solid tumors, including glioma (15), laryngeal squamous cell carcinoma
(16), non-small cell lung cancer
(17), breast cancer (18), renal cell carcinoma (19), colorectal cancer (20), bladder carcinoma (21) and epithelial ovarian cancer
(22). However, the underlying
mechanism has remained elusive. Recent findings have described CD24
as a marker for tumor stem cells in pancreatic and ovarian
carcinomas (23,24).
In this study, we demonstrated that CD24 was
markedly associated with a more aggressive disease course. The data
on CD24 demonstrated that CD24-positive ovarian cancer cells are
susceptible to the induction of EMT, and this study provides new
knowledge on the function of CD24 in tumor metastasis and its
suitability as a therapeutic target in patients with advanced
ovarian cancer.
Materials and methods
Tissue samples
Tissue samples were obtained from 174 Japanese
patients who underwent surgical resection for primary ovarian
carcinomas at Osaka Medical College between January 2001 and
December 2009. All patients underwent a primary staging operation,
such as total abdominal hysterectomy, bilateral
salpingo-oophorectomy, omentectomy, and/or lymphadenectomy.
Furthermore, all patients received 6-course conventional TC
(paclitaxel and carboplatin) therapies as the adjuvant
chemotherapy. The median follow-up duration was 29.2 months (range,
2–122 months). The institutional review board approved this study
(no. 0156) and informed consent was obtained from all of the
patients. We performed a tissue microarray of the assembly of 5-mm
cores retrieved from microscopically selected representative
tissues of the primary tumors. The specimens were fixed in 10%
formalin and embedded in paraffin. Serial sections cut out from
paraffin-embedded blocks were used for routine histopathology. A
4-µm section was cut from a tissue microarray block and was
immunohistochemically analyzed to detect the expression of
CD24.
Immunohistochemistry
Tumor samples were formalin-fixed and embedded in
paraffin. Deparaffinized and rehydrated sections (4-µm) were
autoclaved in 0.01 mol/l citrate buffer (pH 6.0) for 15 min at
121°C for antigen retrieval. Endogenous peroxidase activity was
blocked with 0.3% solution hydrogen peroxidase in methanol for 30
min. Tumor sections were incubated at 4°C for 12 h with a
CD24-specific antibody (1:50 dilution; Thermo Fisher Scientific,
Yokohama, Japan). The sections were washed with 1X
phosphate-buffered saline (PBS) and incubated with Histofine Simple
Stain MAX PO (Multi; Nichirei) for 30 min at room temperature.
Finally, the sections were washed with 1X PBS and the signals were
visualized by incubation with
H2O2/diaminobenzidine substrate solution for
5 min. The sections were counterstained with hematoxylin prior to
dehydration and mounting. The evaluation of the immunohistochemical
data was performed by two independent pathologists who were blinded
to the clinicopathological data. For the immunohistochemistry
score, the expression levels of CD24 were assessed using a
semiquantitative system based on the staining intensity (0,
negative; 1+, weak; 2+, moderate; 3+, strong) and the percentage of
cells positively stained cells (0–100%), with the positive
expression of CD24 defined as ≥50% of tumor cells having a
membranous or cytoplasmic staining intensity of ≥2+, as described
by Kim et al (25). Scoring
was performed three times per slide for three distinct fields, and
the three scores were averaged.
Reagents/antibodies
Cisplatin was also purchased from Sigma-Aldrich (St.
Louis, MO, USA). The anti-phospho-Akt (ser473), anti-Akt
antibodies, anti-phospho-ERK (Thr202/Tyr204), anti-ERK antibodies,
anti-phospho-NF-κB (Ser536), and anti-NF-κB antibodies purchased
from Cell Signaling Technology (Beverly, MA, USA).
Cell culture
One human ovarian mucinous adenocarcinoma cancer
cell line, Caov-3, which was obtained from the American Type
Culture Collection (Rockville, MD, USA), was grown in phenol DMEM
containing 10% dextran-coated, charcoal-treated fetal calf serum in
a humidified atmosphere of 5% CO2 with 95% air at
37°C.
The expression plasmids and cDNA
transfection
We purchased pEZ-M02-CD24 (Gene Copoeia, Rockville,
MD, USA) which used Homo sapiens CD24 signal transducer
mRNA. For the transfection of each sample, oligomer-Lipofectamine
plus complexes were prepared as follows: 4.0 µg of cDNA oligomer
were diluted in 250 µl of Opti-MEM (Invitrogen, CA, USA). The
oligomer-Lipofectamine plus complexes were added to each of the
wells, which contained cells and medium; gentle mixing was then
performed by rocking the plate back and forth. The cells were
incubated at 37°C in a CO2 incubator for 18 h. The cells
were then prepared for each assay.
CD24 mRNA depletion by the plasmid
transfection of siRNA
We purchased the Trilencer-27 siRNA plasmid for
human CD24 (OriGene, Rockville, MD, USA), as well as scrambled
sequence negative control plasmid (OriGene). Caov-3 cells were
transfected using the CD24 siRNA Lipofectin method, according to
the manufacturer's protocol, which was the same method that was
used for the expression of CD24.
The reverse transcription polymerase
chain reaction (RT-PCR)
Caov-3 cells were treated with PBS, TGF-β for 24 h.
Total RNA was isolated from cells using an RNeasy Protect Mini kit
(Qiagen) according to the manufacturer's protocol. The Super Script
II Reverse Transcriptase kit (Invitrogen) was used to synthesize
cDNA. Polymerase chain reactions were performed with a Platinum PCR
SuperMix (Invitrogen) system according to the manufacturer's
protocol. The following primers were used for the PCR: CD24
forward, 5′-ACCCACGCAGATTTATTCCA-3′; reverse,
5′-ACCACGAAGAGACTGGCTGT-3′. Snail forward,
5′-GCCTTCAACTGCAAATACTGC-3′; reverse, 5′-CTTCTTGACATCTGAGTGGGTC-3′.
Ecad forward, 5′-GGTGGGTGACTACAAAATCAATCTC-3′; reverse,
5′-GGGGCAGTAAGGGCTCTTT-3′. β-actin forward,
5′-GAATTCATGTTTGAGACCTTCAA-3′; reverse,
5′-CCGGATCCATCTCTTGCTCGAAGTCCA-3′. The expression levels of β-actin
were assessed as an internal control in all reactions.
Flow cytometry
The cultures from the Caov-3 cell lines were washed
with PBS. For some experiments, single cells dissociated from tumor
spheres were analyzed by the following method. One million
trysinized cells were incubated with 7-amino-actinomycin D (BD
Biosciences, NJ, USA) only or 7-amino-actinomycin D and a mouse
anti-human monoclonal antibody of CD24-FITC (BD Biosciences) with
stain buffer (BD Pharmingen™) for 20 min at room temperature in the
dark. After washing, the cells were analyzed using a BD FACS Aria™.
The fluorescence intensity was analyzed using the BD FACS Diva
software program (BD Biosciences).
Magnetic activated cell sorting
(MACS)
The CD24-positive/CD24-negative Caov-3 cells were
obtained using the MACS kit according to the manufacturer's
instructions (Miltenyi Biotec, Nordrhein-Westfalen, Germany).
Briefly, total populations of adherent cells were enzymatically
dissociated into a single cell suspension and counted to confirm
the quantity of the whole cells. The cells were incubated with 40
µl buffer per 107 total cells, which were directly
conjugated to 10 µl of CD24 Biotin at 4°C for 15 min. After
washing, the cells were incubated with 80 µl buffer directly
conjugated to 20 µl of Anti-Biotin MicroBeads at 4°C for 15 min.
Subsequently, the suspended cells were added to a MACS column that
was placed in the magnetic field of a MACS separator (Miltenyi
Biotech). The labeled CD24-positive cells were retained on the
column and the unlabeled cells were eluted; when the column was
removed from the magnetic field, the magnetically retained
CD24-positive cells were collected as positively selected cells for
further investigation. We also confirmed the expression of CD24
mRNA in CD24-positive and CD24-negative Caov-3 cells using
conventional PCR. All experiments were performed three times.
The soft agar assay
The Caov-3 cells sorted CD24-positive/CD24-negative
by MACS. A 6-well culture plate was coated with 2 ml of bottom agar
mixture (DMEM/F12 with 10% FBS, 0.6% agar). After the bottom layer
solidified, 2 ml top agar-medium mixture (DMEM/F12 with 10% FBS,
0.3% agar) containing 2×104 CD24-positive or
CD24-negative cells was added, and the plate was incubated at 37°C
for 4 weeks. The medium was replenished every 3 days. The plate was
stained with 0.5% crystal violet, and the colonies with a diameter
of >40 µm were counted.
The cell proliferation assay
CD24-positive/CD24-negative Caov-3 cells were
obtained using a MACS kit. CD24-positive and CD24-negative cells
(1×105 cells/well) were seeded in triplicate in a 6-well
plate with serum-free DMEM to achieve cellular starvation. After 24
h, the medium was replaced with 100 ml complete growth medium and
incubated for 1 more week. The cell numbers were counted manually
with an optical microscope after 1, 3, 5 and 7 days. This
experiment was carried out in quadruplicate.
The cell invasion assays
Cell invasion was assessed using 24-well-Transwell
chambers (Corning, NY, USA) with a pore size of 8 µm (Kurabo,
Osaka, Japan). Transwell upper chambers were pre-coated with 5
µg/chamber of solubilized basement membrane (Matrigel; BD
Biosciences). Forty percent fibronectin was added to the bottom
chamber. Ovarian cancer cells (2.5×105 per chamber) were
seeded in the serum-containing media in the upper well of the
Transwell chambers and incubated for 16 h at 37°C in 5%
CO2. The non-invasive cells in the lower chamber (below
the filter surface) were fixed in 100% methanol after 3.7%
formaldehyde, stained with Giemsa staining solution, and counted
under a microscope (×40 magnification). Cells from at least five
fields were counted per chamber.
The chemotherapeutic sensitivity
assay
Cells cultured in 10% growth medium were seeded in
96-well plates at a density of 2×104 cells per well.
After 16 h of starvation, the cells were incubated for 48 h before
being exposed to 0, 10, 50 or 100 µM cisplatin, which was purchased
from Sigma-Aldrich. Then, CellTiter 96 AQueous (MTS) One Solution
reagent (Promega, Tokyo, Japan) was added to each well, and the
absorbance was recorded at 490 nm using a Corona SH-1000 lab
absorbance microplate reader (Corona Electric Co. Inc., Ibaraki,
Japan). The cell numbers were then calculated using a standard
curve of the correlation of absorbance with the cell number counted
under a microscope. All of the experiments were carried out in
quadruplicate, and cell viability was expressed as the ratio of the
number of viable cells with cisplatin treatment to that of
untreated cells.
Western blotting
The cells were serum-starved and stimulated with PBS
or 10 ng/ml TGF-β for 10 min. The cells were then washed three
times in ice-cold PBS and lysed using Pierce RIPA buffer (Thermo
Fisher Scientific) and the cytoplasmic and nuclear fractions were
separated using a Nuclear Extract kit (Active Motif Carlsbad, CA,
USA). To detect all of the proteins, equal amounts of cytoplasmic
proteins were separated by SDS-polyacrylamide gel electrophoresis
and transferred to nitrocellulose membranes. Blocking was performed
in 10% bovine serum albumin in 1X Tris-buffered saline. Western
blotting was performed with the primary antibody of CD24 (1:100
dilution), phosphorylated Akt (1:1,000 dilution), Akt (1:1,000
dilution), phosphorylated ERK (1:1,000 dilution), ERK (1:1,000
dilution), phosphorylated NF-κB (1:1,000 dilution), and NF-κB
(1:1,000 dilution) overnight at 4°C. Finally, the bands were
visualized using enhanced chemiluminescence (ECL Plus, GE
Healthcare Life Sciences, Pittsburgh, PA, USA). All of the western
blots were checked for equal protein loading using Ponceau
staining.
The intraperitoneal xenograft
model
Female 5-week-old athymic nude mice (BALB/c
Slc-nu/nu) were purchased from Japan SLC and maintained in
accordance with the institutional guidelines of Osaka Medical
College. All of the animal studies were carried out according to
approved experimental protocols. To assess tumorigenicity,
80×104 purified CD24-positive and CD24-negative cells
were suspended in 1 ml PBS and were then intraperitoneally injected
into each of the 10 female nude mice (5-week-old). The body weight
and abdominal circumference were measured weekly. On the 50th day
after the initiation of treatment, all of the mice were sacrificed,
and the presence of macroscopic disease and the ascites volumes and
organ weight were examined. Dissemination tumors were resected,
fixed in formalin and paraffin sections were made for hematoxylin
and eosin staining.
Statistical analysis
The statistical analyses in this study were
performed using the JMP software program (SAS Institute, Japan).
Fisher's exact probably test was used to evaluate the correlations
between the immunohistochemical and clinical data. The endpoints
were progression-free survival (defined as the time from the first
day of chemotherapy until disease progression, based on the
findings of imaging studies) and overall survival (defined as the
time from the first day of chemotherapy to death from any cause).
The univariate analyses of the histology, progression-free survival
and overall survival were determined according to the Kaplan-Meier
method using the log-rank test and the Cox proportional hazards
model, respectively. Differences with p-values of <0.05 were
considered to indicate statistical significance.
Results
The expression of CD24 in ovarian
cancer patients
The data that we investigated included the patients'
age, histology, International Federation of Gynecology and
Obstetrics (FIGO) stage, peritoneal cytology, the presence of lymph
node metastasis, the presence of peritoneal metastasis and the
outcome, as shown in Table I. The
174 epithelial ovarian cancers included serous adenocarcinomas
(n=48), mucinous adenocarcinomas (n=21), endometrioid
adenocarcinomas (n=32), clear cell adenocarcinomas (n=32), serous
surface papillary carcinomas (SSPCs; n=26) and others (n=15). The
disease stages of the 174 investigated patients were classified as
follows: stage I, n=56; stage II, n=14; stage III, n=85 and stage
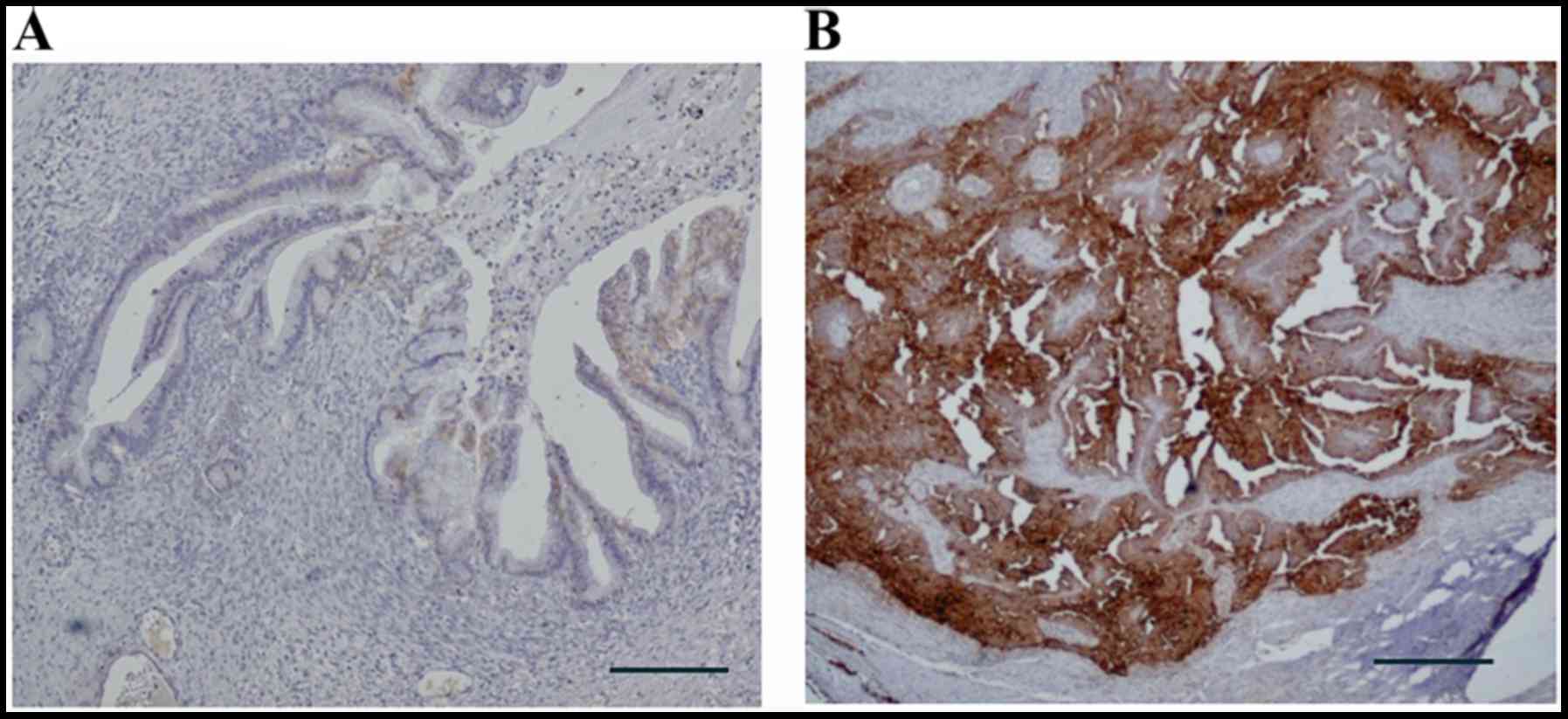
IV, n=19. Representative examples of immunohistochemical staining
with anti-CD24 antibody are shown in Fig. 1. CD24 was expressed in a
significantly higher percentage of patients with serous
adenocarcinoma (87.5%) and SSPCs (84.6%) than among patients with
other histological subtypes (p<0.01). In the analysis of the
FIGO stage, CD24 was expressed in 53.6, 78.6, 77.7 and 79.0% of the
patients with stage I, II, III and IV disease, respectively. CD24
was expressed in 70.8% of the patients with positive peritoneal
cytology and 68.1% of the patients with negative peritoneal
cytology. The expression of CD24 was observed in 78.7% of the
patients with peritoneal metastasis, in comparison to 60.0% of the
patients without peritoneal metastasis. The expression of CD24 was
observed in 87.9% of the patients with lymph node metastasis, in
comparison to 60.7% of the patients without lymph node metastasis.
Significant associations were observed between the CD24 expression
rate and the FIGO stage (p<0.01), and the presence of peritoneal
(p<0.01) and lymph node metastasis (p<0.01).
 | Table I.The immunohistochemistry results. |
Table I.
The immunohistochemistry results.
|
| CD24 |
|
|---|
|
|
|
|
|---|
| Variables | Negative | Positive | P-value |
|---|
| Age, years |
|
|
0.18 |
|
<50 | 18 (37.5) | 30 (62.5) |
|
|
≥50 | 34 (27.0) | 92 (73.0) |
|
| Histology |
|
| <0.01 |
| Serous
adenocarcinoma | 6
(12.5) | 42 (87.5) |
|
| Mucinous
adenocarcinoma | 13 (61.9) | 8
(38.1) |
|
| Clear cell
adenocarcinoma | 12 (37.5) | 20 (62.5) |
|
| Endometrioid
adenocarcinoma | 13 (40.6) | 19 (59.4) |
|
| SSPCs | 4
(15.4) | 22 (84.6) |
|
| Others | 4
(26.6) | 11 (73.4) |
|
| Stage |
|
| <0.001 |
| I | 26 (46.4) | 30 (53.6) |
|
| II | 3
(21.4) | 11 (78.6) |
|
|
III | 19 (22.3) | 66 (77.7) |
|
| IV | 4
(21.0) | 15 (79.0) |
|
| Peritoneal
cytology |
|
|
0.75 |
|
Positive | 38 (29.2) | 92 (70.8) |
|
|
Negative | 14 (31.8) | 30 (68.1) |
|
| Dissemination |
|
| <0.01 |
|
Positive | 20 (21.3) | 74 (78.7) |
|
|
Negative | 32 (40.0) | 48 (60.0) |
|
| Lymph node
metastasis |
|
| <0.01 |
|
Positive | 4
(12.1) | 29 (87.9) |
|
|
Negative | 35 (39.3) | 54 (60.7) |
|
| Recurrence |
|
| <0.01 |
|
Positive | 18 (19.1) | 76 (80.9) |
|
|
Negative | 34 (42.5) | 46 (57.5) |
|
| Died or
survived |
|
|
0.02 |
|
Died | 11 (18.6) | 48 (81.4) |
|
|
Survived | 41 (35.7) | 74 (64.3) |
|
The prognostic impact of the CD24
expression status in patients with ovarian carcinoma
The CD24 expression status was investigated in
relation to patient survival. All patients underwent a primary
staging operation such as total abdominal hysterectomy, bilateral
salpingo-oophorectomy, omentectomy, and/or lymphadenectomy.
Furthermore, all patients received 6-course conventional TC
(paclitaxel and carboplatin) therapies as the adjuvant
chemotherapy. The median follow-up duration was 29.2 months (range,
2–122 months). During the follow-up period, 94 of 174 patients
(54%) had recurrent disease (18 from the CD24-negative group and 76
from the CD24-positive group), and 59 of 174 patients (33.9%) died
(11 from the CD24-negative group and 48 from the CD24-positive
group), as shown in Table I. As
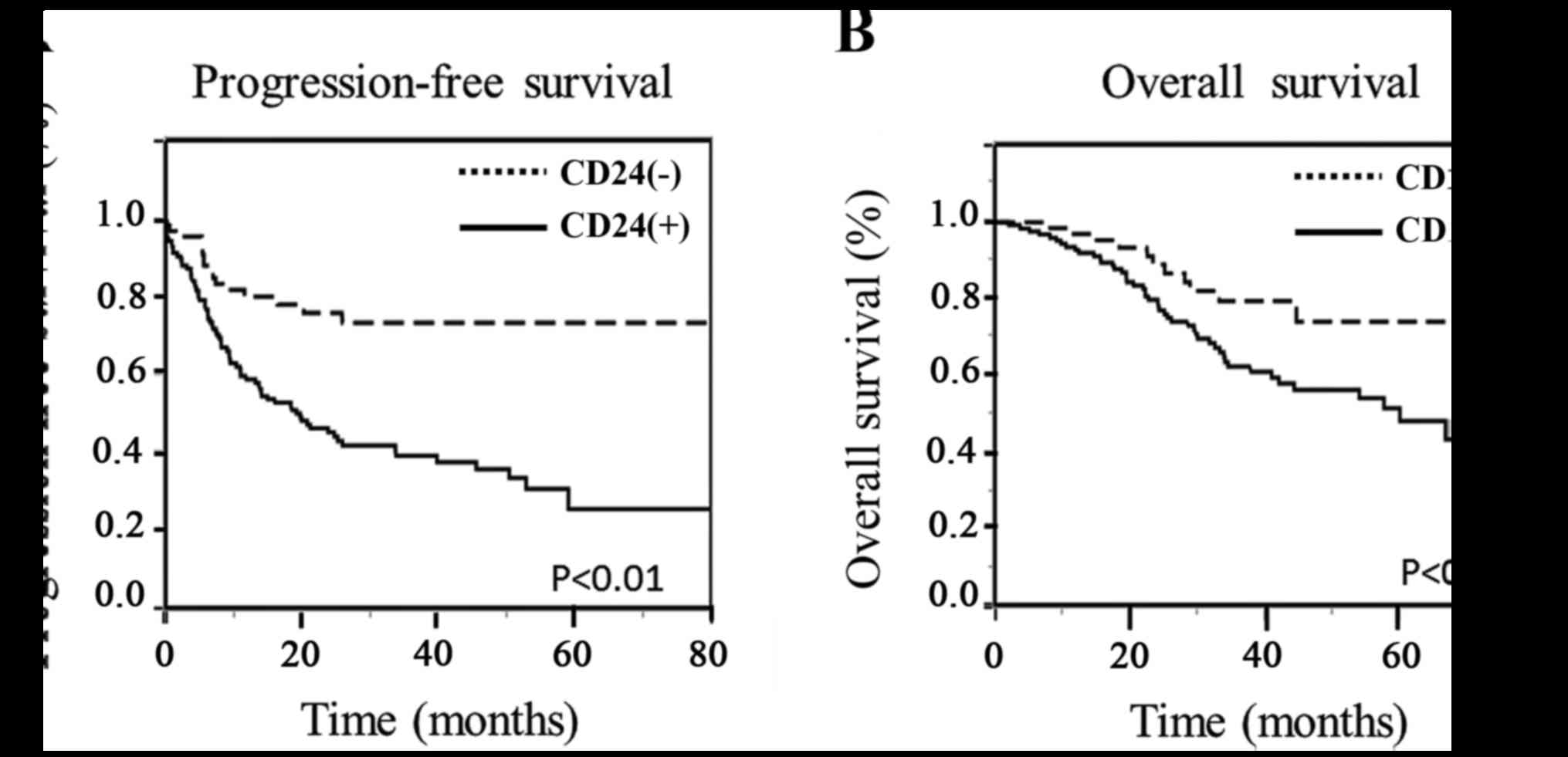
shown in Fig. 2, progression-free
and overall survival were stratified according to the CD24
expression status using the Kaplan-Meier method with the log-rank
test. The CD24 expression status was found to be significantly
associated with both the progression-free survival (p<0.01) and
overall survival (p<0.05).
TGF-β stimulates EMT and CD24
expression
To study the EMT mechanism, we cultured
cisplatin-resistant ovarian cancer cell lines (Caov-3 cells) in
medium containing TGF-β. TGF-β is a major inducer of EMT during
embryonic development, the pathogenesis of fibrotic disorders and
cancer progression (26–28). It was first described as an inducer
of the EMT in normal mammary epithelial cells (29). We examined whether there was a
correlation between the EMT mechanism induced by TGF-β and the
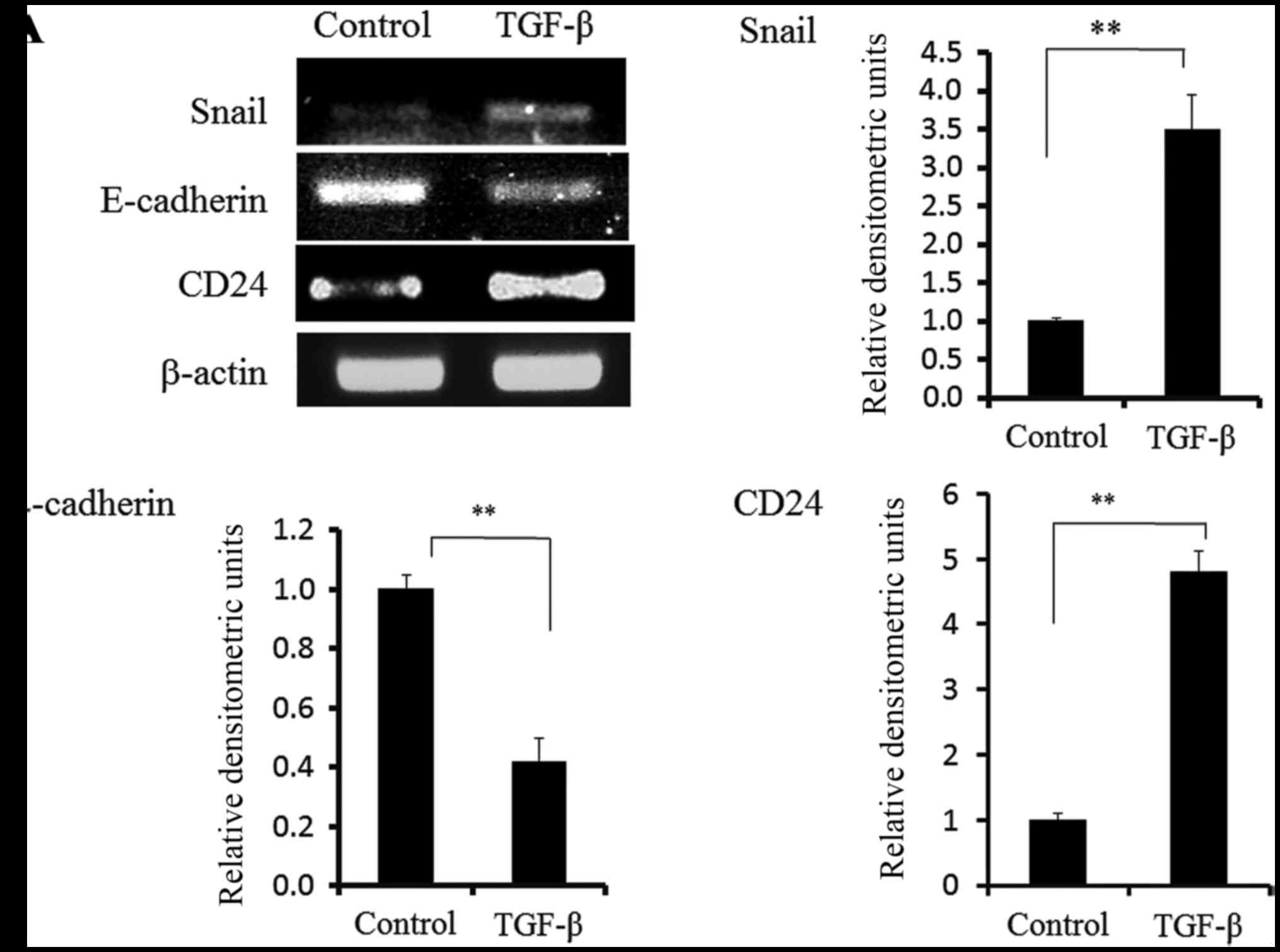
expression of CD24 in Caov-3 cells. The data from a quantitative
PCR indicated that the expression of Snail was increased with TGF-β
in the cultured medium, and that the expression of E-cadherin was
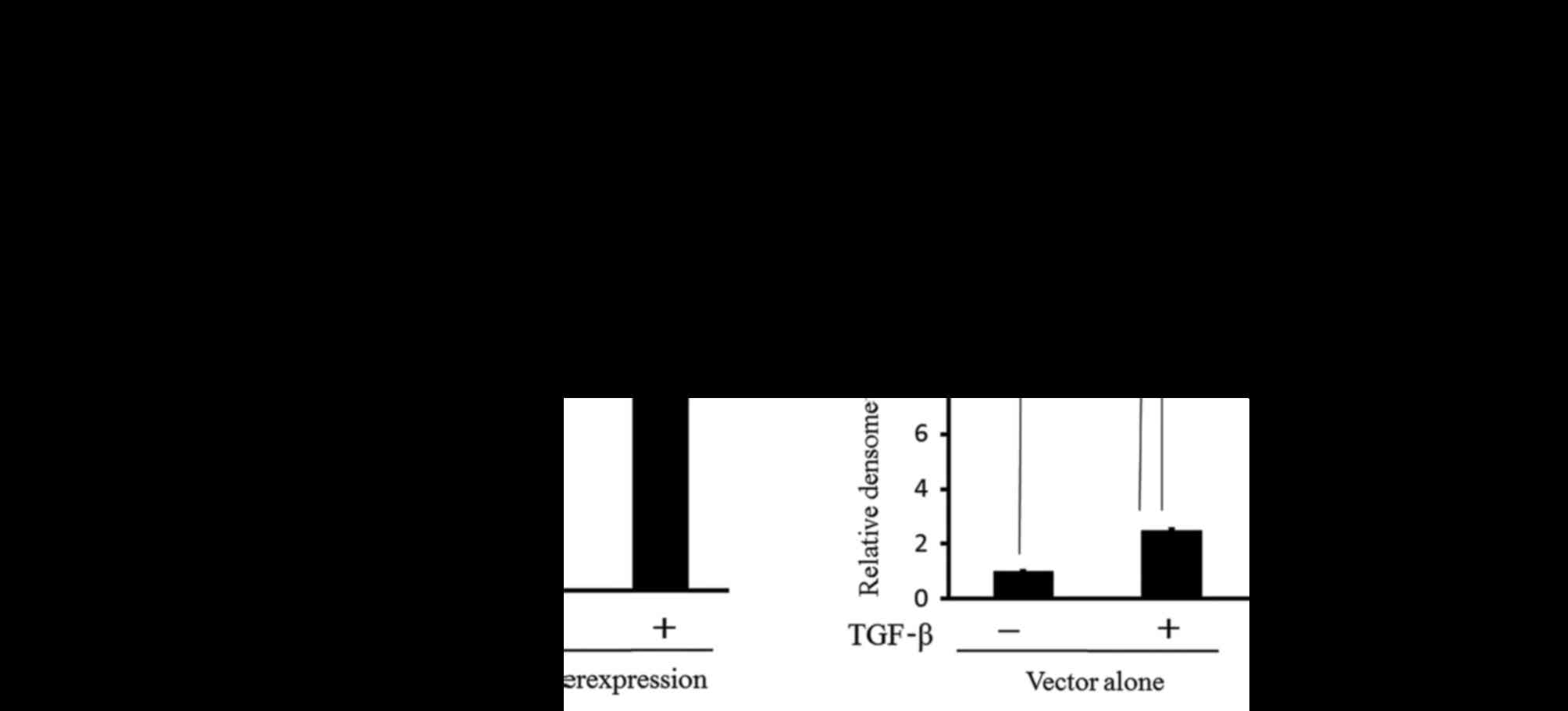
decreased with TGF-β in the cultured medium, as shown in Fig. 3A. The expression of CD24 was also
increased with TGF-β in the cultured medium. These findings suggest
that TGF-β stimulates not only the EMT phenotypes but also the
expression of CD24 in Caov-3 cells.
Hypoxia stimulates EMT and CD24
expression
A hypoxic microenvironment, which is common to
cancer cells emerges as an important factor in the induction of a
pathological EMT, which is a key link in cancer progression
(30). We examined whether there is
a correlation between the EMT mechanism induced by hypoxia and the
expression of CD24 in Caov-3 cells. We cultured Caov-3 cells under
the condition of 1% hypoxia for 12 h. As shown in Fig. 3B, the data from quantitative PCR
indicated that the expression of Snail was increased, and that the
expression of E-cadherin was decreased under 1% hypoxia in
comparison to normoxia. The expression of CD24 was also increased
under 1% hypoxia (Fig. 3B). Next,
we performed flow cytometry to examine whether the CD24 fraction of
the Caov-3 cells was changed under the condition of 1% hypoxia. The
CD24 fraction under normoxia and 1% hypoxia was 52.9 and 72.9%,
respectively (data not shown). These findings suggest that hypoxic
conditions stimulate not only the EMT phenotypes but also the
expression of CD24 in Caov-3 cells.
CD24 induces EMT in ovarian cancer
cells
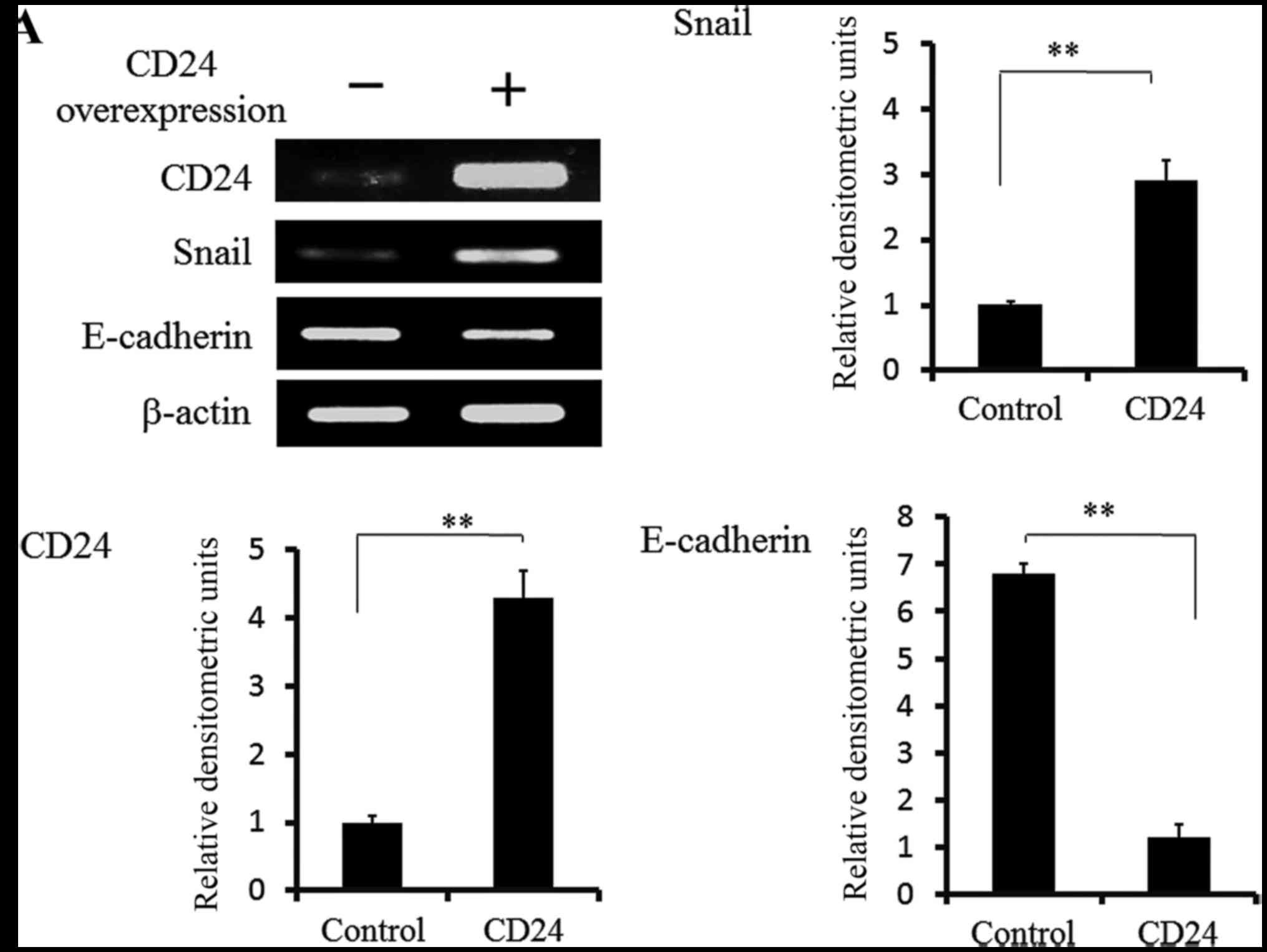
We examined whether cells with the overexpression of
CD24 induced the EMT mechanism in Caov-3 cells. As shown in
Fig. 4A, the data from a
quantitative PCR indicated that the expression of Snail was
increased, and that the expression of E-cadherin was decreased in
the CD24-overexpressing cells in comparison to control cells. We
confirmed that CD24 directly induced the EMT phenotypes in Caov-3
cells and that the CD24-positive Caov-3 cells showed the
acquisition of spindle cell morphology, as shown in Fig. 4B.
CD24-positive cells have enhanced
proliferation, a highly invasive phenotype, colony formation and
are associated with cisplatin resistance in ovarian cancer
cells
Several groups have reported that the EMT
contributes to the properties of cancer stem cells (31,32).
We examined whether CD24 could increase the colony-forming activity
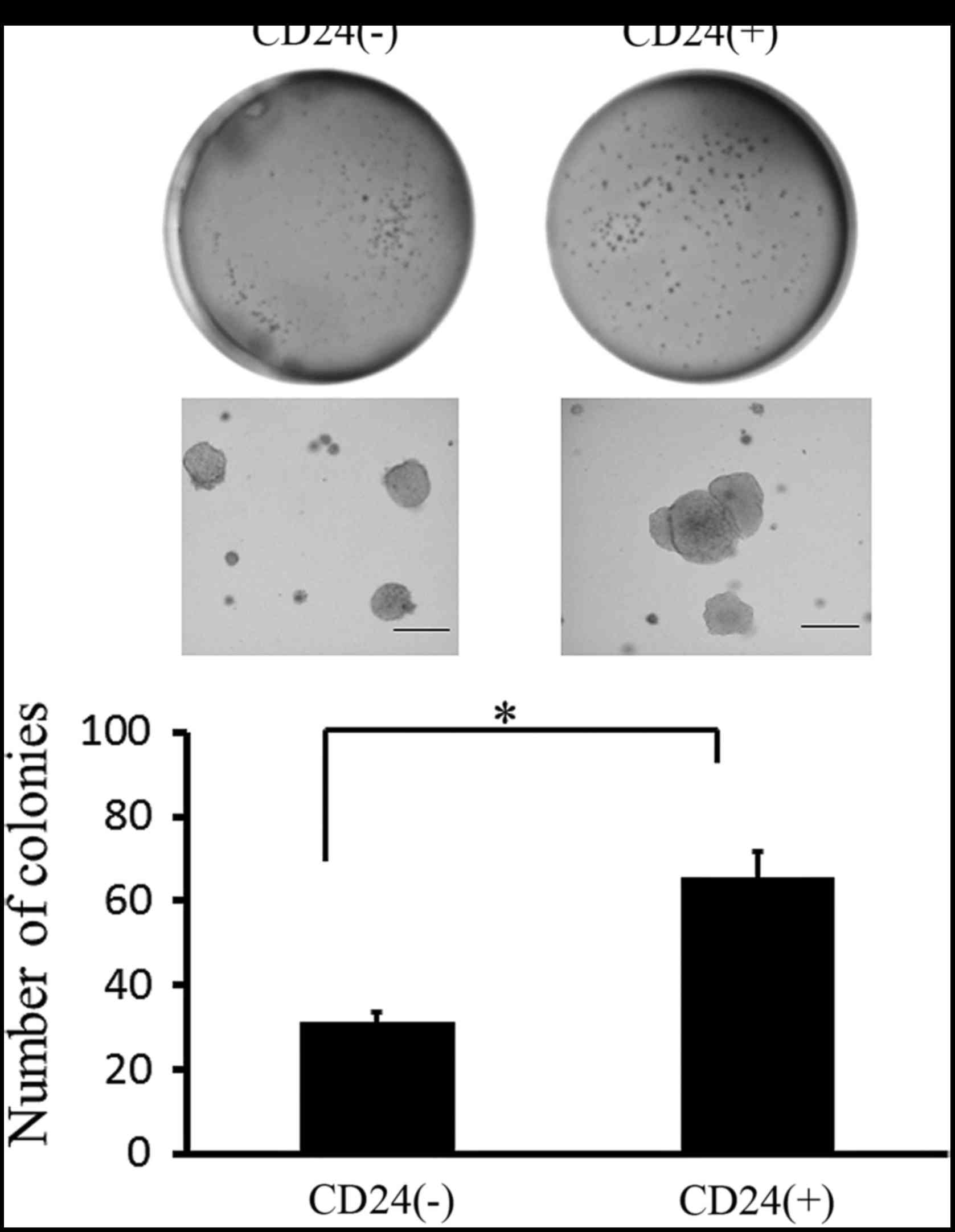
of CD24-positive Caov-3 cells. As shown in Fig. 5, after 28 days of culture,
CD24-positive Caov-3 cells formed more colonies than CD-negative
Caov-3 cells.
To obtain further insight into the biological
relevance of CD24-positive cells, we analyzed their effect on the
proliferative activity of Caov-3 cells by manual cell counting. As
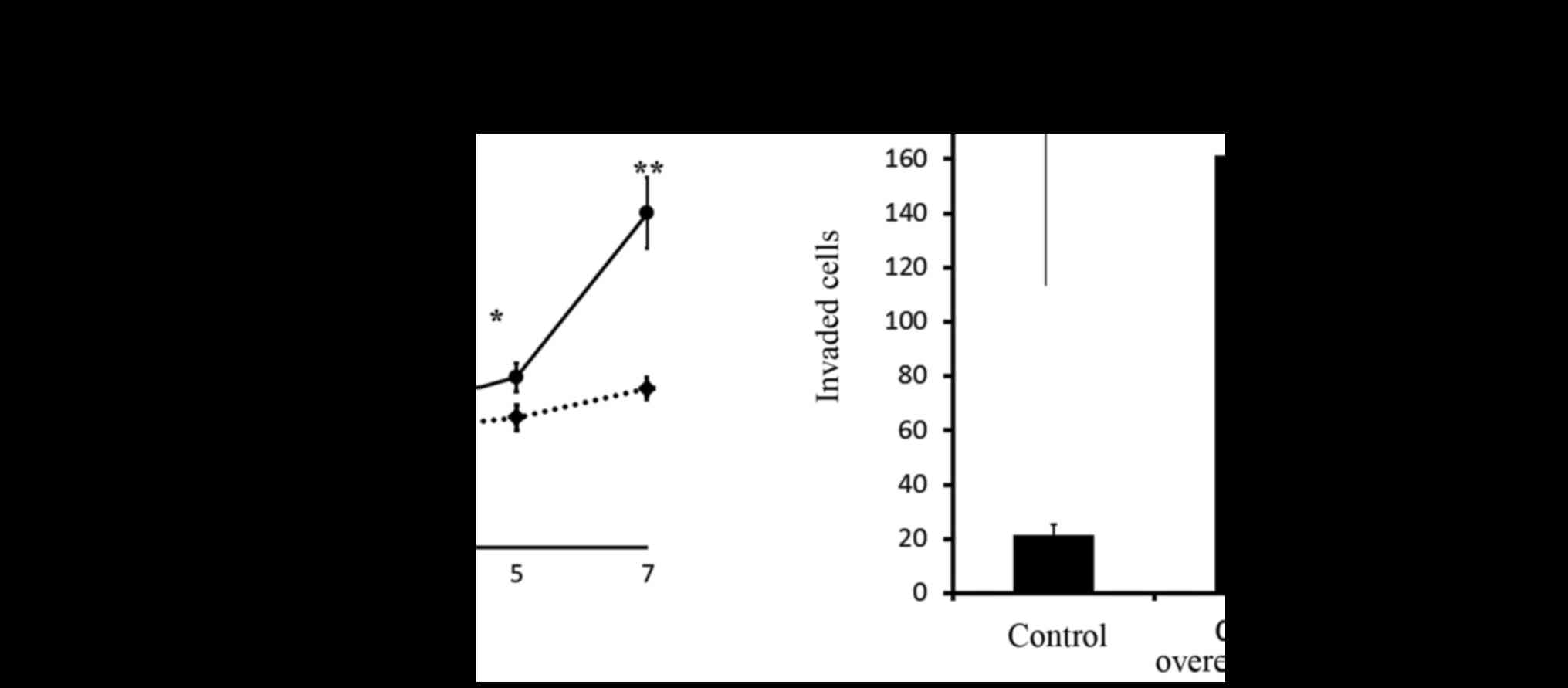
shown in Fig. 6A, CD24-positive
Caov-3 cells showed an enhanced proliferation rate after 7 days of
culture in comparison to CD24-negative Caov-3 cells. We next
examined the correlation between the expression of CD24 and the
invasiveness of ovarian cancer. CD24-overexpressing cells showed
7.5-fold greater invasion in comparison to the control cells
(p<0.01). Moreover, CD24 siRNA-transfected cells significantly
inhibited the invasive activity of ovarian cancer (Fig. 6B).
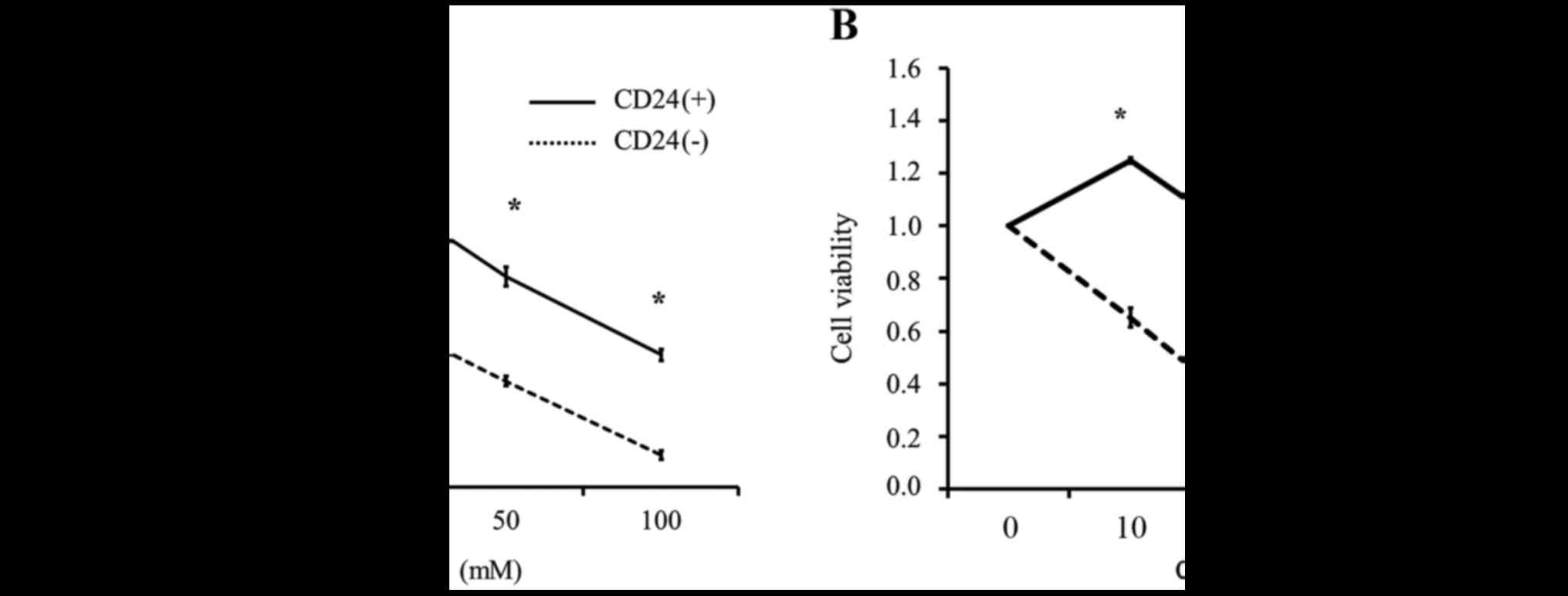
To clarify whether the CD24-positive cells were more
resistant to cisplatin than CD24-negative cells, we divided the
Caov-3 cells into CD24-positive and CD24-negative fractions using
MACS and performed an MTS assay with various concentrations of
cisplatin. As shown in Fig. 7A, the
CD24-positive Caov-3 cells were more resistant to cisplatin
treatment than the CD24-negative Caov-3 cells. We confirmed that
the CD24-overexpressing cells were also more resistant to cisplatin
treatment than the CD24 siRNA-transfected cells (Fig. 7B).
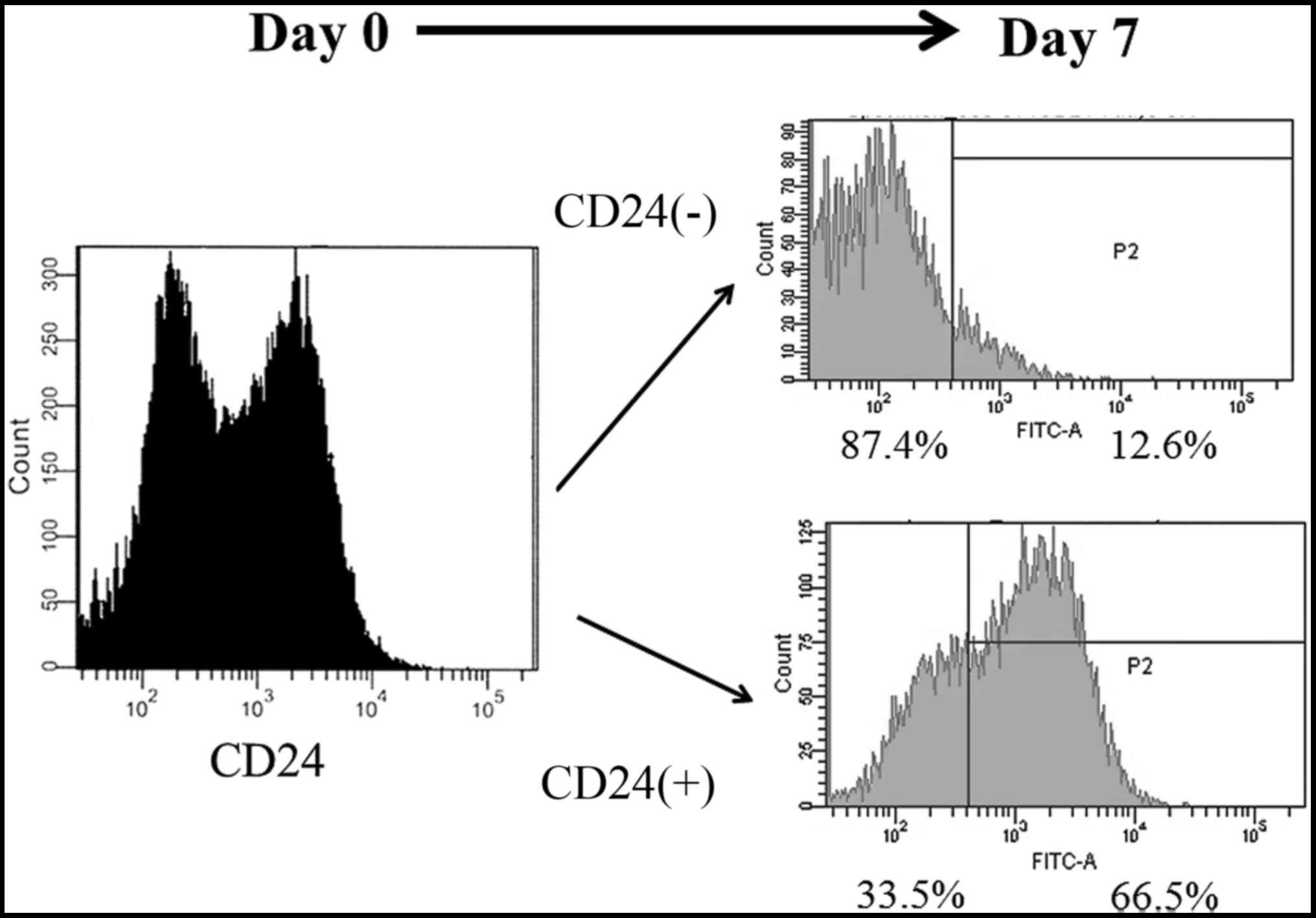
The long-term differentiation
potential of CD24-positive cells
During long-term culture, in addition to increasing
their cell mass, CSCs also undergo asymmetric division to generate
a cell population with heterogeneous phenotypes and low
tumorigenicity (33). We divided
the Caov-3 cells into CD24-positive and CD24-negative fractions
using MACS, and examined whether the CD24-positive and negative
fractions of Caov-3 cells changed in the fraction after 7 days of
culture by flow cytometry, as shown in Fig. 8. After 7 days of culture, 66.5% of
the CD24-positive fractions cells were CD24-positive and 33.5% were
CD24-negative. Interestingly, after 7 days of culture, 87.4% of the
CD24-negative fractions cells were CD24-negative, while only 12.6%
were CD24-positive. These results suggest that CD24-positive cells
show self-renewal potential and cell differentiation potential
after prolonged culture.
CD24 differentially promotes the
activation of the Akt and ERK pathway via the TGF-β signaling
cascade in ovarian cancer cell lines
TGF-β is major inducer of the EMT during embryonic
development, the pathogenesis of fibrotic disorders and cancer
progression (26–28). It was first described as an inducer
of the EMT in normal mammary epithelial cells (29). As the TGF-β signaling pathway is
located upstream of both the PI3k/Akt and MAPK pathways, we
anticipated that there would be a connection between CD24 and the
TGF-β signaling pathway. To determine whether CD24 has any effect
on the TGF-β signaling pathway, the Akt phosphorylation status and
the ERK phosphorylation status were assessed by western blotting.
As shown in Fig. 9, both Akt and
ERK were significantly more phosphorylated in Caov-3 cells that
were treated with TGF-β than in Caov-3 cells that were not treated
with TGF-β. The phosphorylation of both Akt and ERK was
significantly enhanced in CD24-overexpressing Caov-3 cells in
comparison to the scrambled transfected Caov-3 cells. Moreover, we
examined whether CD24 had any effect on the NF-κB phosphorylation
status. NF-κB was significantly more phosphorylated in Caov-3 cells
that were treated with TGF-β, and was significantly enhanced in
CD24-overexpressing Caov-3 cells in comparison to control cells
(data not shown). These data suggest that CD24 somehow enhances the
TGF-β signaling pathway and that the PI3K/Akt and MAPK pathways may
be promoted via this enhanced TGF-β signaling.
CD24 enhances the intra-abdominal
disseminated tumorigenesis of ovarian cancer in vivo
Peritoneal dissemination is the primary route of
progression in human ovarian cancer, and the amount of ascites and
the disseminated tumor burden are correlated with the prognosis of
human patients (34). We therefore
examined the correlation between the expression of CD24 and the
intra-abdominally disseminated tumorigenesis of ovarian cancer. We
divided the Caov-3 cells into CD24-positive and CD24-negative
fractions using MACS, and athymic nude mice were intraperitoneally
inoculated with CD24-positive Caov-3 cells and CD24-negative cells,
as described in Materials and methods. The appearance of the mice
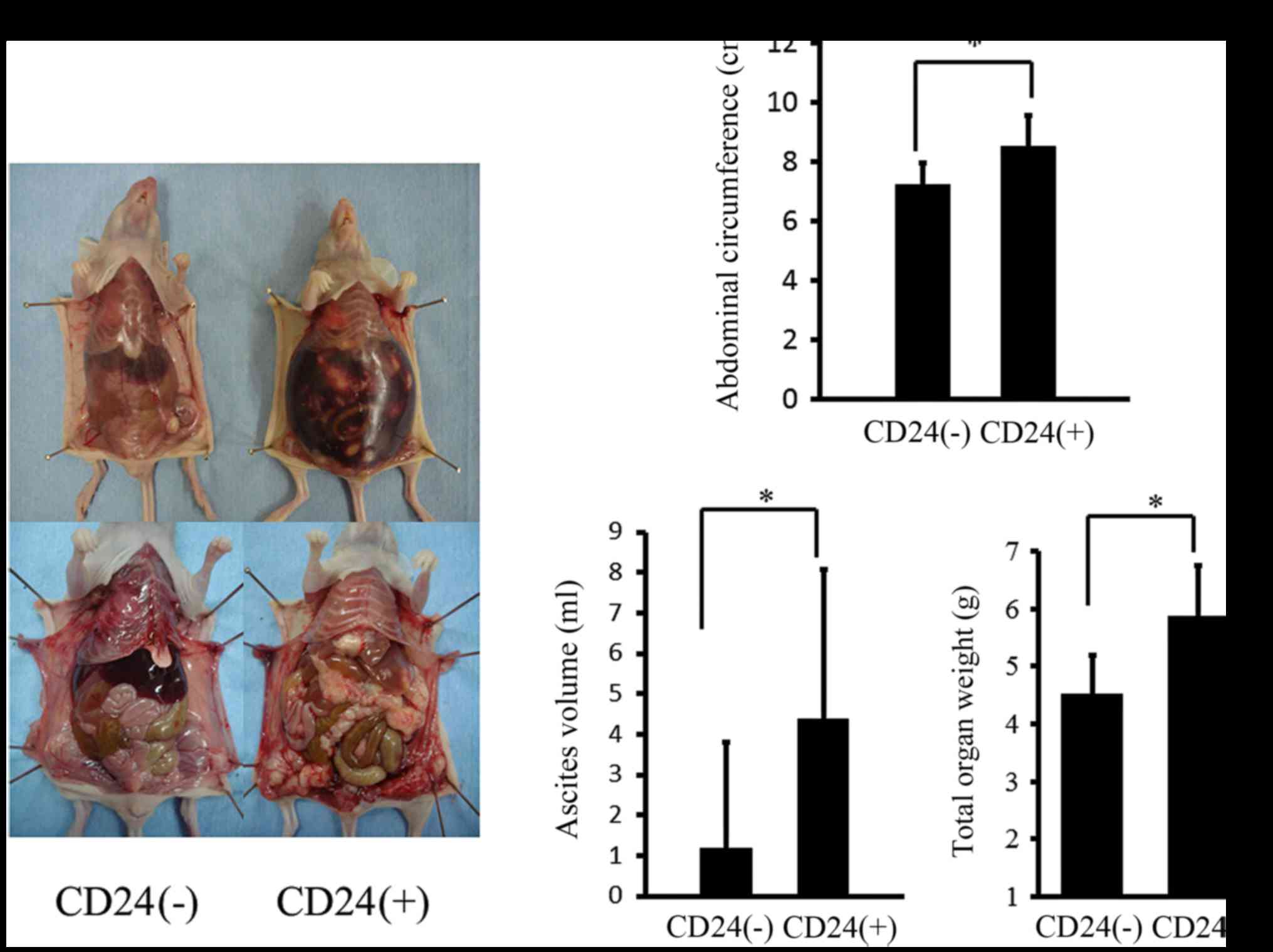
is shown in Fig. 10A. After 50
days, intra-abdominal dissemination was clearly detected in the
athymic nude mice that were intraperitoneally inoculated with
CD24-positive cells in comparison to CD24-negative cells. After
performing a histological examination, the abdominal tumors were
found to be papillary adenocarcinomas, which was consistent with
the characteristics of the Caov-3 cells. The mean abdominal
circumference after 50 days in the athymic nude mice that were
intraperitoneally inoculated with CD24-positive cells was
significantly higher than that observed in the mice with
CD24-negative cells (Fig. 10B).
The mean ascites volume after 50 days in the athymic nude mice that
were intraperitoneally inoculated with CD-24 positive cells was
also significantly higher than that observed in the mice with
CD24-negative cells (Fig. 10B).
The disseminated tumor weight after 50 days in the athymic nude
mice that were intraperitoneally inoculated with CD24-positive
cells was significantly higher than that in mice that were
inoculated wit CD24-negative cells (Fig. 10B).
Discussion
We found that CD24 was expressed in 70.1% of ovarian
cancers and that the expression of CD24 was an independent
predictor of survival in patients with ovarian cancer. This finding
is in line with previous reports (35,36).
Moreover, we demonstrated that the expression of CD24 was
correlated with the FIGO stage (p<0.01), and the presence of
peritoneal (p<0.01) and lymph node metastasis (p<0.01). In
addition, CD24 induced the EMT phenomenon, which was involved in
cell invasion, the highly proliferative phenotype, resistance to
chemotherapy, and the properties of cancer stem-like cells (CSCs).
Moreover, we clarified, for the first time, that the CD24-induced
proliferation and invasiveness in ovarian cancer were dependent on
the activation of PI3K/Akt, NF-κB and ERK.
EMT, which is associated with a key step in tumor
metastasis via the induction of a highly invasive phenotype, has
been intensely studied (4,37). We previously showed that the
expression of EMT-related proteins was correlated with the status
of tumor metastasis or the prognostic value in ovarian carcinoma
(38). Given that 33 of 174
patients (19.0%) had EMT-positive status, as represented by both a
reduced E-cadherin expression and the presence of nuclear Snail
expression [4 (12.1%) in the CD24-negative group and 29 (97.9%) in
the CD24-positive group], we concluded that a strong relationship
existed between the expression of CD24 and EMT status (p<0.05)
(data not shown). In this study, we demonstrated that CD24-positive
ovarian cancer cells consistently acquired an EMT phenotype. These
data indicate that CD24-positive cells are involved in the EMT
process in ovarian cancer. In the present study, we also
demonstrated that CD24-positive ovarian carcinomas have greater
potential for intra-abdominal tumor cell dissemination and ascites
production than CD24-negative ovarian carcinomas in in vivo
models.
EMT is a clinically important multifaceted
pathological program that endows cancer cells with the ability of
invasion, resistance to apoptosis and dissemination. EMT-inducing
transcription factors such as Snail, Twist and Slug are aberrantly
expressed in human cancers and are responsible for a poor prognosis
in patients (5). In the present
study, we demonstrated that CD24 enhanced the expression of Snail,
and decreased the expression of E-cadherin in the Caov-3 cells. We
herein demonstrated, for the first time, that CD24 amplified cell
growth-related intracellular signaling via the PI3K/Akt and MAPK
pathways by affecting the EMT signal pathways. Some groups have
reported that EMT also contributes to the properties of CSCs
(31,32,39),
and CSCs have been found to transiently acquire stem cell-like
properties as a consequence of EMT (40). The CSC hypothesis states that solid
tumors are hierarchically organized and sustained by a minority of
the tumor cell population with stem cell-like properties, such as
self-renewal, multilineage differentiation and tumorigenicity. CD24
is a mucin-like cell surface glycoprotein that is frequently
overexpressed in various human cancers and is correlated with a
poor prognosis (35,41). Recent reports have revealed that
CD24-positive cells function as CSCs in colon cancer,
hepatocellular carcinoma, pancreatic carcinomas, and ovarian
carcinomas (23,24,41,42).
In the present study, we also demonstrated that CD24-positive cells
show self-renewal potential and cell differentiation potential
after prolonged culture, and increase the colony-forming activity,
invasive activity, proliferation activity, and resistance to
cisplatin in comparison to CD24-negative cells. We also
demonstrated that CD24 induced the EMT phenomenon, which was
involved in cell invasion, resistance to chemotherapy, and the
formation of side populations of CSCs (7); however, we could not clarify what
regulated the expression of CD24, and CD24-positive cells were
identical with ovarian cancer stem cells in that study. In the
present study, we indicated that CD24 plays a critical role in
regulating the EMT phenomenon in ovarian cancer. Our study provides
novel insight into the interaction between CSCs and the EMT program
and a better understanding of the mechanism underlying the
involvement of CD24 in ovarian cancer metastasis. Given the
findings in the present study, we believe that CD24 is a key
molecule of metastatic progression in the EMT phenomenon and a
promising therapeutic target for advanced ovarian cancer.
Acknowledgements
This study was supported by a JSPS KAKENHI grant no.
25462621 (to Y. Terai).
Glossary
Abbreviations
Abbreviations:
|
CD24
|
cluster of differentiation 24
|
|
EMT
|
epithelial-mesenchymal-transition
|
|
CSCs
|
cancer stem-like cells
|
|
PI3K-Akt
|
phosphatidylinositol 3-kinase-Akt
|
|
NF-κB
|
nuclear factor-κB
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
MAPK
|
mitogen-activated protein kinase
|
|
RT-PCR
|
reverse transcription polymerase chain
reaction
|
|
MTS assay
|
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
assay
|
|
MASC
|
magnetic activated cell sorting
|
|
FIGO
|
International Federation of Gynecology
and Obstetrics
|
|
SSPCs
|
serous surface papillary
carcinomas
|
References
|
1
|
Ozols RF: Treatment goals in ovarian
cancer. Int J Gynecol Cancer. 15:3–11. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kang Y and Massagué J:
Epithelial-mesenchymal transitions: Twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Krantz SB, Shields MA, Dangi-Garimella S,
Munshi HG and Bentrem DJ: Contribution of epithelial-to-mesenchymal
transition and cancer stem cells to pancreatic cancer progression.
J Surg Res. 173:105–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dalerba P, Cho RW and Clarke MF: Cancer
stem cells: Models and concepts. Annu Rev Med. 58:267–284. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kay R, Rosten PM and Humphries RK: CD24, a
signal transducer modulating B cell activation responses, is a very
short peptide with a glycosyl phosphatidylinositol membrane anchor.
J Immunol. 147:1412–1416. 1991.PubMed/NCBI
|
|
10
|
Aigner S, Ruppert M, Hubbe M, Sammar M,
Sthoeger Z, Butcher EC, Vestweber D, Altevogt P and Kaufmann SHE:
Heat stable antigen (mouse CD24) supports myeloid cell binding to
endothelial and platelet P-selectin. Int Immunol. 7:1557–1565.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee KM, Ju JH, Jang K, Yang W, Yi JY, Noh
DY and Shin I: CD24 regulates cell proliferation and transforming
growth factor β-induced epithelial to mesenchymal transition
through modulation of integrin β1 stability. Cell Signal.
24:2132–2142. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aigner S, Sthoeger ZM, Fogel M, Weber E,
Zarn J, Ruppert M, Zeller Y, Vestweber D, Stahel R, Sammar M, et
al: CD24, a mucin-type glycoprotein, is a ligand for P-selectin on
human tumor cells. Blood. 89:3385–3395. 1997.PubMed/NCBI
|
|
13
|
Sammar M, Aigner S and Altevogt P:
Heat-stable antigen (mouse CD24) in the brain: Dual but distinct
interaction with P-selectin and L1. Biochim Biophys Acta.
1337:287–294. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
King JB, von Furstenberg RJ, Smith BJ,
McNaughton KK, Galanko JA and Henning SJ: CD24 can be used to
isolate Lgr5+ putative colonic epithelial stem cells in
mice. Am J Physiol Gastrointest Liver Physiol. 303:G443–G452. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng J, Gao G, Wang L, Wang T, Yu J and
Zhao Z: CD24 expression as a marker for predicting clinical outcome
in human gliomas. J Biomed Biotechnol. 2012:5171722012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi Y, Gong HL, Zhou L, Tian J and Wang Y:
CD24: A novel cancer biomarker in laryngeal squamous cell
carcinoma. ORL J Otorhinolaryngol Relat Spec. 74:78–85. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kristiansen G, Schlüns K, Yongwei Y,
Denkert C, Dietel M and Petersen I: CD24 is an independent
prognostic marker of survival in nonsmall cell lung cancer
patients. Br J Cancer. 88:231–236. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kristiansen G, Winzer KJ, Mayordomo E,
Bellach J, Schlüns K, Denkert C, Dahl E, Pilarsky C, Altevogt P,
Guski H, et al: CD24 expression is a new prognostic marker in
breast cancer. Clin Cancer Res. 9:4906–4913. 2003.PubMed/NCBI
|
|
19
|
Lee HJ, Kim DI, Kwak C, Ku JH and Moon KC:
Expression of CD24 in clear cell renal cell carcinoma and its
prognostic significance. Urology. 72:603–607. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Su N, Peng L, Xia B, Zhao Y, Xu A, Wang J,
Wang X and Jiang B: Lyn is involved in CD24-induced ERK1/2
activation in colorectal cancer. Mol Cancer. 11:432012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu C, Zheng S, Shen H, Xu K, Chen J, Li
H, Xu Y, Xu A, Chen B, Kaku H, et al: Clinical significance of CD24
as a predictor of bladder cancer recurrence. Oncol Lett. 6:96–100.
2013.PubMed/NCBI
|
|
22
|
Zhu J, Zhang G and Lu H: CD24, COX-2, and
p53 in epithelial ovarian cancer and its clinical significance.
Front Biosci (Elite Ed). 4:2745–2751. 2012.PubMed/NCBI
|
|
23
|
Li C, Heidt DG, Dalerba P, Burant CF,
Zhang L, Adsay V, Wicha M, Clarke MF and Simeone DM: Identification
of pancreatic cancer stem cells. Cancer Res. 67:1030–1037. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao MQ, Choi YP, Kang S, Youn JH and Cho
NH: CD24+ cells from hierarchically organized ovarian
cancer are enriched in cancer stem cells. Oncogene. 29:2672–2680.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim KH, Choi JS, Kim JM, Choi YL, Shin YK,
Lee HC, Seong IO, Kim BK, Chae SW and Kim SH: Enhanced CD24
expression in endometrial carcinoma and its expression pattern in
normal and hyperplastic endometrium. Histol Histopathol.
24:309–316. 2009.PubMed/NCBI
|
|
26
|
Taylor MA, Parvani JG and Schiemann WP:
The pathophysiology of epithelial-mesenchymal transition induced by
transforming growth factor-beta in normal and malignant mammary
epithelial cells. J Mammary Gland Biol Neoplasia. 15:169–190. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zavadil J and Böttinger EP: TGF-beta and
epithelial-to-mesenchymal transitions. Oncogene. 24:5764–5774.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee YH, Albig AR, Regner M, Schiemann BJ
and Schiemann WP: Fibulin-5 initiates epithelial-mesenchymal
transition (EMT) and enhances EMT induced by TGF-beta in mammary
epithelial cells via a MMP-dependent mechanism. Carcinogenesis.
29:2243–2251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Miettinen PJ, Ebner R, Lopez AR and
Derynck R: TGF-beta induced transdifferentiation of mammary
epithelial cells to mesenchymal cells: Involvement of type I
receptors. J Cell Biol. 127:2021–2036. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jiang J, Tang YL and Liang XH: EMT: A new
vision of hypoxia promoting cancer progression. Cancer Biol Ther.
11:714–723. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bao B, Wang Z, Ali S, Kong D, Banerjee S,
Ahmad A, Li Y, Azmi AS, Miele L and Sarkar FH: Over-expression of
FoxM1 leads to epithelial-mesenchymal transition and cancer stem
cell phenotype in pancreatic cancer cells. J Cell Biochem.
112:2296–2306. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Roszkowski P, Wronkowski Z, Szamborski J
and Romejko M: Evaluation of selected prognostic factors in ovarian
cancer. Eur J Gynaecol Oncol. 14 Suppl:140–145. 1993.PubMed/NCBI
|
|
35
|
Kristiansen G, Denkert C, Schlüns K, Dahl
E, Pilarsky C and Hauptmann S: CD24 is expressed in ovarian cancer
and is a new independent prognostic marker of patient survival. Am
J Pathol. 161:1215–1221. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Surowiak P, Materna V, Kaplenko I,
Spaczynski M, Dietel M, Kristiansen G, Lage H and Zabel M:
Unfavorable prognostic value of CD24 expression in sections from
primary and relapsed ovarian cancer tissue. Int J Gynecol Cancer.
16:515–521. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Grünert S, Jechlinger M and Beug H:
Diverse cellular and molecular mechanisms contribute to epithelial
plasticity and metastasis. Nat Rev Mol Cell Biol. 4:657–665. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Takai M, Terai Y, Kawaguchi H, Ashihara K,
Fujiwara S, Tanaka T, Tsunetoh S, Tanaka Y, Sasaki H, Kanemura M,
et al: The EMT (epithelial-mesenchymal-transition)-related protein
expression indicates the metastatic status and prognosis in
patients with ovarian cancer. J Ovarian Res. 7:762014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hollier BG, Tinnirello AA, Werden SJ,
Evans KW, Taube JH, Sarkar TR, Sphyris N, Shariati M, Kumar SV,
Battula VL, et al: FOXC2 expression links epithelial-mesenchymal
transition and stem cell properties in breast cancer. Cancer Res.
73:1981–1992. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Iwatsuki M, Mimori K, Yokobori T, Ishi H,
Beppu T, Nakamori S, Baba H and Mori M: Epithelial-mesenchymal
transition in cancer development and its clinical significance.
Cancer Sci. 101:293–299. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee TK, Castilho A, Cheung VC, Tang KH, Ma
S and Ng IO: CD24(+) liver tumor-initiating cells drive
self-renewal and tumor initiation through STAT3-mediated NANOG
regulation. Cell Stem Cell. 9:50–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yeung TM, Gandhi SC, Wilding JL, Muschel R
and Bodmer WF: Cancer stem cells from colorectal cancer-derived
cell lines. Proc Natl Acad Sci USA. 107:pp. 3722–3727. 2010;
View Article : Google Scholar : PubMed/NCBI
|