Introduction
Colorectal cancer (CRC) is one of the most common
types of cancers in both men and women, worldwide (1). Current treatment of CRC usually
involve surgery; however, ~20% of patients who undergo surgery
ultimately develop metastases over the follow-up period (2). Treatment using 5-fluorouracil
(5-FU)-based regimen has also been used as a standard therapeutic
approach for CRC patients. However, due to drug resistance and
related side-effects, the five-year survival rate is an
unsatisfactory 64.9% (3). Recently,
the use of traditional Chinese medicine (TCM) formulas consisting
of various natural compounds, have been associated with potent
anticancer therapeutic effects with fewer side-effects.
Scutellaria barbata D. Don (SB) is a
well-known TCM formula with strong anticancer effects against
various cancers including CRC (4–7). We
had previously demonstrated that the ethanol extract of SB
inhibited colorectal cancer growth in vivo and in
vitro via promoting apoptosis, while suppressing proliferation
and tumor angiogenesis (7–10). Furthermore, we determined that of
the four polar fractions (petroleum ether, chloroform, ethyl
acetate and N-butanol) of SB, the chloroform fraction of SB (ECSB)
exhibited the most potent inhibitory effect on colorectal cancer
growth, via upregulation of pro-apoptotic Bax/Bcl-2 ratio and
downregulation of pro-proliferative cyclin D1 and cyclin-dependent
kinase 4 (11). However, the
potential mechanism of ECSB exerting its anticancer effects is
still not fully understood.
miRNAs are commonly involved in the
post-transcriptional control of gene expression by targeting mRNAs
for cleavage or translational suppression (12). Over 60% of all protein coding genes
in humans are predicted to be regulated by miRNAs. Dysregulation of
miRNAs is often implicated as a precursor to various human diseases
including cancer (13,14). Among these, miR-34a has been
implicated as a tumor suppressor in numerous cancers (15). Recently, several protein-coding
genes have been identified to be directly targeted by miR-34a,
including Bcl-2, SIRT1, Notch1/2, Jagged1, CDK4/6, cyclin E2,
cyclin D1, E2F, c-Myc and c-MET (16–18).
Activation of miR-34 results in the downregulation of its
downstream target genes, which in turn regulates various cellular
processes such as cell proliferation, apoptosis, senescence,
migration and invasion (18–21).
For example, miR-34a activation has been associated with
downregulation of Notch signaling in colon cancer stem cells
(22). Therefore, this study was
conducted to examine whether ECSB inhibited the growth of HCT-8
cells through regulating miR-34a and its downstream target
genes.
Materials and methods
Material and reagents
RPMI-1640 medium, fetal bovine serum (FBS),
penicillin-streptomycin, trypsin-EDTA, 3-(4,
5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) and
TRIzol reagent were obtained from Invitrogen (Invitrogen Life
Technologies, Carlsbad, CA, USA). Bcl-2 (no. 3498), Jagged1 (no.
2155), Notch1 (no. 2495), Notch2 (no. 2420) and horseradish
peroxidase (HRP)-conjugated secondary antibodies (anti-rabbit IgG,
no. 7074) were purchased from Cell Signaling Technology (Beverly,
MA, USA). Fluorescein isothiocyanate (FITC)-conjugated Annexin V
apoptosis detection kit was provided by Becton-Dickinson (San Jose,
CA, USA). All the other chemicals, unless otherwise stated, were
obtained from Sigma-Aldrich (St. Louis, MO, USA).
Preparation of the SB extract
SB was purchased from Guo Yi Tang Chinese Herbal
Medicine Store (Fujian, China). Different polar fractions of SB
were prepared as previously described (11). The chloroform fraction of SB (ECSB)
was dissolved in 100% dimethylsulfoxide (DMSO) to a stock
concentration of 200 mg/ml and stored at −20°C. The working
concentration of ECSB was obtained by diluting the stock solution
in the cell culture medium. The final concentration of DMSO in the
cell culture medium was ≤0.25% for all experiments.
Cell culture
Human colon cancer HCT-8 cells were obtained from
the Nanjing KeyGen Biotech. Co. Ltd. (Nanjing, Jiangsu, China).
HCT-8 cells were cultured in RPMI-1640 medium, supplemented with
10% fetal bovine serum, 100 U/ml penicillin and 100 mg/ml
streptomycin in a 37°C humidified incubator supplemented with 5%
CO2.
Evaluation of cell viability using MTT
assay
Cell viability was assessed using MTT colorimetric
assay. HCT-8 cells were seeded into 96-well plates at a density of
1×105 cells/ml and treated with various concentrations
of ECSB for 24 or 48 h, respectively. Following removal of cell
culture medium, 100 µl of MTT (0.5 mg/ml) was added to each well
and cells were incubated for an additional 4 h at 37°C.
Subsequently, the MTT formazan precipitate was dissolved in 100 µl
of DMSO and the resulting absorbance was measured at 570 nm using
an ELISA plate reader (model ELX800; BioTek, Winooski, VT, USA).
The cell viability was determined using the formula: Cell viability
(%) = sample optical density (OD)/ control OD × 100.
Observation of morphologic
changes
HCT-8 cells were seeded into 6-well plates at a
density of 2×105 cells/ml and treated with various
concentrations of ECSB for 48 h. Cell morphology was observed using
a phase-contrast microscope (Olympus, Tokyo, Japan). Photographs
were taken at a magnification of ×200.
Colony formation
HCT-8 cells were seeded into 6-well plates at a
density of 2×105 cells/ml and treated with various
concentrations of ECSB for 48 h. Subsequently, cells were harvested
and diluted in fresh medium without ECSB, and reseeded at a density
of 1,000 cells/well. The cell culture medium was replaced with
fresh medium every three days. Following 12 days, cells were fixed
with 10% formaldehyde, stained with 0.01% crystal violet and
counted. Cell survival rate was calculated by normalizing the
survival rate of control cells as 100%.
Detection of apoptosis
After incubation with various concentrations of ECSB
for 48 h, HCT-8 cell apoptosis was determined by flow cytometry
analysis using a fluorescence-activated cell sorting (FACS) caliber
(Becton-Dickinson, CA, USA) and Annexin V-FITC/PI kit
(Becton-Dickinson). Staining was performed according to the
manufacturer's instructions. Annexin V-positive and PI-negative
cells indicated presence of early apoptosis, whereas both Annexin
V-positive and PI-positive cells indicated late apoptosis.
Small interfering RNAs
Anti-miR-34a oligonucleotide and scrambled
oligonucleotide (used as negative control) were obtained from
Invitrogen (Invitrogen Life Technologies). Transfection was
performed using RNAiMAX kit (Invitrogen Life Technologies)
according to the manufacturer's instructions. After transfection
for 6 h, HCT-8 cells were treated with ECSB (150 µg/ml) for 48 h
and total RNAs and protein were extracted for real-time PCR and
western blot analysis.
RNA extraction and real-time PCR
analysis
Total RNA was isolated using TRIzol reagent
(Invitrogen Life Technologies). First-strand cDNA was generated by
reverse transcription of 2 µg total RNA using Oligo (dT) or special
RT-miR-34a primer and SuperScript II reverse transcriptase
according to the manufacturer's instructions. The mRNA levels of
miR-34a, Bcl-2, Notch1, Notch2 and Jagged1 was determined with
real-time PCR analysis using SYBR Premix Ex Taq II (Takara, Dalian,
China) and ABI 7500 Fast PCR system, according to the
manufacturer's instructions. U6 and B2M were used as the
internal controls for miR-34a and other genes, respectively. The
mRNA expression was quantified by comparing the cycle threshold
(Ct) values. The experimental data were analyzed using the
2−∆∆Ct method. All experiments were performed in
triplicate.
Western blot analysis
HCT-8 cells were lysed with mammalian cell lysis
buffer (Pierce Chemical Co., Rockford, IL, USA) containing protease
and phosphatase inhibitor. The protein concentrations were
quantified using the bicinchoninic acid protein assay (Pierce
Chemical Co.), and 50 µg of proteins were separated on 12% SDS-PAGE
gels and transferred onto PVDF membranes (Millipore Corp.,
Billerica, MA, USA). The membranes were blocked with 5% non-fat dry
milk and probed with primary antibodies against Bcl-2, Notch1,
Notch2, Jagged1 and GAPDH (1:1,000) overnight at 4°C. The membranes
were washed three times with Tris-buffered saline with Tween-20
(TBST) prior to incubation with the appropriate HRP-conjugated
secondary antibodies for 1 h at room temperature. Following washing
in TBST, the protein bands were detected using enhanced
chemiluminescence. GAPDH was used as an internal control.
Statistical analysis
Each experiment was performed three times
independently. Data were expressed as mean ± standard deviation.
Statistical analysis was analyzed using SPSS package for Windows
(version 13.0; SPSS Inc., Chicago, IL, USA) by the Student's
t-test. P-values <0.05 were considered as statistically
significant.
Results
ECSB inhibits the growth of HCT-8
cells
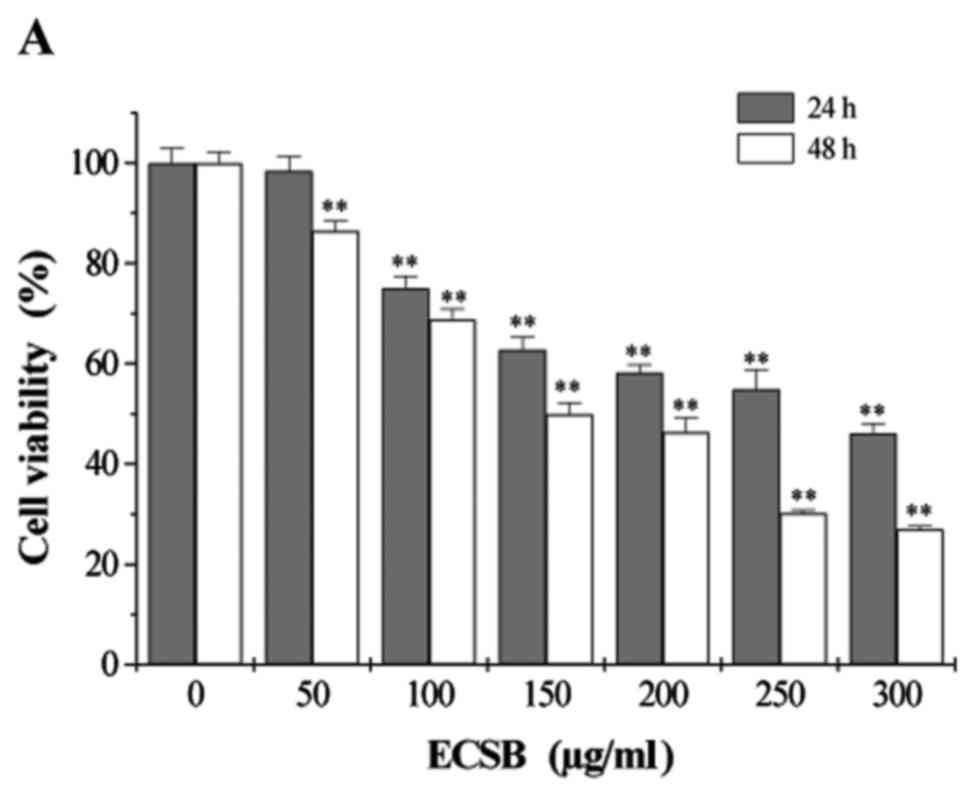
We first examined the effect of ECSB on HCT-8 cell
viability using MTT assay. ECSB treatment significantly inhibited
cell viability in a dose- and time-dependent manner (Fig. 1A). After 24 h, ECSB treatment
decreased HCT-8 cell viability from 98.4 to 36.7%. Similarly, after
48 h, ECSB treatment decreased HCT-8 cell viability from 86.4 to
23.5%. We further examined the effect of ECSB on HCT-8 cell
morphology using phase-contrast microscopy. Untreated control cells
appeared healthy and had a high rate of confluence, whereas ECSB
treatment significantly decreased the confluence and state of HCT-8
cells, in a dose-dependent manner (Fig.
1B). Moreover, ECSB treatment resulted in distinctive rounding
of cells, indicative of cellular apoptosis. Taken together, these
data revealed that ECSB significantly inhibited the growth of HCT-8
cells.
ECSB induces apoptosis and suppresses
proliferation in HCT-8 cells
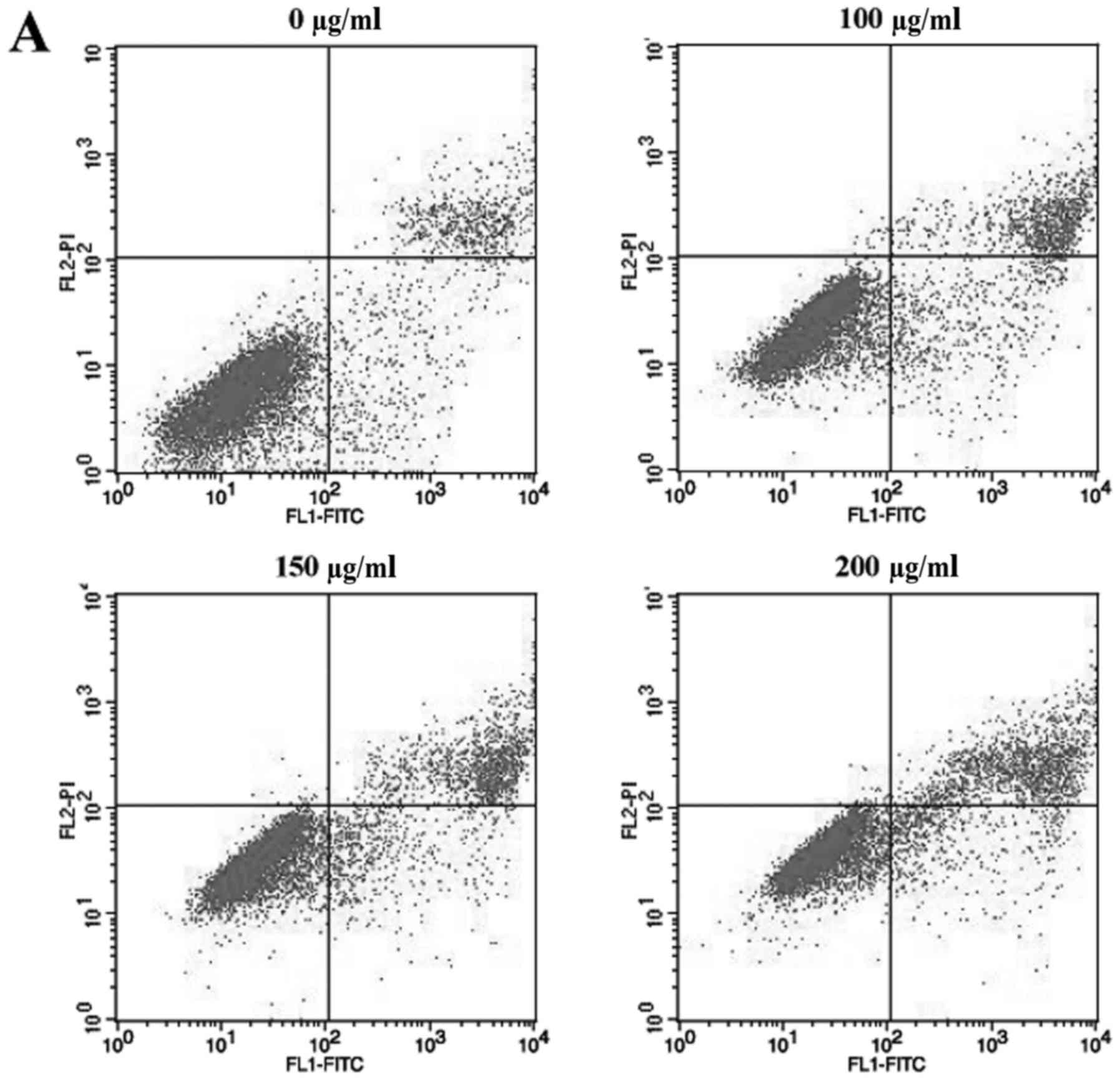
We next examined whether ECSB inhibited cell growth
through inducing cell apoptosis. HCT-8 cells were stained with
Annexin V-FITC/PI and analyzed using flow cytometry analysis
(Fig. 2). Following treatment with
0, 100, 150 and 200 µg/ml ECSB, the percentage of cells undergoing
either early or late apoptosis were 10.42, 16.68, 19.46 and 23.25%,
respectively, which demonstrated that ECSB promoted HCT-8 cell
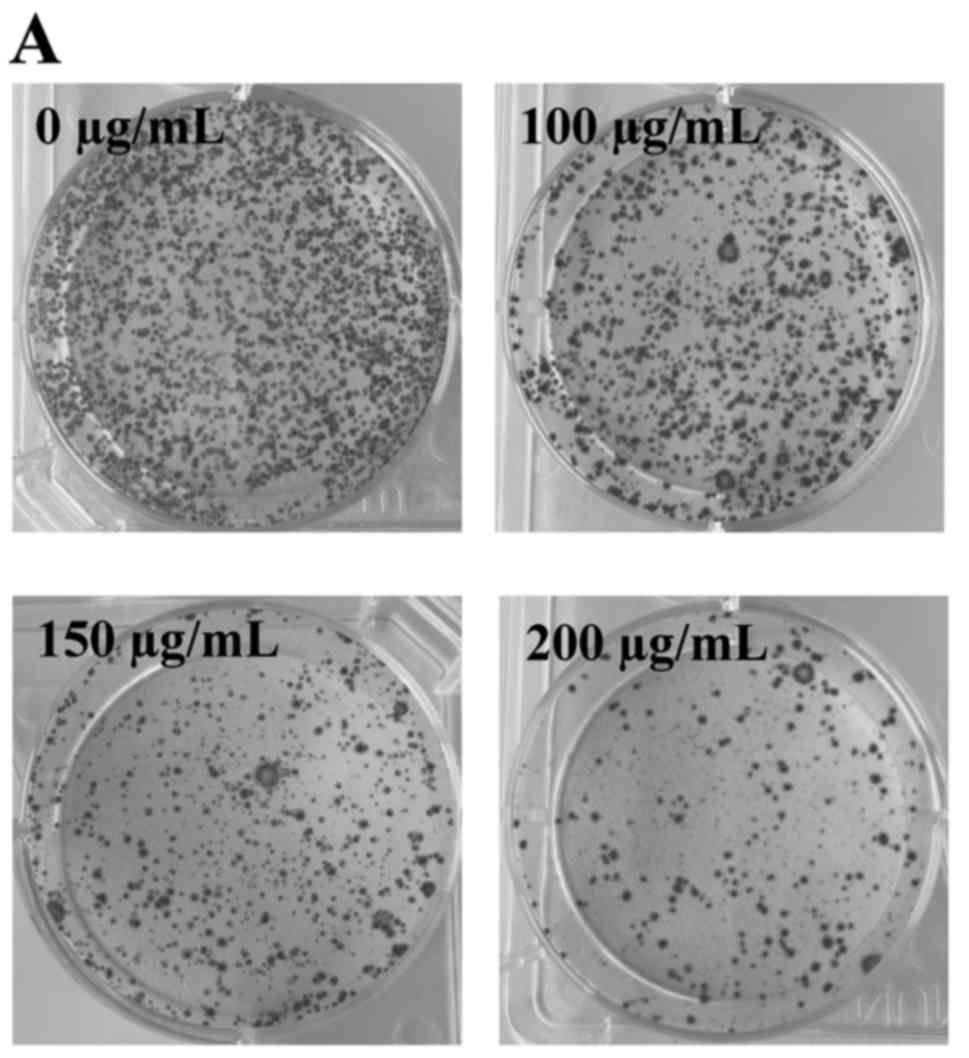
apoptosis in a dose-dependent manner. Furthermore, we performed
colony formation assays to determine the proliferation of HCT-8
cells following ECSB treatment. Treatment with 100, 150 and 200
µg/ml ECSB for 48 h significantly decreased the survival rate of
HCT-8 cells by 27.13, 49.24 and 62.96%, respectively (P<0.05),
which demonstrated that ECSB inhibited the proliferation of HCT-8
cells in a dose-dependent manner (Fig.
3).
ECSB enhances miR-34a expression and
decreases its downstream target genes in HCT-8
miR-34a is a member of the miR-34 family which
functions as a tumor suppressor in numerous cancers including colon
cancer, through inhibition of the genes involved in multiple
oncogenic signaling pathways. In addition, genes which are involved
in cancer cell growth, such as Bcl-2, Notch1, Notch2 and Jagged1
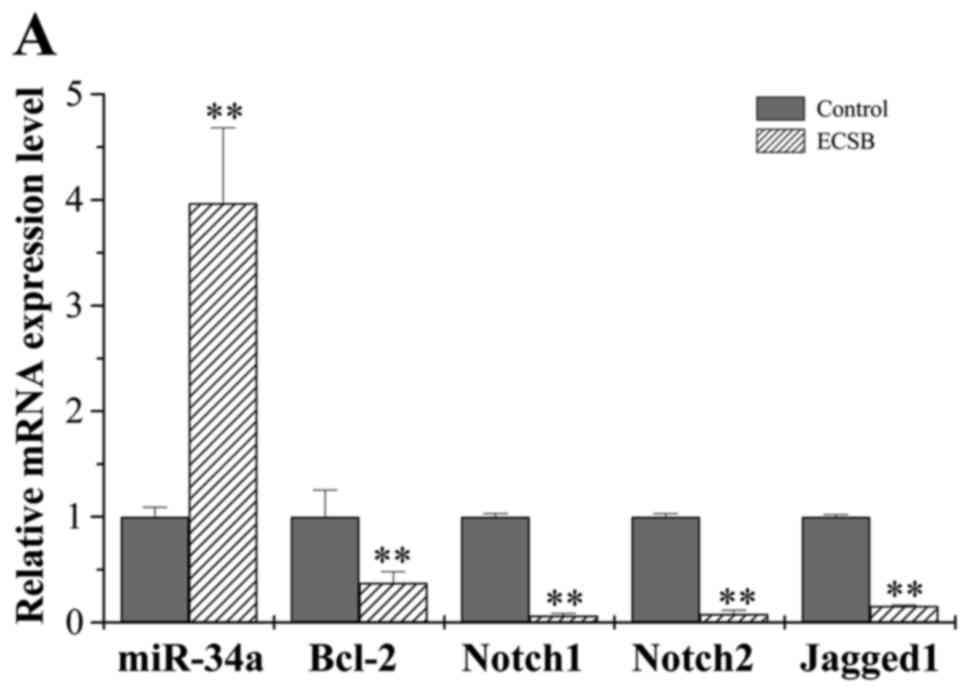
are candidate target genes of miR-34a. We therefore examined
whether ECSB suppresses cancer cell growth through regulation of
miR-34a, using real-time PCR analysis. ECSB treatment (150 µg/ml)
for 48 h significantly increased the level of miR-34a mRNA
expression in HCT-8 cells (Fig.
4A). Furthermore, ECSB treatment also significantly decreased
both the mRNA and protein expression levels of miR-34a target genes
Bcl-2, Notch1/2 and Jagged1 (Fig.
4). These results suggest that the inhibitory effect of ECSB on
cancer cell growth is likely mediated by upregulation of miR-34a
expression and the inhibition of its downstream target genes.
ECSB suppresses cancer cell growth
through directly targeting miR-34a and downregulating its
downstream target genes
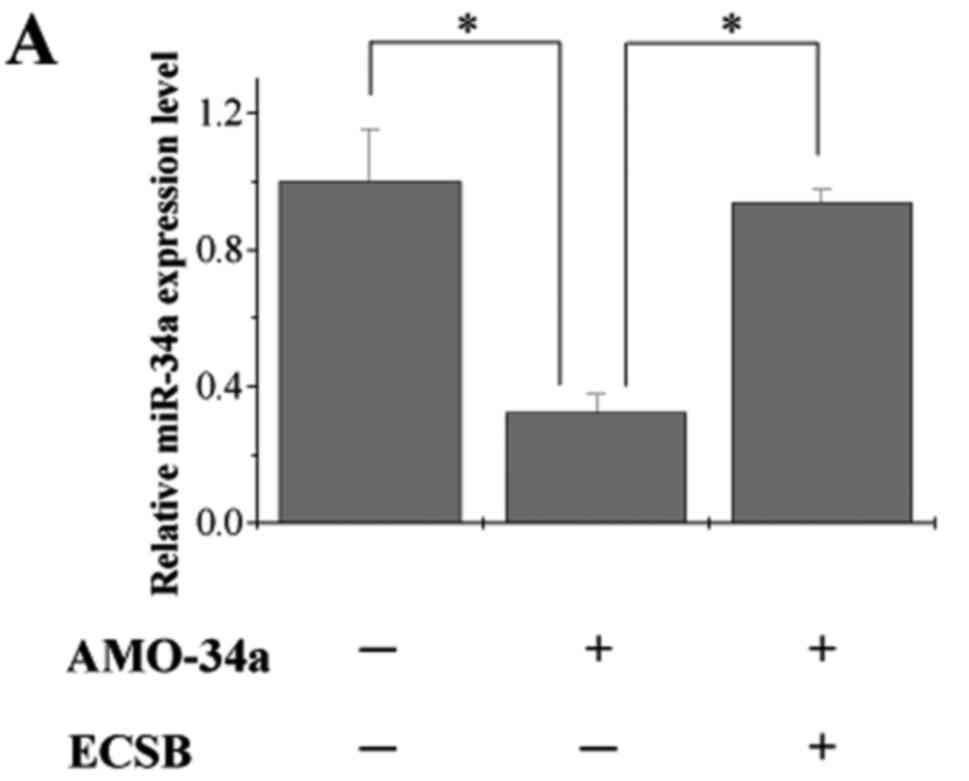
To further verify that ECSB inhibited cancer cell
growth through directly regulating miR-34a and its downstream
target genes (Bcl-2, Notch1, Notch2 and Jagged1), we treated HCT-8
cells with the combination of ECSB and anti-miR-34a oligonucleotide
(AMO-34a) comparing to AMO-34a only and a scrambled oligonucleotide
was used as the negative control. Expression of miR-34a was
significantly decreased following transfection of AMO-34a, which
was rescued by ECSB treatment (Fig.
5A), suggesting that ECSB can directly mediate the expression
of miR-34a. We next examined the mRNA and protein expression of
miR-34a downstream target genes following transfection of AMO-34a
in HCT-8 cells. The mRNA expression of Notch1/2, but not Bcl-2 or
Jagged1 was significantly upregulated whereas the protein
expression of Notch1/2, Bcl-2 and Jagged1 was significantly
upregulated following transfection of AMO-34a in HCT-8 cells.
However, following co-treatment with ECSB, the expression of
Notch1, Notch2, Bcl-2 and Jagged1 was drastically decreased
compared to AMO-34a transfected cells (Fig. 5B-D). Collectively, these findings
reveal that ECSB-induced suppression of cancer cell growth via
directly targeting miR-34a and downregulating its downstream target
genes.
Discussion
The underlying mechanisms of colorectal cancer are
complex and involve a cascade of signaling pathways, including p53,
PI3K, RAS, MAPK, Wnt/β-catenin pathways, and EMT transcription
factors (23). The recent emergence
of systems biology have revealed that a wide range of components
used in TCM exhibit potent anticancer effects (24). SB is known for its anti-inflammatory
and antitumor effects, and is often used in TCM for the treatment
of colon cancer. We had previously demonstrated that SB suppressed
tumor angiogenesis via inhibition of hedgehog pathway in a mouse
model of colorectal cancer; and induced G1/S arrest via modulation
of p53 and Akt pathways in human colon carcinoma cells (8,9). In
addition, SB inhibited colorectal cancer growth and promoted cell
apoptosis via suppression of multiple signaling pathways (7).
Recently, the role of miRNAs in the regulation of
various oncogenic signaling pathways has received significant
attention. For instance, Pien Tze Huang (PZH) inhibited metastasis
of human colon cancer cells via regulation of TGF-β1/ZEB/miR-200
signaling network (25). Similarly,
Hedyotis diffusa plus Scutellaria barbata promoted
bladder cancer cell apoptosis through downregulation of miR-155
expression and inhibition of Akt signaling (26). In this study, we demonstrated that
ECSB significantly inhibited the growth of HCT-8 colon cancer cells
by suppressing cell proliferation and promoting cell apoptosis.
Notably, we revealed that the expression of miR-34a was
significantly increased following ECSB treatment, suggesting that
the inhibitory effect of ECSB on cancer cell growth is likely
mediated by the upregulation of miR-34a.
Notch signaling pathway is involved in a variety of
cellular processes, such as cell proliferation, apoptosis,
migration, invasion and tumor angiogenesis (27), which is activated when a ligand (DLL
or Jagged) interacts with a Notch receptor, resulting in the
cleavage of Notch and subsequent releases the Notch intracellular
domain (NICD) into the nucleus. The NICD acts as a transcription
factor to regulate the expression of Notch target genes, including
Jagged. Consequently, Notch-Jagged signaling forms a positive
feedback loop in the cells (28).
Recent studies have shown that both Notch1/2 receptors and its
ligand Jagged1 are direct downstream targets of miR-34a, which are
downregulated following miR-34a activation (29). The anti-apoptotic protein Bcl-2,
which was triggered to induce cell apoptosis via the activation of
Notch signaling in cancer, is other downstream target gene of
miR-34a (30,31). Furthermore, the increase in miR-34a
expression could also induce apoptosis and inhibit cancer cell
proliferation (16,32). These observations correspond with
this study, which showed that ECSB-induced suppression of cancer
cell growth is mediated by the activation of miR-34a and
downregulation of Notch1/2, Jagged and Bcl-2. Moreover, previous
studies have shown that the drugs genistein and rhamnetin could
also suppress cancer cell growth via activation of miR-34a and
inhibition of Notch signaling pathway (33,34).
miR-34a is transcriptionally induced by the tumor
suppressor p53. It has been proved that the interaction of p53 and
miR-34a is disrupted in various cancer cells (35). Our previous study indicated that SB
can induce the proliferation of human colon carcinoma cells via
modulation of p53 (8). Therefore,
ECSB may directly mediate elevated expression of miR-34a via
upregulation of p53. In addition, there is evidence that aberrant
CpG methylation of miR-34a promoter can result in reduced miR-34a
expression (36). However, the
mechanism whether ECSB is able to reverse hypermethylation of
miR-34a needs to be further explore.
In conclusion, we demonstrated for the first time
that ECSB inhibited colorectal cancer growth in HCT-8 cells via
promotion of cell apoptosis and inhibition of cell proliferation,
which was mediated by activation of miR-34a and likely suppression
of Notch signaling pathway. Our findings also implicate ECSB as an
effective and promising therapeutic agent for the treatment of
CRC.
Acknowledgements
This study was sponsored by Fujian Province Natural
Science Foundation (2015J01687) and the Youth Science Foundation of
Fujian Provincial Health Department (2014-2-29).
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
SB
|
Scutellaria barbata D. Don
|
|
ECSB
|
chloroform fraction of Scutellaria
barbata D. Don
|
|
TCM
|
traditional Chinese medicine
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
DMSO
|
dimethyl sulfoxide
|
|
miR-34a
|
microRNA-34a
|
|
AMO-34a
|
anti-miR-34a oligonucleotide
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cekaite L, Eide PW, Lind GE, Skotheim RI
and Lothe RA: MicroRNAs as growth regulators, their function and
biomarker status in colorectal cancer. Oncotarget. 7:6476–6505.
2016.PubMed/NCBI
|
|
3
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang J, Yang G, Hou G, Liu Q, Hu W, Zhao
PU and He YI: Scutellaria barbata D. Don polysaccharides inhibit
the growth of Calu-3 xenograft tumors via suppression of the HER2
pathway and angiogenesis. Oncol Lett. 9:2721–2725. 2015.PubMed/NCBI
|
|
5
|
Kim KW, Jin UH, Kim DI, Lee TK, Kim MS, Oh
MJ, Kim MS, Kwon DY, Lee YC and Kim CH: Antiproliferative effect of
Scutellaria barbata D. Don. on cultured human uterine leiomyoma
cells by down-regulation of the expression of Bcl-2 protein.
Phytother Res. 22:583–590. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gao J, Lu WF, Dai ZJ, Lin S, Zhao Y, Li S,
Zhao NN, Wang XJ, Kang HF, Ma XB, et al: Induction of apoptosis by
total flavonoids from Scutellaria barbata D. Don in human
hepatocarcinoma MHCC97-H cells via the mitochondrial pathway.
Tumour Biol. 35:2549–2559. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin J, Chen Y, Cai Q, Wei L, Zhan Y, Shen
A, Sferra TJ and Peng J: Scutellaria Barbata D Don Inhibits
Colorectal Cancer Growth via Suppression of Multiple Signaling
Pathways. Integr Cancer Ther. 13:240–248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wei L, Lin J, Wu G, Xu W, Li H, Hong Z and
Peng J: Scutellaria barbata D. Don induces G1/S arrest via
modulation of p53 and Akt pathways in human colon carcinoma cells.
Oncol Rep. 29:1623–1628. 2013.PubMed/NCBI
|
|
9
|
Wei L, Lin J, Xu W, Cai Q, Shen A, Hong Z
and Peng J: Scutellaria barbata D. Don inhibits tumor angiogenesis
via suppression of Hedgehog pathway in a mouse model of colorectal
cancer. Int J Mol Sci. 13:9419–9430. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang Q, Li Q, Chen H, Shen A, Cai Q, Lin
J and Peng J: Scutellaria barbata D. Don inhibits growth and
induces apoptosis by suppressing IL-6-inducible STAT3 pathway
activation in human colorectal cancer cells. Exp Ther Med.
10:1602–1608. 2015.PubMed/NCBI
|
|
11
|
Zhang L, Cai Q, Lin J, Fang Y, Zhan Y,
Shen A, Wei L, Wang L and Peng J: Chloroform fraction of
Scutellaria barbata D. Don promotes apoptosis and suppresses
proliferation in human colon cancer cells. Mol Med Rep. 9:701–706.
2014.PubMed/NCBI
|
|
12
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M,
et al: Human microRNA genes are frequently located at fragile sites
and genomic regions involved in cancers. Proc Natl Acad Sci USA.
101:pp. 2999–3004. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Y, Guessous F, Zhang Y, Dipierro C,
Kefas B, Johnson E, Marcinkiewicz L, Jiang J, Yang Y, Schmittgen
TD, et al: MicroRNA-34a inhibits glioblastoma growth by targeting
multiple oncogenes. Cancer Res. 69:7569–7576. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hermeking H: The miR-34 family in cancer
and apoptosis. Cell Death Differ. 17:193–199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamakuchi M, Ferlito M and Lowenstein CJ:
miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci
USA. 105:pp. 13421–13426. 2008; View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tazawa H, Tsuchiya N, Izumiya M and
Nakagama H: Tumor-suppressive miR-34a induces senescence-like
growth arrest through modulation of the E2F pathway in human colon
cancer cells. Proc Natl Acad Sci USA. 104:pp. 15472–15477. 2007;
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma Y, Bao-Han W, Lv X, Su Y, Zhao X, Yin
Y, Zhang X, Zhou Z, MacNaughton WK and Wang H: MicroRNA-34a
mediates the autocrine signaling of PAR2-activating proteinase and
its role in colonic cancer cell proliferation. PLoS One.
8:e723832013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu J, Wu G, Lv L, Ren YF, Zhang XJ, Xue
YF, Li G, Lu X, Sun Z and Tang KF: MicroRNA-34a inhibits migration
and invasion of colon cancer cells via targeting to Fra-1.
Carcinogenesis. 33:519–528. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bu P, Chen KY, Chen JH, Wang L, Walters J,
Shin YJ, Goerger JP, Sun J, Witherspoon M, Rakhilin N, et al: A
microRNA miR-34a-regulated bimodal switch targets Notch in colon
cancer stem cells. Cell Stem Cell. 12:602–615. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chi Y and Zhou D: MicroRNAs in colorectal
carcinoma - from pathogenesis to therapy. J Exp Clin Cancer Res.
35:432016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Buriani A, Garcia-Bermejo ML, Bosisio E,
Xu Q, Li H, Dong X, Simmonds MS, Carrara M, Tejedor N, Lucio-Cazana
J, et al: Omic techniques in systems biology approaches to
traditional Chinese medicine research: Present and future. J
Ethnopharmacol. 140:535–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen A, Lin W, Chen Y, Liu L, Chen H,
Zhuang Q, Lin J, Sferra TJ and Peng J: Pien Tze Huang inhibits
metastasis of human colorectal carcinoma cells via modulation of
TGF-β1/ZEB/miR-200 signaling network. Int J Oncol. 46:685–690.
2015.PubMed/NCBI
|
|
26
|
Pan L, Sheung Y, Guo W, Rong ZB and Cai
ZM: Hedyotis diffusa plus Scutellaria barbata induce bladder cancer
cell apoptosis by inhibiting Akt signaling pathway through
downregulating miR-155 expression. Evid Based Complement Alternat
Med. 2016:91749032016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo H, Lu Y, Wang J, Liu X, Keller ET, Liu
Q, Zhou Q and Zhang J: Targeting the Notch signaling pathway in
cancer therapeutics. Thorac Cancer. 5:473–486. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Boareto M, Jolly MK, Goldman A, Pietilä M,
Mani SA, Sengupta S, Ben-Jacob E, Levine H and Onuchic JN:
Notch-Jagged signalling can give rise to clusters of cells
exhibiting a hybrid epithelial/mesenchymal phenotype. J R Soc
Interface. 13:132016. View Article : Google Scholar
|
|
29
|
Pang RTK, Leung CON, Ye TM, Liu W, Chiu
PC, Lam KK, Lee KF and Yeung WS: MicroRNA-34a suppresses invasion
through downregulation of Notch1 and Jagged1 in cervical carcinoma
and choriocarcinoma cells. Carcinogenesis. 31:1037–1044. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ye QF, Zhang YC, Peng XQ, Long Z, Ming YZ
and He LY: Silencing Notch-1 induces apoptosis and increases the
chemosensitivity of prostate cancer cells to docetaxel through
Bcl-2 and Bax. Oncol Lett. 3:879–884. 2012.PubMed/NCBI
|
|
31
|
Gao F, Yao M, Shi Y, Hao J, Ren Y, Liu Q,
Wang X and Duan H: Notch pathway is involved in high
glucose-induced apoptosis in podocytes via Bcl-2 and p53 pathways.
J Cell Biochem. 114:1029–1038. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang C, Mo R, Yin B, Zhou L, Liu Y and
Fan J: Tumor suppressor microRNA-34a inhibits cell proliferation by
targeting Notch1 in renal cell carcinoma. Oncol Lett. 7:1689–1694.
2014.PubMed/NCBI
|
|
33
|
Xia J, Duan Q, Ahmad A, Bao B, Banerjee S,
Shi Y, Ma J, Geng J, Chen Z, Rahman KM, et al: Genistein inhibits
cell growth and induces apoptosis through up-regulation of miR-34a
in pancreatic cancer cells. Curr Drug Targets. 13:1750–1756. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jia H, Yang Q, Wang T, Cao Y, Jiang QY, Ma
HD, Sun HW, Hou MX, Yang YP and Feng F: Rhamnetin induces
sensitization of hepatocellular carcinoma cells to a small
molecular kinase inhibitor or chemotherapeutic agents. Biochim
Biophys Acta. 1860:1417–1430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Raver-Shapira N, Marciano E, Meiri E,
Spector Y, Rosenfeld N, Moskovits N, Bentwich Z and Oren M:
Transcriptional activation of miR-34a contributes to p53-mediated
apoptosis. Mol Cell. 26:731–743. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lodygin D, Tarasov V, Epanchintsev A,
Berking C, Knyazeva T, Körner H, Knyazev P, Diebold J and Hermeking
H: Inactivation of miR-34a by aberrant CpG methylation in multiple
types of cancer. Cell Cycle. 7:2591–2600. 2008. View Article : Google Scholar : PubMed/NCBI
|