Introduction
Lung cancer is one of the most common malignancies
with severe mortality worldwide (1,2). There
are ermerging treatments for lung cancer including surgery,
irradiation, chemotherapy and immunotherapy. However, prognosis
remains unsatisfactory (3–5). Recently, comorbid cancer-infection
which represents an independent concomitant microorganism infection
in tumor has attracted new attention, since certain microorganism
infection might induce antitumor immunity responses for a new
treatment strategy. Malaria infection significantly suppresses
murine Lewis lung cancer growth via induction of innate and
adaptive antitumor responses in a mouse model, suggesting that the
malaria parasite may stand for a new strategy or therapeutic
vaccine vector for anti-lung cancer immunotherapy (6). T. gondii infection inhibits
tumor growth in the Lewis lung carcinoma mouse model through the
induction of Th1 immune responses and anti-angiogenic activity
(7).
Mycobacterium tuberculosis (MTB) is an
obligate pathogenic bacterial species in the family
Mycobacteriaceae and the causative agent of tuberculosis
(8). Mononuclear cells recruited to
sites of MTB infection or novel MTB antigens, are exposed to MTB
Toll-like receptor (TLR) ligands. MTB is rich in TLR2 ligands
(9,10), and a role for TLR2 ligand in
expansion of Treg has been previously shown (11). MTB and its components expand
functional CD4+Foxp3+ Treg, which implicates
for effective immunization against MTB (12,13).
It was also reported that active tuberculosis in non-small cell
lung cancer (NSCLC) patients shows better survival outcome,
possibly due to the T lymphocyte infiltration in tumors (14). However, the role of an independent
H37Rv infection in the development of NSCLC is not quite clear.
Here, we demonstrated that independent MTB H37Rv
infection facilitated NSCLC progression. H37Rv cocommitant
infection promoted Treg differentiation and its suppressive
function through enhancing PD-L1 expression on macrophages.
Mechanically, Akt-mTORC1 is responsible for H37Rv sitmulated PD-L1
expression on macrophages. Inactivation of mTORC1 by rapamycin or
knockdown of raptor dereased Treg proportion and further reduced
tumor development enhanced by H37Rv concomitant infection.
Materials and methods
Mice, cells, and bacteria
Female 8- to 10-week-old C57BL/6 mice were purchased
from the SLAC Laboratory (Shanghai, China) and raised in the Animal
Center of the Shanghai Chest Hospital. The animal experiment
facilities were approved by the Shanghai Jiao Tong University
School of Medicine Animal Care and Use Committee. All surgery was
performed under anesthesia, and all efforts were made to minimize
animal suffering. The murine LLC cell line was obtained from the
Chinese Academy of Sciences Cell Bank (Shanghai, China). The H37Rv
strain was obtained from the Shanghai Pulmonary Hospital as a
gift.
Antibodies
Neutralizing antibody to PD-L1 and control IgG were
obtained from BioXcell. Antibodies used in western blotting were
all from Cell Signaling Technology.
Mouse models
C57BL/6 mice were s.c. injected with
2×106 murine LLC cells to establish tumors. At the same
time, the tumor cell-inoculated mice were infected peritoneally
with 2×106 heat-killed H37Rv (H37Rv), while challenged
peritoneal with PBS were used as the control group (Ctr). Animals
were examined daily until the tumors became palpable, after which
the tumor volume was determined daily by measuring the diameter of
the tumors using calipers. The volume was calculated using the
formula, V=(ab2)/2, where a is the long axis, and b is the short
axis.
Rapamycin (Sigma) treatment was performed by
injecting intraperitoneally with 4 mg/kg rapamycin or vehicle
solution twice a week. Rapamycin was first dissolved in 100%
ethanol at 10 mg/ml, diluted in vehicle solution containing 5%
Tween-80 and 5% PEG-400 in PBS to 0.5 mg/ml, and filtered (15).
Flow cytometry
The following antibodies and their corresponding
isotype controls (all purchased from eBioscience, USA) were used
for staining: CD4-Percp, Foxp3-FITC, CD11c-FITC, CD80-PE, MHCII-PE,
PD-L1-PE, F4/80-FITC. CFSE were obtained from Invitrogen, USA.
Cells were washed, fixed and stained according to the
manufacturer's instructions. Samples were run on a FACSCalibur (BD
Biosciences) and analyzed using FlowJo software (TreeStar).
Quantitative RT-PCR
RNA was isolated from cells using the Qiagen RNeasy
Mini kit (Qiagen). cDNA was made using the SuperScript II RT
Reaction kit (Invitrogen) from 2 µg of isolated RNA. Samples were
analyzed on a ABI 9500 RT-PCR System Instrument using SYBR PCR
Master Mix according to the manufacturer's instructions. Specific
primers were as follows: Foxp3 forward, GGCCCTTCTCCAGGACAGA;
reverse, GGCATGGGCATCCACAGT. T-bet forward, ACCT
GTTGTGGTCCAAGTTCAA; reverse, GCCGTCCTTGCT TAGTGATGA. GATA-3
forward, GACCCGAAACCGGAAG ATGT; reverse, CGCGTCATGCACCTTTT. RORrt
forward, TGCGACTGGAGGACCTTCTAC; reverse, TCACCTCCT CCCGTGAAAAG.
CD80 forward, TGGGAAAAACCCCC AGAAG; reverse, CCCCAAAGAGCACAAGTGTGT.
MHCII forward, ACAGCCCAATGTCGTCATCTC; reverse, CCAG
AGTGTTGTGGTGGTTGA. PD-L1 forward, CAGGCCGA GGGTTATCCA; reverse,
CGGGTTGGTGGTCACTGTTT. CD74 forward, CCAACGCGACCTCATCTCTAA; reverse,
AGGGCGGTTGCCCAGTA. CD86 forward, CTGTGGCC CTCCTCCTTGT; reverse,
CTGATTCGGCTTCTTGTGAC ATA. IFN-γ forward, TTGGCTTTGCAGCTCTTCCT;
reverse, TGACTGTGCCGTGGCAGTA. TGF-β forward, GCAGTGGCTGAACCAAGGA;
reverse, AGCAGTGAGCG CTGAATCG. IL-10 forward, GATGCCCCAGGCAGAGAA;
reverse, CACCCAGGGAATTCAAATGC. IL-2 forward, GCAGGCCACAGAATTGAAAGA;
reverse, TGCCGCAG AGGTCCAAGT.
Immunoblotting
Cell lysates were prepared in 2X LSB. Anti-PD-L1
antibody, anti-phospho-AKT (S308), anti-AKT, anti-phospho-S6
(T389), anti-S6K were purchased from Abcam. Anti-β-actin was
purchased from Cell Signaling Technology.
CFSE staining
Cells were washed and resuspended in 5 µM CellTrace
CFSE dilution buffer, and stained for 15 min at 37°C in the dark.
Cells were then centrifuged and washed in PBS containing 2% FBS
twice.
Primary cell culture
The bone marrow-derived macrophages (BMDM) were
prepared as follows: bone marrow cells were fushed from the femurs
and tibias of C57BL/6 mice. The cells were then cultured at
2×106 cells per well in 24-well plates in DMEM
supplemented with 20 ng/ml murine M-CSF. Non-adherent cells were
carefully removed, and fresh medium was added every 2 days. On day
6, the cells were collected for experiments. Naive CD4+
T cells were enriched from splenic mononuclear cells by magnetic
cell sorting using a mouse CD4+ T-cell isolation kit
(Miltenyi Biotec). T cells were cultured with macrophages at a
ratio of 5:1 in the presence of OVA peptide (Sigma), and the
supernatants and cells were analyzed on day 3 of culture.
[3H] proliferation
analysis
CD4+ T cell proliferation was performed
as described (16). In brief,
CD4+ T cells were cocultured with macrophages as
described in 96-well plates for 72 h. Proliferation was assessed
based on the incorporation of [3H]-thymidine (1
µCi/well) during the last 12 h of culture in triplicate wells.
Cells were collected using a cell harvester, and
[3H]-thymidine was quantified using a scintillation
counter.
T cell suppression assay
Primed-T cells were purified from BMDMs-T cell
mixtures by flow cytometry using anti-CD25 and anti-Foxp3
antibodies. The purified T cells were co-cultured with CFSE-labeled
activated CD4+ T cells at 1:1 ratio (primed
T:CFSE-labeled T cell) in 96-well round bottom plates for an
additional 3 days. CFSE intensity was monitored by flow
cytometry.
Statistical analysis
Data in bar graphs are presented as mean ± SEM.
Differences between groups were analyzed with unpaired Student's
t-tests. Statistical significance was set at P<0.05.
Results
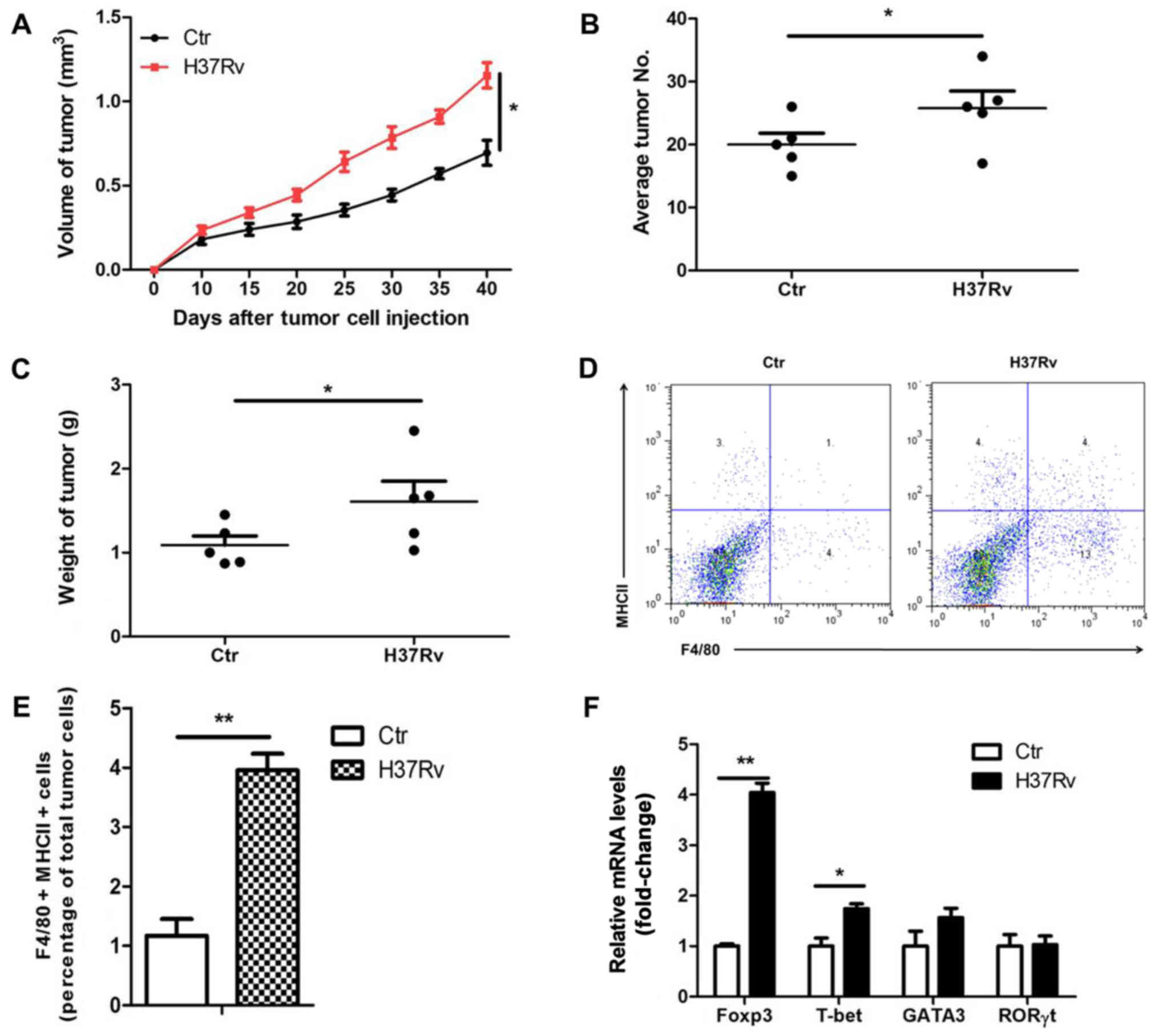
H37Rv infection promotes tumor growth
in a mouse model
To determine the effect of H37Rv infection on the
growth of non-small cell lung cancer (NSCLC), LLC cells were used
to establish a tumor xenograft mouse model. We infected
tumor-bearing mice with H37Rv at the same time. Results showed that
the tumor volumes were clearly increased in the H37Rv-infection
group compared to the non-infection group (Fig. 1A). The number of tumors and tumor
weight were significantly increased in the H37Rv-infection group as
compared with the non-infection group (Fig. 1B and C). Macrophages are the main
cell type infected by H37Rv. Macrophages sense the pathogen,
produce cytokines and activate lymphocytes, thus restraining or
promoting tumor development. We then assessed the changes of immune
responses in the draining lymph nodes after H37Rv infection. H37Rv
infection induced tumor-associated macrophage activation as
assessed by F4/80+MHCII+ by flow cytometry
(Fig. 1D and E). Besides, H37Rv
infection promoted Treg proportion with modest changes in other T
cells types in the drainning lymph nodes (Fig. 1F). These results suggest that H37Rv
infection promotes tumor growth possibly through increasing Treg
proportion.
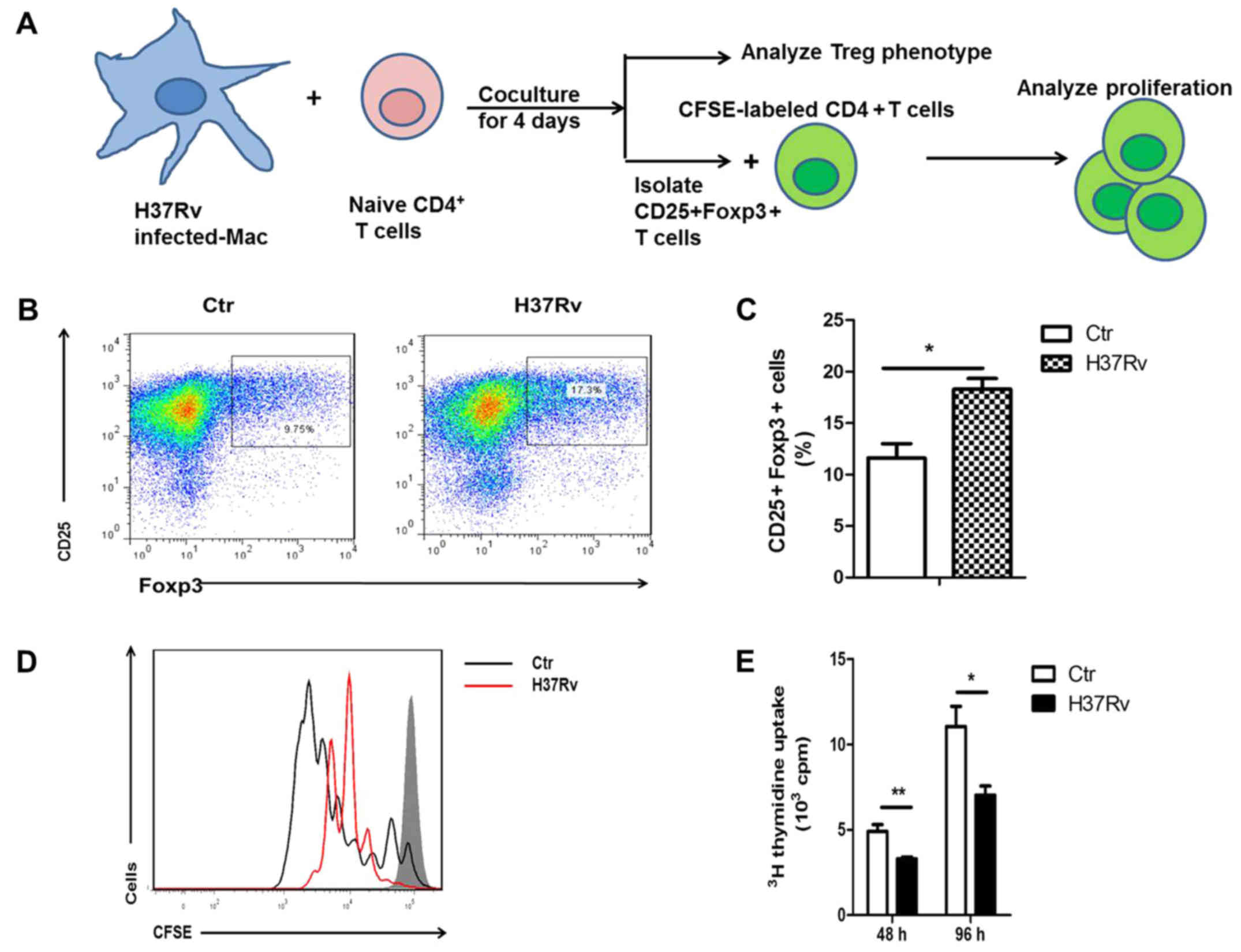
H37Rv-infected macrophages promote
Treg cell differentiation
To determine how H37Rv infection promoted Treg
polarization, we coculture H37Rv-infected BMDMs with naive
CD4+ T cells in vitro for 96 h as described
(Fig. 2A). Of the cultured BMDM
>80% are F4/80+ cells (data not shown). Flow
cytometry analysis showed Treg was enhanced significantly when
co-cultured with H37Rv-infected macrophages, as
CD25+Foxp3+ cell-percentage showed >2-fold
increase (Fig. 2B and C). Generated
Treg cells were also evaluated with a functional assay of T cell
proliferation. We then purified the
CD25+Foxp3+ T cells from the co-culture
system and further co-culture with CFSE-labeled activated
CD4+ T cells for 3 days (Fig. 2A). Naïve T cells primed by
H37Rv-infected macrophges had a stronger inhibitory effect on
anti-CD3 and anti-CD28-stimulated T cells (Fig. 2C). The inhibitory effect was also
observed when similar proliferation was studied by
[3H]-thymidine incorporation (Fig. 2D). These results suggest that
H37Rv-infected macrophages promoted Treg cell differentiation.
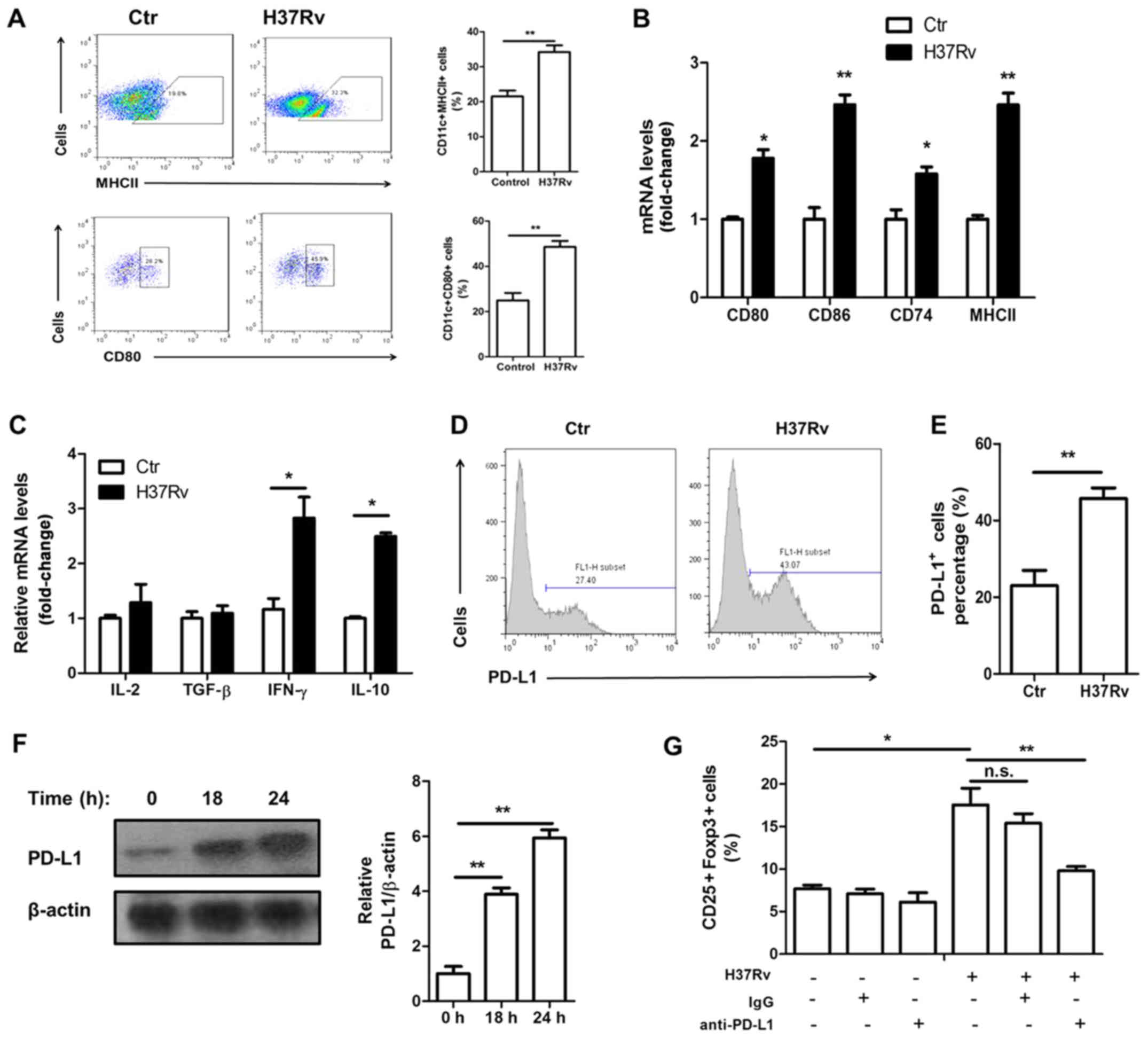
H37Rv promotes macrophage activation
and PD-L1 expression
Macrophage activation is necessary for priming and
polarization of T cells. We found that macrophages were activated
upon H37Rv stimulation. The surface expression of activation
markers MHCII and CD80 was increased after H37Rv stimulation
(Fig. 3A). In addition, their mRNA
levels as well as CD74 and CD86, which both are activation markers,
were all upregulated upon H37Rv infection (Fig. 3B). Treg cell function is controlled
by many signaling pathways, including the IL-2, TGF-β and PD-L1
mediated pathway. H37Rv-infected macrophages showed no increased
IL-2 or TGF-β expression (Fig. 3C).
However, the surface expression of PD-L1 was markedly increased
(Fig. 3D and E). The total protein
levels of PD-L1 were also upregulated after H37Rv infection, with a
time-dependent manner (Fig. 3F).
Blocking PD-L1 using anti-PD-L1 neutralizing antibody significantly
impaired Treg proportion in the coculture system (Fig. 3G).
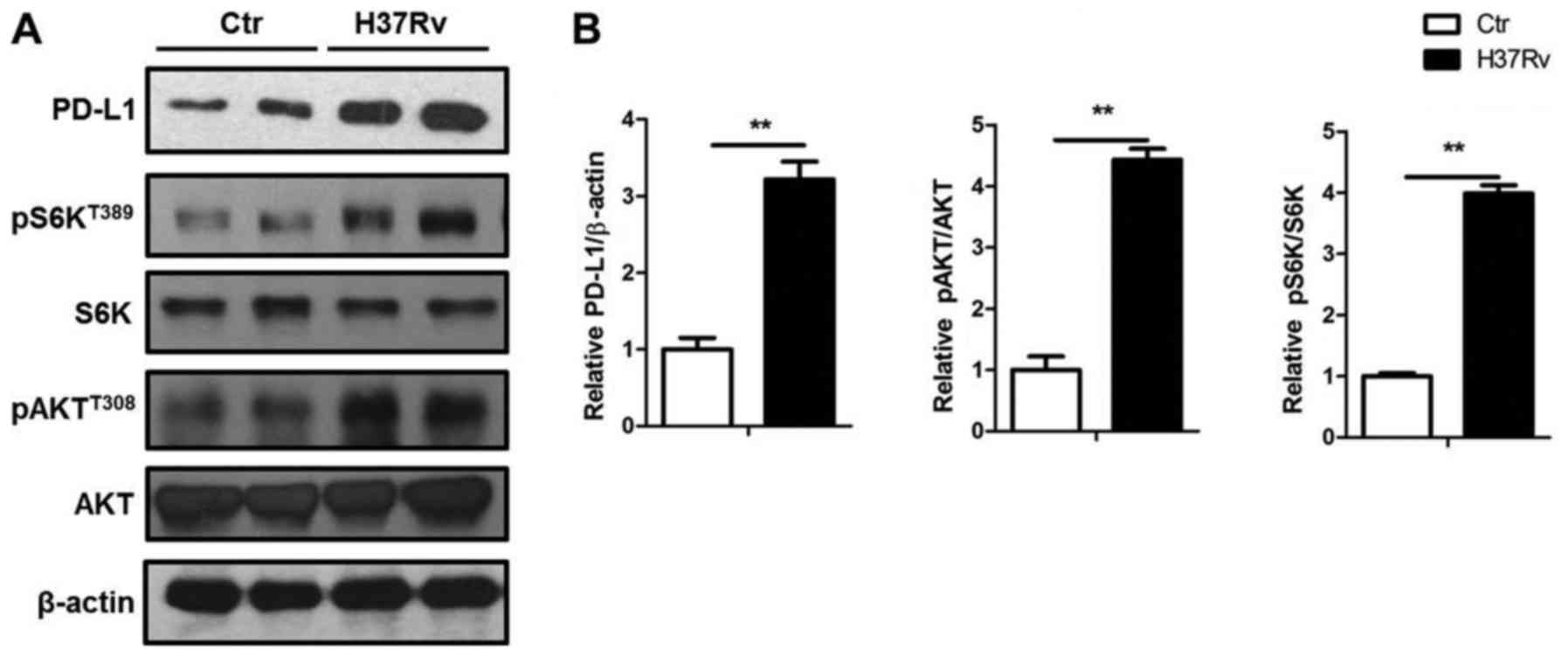
mTORC1 activation contributes to the
PD-L1 expression by H37Rv infection
PD-L1 expression was upregulated by a group of
pro-inflammatory factors such as IL-6 and TNF-α. Also, PD-L1
expression can be downregulated by PTEN (17), which could inactivate the mTORC1
signal. Here, we showed that H37Rv significantly promoted PD-L1
expression with the elevated Akt and S6K phosphorylation, which
both indicated activation of Akt-mTORC1 signal (Fig. 4).
Inhibition of mTORC1 signal attenuates
PD-L1 expression induced by H37Rv stimulation
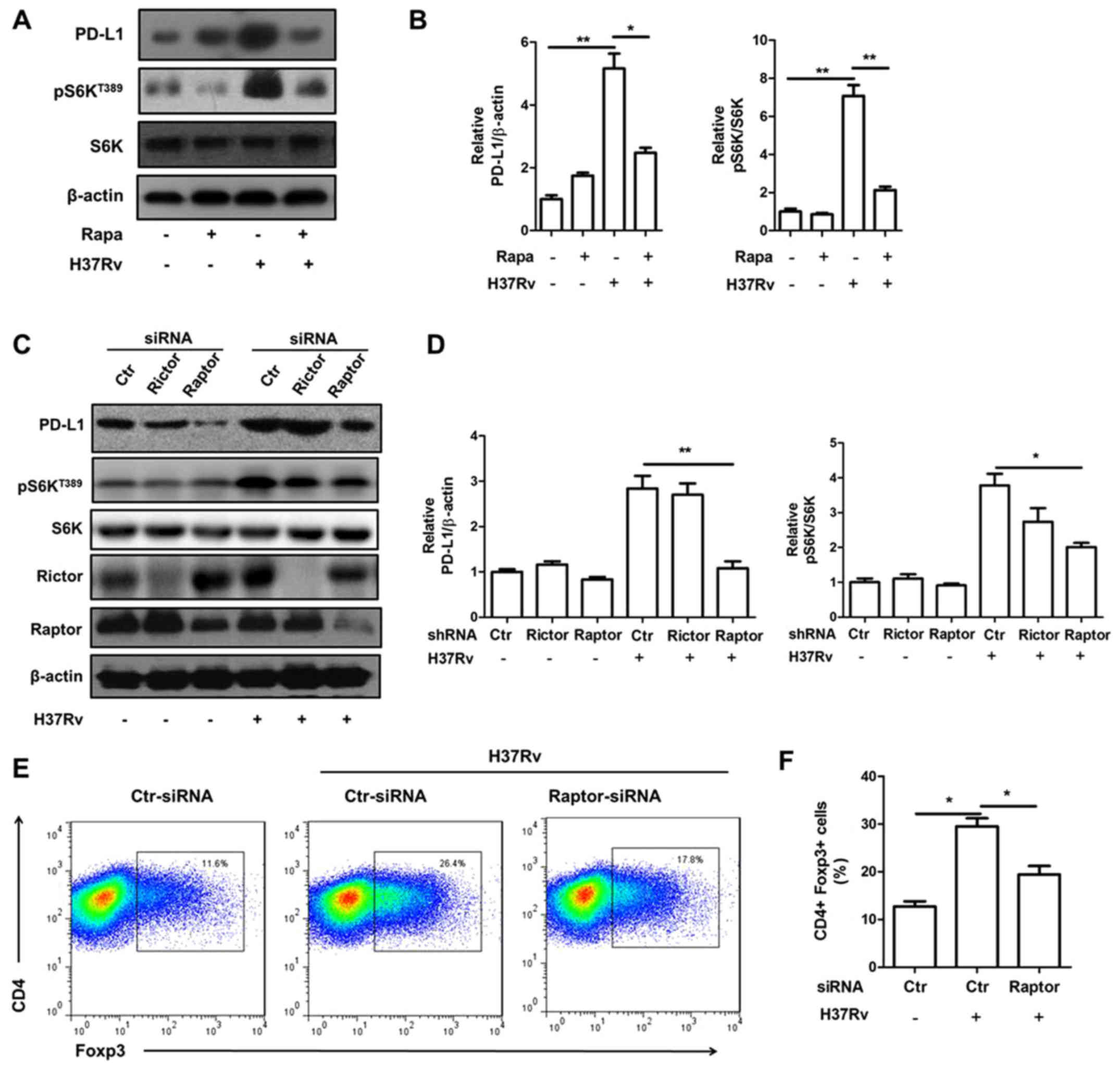
We then tried to determine whether mTORC1 inhibition
will block PD-L1 expression by adopting mTORC1 inhibitor rapamycin.
Rapamycin pre-treatment obviously attenuated PD-L1 expression upon
H37Rv infection (Fig. 5A and B).
Raptor is a specific component protein of mTORC1 while rictor is a
specific component of mTORC2. Knock down of raptor, but not rictor,
significantly lowered mTORC1 activity indicated by S6K
phosphorylation (Fig. 5C and D).
Consistently, raptor, but not rictor, deficiency decreased PD-L1
expression upregulated by H37Rv stimulation (Fig. 5C and D). Moreover, the Treg
promotion ability of macrophages infected by H37Rv was also reduced
with raptor deficiency (Fig.
5E).
mTOR inhibition decreases the ability
of H37Rv infection in tumor acceleration
Considering the critical role of mTORC1 signal in
the PD-L1 promotion and further Treg polarization, we investigated
whether inhibition of mTORC1 could rescue the results of H37Rv
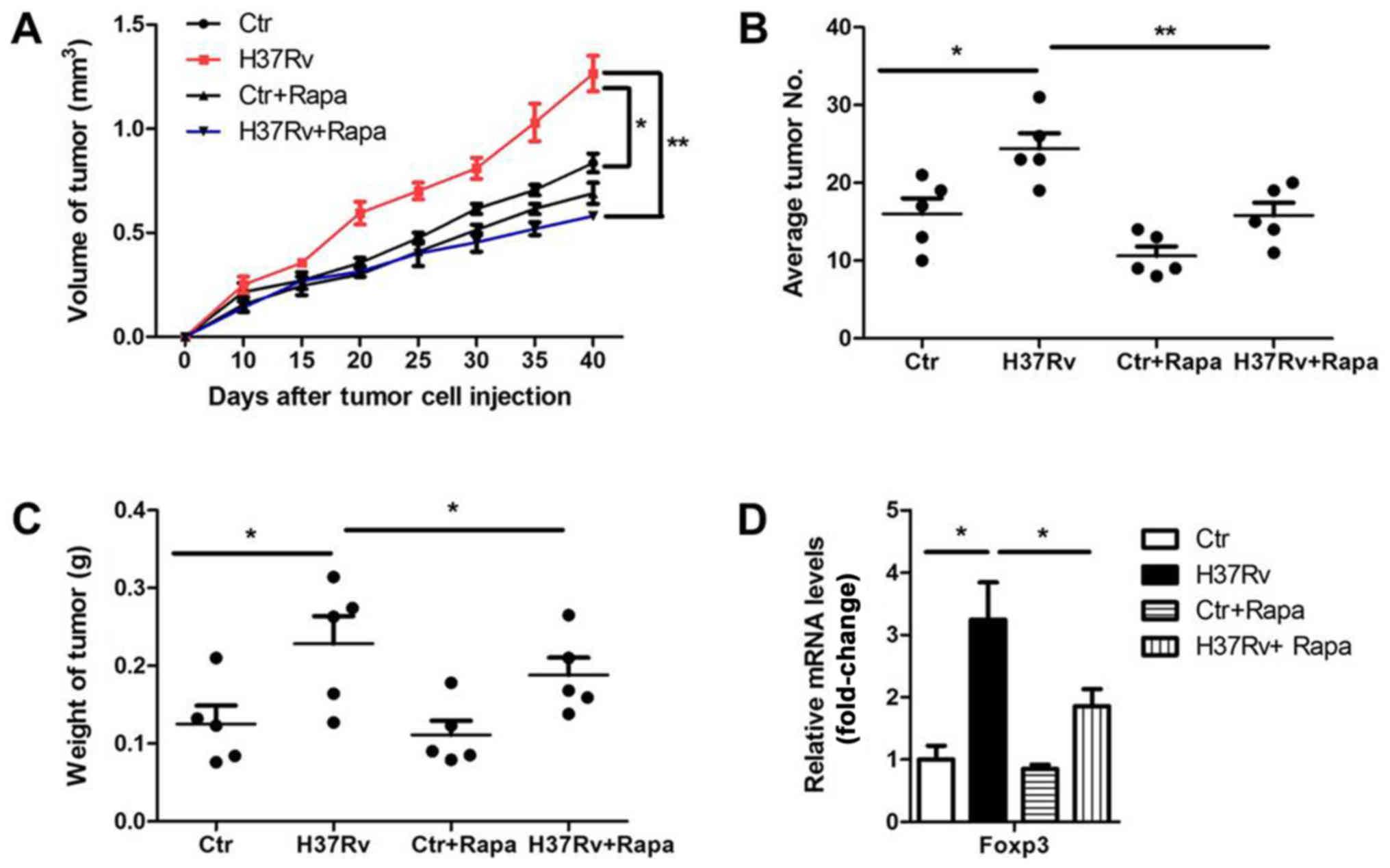
infection in tumor progression. We infected tumor-bearing mice with
H37Rv with or without rapamycin once a week. Tumor volumes were
remarkably suppressed in the rapamycin-treatment group compared to
the non-treatment group after H37Rv-infection, with comparable
levels of non-infection groups (Fig.
6A). Besides, the tumor numbers and tumor weights were also
decreased in the group treated with H37Rv plus rapamycin, as
compared with the H37Rv-infection group (Fig. 6B and C). Rapamycin treatment alone
had a modest effect on tumor volumes, tumor weights and numbers,
but without significant differences (Fig. 6A-C). Accordingly, additional
rapamycin treatment blocked Treg proportion upon H37Rv infection
(Fig. 6D). These results suggest
that mTOCR1 is responsible for tumor acceleration and Treg
promotion after H37Rv infection.
Discussion
As one of the major antigen presenting cells (APC),
macrophage activation delivers costimulation signals that lead to
the activation of CD4+ T cells. CD4+ effector
T cells are a heterogeneous population that can be divided into
multiple subsets defined by their cytokine profiles. Th1 cells,
which express the master transcription factor T-bet and secrete
IFN-γ, is responsible for anti-bacteral and antitumor immunity
(18,19). Tregs, express high levels of CD25
and the transcription factor Foxp3, restrain immune responses
including antitumor immunity (20–22).
The tumor microenvironment is potently immunosuppressive as a
result of inability of effector Th1 cell infiltration and
suppressive Treg accumulation. Therefore, the immunoadjuvant
component is needed to create a proinflammatory milieu, thereby
enhancing costimulation and T lymphocyte activation (23). The ability of various infections to
affect tumor growth has been well determined (24,25).
Immune adjuvants, such as tuberculosis Bacillus Calmette-Guérin
(BCG) and heat-killed Mycobacterium vaccae suspension
(SRL172), have been used as non-specific immunostimulators against
various human cancers with suboptimal clinical response (26–28).
In this study, we intended to determine whether MTB
infection could initiate effective antitumor immune responses. We
first successfully infected mice with H37Rv (data not shown).
Surprisingly, we found that H37Rv infection did not improve but
promoted tumor development. We detected obvious Th1 cytokine
profile that leads to an immune response against tumor cells in
macrophges after H37Rv stimulation. In the in vitro
coculture system, Th1 and Treg proportion both were enhanced with a
high Treg/Th1 ratio after H37Rv infection, which might to some
extent explain the acceleration of tumor progression. Indeed, in
Mycobacterium tuberculosis infection, Tregs proliferate and
accumulate at sites of infection and prevent bacillary clearance in
mice (29,30). MTB promotes regulatory T-cell
expansion via induction of PD-L1 (31–33).
Depletion of Tregs enhances M. tuberculosis-induced IFN-γ
production by peripheral blood mononuclear cells (PBMCs) (9,34).
Although macrophages showed upregulated MHCII, CD80
and CD86 expression upon H37Rv infection, the levels of PD-L1,
which could interacts with the immune receptor programmed cell
death-1 (PD-1) stimulated Foxp3 expression and Treg differentiation
(35), was also markedly enhanced
upon H37Rv stimulation, reflecting a negtive feed back mechanism to
restrain robust inflammation. In fact, a direct correlation between
PD-1 and IFN-γ expression on NK cells was observed upon MTB
infection (33). PD-L1
transcription is regulated by c-Jun and Stat3 (36). Akt-mTORC1 signal which is upstream
of Stat3 has been reported to affect PD-L1 expression in various
cells. Loss of Lkb1 and Pten leads to lung squamous cell carcinoma
with elevated PD-L1 expression (17). In a human-mouse chimeric model of
allograft rejection, rapamycin pretreatment of human arterial
allografts increased graft endothelial cells expression of PD-L1
and PD-L2 and reduced subsequent infiltration of allogeneic
effector T cells into the artery intima and intimal expansion
(37). Oncogenic activation of the
AKT-mTOR pathway promotes immune escape by driving expression of
PD-L1 in syngeneic and genetically engineered mouse models of lung
cancer (38). In our study, we
found that H37Rv infection stimulated mTORC1 activation. Deficiency
of mTORC1 by rapamycin or knockdown of raptor obviously abolished
PD-L1 expression induced by H37Rv. Rapamycin treatment also
reversed the ability of H37Rv in tumor promotion.
Taken together, our results further strengthen that
mTORC1 in macrophages controls Treg promotion and for the first
time pointed out that concomitant H37Rv infection will accelerate
NSCLC progression. This study provides new insights for clinical
treatment of NSCLC.
Acknowledgements
This study was supported by Science and Technology
Commission guidance of Shanghai (no. 124119a6300); International
Science and Technology Cooperation of Shanghai (no. 14430723300);
National Natural Science Foundation of China (no. 81472642);
2014YZDC20700; Shanghai Municipal Education Commision-Gaofeng
Clininal Medicine Grant Support (no. 20161434); YZ2015-ZX12.
References
|
1
|
Paleiron N, Bylicki O, André M, Rivière E,
Grassin F, Robinet G and Chouaïd C: Targeted therapy for localized
non-small-cell lung cancer: A review. Onco Targets Ther.
9:4099–4104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Whiteman DC and Wilson LF: The fractions
of cancer attributable to modifiable factors: A global review.
Cancer Epidemiol. 44:203–221. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hamilton G, Rath B and Ulsperger E: A
review of the role of surgery for small cell lung cancer and the
potential prognostic value of enumeration of circulating tumor
cells. Eur J Surg Oncol. 42:1296–1302. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kuribayashi K, Funaguchi N and Nakano T:
Chemotherapy for advanced non-small cell lung cancer with a focus
on squamous cell carcinoma. J Cancer Res Ther. 12:528–534. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Giaj-Levra N, Ricchetti F and Alongi F:
What is changing in radiotherapy for the treatment of locally
advanced nonsmall cell lung cancer patients? A review. Cancer
Invest. 34:80–93. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen L, He Z, Qin L, Li Q, Shi X, Zhao S,
Chen L, Zhong N and Chen X: Antitumor effect of malaria parasite
infection in a murine Lewis lung cancer model through induction of
innate and adaptive immunity. PLoS One. 6:e244072011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim JO, Jung SS, Kim SY, Kim TY, Shin DW,
Lee JH and Lee YH: Inhibition of Lewis lung carcinoma growth by
Toxoplasma gondii through induction of Th1 immune responses and
inhibition of angiogenesis. J Korean Med Sci. 22:(Suppl). S38–S46.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Martínez A, Torello S and Kolter R:
Sliding motility in mycobacteria. J Bacteriol. 181:7331–7338.
1999.PubMed/NCBI
|
|
9
|
Garg A, Barnes PF, Roy S, Quiroga MF, Wu
S, García VE, Krutzik SR, Weis SE and Vankayalapati R:
Mannose-capped lipoarabinomannan- and prostaglandin E2-dependent
expansion of regulatory T cells in human Mycobacterium
tuberculosis infection. Eur J Immunol. 38:459–469. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gehring AJ, Rojas RE, Canaday DH, Lakey
DL, Harding CV and Boom WH: The Mycobacterium tuberculosis
19-kilodalton lipoprotein inhibits gamma interferon-regulated
HLA-DR and Fc gamma R1 on human macrophages through Toll-like
receptor 2. Infect Immun. 71:4487–4497. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu H, Komai-Koma M, Xu D and Liew FY:
Toll-like receptor 2 signaling modulates the functions of
CD4+ CD25+ regulatory T cells. Proc Natl Acad
Sci USA. 103:7048–7053. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hirsch CS, Rojas R, Wu M and Toossi Z:
Mycobacterium tuberculosis induces expansion of Foxp3
positive CD4 T-cells with a regulatory profile in tuberculin
non-sensitized healthy subjects: Implications for effective
immunization against TB. J Clin Cell Immunol. 7:72016. View Article : Google Scholar
|
|
13
|
He XY, Xiao L, Chen HB, Hao J, Li J, Wang
YJ, He K, Gao Y and Shi BY: T regulatory cells and Th1/Th2
cytokines in peripheral blood from tuberculosis patients. Eur J
Clin Microbiol Infect Dis. 29:643–650. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kuo CH, Lo CY, Chung FT, Lee KY, Lin SM,
Wang CH, Heh CC, Chen HC and Kuo HP: Concomitant active
tuberculosis prolongs survival in non-small cell lung cancer: A
study in a tuberculosis-endemic country. PLoS One. 7:e332262012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen C, Liu Y, Liu Y and Zheng P: mTOR
regulation and therapeutic rejuvenation of aging hematopoietic stem
cells. Sci Signal. 2:ra752009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou H, Wang Y, Lian Q, Yang B, Ma Y, Wu
X, Sun S, Liu Y and Sun B: Differential IL-10 production by DCs
determines the distinct adjuvant effects of LPS and PTX in EAE
induction. Eur J Immunol. 44:1352–1362. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu C, Fillmore CM, Koyama S, Wu H, Zhao Y,
Chen Z, Herter-Sprie GS, Akbay EA, Tchaicha JH, Altabef A, et al:
Loss of Lkb1 and Pten leads to lung squamous cell carcinoma with
elevated PD-L1 expression. Cancer Cell. 25:590–604. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zamarron BF and Chen W: Dual roles of
immune cells and their factors in cancer development and
progression. Int J Biol Sci. 7:651–658. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wieder T, Braumüller H, Kneilling M,
Pichler B and Röcken M: T cell-mediated help against tumors. Cell
Cycle. 7:2974–2977. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sakaguchi S, Miyara M, Costantino CM and
Hafler DA: FOXP3+ regulatory T cells in the human immune
system. Nat Rev Immunol. 10:490–500. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maynard CL, Hatton RD, Helms WS, Oliver
JR, Stephensen CB and Weaver CT: Contrasting roles for all-trans
retinoic acid in TGF-beta-mediated induction of Foxp3 and Il10
genes in developing regulatory T cells. J Exp Med. 206:343–357.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roychoudhuri R, Eil RL and Restifo NP: The
interplay of effector and regulatory T cells in cancer. Curr Opin
Immunol. 33:101–111. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yasumoto K, Hanagiri T and Takenoyama M:
Lung cancer-associated tumor antigens and the present status of
immunotherapy against non-small-cell lung cancer. Gen Thorac
Cardiovasc Surg. 57:449–457. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alexandroff AB, Jackson AM, O'Donnell MA
and James K: BCG immunotherapy of bladder cancer: 20 years on.
Lancet. 353:1689–1694. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hibbs JB Jr, Lambert LH Jr and Remington
JS: Resistance to murine tumors conferred by chronic infection with
intracellular protozoa, Toxoplasma gondii and Besnoitia jellisoni.
J Infect Dis. 124:587–592. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Grange JM, Bottasso O, Stanford CA and
Stanford JL: The use of mycobacterial adjuvant-based agents for
immunotherapy of cancer. Vaccine. 26:4984–4990. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Malmström PU, Wijkström H, Lundholm C,
Wester K, Busch C and Norlén BJ: Swedish-Norwegian Bladder Cancer
Study Group: 5-year followup of a randomized prospective study
comparing mitomycin C and bacillus Calmette-Guerin in patients with
superficial bladder carcinoma. J Urol. 161:1124–1127. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stanford JL, Stanford CA, O'Brien ME and
Grange JM: Successful immunotherapy with Mycobacterium
vaccae in the treatment of adenocarcinoma of the lung. Eur J
Cancer. 44:224–227. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ordway D, Henao-Tamayo M, Harton M,
Palanisamy G, Troudt J, Shanley C, Basaraba RJ and Orme IM: The
hypervirulent Mycobacterium tuberculosis strain HN878
induces a potent TH1 response followed by rapid down-regulation. J
Immunol. 179:522–531. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Scott-Browne JP, Shafiani S, Tucker-Heard
G, Ishida-Tsubota K, Fontenot JD, Rudensky AY, Bevan MJ and Urdahl
KB: Expansion and function of Foxp3-expressing T regulatory cells
during tuberculosis. J Exp Med. 204:2159–2169. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Trinath J, Maddur MS, Kaveri SV, Balaji KN
and Bayry J: Mycobacterium tuberculosis promotes regulatory
T-cell expansion via induction of programmed death-1 ligand 1
(PD-L1, CD274) on dendritic cells. J Infect Dis. 205:694–696. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Periasamy S, Dhiman R, Barnes PF,
Paidipally P, Tvinnereim A, Bandaru A, Valluri VL and Vankayalapati
R: Programmed death 1 and cytokine inducible SH2-containing protein
dependent expansion of regulatory T cells upon stimulation With
Mycobacterium tuberculosis. J Infect Dis. 203:1256–1263.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alvarez IB, Pasquinelli V, Jurado JO,
Abbate E, Musella RM, de la Barrera SS and García VE: Role played
by the programmed death-1-programmed death ligand pathway during
innate immunity against Mycobacterium tuberculosis. J Infect
Dis. 202:524–532. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li L, Lao SH and Wu CY: Increased
frequency of CD4(+)CD25(high) Treg cells inhibit BCG-specific
induction of IFN-gamma by CD4(+) T cells from TB patients.
Tuberculosis (Edinb). 87:526–534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Francisco LM, Salinas VH, Brown KE,
Vanguri VK, Freeman GJ, Kuchroo VK and Sharpe AH: PD-L1 regulates
the development, maintenance, and function of induced regulatory T
cells. J Exp Med. 206:3015–3029. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiang X, Zhou J, Giobbie-Hurder A, Wargo J
and Hodi FS: The activation of MAPK in melanoma cells resistant to
BRAF inhibition promotes PD-L1 expression that is reversible by MEK
and PI3K inhibition. Clin Cancer Res. 19:598–609. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang C, Yi T, Qin L, Maldonado RA, von
Andrian UH, Kulkarni S, Tellides G and Pober JS: Rapamycin-treated
human endothelial cells preferentially activate allogeneic
regulatory T cells. J Clin Invest. 123:1677–1693. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lastwika KJ, Wilson W III, Li QK, Norris
J, Xu H, Ghazarian SR, Kitagawa H, Kawabata S, Taube JM, Yao S, et
al: Control of PD-L1 expression by oncogenic activation of the
AKT-mTOR pathway in non-small cell lung cancer. Cancer Res.
76:227–238. 2016. View Article : Google Scholar : PubMed/NCBI
|