Introduction
Cigarette smoking is strongly associated with lung
cancer incidence and mortality (1,2). As an
important toxic component of cigarette smoke (3), benzo(a)pyrene (BaP) alone is
sufficient to induce lung cancer (4, 5).
However, BaP promotes lung cancer through different mechanisms
(6–10), and one of these mechanisms is the
induction of epithelial-mesenchymal transition (EMT) (10).
EMT is a process by which epithelial cells gain
migratory and invasive characteristics to become motile mesenchymal
stem cells. In physiological conditions, EMT is integral in embryo
implantation, organ development and wound healing (11,12).
However, under pathological conditions, EMT contributes to fibrosis
and cancer progression (11,12).
It is an important process in the development and progression of
lung cancer induced by tobacco smoke (13), and it is an important cause of drug
resistance and metastasis in lung cancer (14). In addition, the EMT secretory
phenotype can also predict survival in patients with lung cancer
(15).
Twist family BHLH transcription factor 1 (Twist1) is
a basic helix-loop-helix domain-containing transcription factor
that is encoded by the Twist1 gene (16), and is essential for embryonic
differentiation during physiological conditions (16). Upregulated Twist1 expression is
associated with the progression of lung cancer (17) and other types of cancer, such as
gastric (18), breast (19), colorectal (20), endometrial (21) and prostate (22) cancer. In addition, Twist1 can induce
EMT in cancer cells (23,24). Particularly, inhibiting the
expression of Twist1 in lung cancer cells suppresses cell
proliferation (17,25) and metastasis (25), activates oncogene-induced senescence
(26) and suppresses the EMT
process (25).
In the present study, we aimed to study the effects
of BaP on migration ability, the EMT process and Twist1 expression
in adenocarcinoma A549 cells. We found that BaP can enhance the
migration of lung adenocarcinoma A549 cells through the promotion
of EMT by upregulation of Twist1.
Materials and methods
Cell culture and treatments
Human A549 cells (Xiangya Cells Center, Central
South University, Changsha, Hunan, China) were cultured in
RPMI-1640 medium (HyClone, Logan, UT, USA) containing 10% fetal
bovine serum (FBS) (Biological Industries, Beit Haemek, Israel) and
1% penicillin/streptomycin and incubated in a humidified incubator
at 37°C with 5% CO2.
BaP (purity ≥96%; Sigma-Aldrich, St. Louis, MO, USA)
was dissolved in dimethyl sulfoxide (DMSO) and added to the culture
medium at a final concentration of 1 µM. Before proceeding with
further experiments, A549 cells were treated with 1 µM BaP for 24,
48 and 72 h; 1, 2 and 4 weeks.
Wound healing assay
Wound healing assays were conducted to evaluate the
lateral migration capacity of cells. After cells were seeded into
24-well plates and had grown into a monolayer, a ‘scratch’ was
scraped in a straight line using a p200 pipet tip. The cells were
washed 3 times with growth medium to smooth the edges of the
scratch and remove debris, and the medium was replaced with fresh
culture medium. The cells were then allowed to recover for 12 h.
Before and after the 12 h incubation, the initial scratch and the
recovery area, respectively, were photographed under a microscope.
The wound closure rate was equal to the recovered distance divided
by the original width of the scratch.
Transwell migration assay
Transwell plates (6.5 mm in diameter at the lower
surface and 8-µm pore filters; Corning Costar, Cambridge, MA, USA)
without Matrigel were used to detect the longitudinal migration
ability of cells. Briefly, 5×104 cells in 200 µl of 0.1%
FBS culture medium were seeded in the upper chamber and 800 µl of
medium with 10% FBS was placed in the lower chamber. Following
incubation for 24 h, a cotton swab was used to remove the cells
adhering to the membrane of the upper chamber. The migrated cells
on the lower side of the filter membrane surface were stained with
0.1% crystal violet, and the cells were counted under a light
microscope.
Reverse transcription PCR
(RT-PCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA), and cDNA was synthesized using a
PrimeScript First Strand cDNA Synthesis kit (Takara Biotech,
Kusatsu, Shiga, Japan) according to the manufacturer's
instructions. The following are the primers used for PCR
amplification: Twist1 forward, 5′-CTCAGCTACGCCTTCTCGGT-3′ and
reverse, 5′-AGTCCATAGTGATGCCTTTC-3′; N-cadherin forward,
5′-ATCCAGACCGACCCAAACAG-3 and reverse, 5′-GGAATTCCATTGTCAGAAGC-3;
vimentin forward, 5′-CCAGGCAAAGCAGGAGTC-3 and reverse,
5′-CGAAGGTGACGAGCCATT-3; E-cadherin forward,
5′-CTGAGAACGAGGCTAACGTCG-3 and reverse, 5′-AGTGTAGGATGTGATTTCCTG-3;
β-actin forward, 5′-AGAGCTACGAGCTGCCTGAC-3 and reverse,
5′-AGCACTGTGTTGGCGTACAG-3.
Western blotting
Whole cell protein was prepared using ice-cold RIPA
buffer (Millipore, Temecula, CA, USA) mixed with a protease
inhibitor cocktail. The protein concentrations were determined
after cell lysates were centrifuged at 12,000 × g at 4°C for 20 min
to clear the cell debris. Equal amounts of protein (50 µg) from
each sample were loaded/lane and separated on 10% SDS-PAGE,
followed by transfer to polyvinylidene difluoride (PVDF) membranes
(Millipore) via electroblotting. Then, 5% non-fat milk was used to
block the membranes for 2 h. Primary antibodies, diluted according
to the manufacturer's recommendations, were incubated with the
membranes overnight at 4°C. The antibodies used included antibodies
against Twist1 (Santa Cruz, Heidelberg, Germany), N-cadherin (BD
Biosciences, San Jose, CA, USA), vimentin (Santa Cruz) and
E-cadherin (BD Biosciences), and β-actin (Cell Signaling
Technology, Beverly, MA, USA) served as the loading control. The
membranes were washed 3 times (10 min/time) in Tris-buffered saline
with Tween-20 (TBST) buffer, followed by incubation with
horseradish peroxidase-conjugated secondary antibodies for 2 h at
room temperature. The protein bands were detected using an enhanced
chemiluminescence (ECL) reagent kit (Thermo Scientific Pierce,
Rockford, IL, USA).
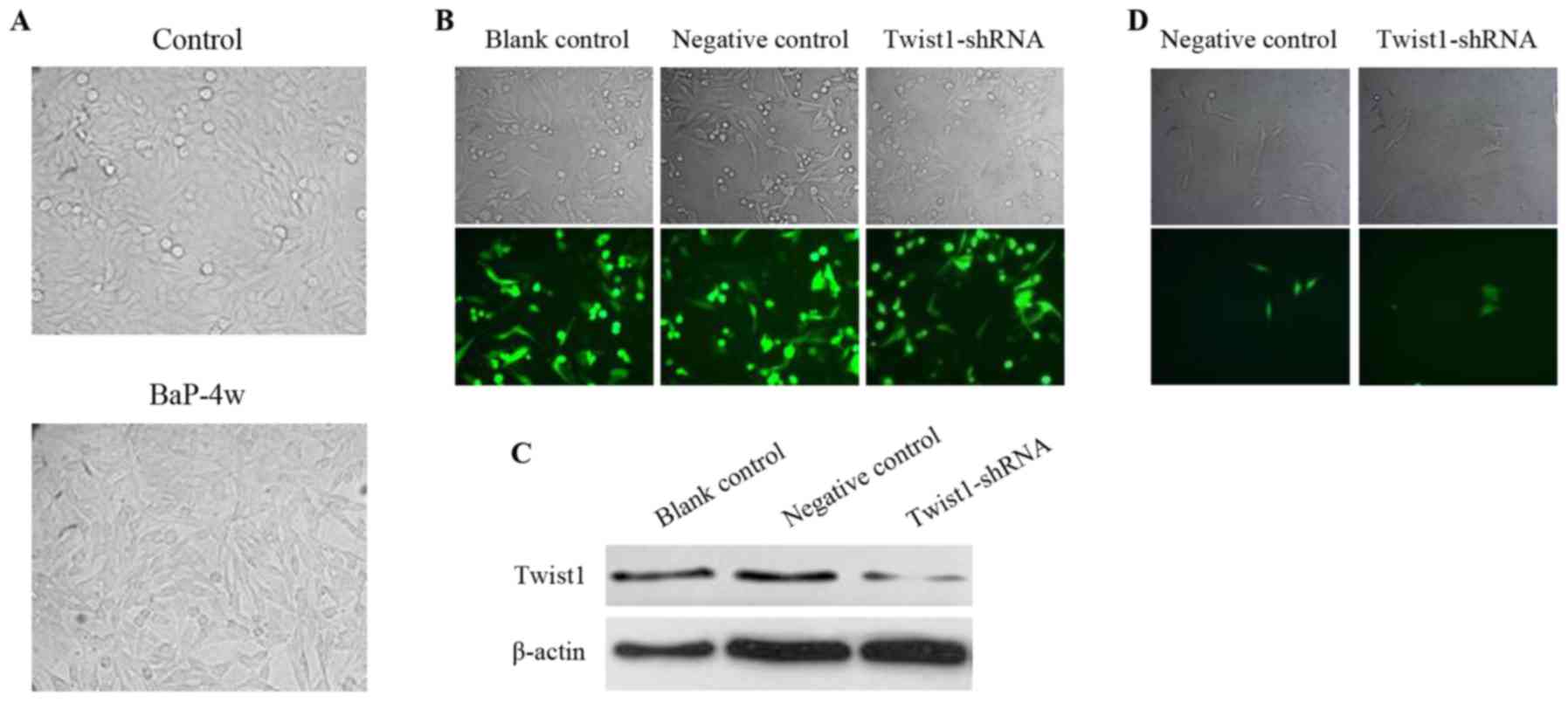
Construction of plasmid-expressing
short hairpin RNA targeting Twist1
A short hairpin RNA sequence (GCTGAGC
AAGATTCAGACCCTTTCAAGAGAAGGGTCTGAATCT TGCTCAGC) targeting Twist1
(Twist1-shRNA) was inserted into a pGCsilencer™ U6/Neo/GFP/RNAi
plasmid (GeneChem, Shanghai, China). A negative control shRNA
(TTCTCCGAAC GTGTCACGT) vector was also constructed using a
pGCsilencer™ U6/Neo/GFP/RNAi plasmid to verify the sequence
specificity of Twist1-shRNA. The cells were transfected with
plasmids using Lipofectamine™ 2000 reagent (Invitrogen). The
transfection efficiency was observed under a fluorescence
microscope after transfection for 24 h. The knockdown efficiency of
Twist1-shRNA was detected with western blotting after transfection
for 48 h.
Statistical analysis
Statistical analyses were conducted using SPSS 18.0
software (SPSS, Inc., Chicago, IL, USA) with a Windows operating
system. Repeated measurement data were presented as the mean ±
standard deviation (SD). When two sets of data were compared,
differences were calculated using Student's t-test (two-tailed).
Statistical significance was defined as p<0.05.
Results
BaP promotes the migration of A549
cells
To study the effect of BaP on the migration of A549
cells, we first treated A549 cells with 1 µM BaP for different
lengths of time (24, 48 and 72 h; 1, 2 and 4 weeks). Thereafter, we
used a wound healing assay to study the lateral migration ability
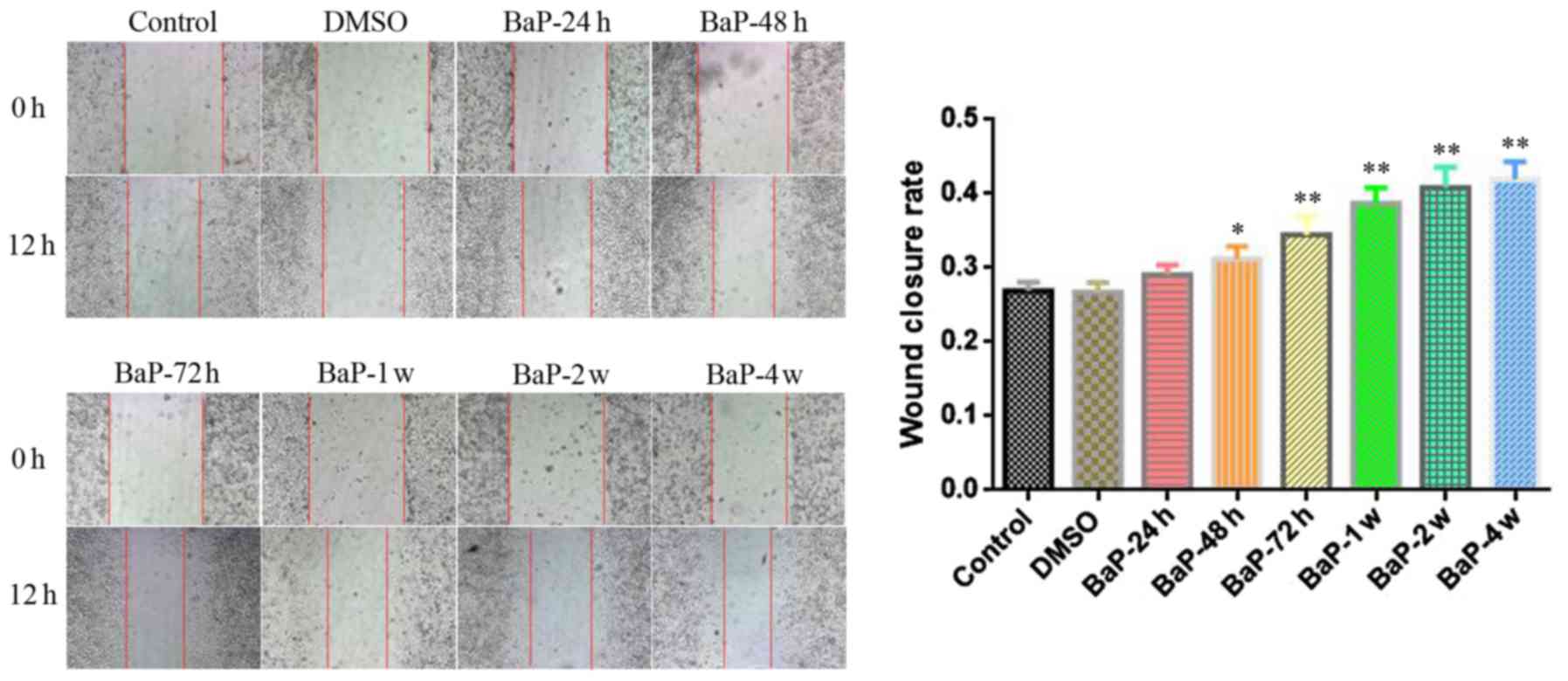
of BaP-treated A549 cells. As shown in Fig. 1, after the cells in the wound
healing assay were allowed to recover for 12 h, BaP enhanced the
lateral migration ability of A549 cells when the treatment time was
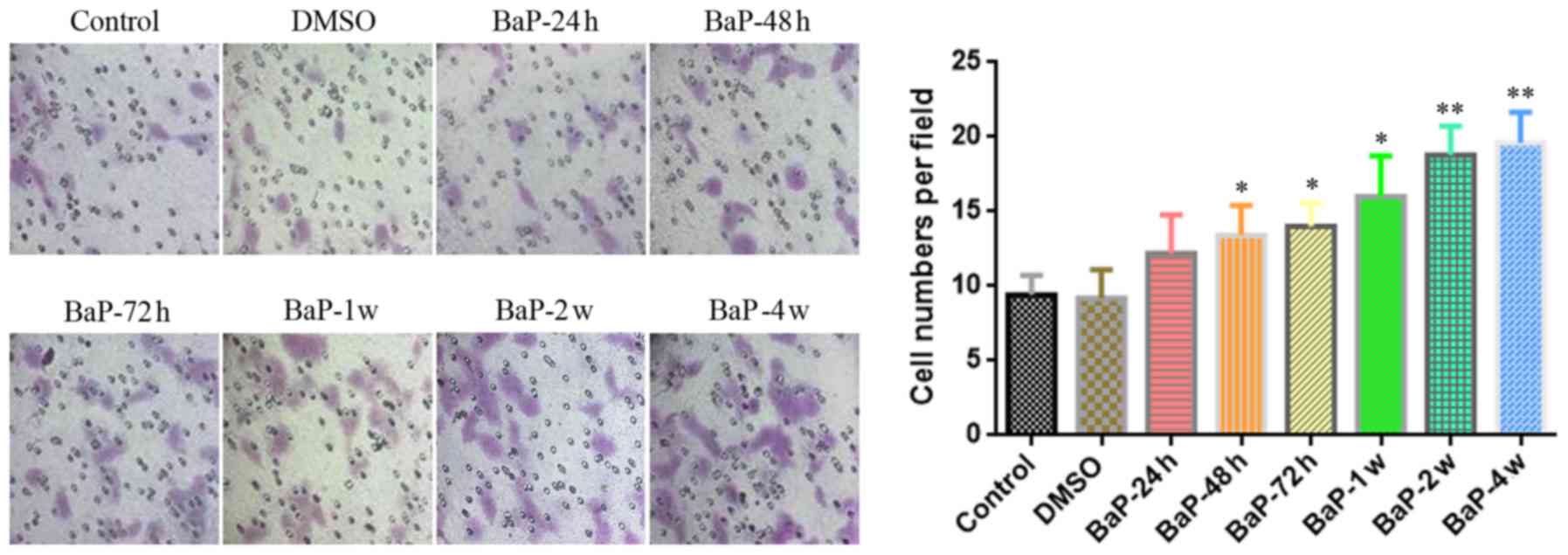
at least 48 h. Furthermore, we used Transwell migration assays to
research the longitudinal migration ability of BaP-treated A549
cells. As shown in Fig. 2, after
the Transwell migration assay was performed for 24 h, BaP enhanced
the longitudinal migration ability of A549 cells when the
intervention time was at least 48 h, similar to the enhancement
effect observed on lateral migration. In addition, we found that
the effect of BaP on the migration of A549 cells was gradually
enhanced with prolonged treatment time (Figs. 1 and 2).
Effect of BaP treatment on the
expression of Twist1, N-cadherin, vimentin and E-cadherin in A549
cells
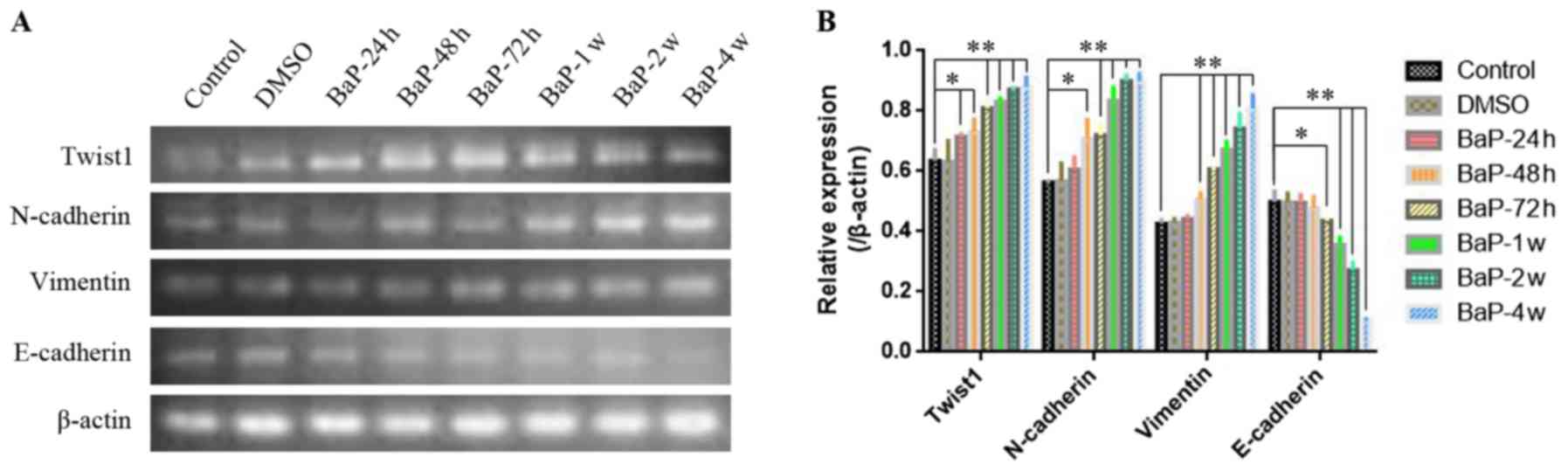
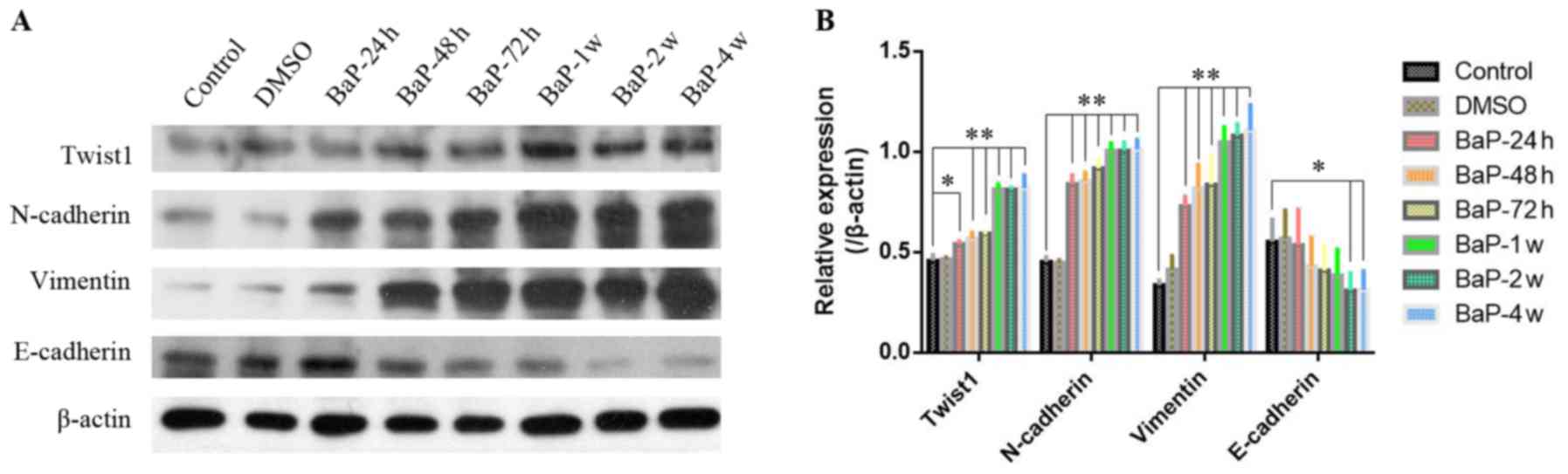
As shown in Figs. 3
and 4, after A549 cells were
exposed to 1 µM BaP for different durations of time (24, 48 and 72
h; 1, 2 or 4 weeks), both the mRNA (Fig. 3) and protein expression (Fig. 4) of Twist1 were gradually increased
with prolonged intervention times. Accordingly, the expression of
N-cadherin and vimentin were also gradually increased with
prolonged BaP intervention (Figs. 3
and 4). However, conversely, the
expression of E-cadherin gradually decreased as the BaP
intervention time was extended (Figs.
3 and 4).
Downregulation of Twist1 inhibits the
migration ability of A549BaP-4w cells
Compared with A549 cells without BaP intervention,
Twist1 was highly expressed in A549 cells treated with BaP for 4
weeks (A549BaP-4w) (Figs.
3 and 4), and the migration
capacity of A549BaP-4w cells was significantly enhanced
(Figs. 1 and 2). We hypothesized that downregulation of
Twist1 expression may decrease the migration of
A549BaP-4w cells. Thus, we applied wound healing and
Transwell migration assays to assess A549BaP-4w cells
that were transfected with recombinant plasmid containing short
hairpin RNA (Twist1-shRNA). The results of both the wound healing
(Fig. 5A) and Transwell migration
assay (Fig. 5B) revealed that
downregulation of Twist1 inhibited the migration ability of
A549BaP-4w cells.
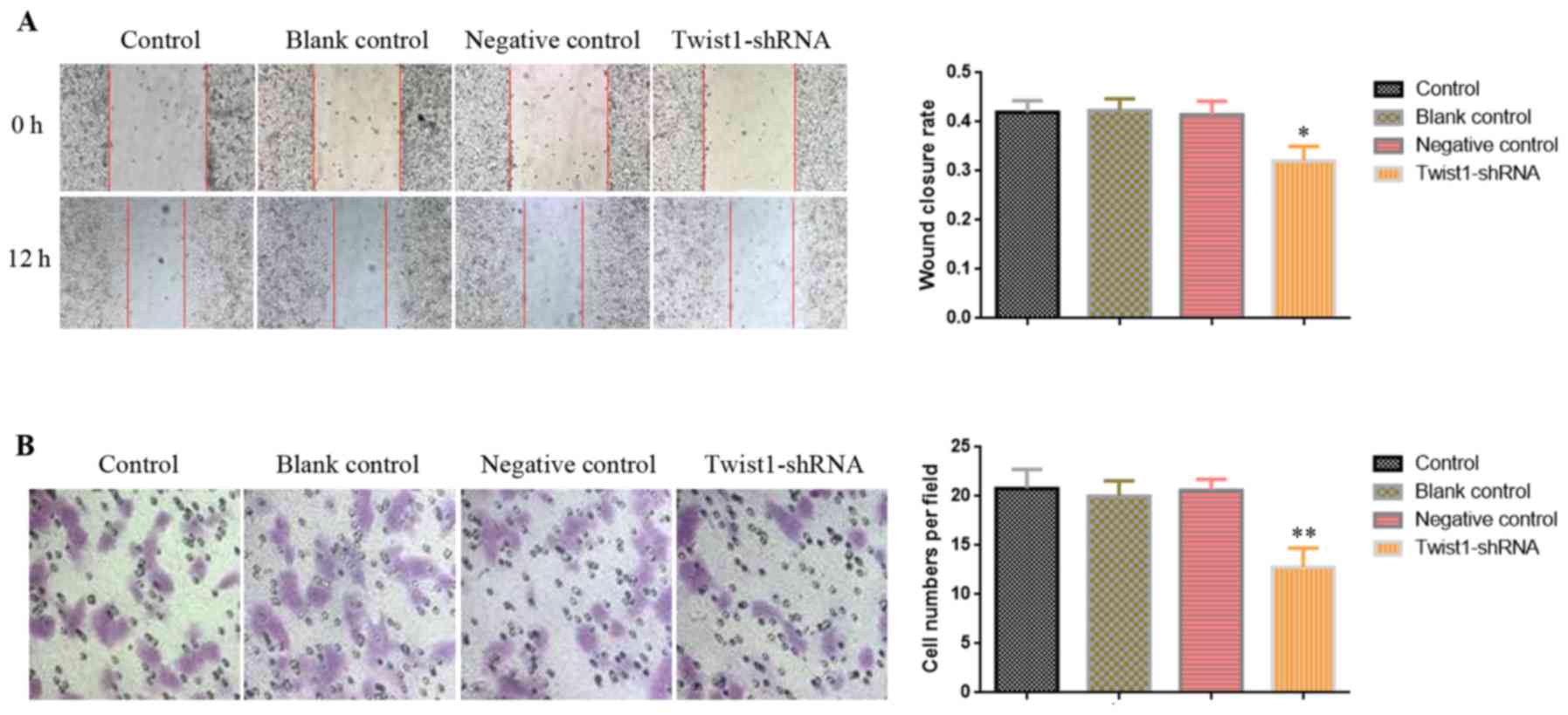 | Figure 5.Downregulation of Twist1 in
A549BaP-4w cells inhibits the migration ability. After
downregulation of Twist1 using Twist1-shRNA, (A) a wound healing
assay was performed with 12 h of recovery and (B) a Transwell
migration assay was performed at 24 h (the microscopic fields are
shown at a magnification of ×200). The graphics display the (A)
wound closure rate and (B) the number of cells/field between the
different groups. The statistical results are presented as the mean
± SD of 3 independent experiments; *p<0.05, **p<0.01.
A549BaP-4w, A549 cell model incubated with 1 µM BaP for
4 weeks, resulting in the upregulation of the expression of Twist1;
control, A549BaP-4w cell group without plasmid
transfection; blank control, A549BaP-4w cell group
transfected with blank plasmid; negative control,
A549BaP-4w cell group transfected with negative plasmid;
Twist1-shRNA, A549BaP-4w cell group transfected with
recombinant plasmid containing short hairpin RNA to downregulate
Twist1 gene expression. BaP, benzo(a)pyrene; Twist1, Twist family
BHLH transcription factor 1. |
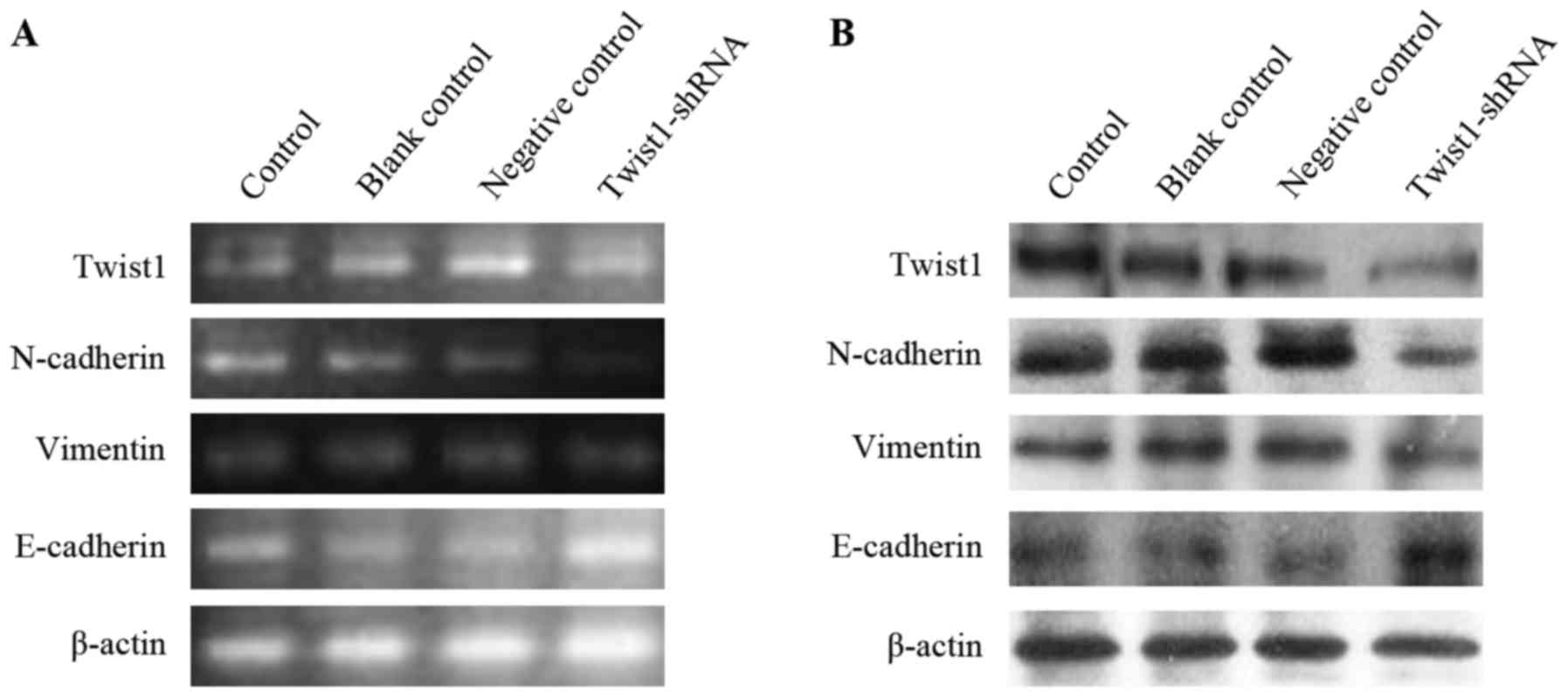
Decreased expression of Twist1 in
A549BaP-4w cells results in downregulation of N-cadherin
and vimentin and upregulation of E-cadherin
As shown in Fig. 6,
the expression of Twist1 in A549BaP-4w cells was
obviously downregulated after transfection with Twist1-shRNA.
Accordingly, the expression of N-cadherin and vimentin was also
markedly decreased after A549BaP-4w cells were
transfected with Twist1-shRNA (Fig. 6A
and B). However, decreased expression of Twist1 in
A549BaP-4w cells resulted in the upregulation of
E-cadherin expression (Fig. 6A and
B).
BaP induces EMT in A549 cells by
upregulating Twist1
Under a microscope, epithelial cells are
characterized by a flat and polygonal shape, and mesenchymal cells
are characterized by a relatively small cell body that is long and
thin. As shown in Fig. 7A, after 4
weeks of 1 µM BaP intervention, most of the of
A549BaP-4w cells transformed from cells with epithelial
characteristics to cells with mesenchymal characteristics. However,
after transfection of A549BaP-4w cells with Twist1-shRNA
(Fig. 7B) to downregulate the
expression of Twist1 (Fig. 7C), the
A549BaP-4w cells were transformed from cells with
mesenchymal characteristics back into cells with epithelial
characteristics (Fig. 7D).
Consequently, we concluded that BaP induced EMT in A549 cells by
upregulating Twist1.
Discussion
In the present study, we first found that BaP
promotes the migration of lung adenocarcinoma A549 cells in a
time-dependent manner. Increased expression of Twist1 in A549 cells
resulted in upregulation of N-cadherin and vimentin and
downregulation of E-cadherin. Then, after the expression of Twist1
was knocked down in A549 cells that were treated with BaP for 4
weeks (A549BaP-4w), the resulting decrease in the
expression of Twist1 inhibited the migration capacity of
A549BaP-4w cells, downregulated the expression of
N-cadherin and vimentin and upregulated the expression of
E-cadherin. Finally, along with the morphological results observed
under the microscope, we concluded that BaP enhanced the migration
of lung adenocarcinoma A549 cells through the promotion of EMT by
upregulation of Twist1.
Prior to the present study, Yoshino et al
(10) reported that BaP can induce
the EMT of lung cancer cells. In addition, D'Angelo et al
(23) and Yoon et al
(24) found that Twist1 can induce
the EMT of cancer cells. We therefore hypothesized that Twist1 may
be the target of BaP-treated lung adenocarcinoma A549 cells, and we
thus demonstrated this hypothesis in the present study. However, as
the mechanism of cigarette smoking-induced lung cancer is very
complex, we surmised that Twist1 cannot be the only target of BaP
in lung cancer cells. More studies on the mechanism of cigarette
smoking/BaP-induced lung cancer need to be conducted in the
future.
Yoshino et al (10) exposed A549 cells to 1 µM BaP for a
long period of time (24 weeks) and used a gene chip to conduct a
microarray analysis. They reported that the mRNA expression of
Twist was upregulated and the mRNA expression of E-cadherin was
downregulated. However, they did not observe morphological changes
under these experimental conditions (10). In the present study, by treating
A549 cells with 1 µM BaP for different lengths of time (24, 48 and
72 h; 1, 2 and 4 weeks), we found that the expression of Twist1,
N-cadherin and vimentin were increased at both the mRNA and protein
levels, while the expression of E-cadherin was decreased.
Furthermore, we discovered through observation under a microscope
that most of the A549 cells were transformed from cells with
epithelial characteristics to cells with mesenchymal
characteristics when treated with BaP for 4 weeks. Through a
comparative analysis, we hypothesized that a relatively short
period of BaP intervention can promote EMT in A549 cells, whereas a
long duration of intervention may reverse this effect. However, the
related mechanisms of this phenomenon remain to be clarified.
Wang et al (27) incubated A549 cells with 8 µM BaP for
24 h and reported that BaP promoted A549 cell migration. They found
that the mRNA and protein expression of Twist were upregulated in
A549 cells with treatment with 8 µM BaP for 24, 48 and 72 h
(27). In addition, when the
expression of Twist was knocked down in A549 cells, the migration
capacity was blocked by intervention with 8 µM BaP for 24 h
(27). In the present study, after
treating A549 cells of 1 µM BaP for at least 48 h, we obtained
results similar to those of Wang et al, specifically,
migration promotion and Twist1 upregulation. Similarly, we also
revealed that downregulation of Twist1 can inhibit the migration
capacity of A549BaP-4w cells, indicating that Twist1
indeed plays an important role in the progression of smoke-induced
lung cancer.
Generally, N-cadherin and vimentin are considered to
be mesenchymal markers (28–30)
while E-cadherin is regarded as an epithelial marker (28,29,31).
In the present study, notably, we found that the expression trend
of Twist1 was consistent with that of N-cadherin and vimentin but
contrary to that of E-cadherin. Furthermore, we also noted a
similar situation in many previous cancer studies (32–35).
Therefore, we speculate that Twist1 is a new potential mesenchymal
biomarker in the process of cancer progression. However, future
studies are warranted to confirm this speculation.
Previous studies have shown that Twist1 is an
oncogene (36–38). It promotes proliferation (17,39–41)
and inhibits apoptosis (42) of
cancer cells. Furthermore, emerging evidence suggests that Twist1
significantly enhances EMT-associated cell migration and invasion
to promote cancer metastasis (16).
It also plays a role in chemotherapeutic resistance (16,43).
Therefore, Twist1 is considered to be a potential therapeutic
target for cancer (16,44,45).
Since lung cancer is reported to be the leading cause of
cancer-related deaths (46), and
cigarette smoking is the main factor leading to lung cancer
(47), the development of
therapeutic strategies targeting Twist1 is particularly important
in the treatment of lung cancer.
In conclusion, the results from the present study
indicate that BaP enhances EMT-associated migration of lung
adenocarcinoma A549 cells by upregulating Twist1.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81572284) and the
Important Research and Development Plan of Hunan Provincial Science
and Technology Department (grant no. 2015SK20662).
References
|
1
|
Freedman ND, Leitzmann MF, Hollenbeck AR,
Schatzkin A and Abnet CC: Cigarette smoking and subsequent risk of
lung cancer in men and women: Analysis of a prospective cohort
study. Lancet Oncol. 9:649–656. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Islami F, Torre LA and Jemal A: Global
trends of lung cancer mortality and smoking prevalence. Transl Lung
Cancer Res. 4:327–338. 2015.PubMed/NCBI
|
|
3
|
Xu D, Penning TM, Blair IA and Harvey RG:
Synthesis of phenol and quinone metabolites of
benzo[a]pyrene, a carcinogenic component of tobacco smoke
implicated in lung cancer. J Org Chem. 74:597–604. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalabus JL, Cheng Q, Jamil RG, Schuetz EG
and Blanco JG: Induction of carbonyl reductase 1 (CBR1)
expression in human lung tissues and lung cancer cells by the
cigarette smoke constituent benzo[a]pyrene. Toxicol Lett.
211:266–273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kasala ER, Bodduluru LN, Barua CC, Sriram
CS and Gogoi R: Benzo(a)pyrene induced lung cancer: Role of dietary
phytochemicals in chemoprevention. Pharmacol Rep. 67:996–1009.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kometani T, Yoshino I, Miura N, Okazaki H,
Ohba T, Takenaka T, Shoji F, Yano T and Maehara Y: Benzo[a]pyrene
promotes proliferation of human lung cancer cells by accelerating
the epidermal growth factor receptor signaling pathway. Cancer
Lett. 278:27–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shinmura K, Iwaizumi M, Igarashi H, Nagura
K, Yamada H, Suzuki M, Fukasawa K and Sugimura H: Induction of
centrosome amplification and chromosome instability in
p53-deficient lung cancer cells exposed to benzo[a]pyrene
diol epoxide (B[a]PDE). J Pathol. 216:365–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tang SC, Sheu GT, Wong RH, Huang CY, Weng
MW, Lee LW, Hsu CP and Ko JL: Expression of glutathione
S-transferase M2 in stage I/II non-small cell lung cancer and
alleviation of DNA damage exposure to benzo[a]pyrene. Toxicol Lett.
192:316–323. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang BY, Wu SY, Tang SC, Lai CH, Ou CC, Wu
MF, Hsiao YM and Ko JL: Benzo[a]pyrene-induced cell cycle
progression occurs via ERK-induced Chk1 pathway activation in human
lung cancer cells. Mutat Res. 773:1–8. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yoshino I, Kometani T, Shoji F, Osoegawa
A, Ohba T, Kouso H, Takenaka T, Yohena T and Maehara Y: Induction
of epithelial-mesenchymal transition-related genes by
benzo[a]pyrene in lung cancer cells. Cancer. 110:369–374. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dasari V, Gallup M, Lemjabbar H, Maltseva
I and McNamara N: Epithelial-mesenchymal transition in lung cancer:
Is tobacco the ‘smoking gun’? Am J Respir Cell Mol Biol. 35:3–9.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nurwidya F, Takahashi F, Murakami A and
Takahashi K: Epithelial mesenchymal transition in drug resistance
and metastasis of lung cancer. Cancer Res Treat. 44:151–156. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Reka AK, Chen G, Jones RC, Amunugama R,
Kim S, Karnovsky A, Standiford TJ, Beer DG, Omenn GS and Keshamouni
VG: Epithelial-mesenchymal transition-associated secretory
phenotype predicts survival in lung cancer patients.
Carcinogenesis. 35:1292–1300. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qin Q, Xu Y, He T, Qin C and Xu J: Normal
and disease-related biological functions of Twist1 and underlying
molecular mechanisms. Cell Res. 22:90–106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han Z, Liu L, Liu Y and Li S: Sirtuin
SIRT6 suppresses cell proliferation through inhibition of Twist1
expression in non-small cell lung cancer. Int J Clin Exp Pathol.
7:4774–4781. 2014.PubMed/NCBI
|
|
18
|
Sakamoto A, Akiyama Y, Shimada S, Zhu WG,
Yuasa Y and Tanaka S: DNA methylation in the exon 1 region and
complex regulation of Twist1 expression in gastric cancer cells.
PLoS One. 10:e01456302015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu Y, Qin L, Sun T, Wu H, He T, Yang Z, Mo
Q, Liao L and Xu J: Twist1 promotes breast cancer invasion and
metastasis by silencing Foxa1 expression. Oncogene. 36:1157–1166.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Galván JA, Helbling M, Koelzer VH, Tschan
MP, Berger MD, Hädrich M, Schnüriger B, Karamitopoulou E, Dawson H,
Inderbitzin D, et al: TWIST1 and TWIST2 promoter methylation and
protein expression in tumor stroma influence the
epithelial-mesenchymal transition-like tumor budding phenotype in
colorectal cancer. Oncotarget. 6:874–885. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bing L, Hong C, Li-Xin S and Wei G:
MicroRNA-543 suppresses endometrial cancer oncogenicity via
targeting FAK and TWIST1 expression. Arch Gynecol Obstet.
290:533–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cho KH, Jeong KJ, Shin SC, Kang J, Park CG
and Lee HY: STAT3 mediates TGF-β1-induced TWIST1 expression and
prostate cancer invasion. Cancer Lett. 336:167–173. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
D'Angelo RC, Liu XW, Najy AJ, Jung YS, Won
J, Chai KX, Fridman R and Kim HR: TIMP-1 via TWIST1 induces EMT
phenotypes in human breast epithelial cells. Mol Cancer Res.
12:1324–1333. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoon NA, Jo HG, Lee UH, Park JH, Yoon JE,
Ryu J, Kang SS, Min YJ, Ju SA, Seo EH, et al: Tristetraprolin
suppresses the EMT through the down-regulation of Twist1 and Snail1
in cancer cells. Oncotarget. 7:8931–8943. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li L and Wu D: miR-32 inhibits
proliferation, epithelial-mesenchymal transition, and metastasis by
targeting TWIST1 in non-small-cell lung cancer cells. Onco Targets
Ther. 9:1489–1498. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Burns TF, Dobromilskaya I, Murphy SC,
Gajula RP, Thiyagarajan S, Chatley SN, Aziz K, Cho YJ, Tran PT and
Rudin CM: Inhibition of TWIST1 leads to activation of
oncogene-induced senescence in oncogene-driven non-small cell lung
cancer. Mol Cancer Res. 11:329–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Y, Zhai W, Wang H, Xia X and Zhang C:
Benzo(a)pyrene promotes A549 cell migration and invasion through
up-regulating Twist. Arch Toxicol. 89:451–458. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hänze J, Henrici M, Hegele A, Hofmann R
and Olbert PJ: Epithelial mesenchymal transition status is
associated with anti-cancer responses towards receptor
tyrosine-kinase inhibition by dovitinib in human bladder cancer
cells. BMC Cancer. 13:5892013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li B, Zheng YW, Sano Y and Taniguchi H:
Evidence for mesenchymal-epithelial transition associated with
mouse hepatic stem cell differentiation. PLoS One. 6:e170922011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shirakihara T, Saitoh M and Miyazono K:
Differential regulation of epithelial and mesenchymal markers by
deltaEF1 proteins in epithelial mesenchymal transition induced by
TGF-beta. Mol Biol Cell. 18:3533–3544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zeisberg M and Neilson EG: Biomarkers for
epithelial-mesenchymal transitions. J Clin Invest. 119:1429–1437.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen SF, Nieh S, Jao SW, Wu MZ, Liu CL,
Chang YC and Lin YS: The paracrine effect of cancer-associated
fibroblast-induced interleukin-33 regulates the invasiveness of
head and neck squamous cell carcinoma. J Pathol. 231:180–189. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Feng J, Fu Z, Guo J, Lu W, Wen K, Chen W,
Wang H, Wei J and Zhang S: Overexpression of peroxiredoxin 2
inhibits TGF-β1-induced epithelial-mesenchymal transition and cell
migration in colorectal cancer. Mol Med Rep. 10:867–873. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo G, Yao W, Zhang Q and Bo Y: Oleanolic
acid suppresses migration and invasion of malignant glioma cells by
inactivating MAPK/ERK signaling pathway. PLoS One. 8:e720792013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee YJ, Han ME, Baek SJ, Kim SY and Oh SO:
MED30 regulates the proliferation and motility of gastric cancer
Cells. PLoS One. 10:e01308262015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gort EH, van Haaften G, Verlaan I, Groot
AJ, Plasterk RH, Shvarts A, Suijkerbuijk KP, van Laar T, van der
Wall E, Raman V, et al: The TWIST1 oncogene is a direct
target of hypoxia-inducible factor-2alpha. Oncogene. 27:1501–1510.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Laursen KB, Mielke E, Iannaccone P and
Füchtbauer EM: Mechanism of transcriptional activation by the
proto-oncogene Twist1. J Biol Chem. 282:34623–34633. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nairismägi ML, Füchtbauer A, Labouriau R,
Bramsen JB and Füchtbauer EM: The proto-oncogene TWIST1 is
regulated by microRNAs. PLoS One. 8:e660702013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen H, Hu L, Luo Z, Zhang J, Zhang C, Qiu
B, Dong L, Tan Y, Ding J, Tang S, et al: A20 suppresses
hepatocellular carcinoma proliferation and metastasis through
inhibition of Twist1 expression. Mol Cancer. 14:1862015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qian J, Luo Y, Gu X, Zhan W and Wang X:
Twist1 promotes gastric cancer cell proliferation through
up-regulation of FoxM1. PLoS One. 8:e776252013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qiang L, Zhao B, Ming M, Wang N, He TC,
Hwang S, Thorburn A and He YY: Regulation of cell proliferation and
migration by p62 through stabilization of Twist1. Proc Natl Acad
Sci USA. 111:9241–9246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Orlandella FM, Di Maro G, Ugolini C,
Basolo F and Salvatore G: TWIST1/miR-584/TUSC2
pathway induces resistance to apoptosis in thyroid cancer cells.
Oncotarget. 7:70575–70588. 2016.PubMed/NCBI
|
|
43
|
Ren H, Du P, Ge Z, Jin Y, Ding D, Liu X
and Zou Q: TWIST1 and BMI1 in cancer metastasis and
chemoresistance. J Cancer. 7:1074–1080. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
da Silva SD, Alaoui-Jamali MA, Soares FA,
Carraro DM, Brentani HP, Hier M, Rogatto SR and Kowalski LP: TWIST1
is a molecular marker for a poor prognosis in oral cancer and
represents a potential therapeutic target. Cancer. 120:352–362.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tran PT, Shroff EH, Burns TF, Thiyagarajan
S, Das ST, Zabuawala T, Chen J, Cho YJ, Luong R, Tamayo P, et al:
Twist1 suppresses senescence programs and thereby
accelerates and maintains mutant Kras-induced lung
tumorigenesis. PLoS Genet. 8:e10026502012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dela Cruz CS, Tanoue LT and Matthay RA:
Lung cancer: Epidemiology, etiology, and prevention. Clin Chest
Med. 32:605–644. 2011. View Article : Google Scholar : PubMed/NCBI
|