Introduction
Glioma is the most common, aggressive and lethal
type of malignant tumor in the central nervous system (1). Its relapse and mortality rate are
increasing due to the inefficiency of current treatment leading to
a poor prognosis despite the combination of multidisciplinary
therapies including surgical resection, chemotherapy and
radiotherapy (2,3). Despite the diverse therapeutic
efforts, the median survival of glioma patients has not obviously
changed (4). The poor prognosis
highlights the urgent requirement for elucidating the detailed
molecular mechanisms for the development of novel therapeutic tools
against glioma and the identification of diagnostic and prognostic
markers of glioma (5).
As a class of small non-coding RNAs, microRNAs
(miRNAs) target the 3′UTR regions of multiple mRNAs and block
translational process, which leads to gene silence (6). One of the gene clusters targeted by
miRNAs has been confirmed to be associated with tumor growth and
aggressiveness (7). Accumulating
evidence indicates that miR-33a functions as either an oncogene or
tumor suppressor in various types of human cancer. miR-33a was
found to function as a tumor suppressor and to inhibit tumor growth
by repressing Pim-1 (8).
Furthermore, miR-33a regulation by Twist1 restrained
epithelial-to-mesenchymal transition and metastasis in
non-small-cell lung carcinoma (NSCLC) (9). Previous research indicates that
miR-33a expression is inhibited in various cancer types including
NSCLC, gallbladder cancer, hepatocellular carcinoma (HCC), breast
cancer, melanoma and pancreatic cancer (9–16).
However, overexpression of miR-33a has been reported in prostate
cancer (17). Furthermore, miR-33a
was found to contribute to osteosarcoma chemoresistant by reducing
cisplatin-induced cell apoptosis (18). The high expression of miR-33a has
been previously detected in glioblastoma specimens and is
significantly correlated with a poor prognosis (19). miR-33a facilitates the malignant
behaviors of glioma-initiating cells including growth and
self-renewal by targeting protein kinase A (PKA) and Notch
signaling pathways (19). However,
the clinical significance and precise mechanisms underlying the
dysfunction of miR-33a in glioma have not been well investigated in
previous studies.
In this study, miR-33a was revealed as an upstream
regulator of sirtuin 6 (SIRT6) which reversed the malignant
potential and reactive oxygen species (ROS) resistance of glioma.
High levels of miR-33a accompanied by decreased SIRT6 expression
were detected in glioma cells and tissues, compared with these
levels in normal cells and tissues. Further analysis suggested that
restoration of SIRT6 in glioma cells resulted in reduced cell
survival, oxidative stress-induced apoptosis as well as repression
of Janus kinase 2 (JAK2)/signal transducer and activator of
transcription 3 (STAT3) pathways. Therefore, our study demonstrated
that aberrant overexpression of miR-33a disrupts the inhibitory
effect of SITR6 and consequently promotes the malignant phenotypes
of glioma.
Materials and methods
Patients
Glioma specimens and the corresponding normal
tissues were acquired from 60 glioma patients who underwent surgery
at Xi'an Central Hospital, Xi'an Jiaotong University School of
Medicine. Patients who received immunotherapy, chemotherapy or
radiotherapy before surgical treatment were excluded. Signed
informed consent was provided by each patient before clinical
specimens were collected and used. Tissue specimens were conserved
in liquid nitrogen or 10% formalin until use. All
clinicopathological information of the glioma patients is
documented in Table I. The study
was performed following the approval of the Ethics Committee of the
Xi'an Jiaotong University School of Medicine.
 | Table I.Correlation between the
clinicopathological characteristics of the glioma cases and miR-33a
expression. |
Table I.
Correlation between the
clinicopathological characteristics of the glioma cases and miR-33a
expression.
|
|
| miR-33a
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Total n=60 | High n=30 | Low n=30 | P-value |
|---|
| Age (years) |
|
<50 | 28 | 12 | 16 | 0.301 |
|
≥50 | 32 | 18 | 14 |
|
| Sex |
|
Male | 33 | 15 | 18 | 0.436 |
|
Female | 27 | 15 | 12 |
|
| Tumor size
(cm) |
|
<5 | 22 | 7 | 15 | 0.032a |
| ≥5 | 38 | 23 | 15 |
|
| KPS score |
|
<80 | 34 | 14 | 20 | 0.118 |
|
≥80 | 26 | 16 | 10 |
|
| WHO grade |
|
I+II | 20 | 6 | 14 | 0.028a |
|
III+IV | 40 | 24 | 16 |
|
Cell culture and reagents
Human glioma cell lines, U87, T98, A172 and U251,
and a normal human astrocyte (NHA) cell line were purchased from
the Cell Bank of Shanghai Institute of Cell Biology (Chinese
Academy of Medical Science, Shanghai, China) and cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS) (both from Gibco, Grand Island, NY, USA)
at 37°C with 5% CO2.
miR-33a mimic and inhibitor as well as their
corresponding negative control (NC) vectors were purchased from
GenePharma (Shanghai, China). pcDNA3.1-SIRT6 and the empty vector
were obtained from Shanghai Genechem Co., Ltd., (Shanghai, China).
Vectors were transferred into cells using Lipofectamine 2000
(Invitrogen, Carlsbad, CA, USA) on the basis of the manufacturer's
recommendation. H2O2 (30%) was purchased from
Sigma-Aldrich (St. Louis, MO, USA).
Immunohistochemistry
All specimens were fixed in 10% neutral formalin,
embedded in paraffin and cut into 4-µm sections for
immunohistochemical staining. The EnVision™ two-step method was
used (Dako, Hamburg, Germany), as well as the following antibody:
SIRT6 primary antibody (Abcam, Cambridge, MA, USA).
Immunoblotting
Cells were dissociated in RIPA lysis buffer (P0013D;
Beyotime, Haimen, China) and PMSF (ST506) (Beyotime). A Bradford
protein assay kit (P0006; Beyotime) was used to analyzed protein
concentrations, and the proteins were loaded for 10% SDS-PAGE
electrophoresis, and then the proteins after separation were
transferred onto PVDF membranes (Sigma). Then the PVDF membranes
were blocked with 5% skim milk (Guangming, Shanghai, China) and
incubated with the primary antibody at 4°C overnight. Then,
specimens were incubated with a secondary antibody conjugated with
HRP (Cell Signaling Technology, Beverly, MA, USA). β-actin (Santa
Cruz Biotechnology, Santa Cruz, CA, USA) was used as a loading
control. SIRT6 primary antibody was obtained from Abcam. Bax,
Bcl-2, caspase-8, p-JAK2, JAK2, p-STAT3 and STAT3 primary
antibodies were purchased from Cell Signaling Technology.
RNA extraction and quantitative
PCR
qRT-PCR was carried out as previously described
(9). Total RNA was isolated from
clinical tissue samples with a Total RNA isolation kit
(AP-MN-MS-RNA; Axygen, Union City, CA, USA) as described by the
manufacturer. Total RNA from cells was isolated with TRIzol reagent
(Invitrogen) and miRNAs were isolated using a microRNA purification
kit (Norgen Biotek, Thorold Ontario, Canada), according to the
manufacturer's protocol. miRNA-specific quantitative PCR was
performed with Taqman microRNA assay primers (Applied Biosystems,
Foster City, CA, USA) according to the manufacturer's instructions.
The levels of miRNAs expression were normalized by U6 RNA. qPCR for
SIRT6 mRNA was performed with SYBR-Green PCR Master Mix (Applied
Biosystems). β-actin was employed as the internal control. The
primers were synthesized and purchased from Sangon Biotech
(Shanghai, China).
Colony formation assay
In regards to the colony formation assay, 2,000
glioma cells were seeded on 6-well plates. Fourteen to twenty-one
days after cell seeding, cell colonies with crystal violet staining
were counted.
Cell cytotoxicity assay
The indicated cells were seeded on 96-well plates
and cultured for 24 h followed by H2O2 at
different concentrations for 24 h. CCK-8 cell viability and lactate
dehydrogenase (LDH) assay were performed as previously reported
(20). ROS levels were measured as
previously described (21).
Flow cytometric detection of
apoptosis
The indicated cells were collected for detection of
apoptosis using the Annexin V FLUOS kit (Roche, Indianapolis, IN,
USA) according to the manufacturer's instructions. All of the
samples were assayed in triplicate.
Experimental animals
Six-week-old male athymic BALB/c nude mice were
subcutaneously injected with 2×106 U251 cells with NC
inhibitor or miR-33a inhibitor transfection (n=6, respectively).
The tumor volumes were calculated using the standard formula: tumor
volume (mm3) = longer diameter × (shorter
diameter)2/2. The in vivo study was approved by
the Institutional Animal Care and Use Committee of Xi'an Jiaotong
University School of Medicine.
Luciferase reporter assay
Wild-type (wt) or mutant (mt) 3′UTR of SIRT6 was
amplified and cloned into pmiR-RB-REPORT™ luciferase. Luciferase
reporter containing the potential binding sequence of 3′UTR of
SIRT6 was co-transfected with miR-33a mimic or negative control
mimic in U87 cells in a 96-well plate. Two days later,
dual-luciferase reporter assay system (Promega, Madison, WI, USA)
was used to measure the alteration of luciferase. Firefly
luciferase activity was normalized to Renilla luciferase
activity.
Statistical analysis
All statistical analyses were carried out using
GraphPad Prism 5 software (GraphPad Software, Inc., San Diego, CA,
USA). Experimental data are presented as mean ± SEM from at least
three independent experiments. The data were analyzed by the
Chi-squared test, Student's t-test, ANOVA, log-rank test and
Pearson's correlation test. A p-value <0.05 was considered
statistically significant.
Results
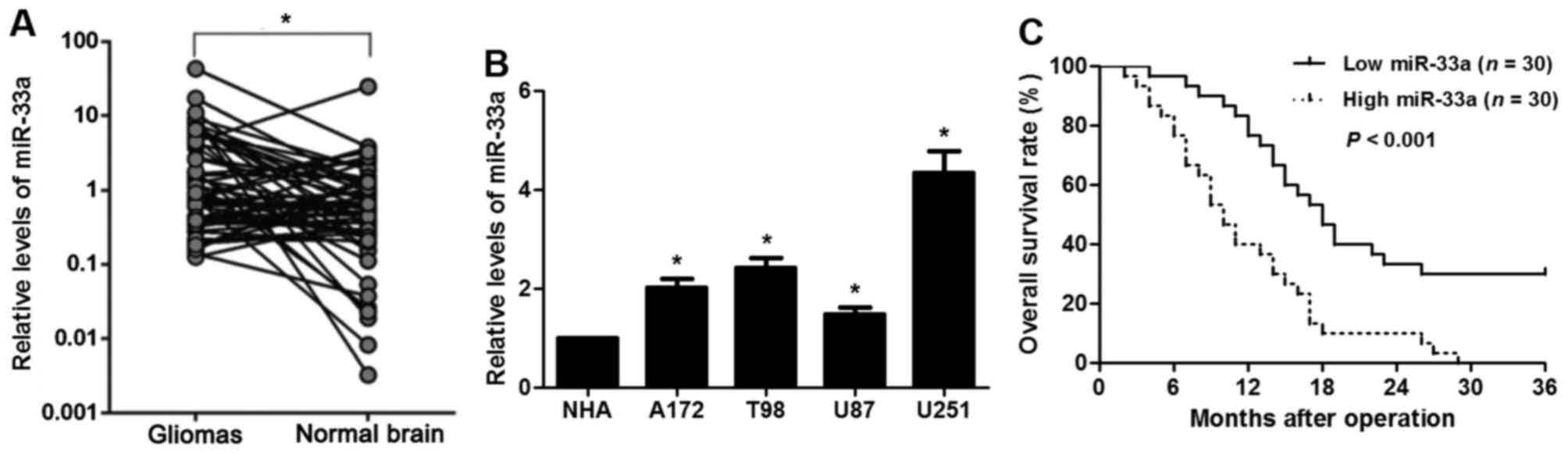
Upregulation of miR-33a is observed in
clinical glioma tissues
Consistent with the dysregulation of miR-33a in
other tumor types, the levels of miR-33a were overexpressed in the
glioma samples when compared with levels in the normal tissues
(P<0.05, Fig. 1A). In addition,
the expression of miR-33a in glioma cell lines (U87, T98, A172 and
U251) was higher than that in a NHA cell line (P<0.05,
respectively, Fig. 1B). Then we
defined the cutoff value between the miR-33a low and high
expression group as the median expression level of miR-33a in
glioma. Further analysis of the clinical specimens showed that high
expression of miR-33a was positively correlated with large tumor
size and advanced World Health Organization (WHO) grade (P<0.05,
respectively) but not with age, sex and Karnofsky performance
status (KPS) score (Table I). In
addition, the 3-year overall survival (OS) rate of the miR-33a high
expression group was significantly lower than that of the miR-33a
low expression group (P<0.001, Fig.
1C). These data suggest that overexpression of miR-33a
indicates a poor prognosis for glioma patients.
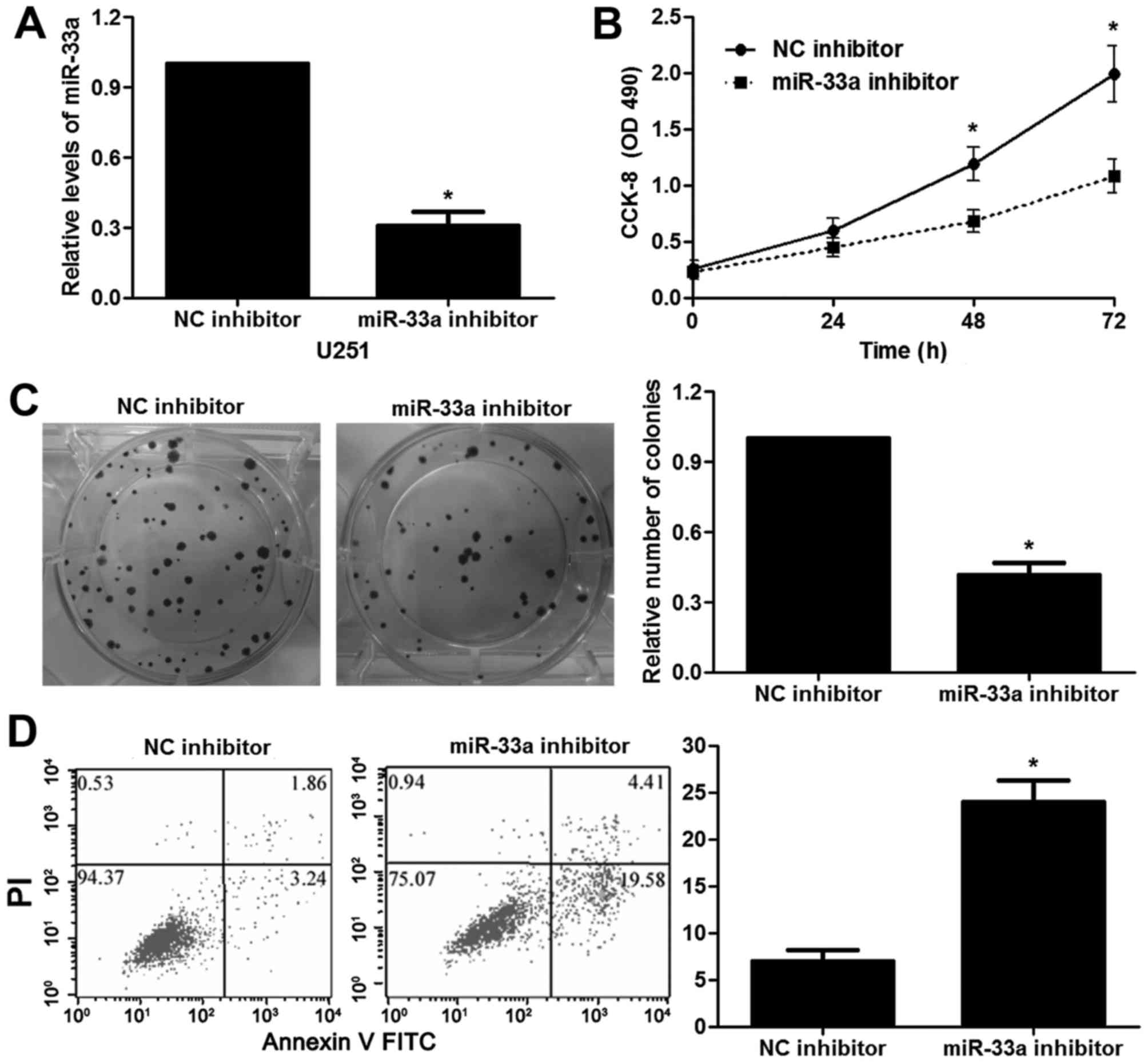
miR-33a regulates the proliferation
and apoptosis of glioma cells
The roles of miR-33a in cancer progression and
metastasis have been discussed in different types of tumor. Thus,
we aimed to elucidate whether silencing of miR-33a prohibits the
malignant phenotypes of glioma. miR-33a inhibitor transfection
resulted in obvious downregulation of miR-33a in the U251 cells
(P<0.05, Fig. 2A). The results
of CCK-8 and colony formation assays showed that U251 cells
transfected with the miR-33a inhibitor presented a reduced cell
proliferation, compared with the control cells (P<0.05,
respectively, Fig. 2B and C). In
addition, stable knockdown of miR-33a induced apoptosis of the U251
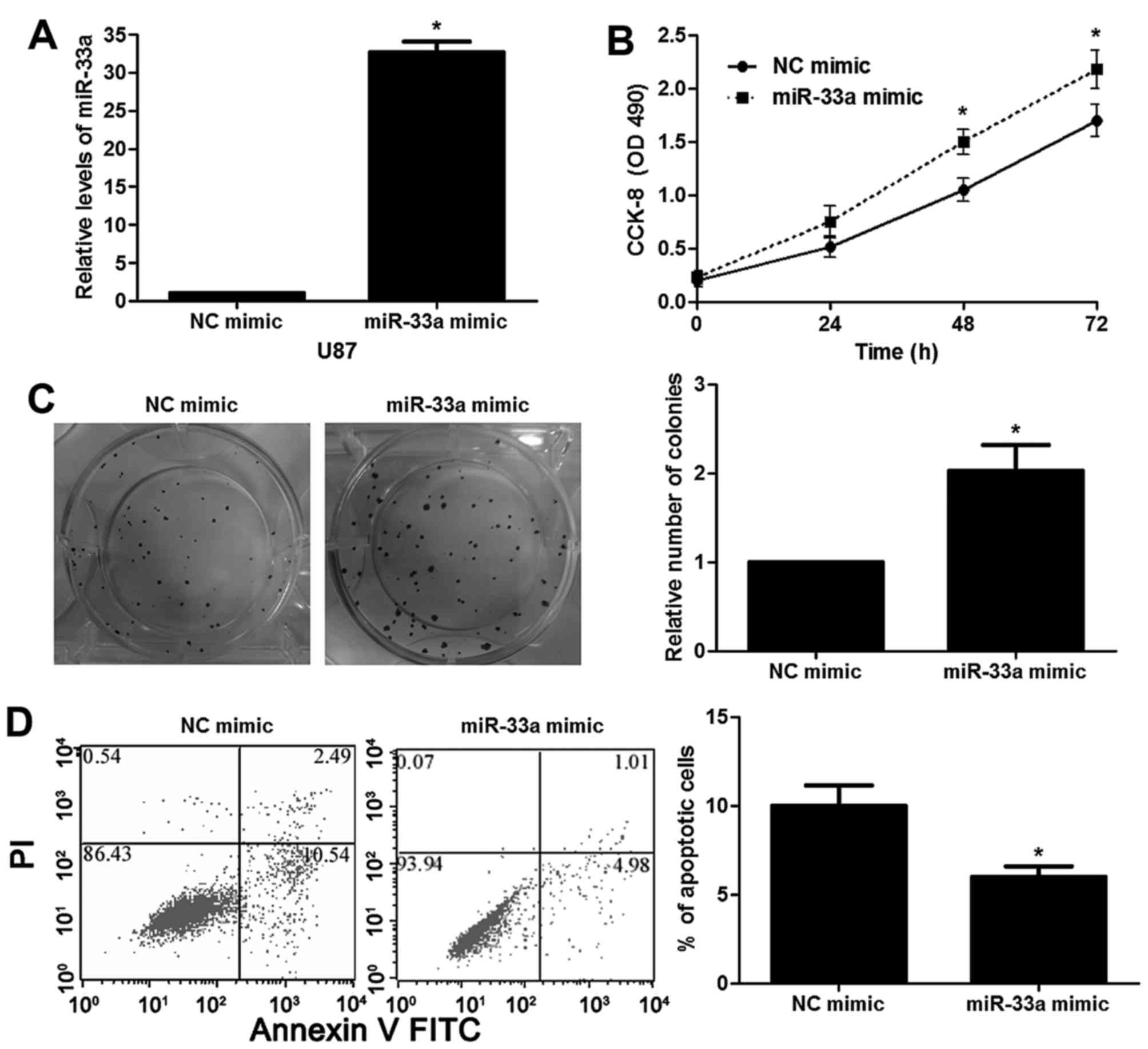
cells (P<0.05, Fig. 2D). In
contrast, the expression of miR-33a was restored by miR-33a mimic
in the U87 cells (P<0.05, Fig.
3A). Notably, miR-33a overexpression enhanced U87 cell growth
and reduced apoptosis in vitro (P<0.05, respectively,
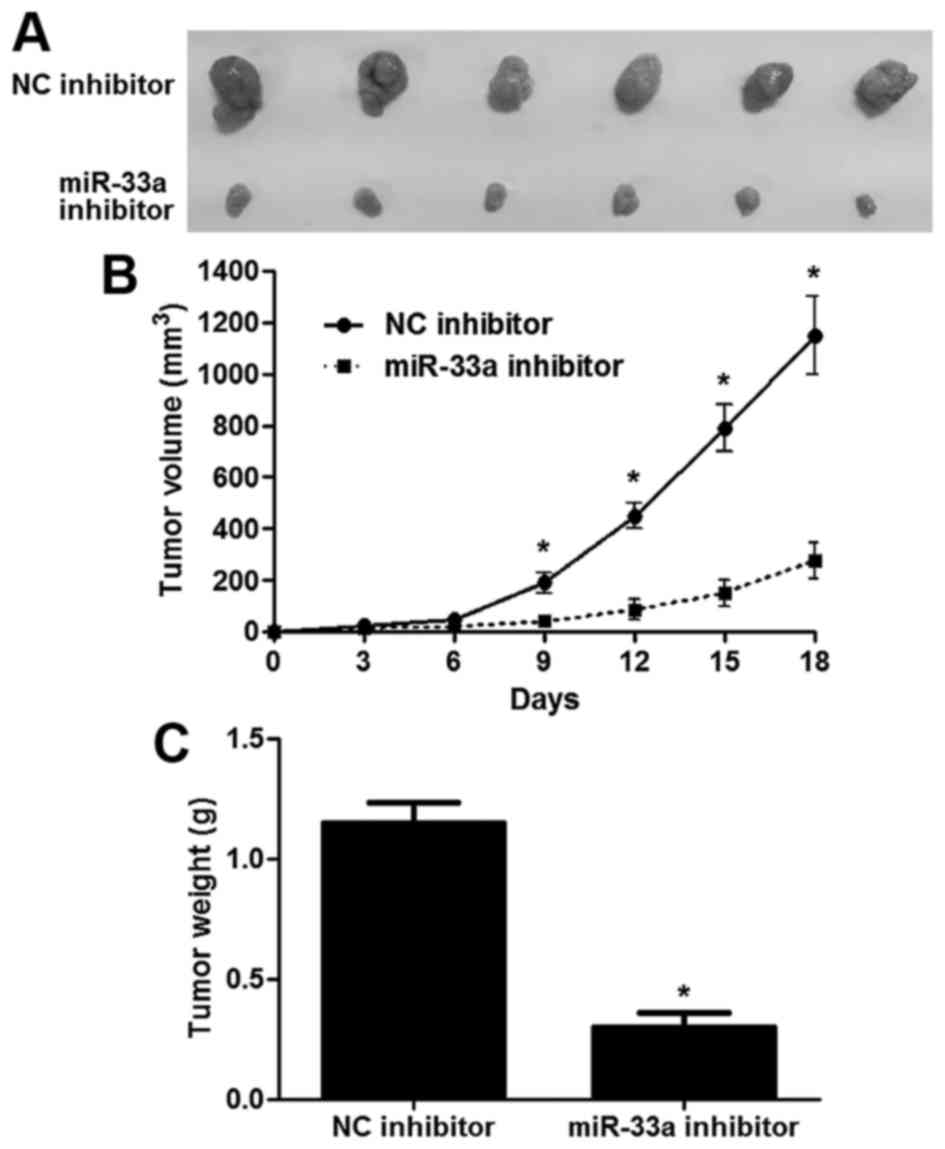
Fig. 3B-D). Next, a subcutaneous
implantation model of human glioma was established in nude mice.
Tumor growth curves and tumor weight revealed that the miR-33a
inhibitor resulted in reduced in vivo growth of glioma cells
compared with the control cells (P<0.05, respectively, Fig. 4A-C). These data indicate that
miR-33a influences the proliferative and apoptotic capabilities of
glioma cells.
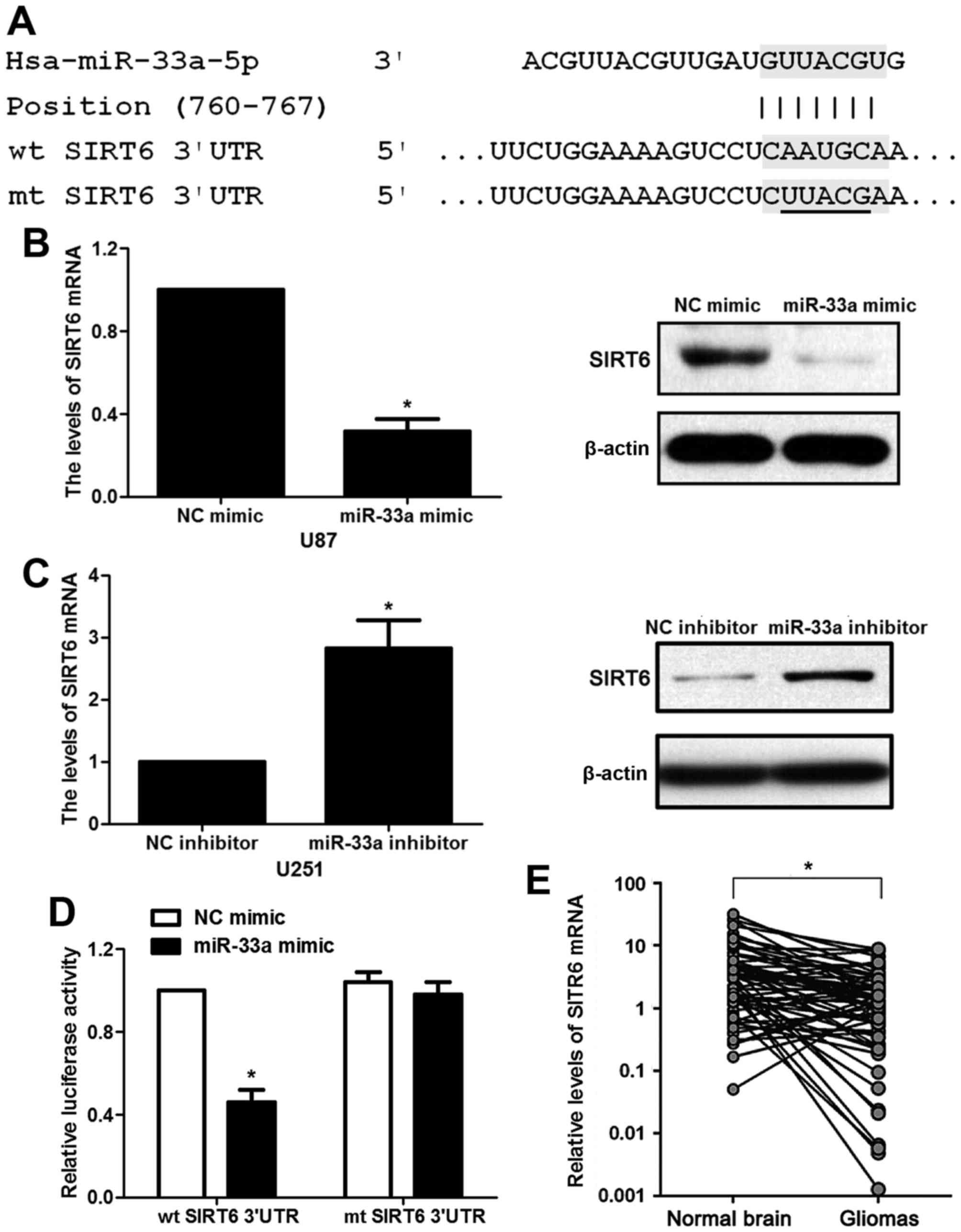
mRNA of SIRT6 is a target of miR-33a
in glioma cells
According to the prediction of bioinformatic
software (Targetscan), SIRT6 was considered as one of the
candidates with which miR-33a could bind directly (Fig. 5A). To experimentally validate the
target prediction, U87 and U251 cells were transfected with miR-33a
mimic or inhibitor. miR-33a mimic was shown to significantly
decrease the expression of SIRT6 at both the mRNA and protein
levels in the U87 cells (P<0.05, Fig. 5B). Consistently, miR-33a inhibitor
displayed a promotive effect on the mRNA and protein levels of
SIRT6 in the U251 cells (P<0.05, Fig. 5C). More importantly, transfection of
miR-33a mimic greatly attenuated the luciferase activity of the wt
SIRT6-3′UTR reporter in the U87 cells while mutation in SIRT6-3′UTR
evidently damaged the response of luciferase activity to miR-33a
mimic (P<0.05, Fig. 5D). Next,
the levels of SIRT6 mRNA were detected by qRT-PCR in glioma and
corresponding normal tissues. Underexpression of SIRT6 mRNA was
found in glioma specimens compared to that noted in the
corresponding normal tissues (P<0.05, Fig. 5E). Spearman's correlation analysis
revealed that the levels of miR-33a were inversely correlated with
SIRT6 mRNA expression in the glioma tissues (r=−0.573, P<0.001).
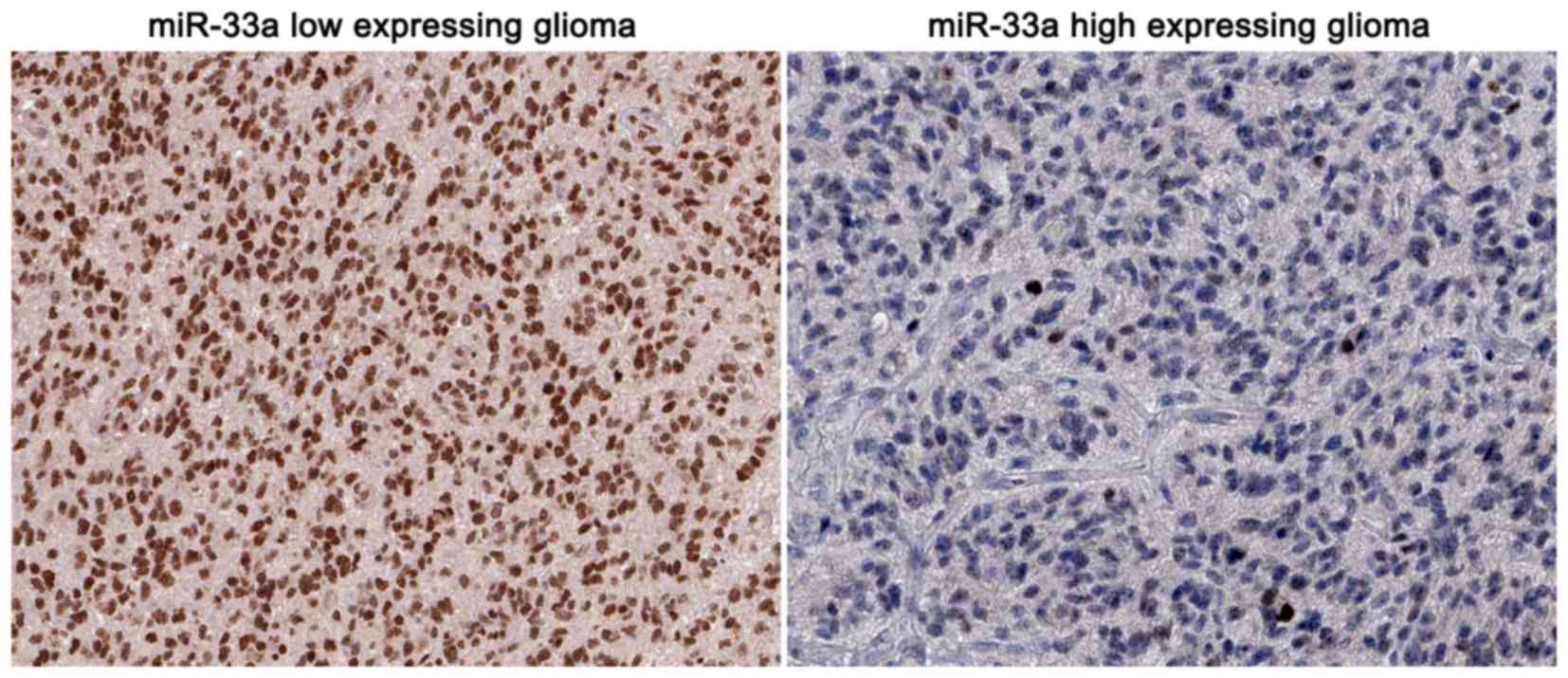
Moreover, representative immunohistochemical staining showed that
miR-33a low-expressing tumors showed strong staining of SIRT6,
while a weak signal of SIRT6 was observed in the miR-33a
high-expressing cases (Fig. 6).
Taken together, miR-33a strongly regulates the expression of SIRT6
by interacting with the 3′UTR region of SIRT6 mRNA in glioma
cells.
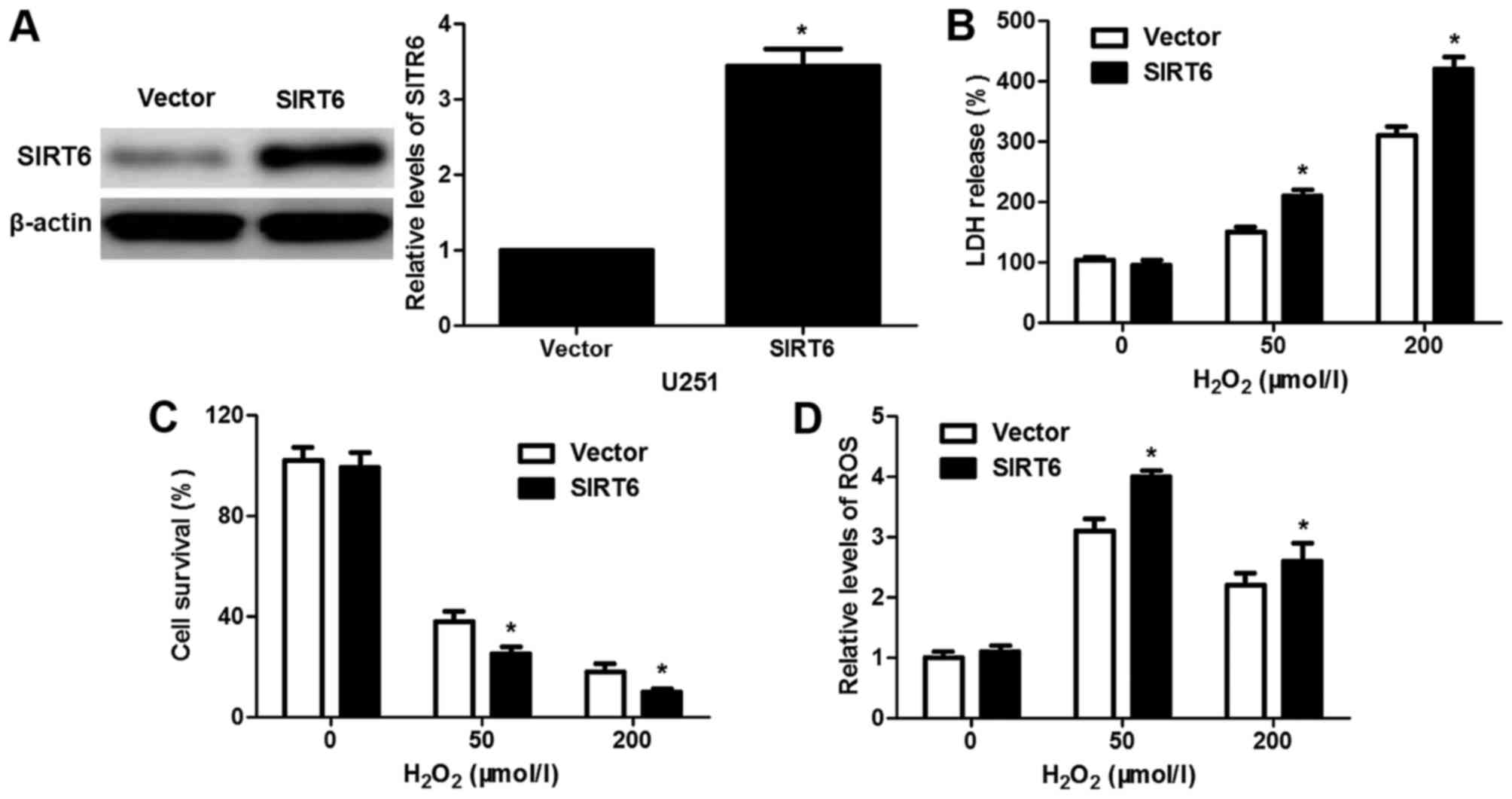
SIRT6 enhances
H2O2-induced oxidative stress and apoptosis
possibly by suppressing the JAK2/STAT3 pathway
Previous research revealed that SIRT6 overexpression
resulted in reduced cell survival and increased ROS production
under H2O2-induced oxidative stress in
neuronal cells (20). To disclose
the potential effect of SIRT6 on H2O2-induced
cancer cell injury, U251 cells that were transfected with
pcDNA3.1-SIRT6 or empty vector were treated with
H2O2 for 1 h and then cultured for 24 h. The
restoration of SIRT6 was confirmed by immunoblotting in the U251
cells (P<0.05, Fig. 7A). Our
results revealed that SIRT6 overexpression increased the levels LDH
and ROS while reducing cell survival compared to the control cells
(P<0.05, respectively, Fig.
7B-D). The Fas ligand/Fas death receptor pathways and
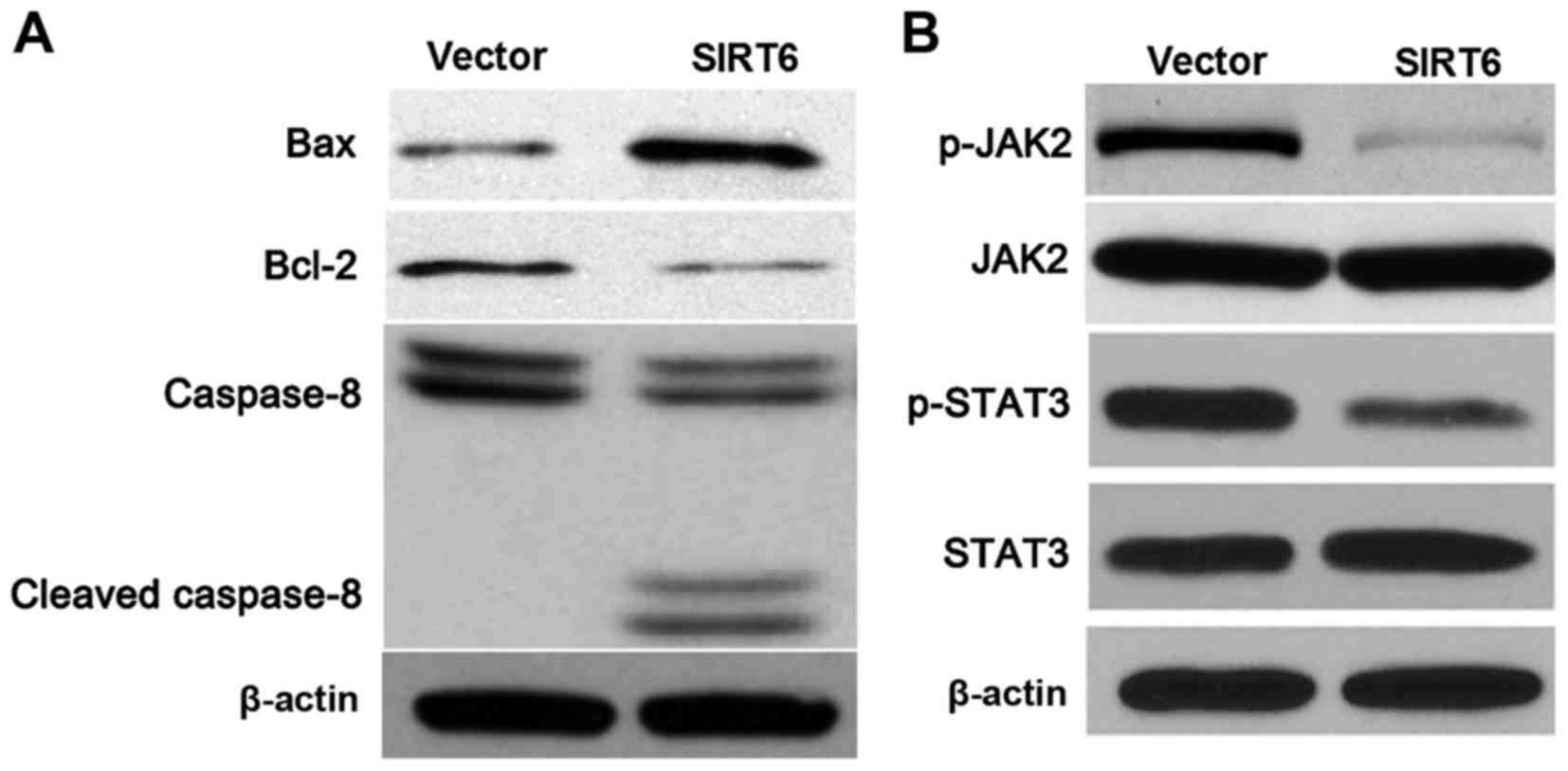
mitochondrial pathway are involved in ROS-induced apoptosis
(22). Next, western blotting
results indicated that SIRT6 overexpression upregulated the levels
of Bax and cleaved caspase-8 while downregulating Bcl-2 expression
in the U251 cells (Fig. 8A).
JAK2/STAT3 pathway was found to enhance cell proliferation and to
prohibit apoptosis, and blocking this pathway increased the
sensitivity to oxidative stress-induced cell injury (23). Our data revealed that SIRT6
restoration repressed the activation of the JAK2/STAT3 pathway with
reduced levels of phosphorylated JAK2 and STAT3 in the U251 cells
(Fig. 8B). Altogether, SIRT6
enhanced oxidative stress-induced cell injury and apoptosis
possibly by inhibition of the JAK2/STAT3 pathway.
Discussion
miRNAs are identified as critical regulators of
cancer growth and metastasis (24,25).
Among numerous miRNAs, miR-33a has been confirmed as a critical
tumor-suppressive miRNA and a prognostic marker of human cancers
including NSCLC (9,26) and pancreatic cancer (16), while, miR-33a exerts an oncogenic
role in prostate cancer (17) and
glioblastoma (19). Here, our data
indicated that overexpression of miR-33a was common in glioma
tissues. In addition, the levels of miR-33a were intensively
elevated in glioma cell lines compared to that noted in the NHA
cells. Glioma patients with a large tumor size and advanced WHO
grade showed prominently higher levels of miR-33a. Furthermore, a
high level of miR-33a was implicated in poor prognosis prediction,
which was consistent with a previous study of glioblastoma reported
by Wang et al (19). Thus,
overexpression of miR-33a may be used as a potential indicator for
the poor prognosis of glioma patients. The roles of miR-33a in the
modulation of cell growth and self-renewal have been reported in
glioma-initiating cells (19). In
our recent study, loss of miR-33a reduced proliferation and
increased apoptosis of U251 cells. The opposite data were obtained
with miR-33a mimic treatment in U87 cells. These data reveal that
miR-33a exerts its oncogenic role by regulating proliferation and
apoptosis in glioma.
SIRT6 has been confirmed as an important tumor
suppressor and regulates the progression and metastasis of
pancreatic cancer (27). SIRT6 is
post-transcriptionally modulated by miR-122 in HCC (28). A previous study reported that
miR-33a/b regulate fatty acid oxidation by targeting SIRT6 in
hepatocytes (29). In this study,
our data from the luciferase reporter assay, qRT-PCR and
immunoblotting demonstrated that SIRT6 is a direct target of
miR-33a in glioma cells. Furthermore, SIRT6 was underexpressed and
was negatively correlated with miR-33a expression in the glioma
specimens. In contrast, upregulation of SIRT6 was reported in NSCLC
and HCC (30,31), which was consistent with the
downregulation of miR-33a in these cancers (9,12). All
these data suggest an inverse correlation between miR-33a and SIRT6
expression. SIRT6 facilitates the sensitivity of neuroblastoma
cells to oxidative stress (20).
Thus, we focused on the correlation between SIRT6 expression and
oxidative stress-induced cell injury in glioma. Notably, SIRT6
restoration increased the levels of LDH and ROS and reduced cell
survival under H2O2 treatment in glioma
cells. A previous study reported that SIRT6 restrained the growth
of glioma cells by inducing apoptosis and repressing the JAK2/STAT3
pathway (23). Here, we showed that
SIRT6 induced apoptosis with increased levels of Bax and cleaved
caspase-8, and decreased expression of Bcl-2 in glioma cells.
Researchers report that ROS activate caspase-8 to induce apoptosis.
In addition, ROS increase the ratio of Bax/Bcl-2 via downregulation
of Bcl-2 and upregulation of Bax and subsequently participate in
the mitochondrial pathways of apoptosis. Furthermore, our data
disclosed that SIRT6 overexpression reduced the activation of the
JAK2/STAT3 pathway, which functions in an oxidative stress
resistant role by promoting proliferation and repressing apoptosis.
Taken together, SIRT6 facilitates oxidative stress-induced growth
arrest and apoptosis by repressing the JAK2/STAT3 pathway in
glioma.
In summary, our findings highlight a miR-33a/SIRT6
pathway, the dysregulation of which leads to the proliferation and
apoptotic resistance of glioma. Furthermore, our study provides a
detailed insight into the mechanism underlying the role of SIRT6 in
glioma. Thus, more research is needed to explore the potential of
miR-33a and SIRT6 as novel therapeutic targets for the treatment of
glioma.
Acknowledgements
This study was supported by the Natural Science
Basic Research Plan in Shaanxi Province of China (program no.
2014JM4139).
References
|
1
|
Awad AJ, Burns TC, Zhang Y and Abounader
R: Targeting MET for glioma therapy. Neurosurg Focus. 37:E102014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katsetos CD, Reginato MJ, Baas PW,
D'Agostino L, Legido A, Ski Tuszyn JA, Dráberová E and Dráber P:
Emerging microtubule targets in glioma therapy. Semin Pediatr
Neurol. 22:49–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klein M: Treatment options and
neurocognitive outcome in patients with diffuse low-grade glioma. J
Neurosurg Sci. 59:383–392. 2015.PubMed/NCBI
|
|
4
|
Kang JH and Adamson C: Novel
chemotherapeutics and other therapies for treating high-grade
glioma. Expert Opin Investig Drugs. 24:1361–1379. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brandner S and von Deimling A: Diagnostic,
prognostic and predictive relevance of molecular markers in
gliomas. Neuropathol Appl Neurobiol. 41:694–720. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang BC and Ma J: Role of microRNAs in
malignant glioma. Chin Med J (Engl). 128:1238–1244. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Thomas M, Lange-Grünweller K, Weirauch U,
Gutsch D, Aigner A, Grünweller A and Hartmann RK: The
proto-oncogene Pim-1 is a target of miR-33a. Oncogene. 31:918–928.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang L, Yang J, Li J, Shen X, Le Y, Zhou
C, Wang S, Zhang S, Xu D and Gong Z: MircoRNA-33a inhibits
epithelial-to-mesenchymal transition and metastasis and could be a
prognostic marker in non-small cell lung cancer. Sci Rep.
5:136772015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Du M, Zhang Y, Mao Y, Mou J, Zhao J, Xue
Q, Wang D, Huang J, Gao S and Gao Y: miR-33a suppresses
proliferation of NSCLC cells via targeting METTL3 mRNA. Biochem
Biophys Res Commun. 482:582–589. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang M, Gong W, Zuo B, Chu B, Tang Z,
Zhang Y, Yang Y, Zhou D, Weng M, Qin Y, et al: The microRNA miR-33a
suppresses IL-6-induced tumor progression by binding Twist in
gallbladder cancer. Oncotarget. 7:78640–78652. 2016.PubMed/NCBI
|
|
12
|
Han SY, Han HB, Tian XY, Sun H, Xue D,
Zhao C, Jiang ST, He XR, Zheng WX, Wang J, et al: MicroRNA-33a-3p
suppresses cell migration and invasion by directly targeting PBX3
in human hepatocellular carcinoma. Oncotarget. 7:42461–42473.
2016.PubMed/NCBI
|
|
13
|
Zhang C, Zhang Y, Ding W, Lin Y, Huang Z
and Luo Q: miR-33a suppresses breast cancer cell proliferation and
metastasis by targeting ADAM9 and ROS1. Protein Cell. 6:881–889.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou J, Xu D, Xie H, Tang J, Liu R, Li J,
Wang S, Chen X, Su J, Zhou X, et al: miR-33a functions as a tumor
suppressor in melanoma by targeting HIF-1α. Cancer Biol Ther.
16:846–855. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wolfe AR, Bambhroliya A, Reddy JP, Debeb
BG, Huo L, Larson R, Li L, Ueno NT and Woodward WA: miR-33a
decreases high-density lipoprotein-induced radiation sensitivity in
breast cancer. Int J Radiat Oncol Biol Phys. 95:791–799. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang C, Yu XJ, Guo XZ, Sun MH, Wang Z,
Song Y, Ni QX, Li HY, Mukaida N and Li YY: MicroRNA-33a-mediated
downregulation of Pim-3 kinase expression renders human pancreatic
cancer cells sensitivity to gemcitabine. Oncotarget. 6:14440–14455.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Q, Lu S, Li X, Hou G, Yan L, Zhang W
and Qiao B: Biological function and mechanism of miR-33a in
prostate cancer survival and metastasis: via downregulating
Engrailed-2. Clin Transl Oncol. 19:562–570. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou Y, Huang Z, Wu S, Zang X, Liu M and
Shi J: miR-33a is up-regulated in chemoresistant osteosarcoma and
promotes osteosarcoma cell resistance to cisplatin by
down-regulating TWIST. J Exp Clin Cancer Res. 33:122014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang H, Sun T, Hu J, Zhang R, Rao Y, Wang
S, Chen R, McLendon RE, Friedman AH, Keir ST, et al: miR-33a
promotes glioma-initiating cell self-renewal via PKA and NOTCH
pathways. J Clin Invest. 124:4489–4502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shao J, Yang X, Liu T, Zhang T, Xie QR and
Xia W: Autophagy induction by SIRT6 is involved in oxidative
stress-induced neuronal damage. Protein Cell. 7:281–290. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song J, Ke SF, Zhou CC, Zhang SL, Guan YF,
Xu TY, Sheng CQ, Wang P and Miao CY: Nicotinamide
phosphoribosyltransferase is required for the calorie
restriction-mediated improvements in oxidative stress,
mitochondrial biogenesis, and metabolic adaptation. J Gerontol A
Biol Sci Med Sci. 69:44–57. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jeong CH and Joo SH: Downregulation of
reactive oxygen species in apoptosis. J Cancer Prev. 21:13–20.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Feng J, Yan PF, Zhao HY, Zhang FC, Zhao WH
and Feng M: SIRT6 suppresses glioma cell growth via induction of
apoptosis, inhibition of oxidative stress and suppression of
JAK2/STAT3 signaling pathway activation. Oncol Rep. 35:1395–1402.
2016.PubMed/NCBI
|
|
24
|
Jansson MD and Lund AH: MicroRNA and
cancer. Mol Oncol. 6:590–610. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nugent M: microRNA and Bone Cancer. Adv
Exp Med Biol. 889:201–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kuo PL, Liao SH, Hung JY, Huang MS and Hsu
YL: MicroRNA-33a functions as a bone metastasis suppressor in lung
cancer by targeting parathyroid hormone related protein. Biochim
Biophys Acta. 1830:3756–3766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kugel S, Sebastián C, Fitamant J, Ross KN,
Saha SK, Jain E, Gladden A, Arora KS, Kato Y, Rivera MN, et al:
SIRT6 suppresses pancreatic cancer through control of Lin28b. Cell.
165:1401–1415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Elhanati S, Ben-Hamo R, Kanfi Y, Varvak A,
Glazz R, Lerrer B, Efroni S and Cohen HY: Reciprocal regulation
between SIRT6 and miR-122 controls liver metabolism and predicts
hepatocarcinoma prognosis. Cell Reports. 14:234–242. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dávalos A, Goedeke L, Smibert P, Ramírez
CM, Warrier NP, Andreo U, Cirera-Salinas D, Rayner K, Suresh U,
Pastor-Pareja JC, et al: miR-33a/b contribute to the regulation of
fatty acid metabolism and insulin signaling. Proc Natl Acad Sci
USA. 108:9232–9237. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee N, Ryu HG, Kwon JH, Kim DK, Kim SR,
Wang HJ, Kim KT and Choi KY: SIRT6 depletion suppresses tumor
growth by promoting cellular senescence induced by DNA damage in
HCC. PLoS One. 11:e01658352016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bai L, Lin G, Sun L, Liu Y, Huang X, Cao
C, Guo Y and Xie C: Upregulation of SIRT6 predicts poor prognosis
and promotes metastasis of non-small cell lung cancer via the
ERK1/2/MMP9 pathway. Oncotarget. 7:40377–40386. 2016.PubMed/NCBI
|