Introduction
Laryngeal squamous cell carcinoma (LSCC) is one of
the most common types of tumor in the head and neck which accounts
for approximately 25% of the total (1,2).
During the last decade, great advance has been made in the study on
laryngeal carcinoma. The treatment for laryngeal carcinoma is the
combination of primary surgery with non-surgical radiotherapy or
chemoradiotherapy. However, the 5-year survival rate of laryngeal
carcinoma (60%) has not been improved. To identify novel biomarkers
that indicate the early stages of laryngeal carcinoma, or specific
biomarkers for various individuals, is urgently required for the
early detection of laryngeal carcinoma and the development of
individualized therapies.
MicroRNAs (miRNAs) are a class of small, non-coding
RNAs that regulate gene expression through targeting the
3′-untranslated region of target mRNA (3–5).
Increasing evidence indicates that miRNAs have significant roles in
various cell biological processes including cell proliferation,
cell cycle, cell apoptosis, cell migration and invasion (6,7). In
the carcinogenesis and development of laryngeal carcinoma, a number
of miRNAs have been reported to be dysregulated such as miR-135,
miR-144, miR-205, miR-296-5p, miR-206, miR-375, miR-101 and miR-150
(8–13). Recent studies suggest that miR-195
may act as a suppressor gene in various types of cancers (14–17),
indicating that miR-195 may be critical in cancer development.
However, the biological roles of miR-195 in laryngeal carcinoma
still remain to be established.
In the present study, we predicted DCUN1D1 is the
target gene of miR-195 by bioinformatic prediction and investigated
the effect of miR-195 on the biological function of LSCC cells and
identified the correlation between miR-195 and its target
DCUN1D1.
Materials and methods
Patient tissues
The LSCC tissues and adjacent normal tissues were
obtained from 122 patients at the Second Hospital of Tianjin
Medical University (Tianjin, China) following surgical resection,
and stored immediately in liquid nitrogen. The study was approved
by the ethics committee of the Second Hospital of Tianjin Medical
University and written informed consent was obtained from all
patients.
RNA isolation and qRT-PCR
Total RNA was extracted from the collected fresh
frozen tissues and cells using the standard TRIzol method. For the
detection of miRNA expression, cDNA were prepared by reversely
transcription of RNA using one Step PrimeScript miRNA cDNA
Synthesis kit (Takara, Dalian, China). A SYBR® Premix Ex
Taq™ II kit (Takara) was used for qRT-PCR in an ABI 7500 Real-time
PCR system (Applied Biosystems, Foster City, CA, USA). Relative
gene expression was calculated using 2−∆∆Ct method. U6
and GAPDH were selected as internal controls for miRNA and mRNA,
respectively. miR-195 mimics Mature sequence
5′-UAGCAGCACAGAAAUAUUGGC-3′; DCUN1D1 (forward:
5′-ACTCGATCCAGCCAGCATTA-3′, reverse: 5′-TGTTTGGAGAACTCGCACTG-3′);
U6 (forward: 5′-CTCGCTTCGGCAGCACA-3′, reverse:
5′-AACGCTTCACGAATTTGCGT-3′); GAPDH (forward:
5′-CGCTCTCTGCTCCTCCTGTT-3′, reverse:
5′-CCATGGTGTCTGAGCGATGT-3′).
Cell culture and transfections
The human laryngeal cancer cell line (TU-177) and
human normal bronchial epithelial cell line (16HBE) were obtained
from ATCC (Manassas, VA, USA) and maintained in RPMI supplemented
with 10% fetal bovine serum (HyClone, Logan, UT, USA). All the cell
lines were cultured in a humidified incubator with 5%
CO2 at 37°C. Transfections were done using Lipofectamine
2000/Lipofectamine LTX-Plus reagent (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer's instructions. miR-195 mimics
(forward: 5′-UAGCAGCACAGAAAUAUUGGC-3′, reverse:
5′-CAAUAUUUCUGUGCUGCUAUU-3′); Control (forward:
5′-UUCUCCGAACGUGUCACGU-3′, reverse: 5′-ACGUGACACGUUCGGAGAA-3′).
miR-195 inhibitor (5′-GCCAAUAUUUCUGUGCUGCUA-3′), Inhibitor control
(5′-CAAUAUUUCUGUGCUGCUAUU-3′).
Oligonucleotide and plasmid
transfection
The miRNA mimics and corresponding negative control
that were used for transient transfection were designed and
synthesized by GenePharma China. The DCUN1D1 cDNA containing the
coding sequence was cloned by PCR, and the PCR product was cloned
into the pcDNA3.1 vector (Invitrogen). The insert was confirmed by
DNA sequencing. The siRNA against DCUN1D1 (DCUN1D1 siRNA) was
synthesized by GenePharma China. The sequence of DCUN1D1 siRNA was
as follows (forward: 5′-GCAGATGACATGTCTAATT-3′, reverse:
5′-AAUUAGACAUGUCAUCUGC-3′). TU-177 cells were transfected with
miRNA mimic, siRNA, and pCDNA3.1-DCUN1D1 using Lipofectamine
2000/Lipofectamine LTX-Plus Reagent (Invitrogen) according to the
manufacturer's protocol.
Cell viability assay
Cell viability was determined by thiazolyl blue
tetrazolium bromide (MTT) assay. TU-177 cells were seeded into
96-well plates at a density of 4×104 cells/ml and
incubated for 24, 48 and 72 h. At each time point, 0.5% MTT
solution were respectively added to TU-177 cells, and incubated for
4 h at 37°C. Then the cell supernatants were discarded and 150 µl
DMSO were added to dissolve the formazan. The optical density of
each group was measured using a microplate reader (Bio-Rad
Laboratories, Hercules, CA, USA) at a wavelength of 490 nm.
Colony formation assay
Cells were seeded on 6-well plates at a density of
400 cells per well then cultured at 37°C in a incubator with 5%
CO2 for 14 days. The cells were fixed with methanol for
20 min at room temperature and stained with crystal violet solution
(Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The
colonies of more than 50 cells were counted.
Wound healing assay
Cells were seeded in 6-well plates at a density of
1×105 cells/well and cultured to confluence. Cell
monolayer was scraped with a 200 µl pipette tip to generate wounds
and then washed with DMEM media twice to remove cell debris. The
cells were grown in DMEM supplemented with 3% FBS for additional 48
h. The cell motility was measured by photographing five random
fields at the time of 0 and 24 h after wounding.
Transwell invasion assays
TU-177 cells were seeded into the upper chamber of a
Transwell insert (Millipore Corp., Billerica, MA, USA) and
incubated in an incubator with 5% CO2 at 37°C. Lower
chamber contains serum-loaded medium as attractant. The invasive
assay was performed using Transwell inserts coated with Matrigel
(BD Biosciences, San Diego, CA, USA). After incubation for 24 h,
the cells on upper surfaces of the Transwell chambers were removed
with cotton swabs, and the invaded cells were fixed with ethanol
and stained using 0.05% crystal violet solution. Images were
captured under a wide-field microscope (Nikon, Melville, NY,
USA).
Flow cytometry assay
For the apoptosis assay, cells were analyzed by flow
cytometry using a FITC-Annexin V/PI kit according to the
manufacturer's instructions (BD Pharmingen, Franklin Lakes, NJ,
USA). In detail, TU-177 cells were seeded into 6-well plates at a
density of 4×104 cells/ml and incubated for 24, 48 and
72 h. At each time point, TU-177 cells were collected and
resuspended with the PBS buffer. Then cells were incubated with 5
µl FITC-Annexin V and 5 µl PI in the dark at room temperature. The
apoptosis rates of cells were analyzed using a FACS Verse flow
cytometry (BD Biosciences) according to the fluorescence
signals.
Luciferase activity assay
The 3′UTR sequence of DCUN1D1 was amplified and
subcloned into the pmirGLO luciferase reporter vector (Promega,
Sunnyvale, CA, USA). HEK293T cells (5×104) were seeded
in 24-well plates and incubated for 24 h. Thereafter, cells were
co-transfected with wild-type (WT) or mutant (Mut) 3′UTR vectors
and miR-195 mimics using Lipofectamine 2000. After 48 h, the cells
were assayed for luciferase activity using the Dual-Luciferase
Reporter Assay System (Promega) following the manufacturer's
protocol. The firefly luciferase activities were normalized to
Renilla luciferase activity.
Immunohistochemistry assay
Sections (4 µm) were cut from formalin-fixed
paraffin-embedded tissue. After dewaxing and rehydration sections
were heated in a microwave oven for 2 min in citrate buffer at
middle power. Subsequently, sections were subjected to blockade of
endogenous peroxidase activity by incubating in 3%
H2O2 solution for 25 min. The slides were
then incubated with the rabbit polyclonal antibody against DCUN1D1
(Cell Signaling Technology, Inc., Danvers, MA, USA) at 4°C
overnight. After sequential incubation with a secondary antibody
(Abcam, Cambridge, UK) at room temperature for 1 h, sections were
washed and stained with 3,3′-diaminobenzidine, and visualized under
a microscope (Nikon, Japan). Brown staining in cytoplasm was
considered as positive immunoreactivity, and was evaluated as
percentage staining over the whole preparation. DCUN1D1 protein
level was classified semiquantitatively combining the proportion
and intensity of positively stained immunoreactive cells (18). The percentage of positive-staining
cells was scored as follows: 0 (<5% positive cells), 1 (5–50%
positive cells), and 2 (>50% positive cells). Staining intensity
was scored as follows: 0 (no staining or only weak staining); 1
(moderate staining); and 2 (strong staining). The sum of the
staining intensity score and the percentage score was used to
define the DCUN1D1 protein expression levels: 0–2, low expression
and 3–4, high expression.
Protein analysis
Cells were lysed in RIPA buffer (Beyotime Institute
of Biotechnology, Haimen, China) and the protein extracts were
quantified using a BCA-protein quantification kit (Beyotime
Institute of Biotechnology). Proteins were separated by SDS-PAGE,
transferred to PVDF membranes and then blocked with 10% non-fat
milk for 1.5 h and incubated overnight at 4°C with primary
antibodies against DCUN1D1, caspase-3 (Cell Signaling Technology,
Inc.), MMP-9, MMP-2, GAPDH (Santa Cruz Biotechnology, Santa Cruz,
CA, USA) at dilutions specified by the manufacturer. After being
washed with TBST, the membranes were incubated with the
HRP-conjugated secondary antibodies for 120 min at room
temperature. Then proteins were detected by chemiluminescence using
the ECL kit (Beyotime Institute of Biotechnology) and imaged with a
digital chemiscope (Qinxiang, Shanghai, China). Band intensity
quantification was calculated by ImageJ software, and GAPDH was
used as a loading control.
Statistical analysis
All statistical analyses were carried out using the
SPSS 17.0 statistical software package. Experimental data are
presented as mean ± SD. An independent Student's two-tailed t-test
and one-way ANOVA were performed to compare the difference. The
expression of miR-195 and DCUN1D1 related with various
clinicopathological characteristics were assessed using the
χ2 test or Fisher's exact test. The Spearman's
correlation was calculated between the expression levels of miR-195
and DCUN1D1 in LSCC. Survival curves were carried out by the
Kaplan-Meier method and Cox regression analysis was used for the
univariate and multivariate analysis. P-values <0.05 were
considered statistically significant.
Results
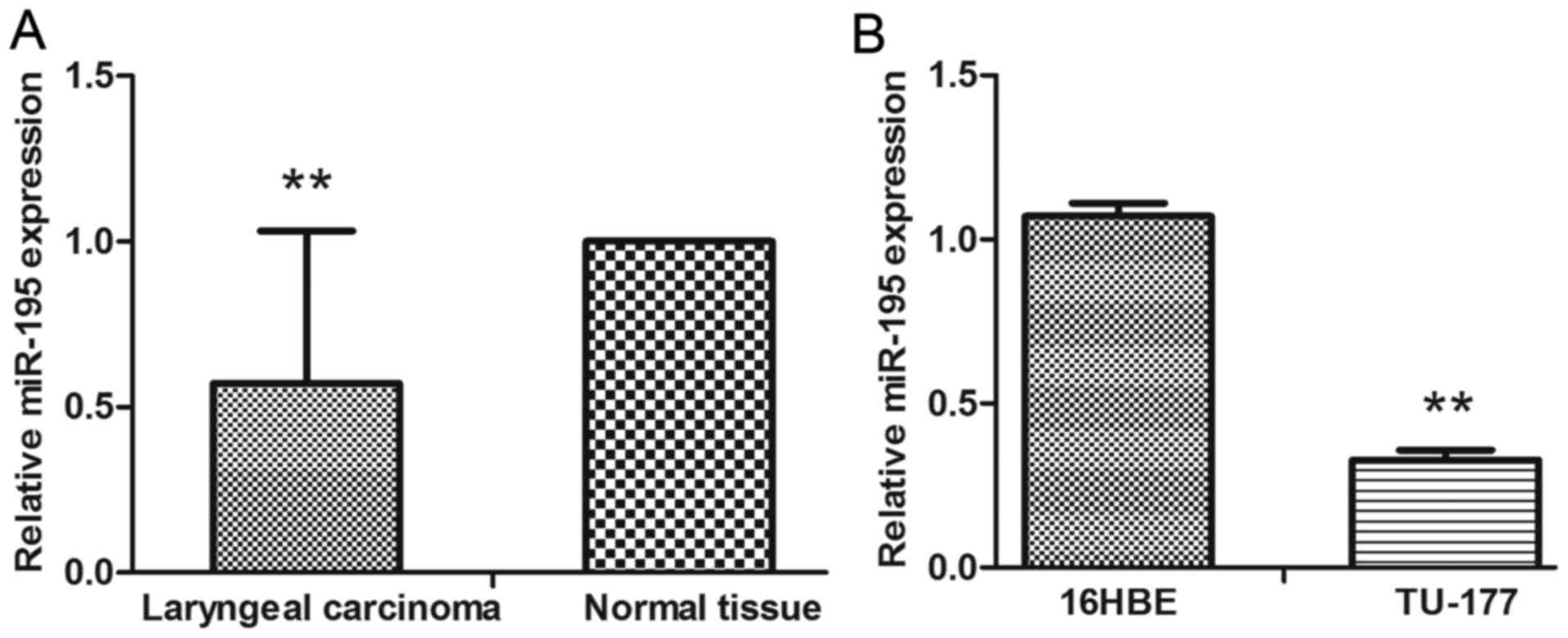
miR-195 was downregulated in LSCC and
cell lines, and was associated with cancer progression
The expression of miR-195 was evaluated using
qRT-PCR. We found that the expression levels of miR-195 in LSCC
were significantly decreased when compared with the adjacent normal
tissue and normal human bronchial epithelial cells (16HBE)
(Fig. 1A and B). To determine
whether the expression of miR-195 was associated with
clinicopathological characteristics of LSCC patients, the data
shown in Table I revealed that
miR-195 downregulation was frequently found in LSCC with high T
stage (P<0.001), N+ (P<0.05), and high clinical
stage (P<0.001). Decreased expression of miR-195 predicted poor
prognosis of LSCC.
 | Table I.Correlation between miR-195, DCUN1D1
expression and clinicopathological parameter in 122 LSCC
patients. |
Table I.
Correlation between miR-195, DCUN1D1
expression and clinicopathological parameter in 122 LSCC
patients.
|
|
| Expression of
miR-195 |
|
| Expression of
DCUN1D1 |
|
|
|---|
|
|
|
|
|
|
|
|
|
|---|
| Parameters | n | Low (n, %) | High (n, %) | χ2 | P-value | Low (n, %) | High (n, %) | χ2 | P-value |
|---|
| Age (years) |
|
≤60 | 69 | 35 (50.7) | 34 (49.3) | 0.033 | 0.855 | 33 (47.8) | 36 (52.2) | 0.078 | 0.780 |
|
>60 | 53 | 26 (49.1) | 27 (50.9) |
|
| 24 (45.3) | 29 (54.7) |
|
|
| Sex |
|
Female | 42 | 22 (52.4) | 20 (47.6) | 0.145 | 0.703 | 19 (45.2) | 23 (54.8) | 0.057 | 0.812 |
| Male | 80 | 39 (48.8) | 41 (51.2) |
|
| 38 (47.5) | 42 (52.5) |
|
|
| Primary site |
|
Glottis | 61 | 31 (50.8) | 30 (49.2) | 0.871 | 0.647 | 22 (36.1) | 39 (63.9) | 6.917 | 0.031 |
|
Supraglottis | 42 | 19 (45.2) | 23 (54.8) |
|
| 22 (52.4) | 20 (47.6) |
|
|
|
Subglottis | 19 | 11 (57.9) | 8 (42.1) |
|
| 13 (68.4) | 6 (31.6) |
|
|
| Differentiation
level |
|
High | 77 | 38 (49.4) | 39 (50.6) | 0.035 | 0.851 | 33 (42.9) | 44 (57.1) | 1.252 | 0.263 |
|
Medium-low | 45 | 23 (51.1) | 22 (48.9) |
|
| 24 (53.3) | 21 (46.7) |
|
|
| T stage |
|
T1+T2 | 30 | 1 (3.3) | 29 (96.7) |
|
<0.001a | 20 (66.7) | 10 (33.3) | 6.358 | 0.012 |
|
T3+T4 | 92 | 60 (65.2) | 32 (34.8) |
|
| 37 (40.2) | 55 (59.8) |
|
|
| Lymph node
metastases |
| N0 | 81 | 35 (43.2) | 46 (56.8) | 4.445 | 0.035 | 54 (66.7) | 27 (33.3) |
|
<0.001a |
| N+ | 41 | 26 (63.4) | 15 (36.6) |
|
| 3 (7.3) | 38 (92.7) |
|
|
| Distant
metastasis |
| M0 | 118 | 57 (48.3) | 61 (51.7) |
| 0.059a | 57 (48.3) | 61 (51.7) |
| 0.077a |
| M1 | 4 | 4 (100.0) | 0 (0.0) |
|
| 0 (0.0) | 4 (100.0) |
|
|
| Clinical stage |
|
I+II | 23 | 1 (4.3) | 22 (95.7) |
|
<0.001a | 18 (78.3) | 5 (21.7) | 11.326 | 0.001 |
|
III+IV | 99 | 60 (60.6) | 39 (39.4) |
|
| 39 (39.4) | 60 (60.6) |
|
|
| Preoperative
smoking |
| No | 23 | 15 (65.2) | 8 (34.8) | 2.625 | 0.105 | 8 (34.8) | 15 (65.2) | 1.623 | 0.203 |
|
Yes | 99 | 46 (46.5) | 53 (53.5) |
|
| 49 (49.5) | 50 (50.5) |
|
|
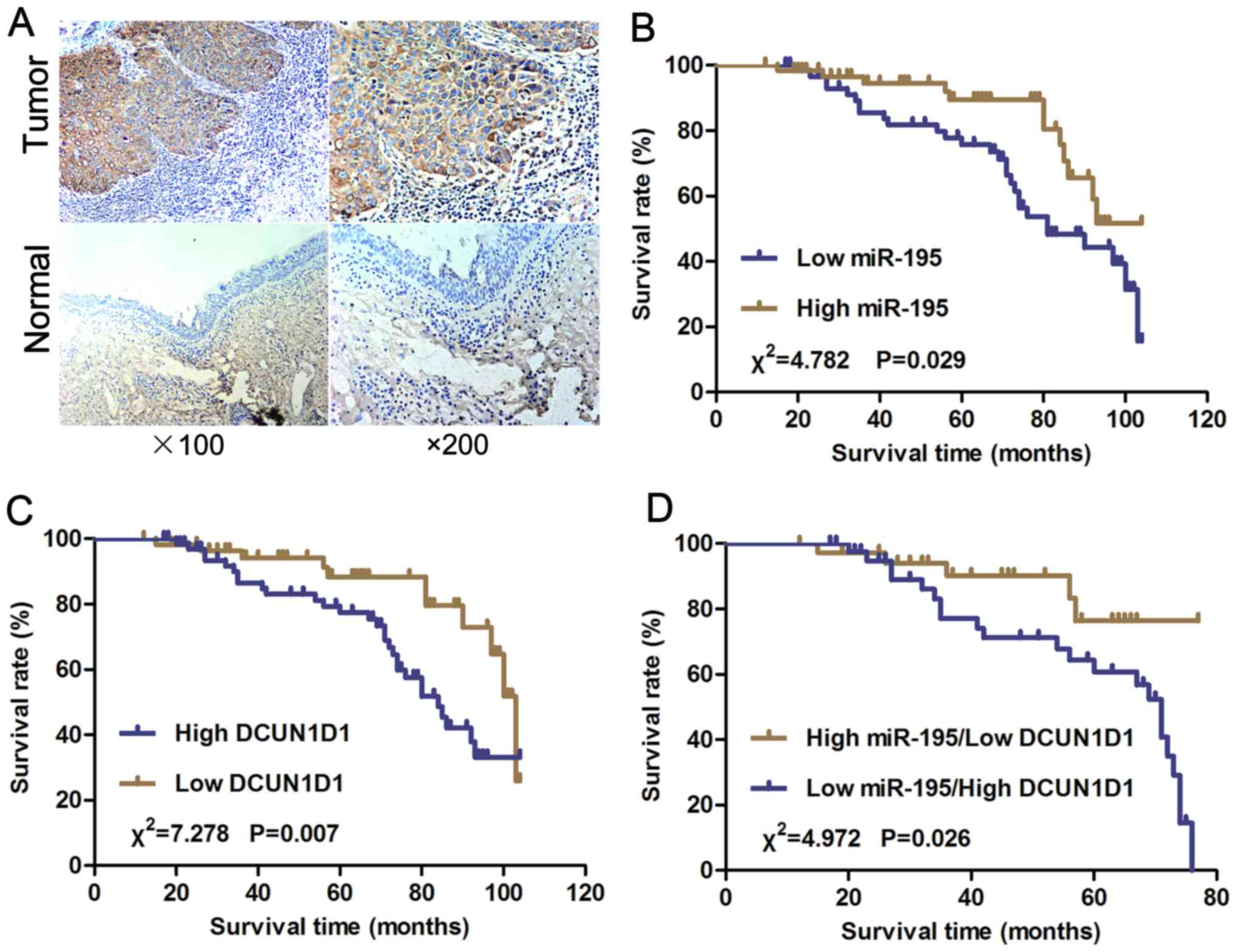
Upregulated DCUN1D1 expression was
associated with cancer progression and was inversely correlated
with miR-195 expression in LSCC
We examined the expression of the DCUN1D1 protein in
paraffin sections of LSCC and nonmalignant samples using
immunohistochemistry. The DCUN1D1 staining in LSCC adjacent
nonmalignant tissues was generally of reduced intensity (Fig. 2A). To understand the roles of
DCUN1D1 in LSCC progression, we analyzed the association of the
expression of DCUN1D1 with clinicopathological characteristics of
LSCC patients. As shown in Table I,
DCUN1D1 upregulation was frequently found in LSCC with high T stage
(P=0.012), N+ (P<0.001), and high clinical stage
(P=0.001). Increased expression of DCUN1D1 predicted poor prognosis
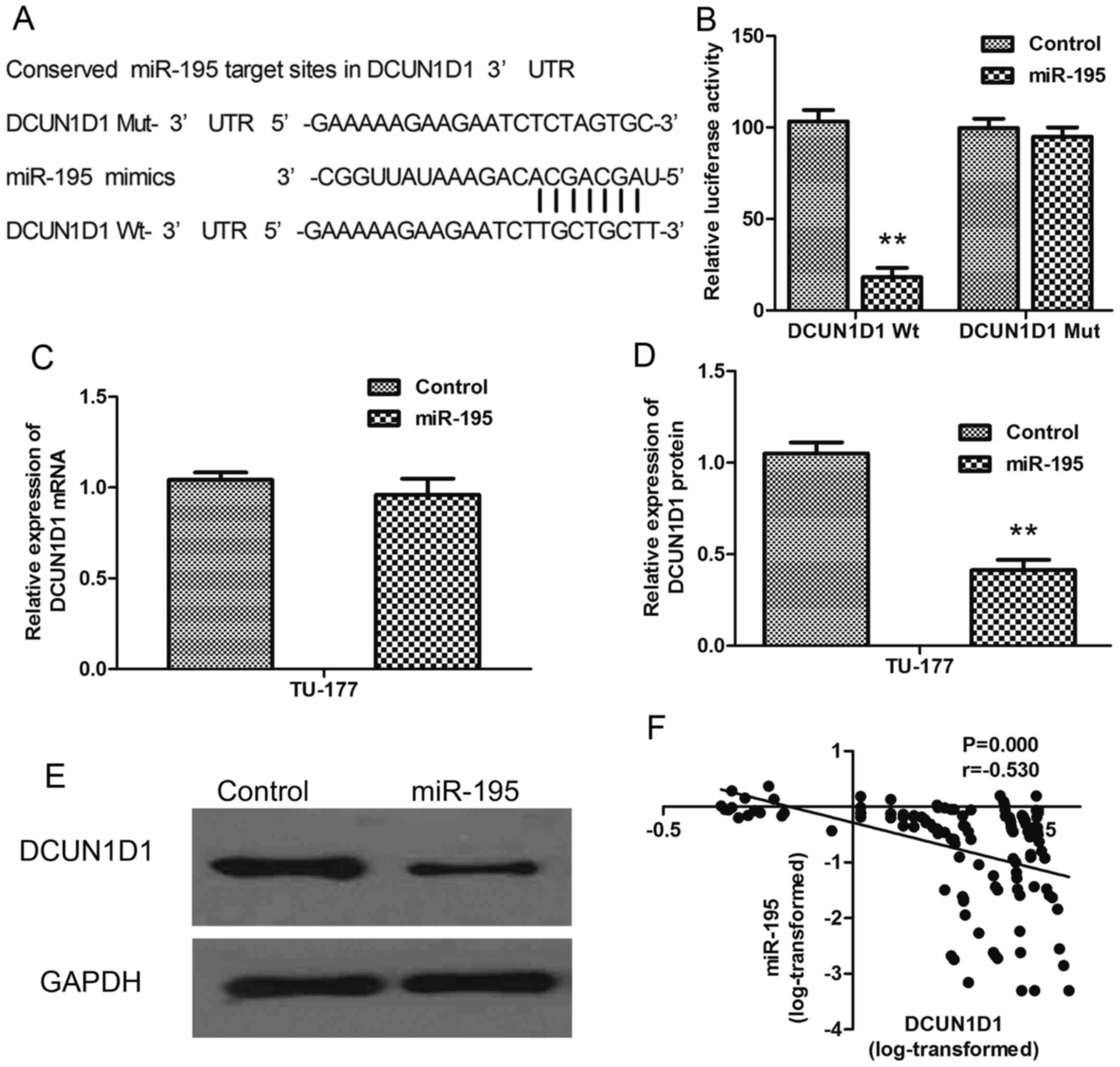
of LSCC. Spearman's correlation analysis showed expression levels
of DCUN1D1 were inversely correlated with miR-195 expression levels
in LSCC tissues (Table II,
Fig. 6F; r= −0.530, P<0.001).
Therefore, high DCUN1D1 mRNA expression in LSCC tissues was
correlated with decreased miR-195 expression.
| Table II.Correlation between miR-195 and
DCUN1D1. |
Table II.
Correlation between miR-195 and
DCUN1D1.
|
|
| Expression of
DCUN1D1 |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| Expression of
miR-195 | n | Low (n, %) | High (n, %) | rs | χ2 | P-value |
|---|
| Low (n, %) | 61 | 20 (32.8) | 41 (67.2) | −0.279 | 9.516 | 0.002 |
| High (n, %) | 61 | 37 (60.7) | 24 (39.3) |
|
|
|
Decreased miR-195 expression and
increased DCUN1D1 expression were associated with poor overall
survival in LSCC patients
We further analyzed the correlation of miR-195
expression and postoperative overall survival of LSCC patients. We
used the mean fold change (T/N=0.436) in miR-195 expression chosen
as the cut-off point and the patients were divided into a high
miR-195 expression group (T/N fold change >0.436) and a low
miR-195 expression group (T/N fold change <0.436). The overall
survival of patients with high miR-195 expression was longer than
those with low miR-195 expression (Fig.
2B; P=0.029). The overall survival of patients with low DCUN1D1
expression was statistically significantly longer than those with
high DCUN1D1 expression (Fig. 2C;
P=0.007). Furthermore, the overall survival of patients with high
miR-195 expression accompanied by low expression of DCUN1D1 was
longer than those with low miR-195 expression accompanied by high
DCUN1D1 expression (Fig. 2D;
P=0.026). A univariate and multivariable Cox proportional hazards
analysis were performed. The final multivariable Cox regression
model showed that the expression of miR-195 (P=0.041, relative
risk=0.358) and the expression of DCUN1D1 (P=0.021, relative
risk=4.253) were associated with poor prognosis according to
overall survival in LSCC (Table
III), suggesting that miR-195 and DCUN1D1 might be used as
independent prognostic factors for LSCC.
| Table III.Prognostic value of miR-195
expression and DCUN1D1 expression for the overall survival in
univariate and multivariate analyses by Cox regression. |
Table III.
Prognostic value of miR-195
expression and DCUN1D1 expression for the overall survival in
univariate and multivariate analyses by Cox regression.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Covariant | Exp (B) | 95% CI | P-value | Exp (B) | 95% CI | P-value |
|---|
| miR-195 | 0.476 | 0.241–0.942 | 0.033 | 0.358 | 0.134–0.959 | 0.041 |
| DCUN1D1 | 2.588 | 1.264–5.297 | 0.009 | 4.253 | 1.243–14.557 | 0.021 |
| Differentiation
level | 2.014 | 1.065–3.810 | 0.031 |
|
|
|
| T stage | 3.344 | 1.184–9.449 | 0.023 |
|
|
|
| Lymph node
metastasis | 2.809 | 1.474–5.350 | 0.002 |
|
|
|
| Distant
metastasis | 3.071 | 0.920–10.253 | 0.068 |
|
|
|
| Clinical stage | 3.586 | 0.859–14.976 | 0.080 |
|
|
|
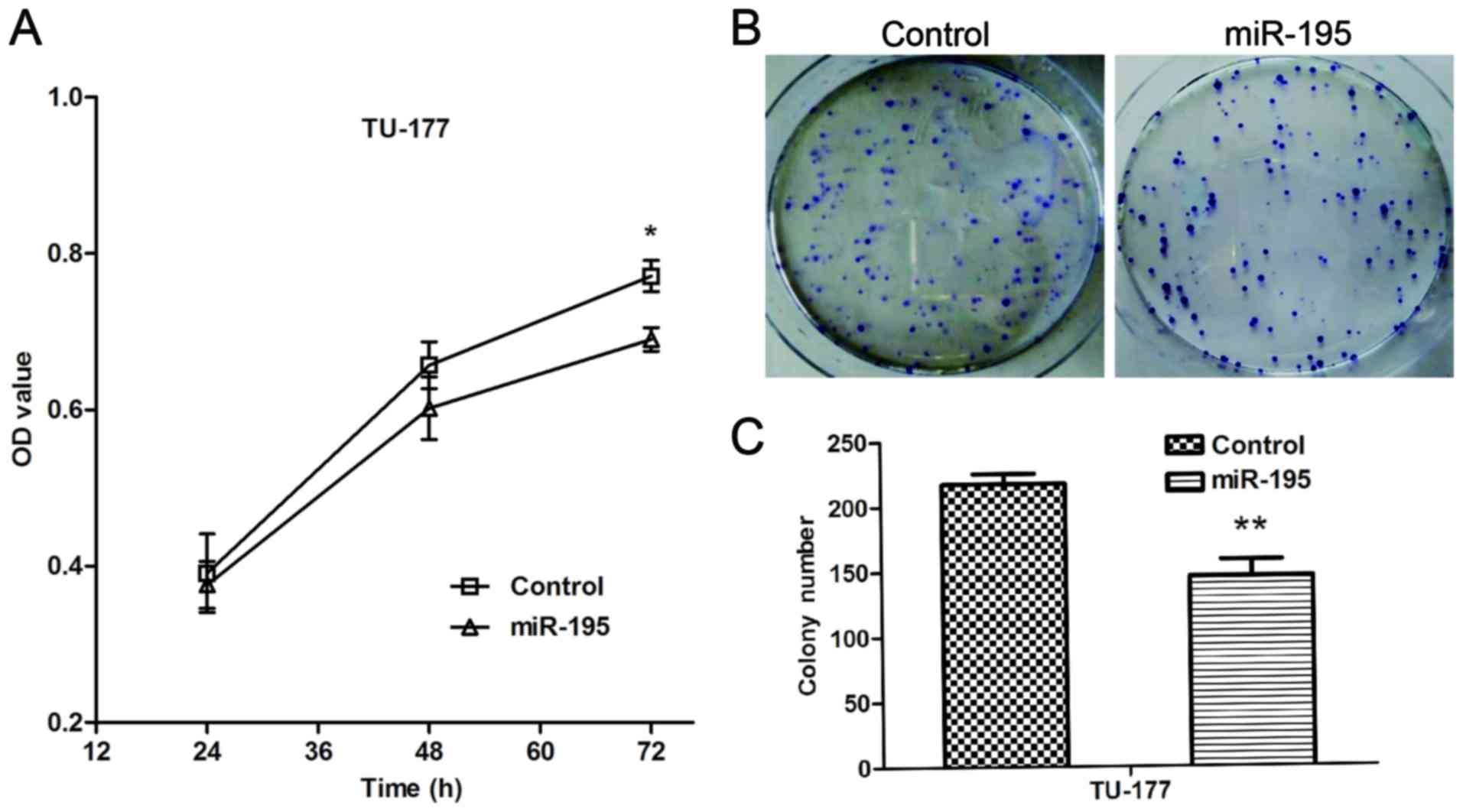
miR-195 inhibits the proliferation and
colony formation ability of LSCC cells
To investigate the biological functions of miR-195
in LSCC cells, we elevated the expression of miR-195 in the TU-177
cells. The MTT assay was performed to evaluate the effect of
miR-195 on the proliferation of TU-177 cells. The results indicated
that ectopic expression of miR-195 significantly reduced
proliferation of TU-177 cells (Fig.
3A). The colony formation assay was performed to determine
forced expression of miR-195 on the proliferation of TU-177 cells.
The results indicated miR-195 significantly inhibited colony
formation ability of TU-177 cells (Fig.
3B and C). Taken together, these data show that miR-195 may
function as a negative regulator on the proliferation of LSCC
cells.
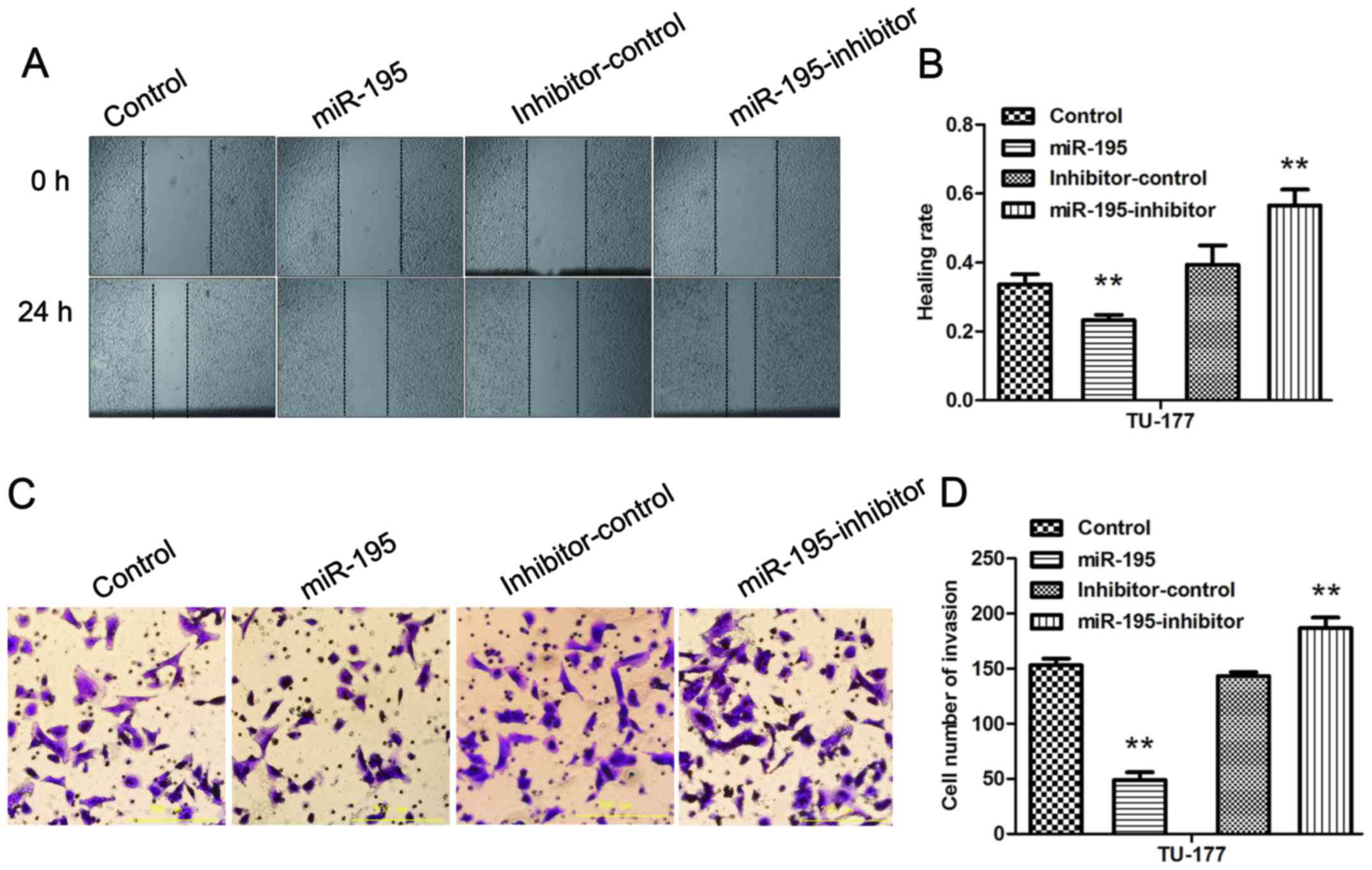
miR-195 inhibits the migration and
invasion ability of LSCC cells, and inhibition of miR-195 partially
reverses the effects
The wound healing assay was performed to evaluate
the effect of miR-195 on the migration ability of TU-177 cells. The
results indicated that transfection of mimics or inhibitors for
miR-195 could significantly inhibit or promote migration ability of
TU-177 cells (Fig. 4A and B). The
Transwell assay was conducted to investigate effect of miR-195 on
the invasion ability of TU-177 cells. The results showed that
transfection of mimics or inhibitors for miR-195 could
significantly reduce or increase the invasion ability of TU-177
cells (Fig. 4C and D). Thus, we
confirmed that miR-195 inhibitor could reverse the effects of
over-expression of miR-195 on the migration and invasion abilities
of LSCC cells.
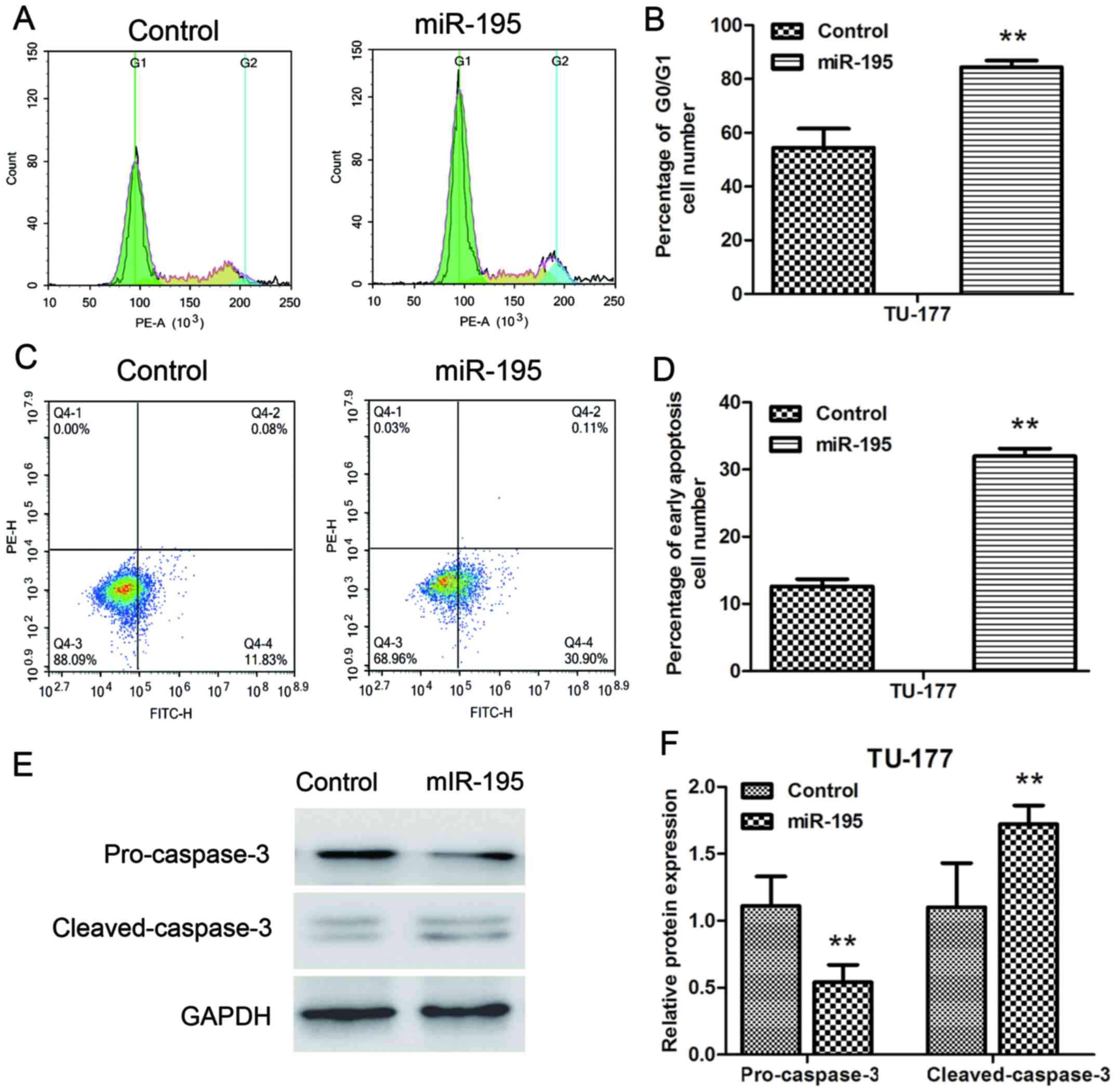
miR-195 induces G1 cell cycle arrest
and promoted apoptosis in LSCC cells
We further analyzed the effect of miR-195 on cell
cycle and apoptosis of TU-177 cells by flow cytometry. The cell
cycle distribution results indicated that miR-195 overexpression
significantly blocked the G1 phase of TU-177 cells (Fig. 5A and B). Moreover, miR-195
overexpression significantly increased early apoptotic cell number
of TU-177 cells (Fig. 5C and D). In
addition, the effect of miR-195 on endogenous expression levels of
pro-caspase-3 and cleaved-caspase-3 were assessed by western
blotting. Transfection of miR-195 significantly reduced the
expression of pro-caspase-3 and upregulated the expression of
cleaved-caspase-3 at the protein level in TU-177 cells (Fig. 5E and F). These results indicated
that overexpression of miR-195 mainly inhibits the growth of TU-177
cells by inducing G1 phase cell cycle arrest and apoptosis.
DCUN1D1 is directly targeted by
miR-195
In this study, DCUN1D1 was predicted to be a
potential target of miR-195 after computational analysis using
three microRNA target gene prediction software (TargetScan, miRDB,
DIANAmT) (Fig. 6A). To confirm
whether the 3′UTR of DCUN1D1 was a functional target of miR-195, we
constructed luciferase reporter plasmids containing
pGL3-DCUN1D1-3′UTR Wt and pGL3-DCUN1D1-3′UTR Mut and carried out
luciferase reporter assay. The results indicated that significant
decreases in luciferase activities were observed in the presence of
miR-195 in the HEK293T cells cotranfected with pGL3-DCUN1D1-3′UTR
Wt (P<0.01), but not with pGL3-DCUN1D1-3′UTR Mut (Fig. 6B). Furthermore, qRT-PCR and western
blot analysis showed a notable downregulation of DCUN1D1 expression
treated with overexpression of miR-195 in TU-177 cells at the
post-transcriptional level. (Fig.
6C-E). Above all, DCUN1D1 was the direct target of miR-195.
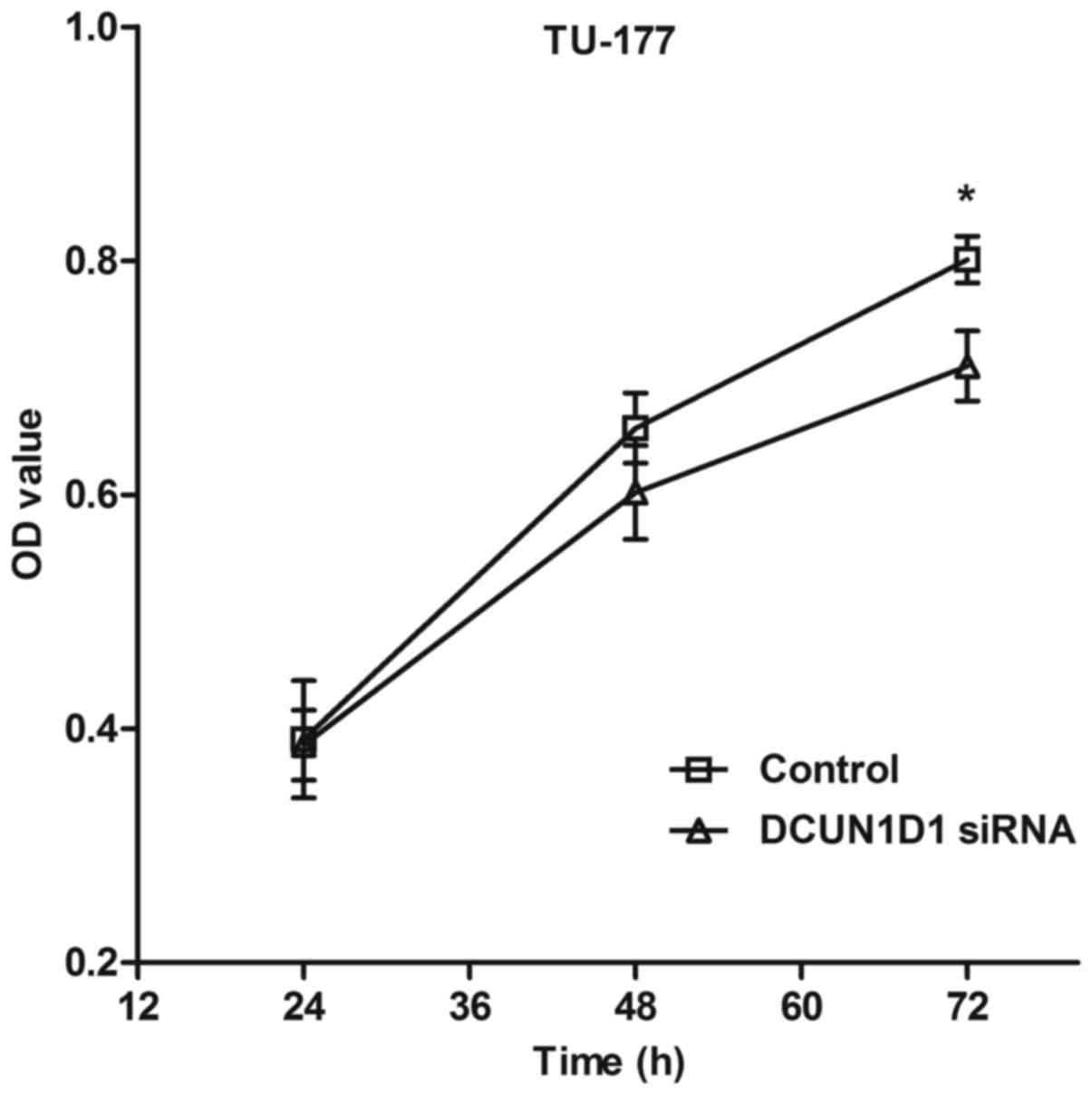
Knockdown of DCUN1D1 inhibits the
proliferation of LSCC cells
The MTT assay was performed to evaluate the effect
of DCUN1D1 siRNA on the proliferation of TU-177 cells. The results
indicated that ectopic expression of DCUN1D1 siRNA significantly
reduced proliferation of LSCC cells (Fig. 7).
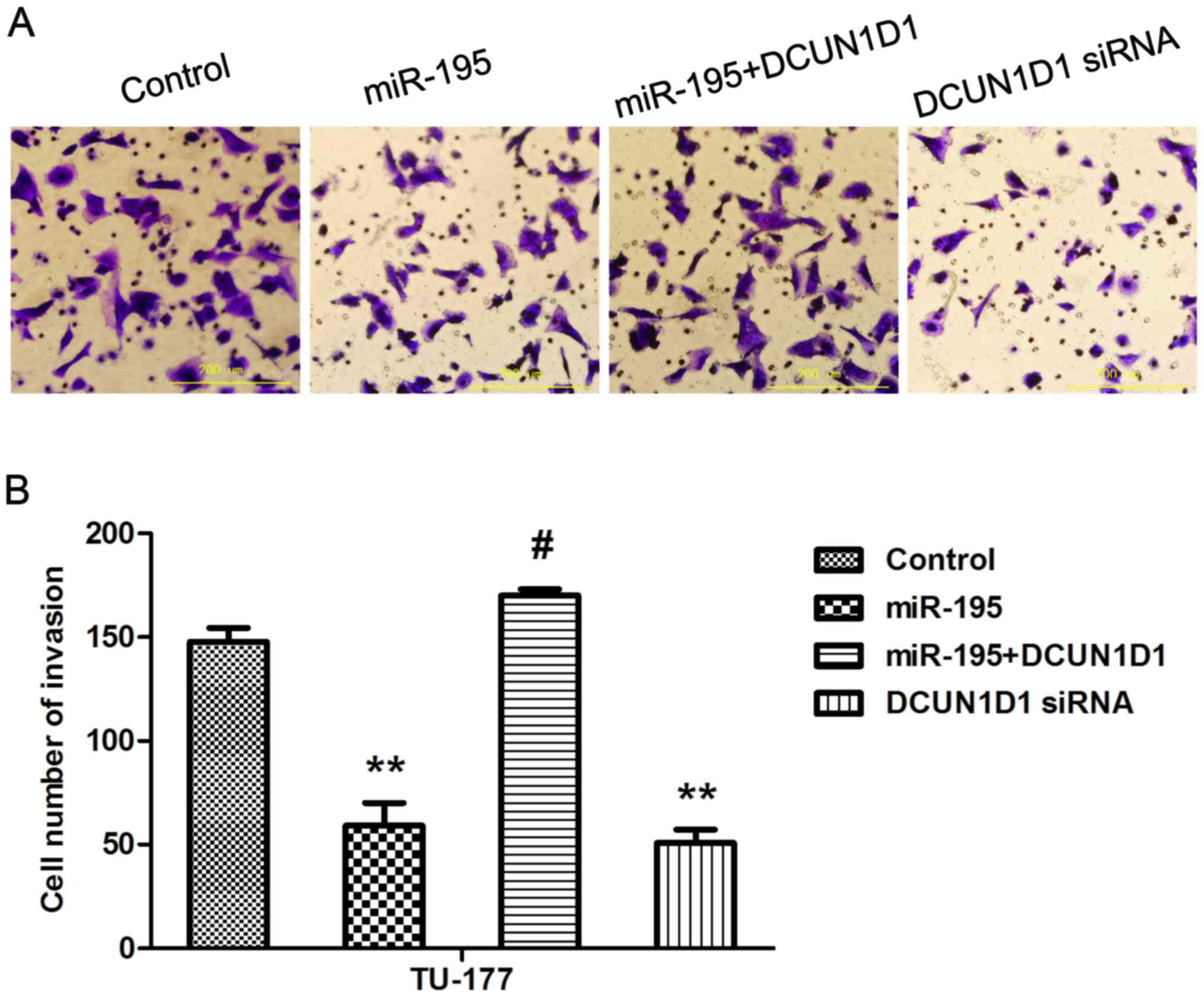
Overexpression of DCUN1D1 restores
invasive capacity of LSCC cells and knockdown of DCUN1D1 simulates
the effect of miR-195
To confirm that miR-195 inhibited the metastasis of
LSCC cells by targeting DCUN1D1, we next performed cell invasion
assays in TU-177 cells that were co-transfected with miR-195 and
pcDNA3.1-DCUN1D1, which lacked the 3′UTR of DCUN1D1 and therefore
cannot be restrained by miR-195. The results showed that forced
DCUN1D1 expression promoted the invasive capability of TU-177
cells. Importantly, this restoration of DCUN1D1, partially, but
significantly, rescued the invasion capability of
miR-195-transfected cells. Knockdown of DCUN1D1 exerted similar
effect as miR-195 (Fig. 8A and B).
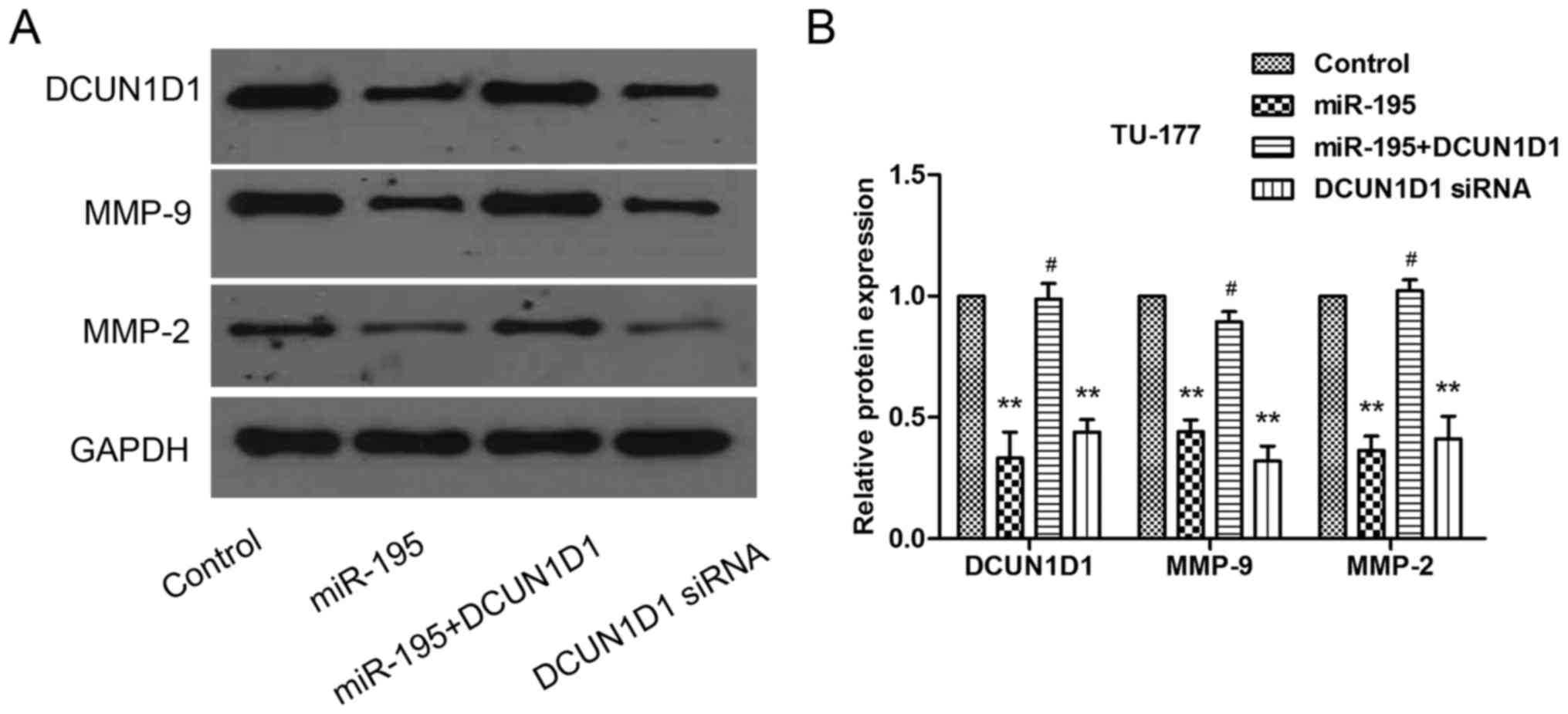
Besides, the DCUN1D1, MMP-9 and MMP-2 protein expression levels
confirmed by western blot analysis were consistent with Transwell
assay (Fig. 9A and B). Taken
together, these data suggested that the effects of miR-195 on cell
invasion were partly regulated by DCUN1D1.
Discussion
Extensive studies have demonstrated that miRNAs play
important roles in tumor initiation, progression, and metastasis
particularly through regulating their corresponding target genes
(19,20). One of them, miR-195 has been
regarded as a tumor suppressor in various cancers including
non-small cell lung cancer (NSCLC) (21), colorectal cancer (22), gastric cancer (23), hepatocellular carcinoma (24), and prostate cancer (25).
In the present study, we found that miR-195 was
downregulated significantly in LSCC tissue and cell lines compared
to the normal tissue and human bronchial epithelial cells which is
consistent with a previous study (26). We also found that miR-195
downregulation was frequently found in LSCC with high T stage,
N+, and high clinical stage. However, the role of
miR-195 in LSCC remains unknown. For the first time, overexpression
of miR-195 was used to observe its effect on LSCC cells by
transfection technique. As expected, miR-195 inhibited the
proliferation, colony formation, migration and invasion ability and
induced G1 cell cycle arrest and promoted apoptosis in LSCC cell
lines.
miRNAs perform biological functions by directly
binding to the 3′UTR of mRNAs and inhibits protein translation. For
instance, CARMA3 and Bcl-2 are direct targets of miR-195 in
colorectal cancer (27,28). IGF1R and HDGF are direct targets of
miR-195 in NSCLC (29). We
predicted the target gene of miR-195 using bioinformatics analysis
and selected DCUN1D1 as the potential target gene. DCUN1D1 is also
known as squamous cell carcinoma-related oncogene (SCCRO). It is a
novel gene initially identified as a result of 3q26-27 locus
amplification of in head and neck squamous cell carcinoma (30). DCUN1D1 can enhance recruitment of
Ubc12 to Cul1 to promote neddylation in the nucleus. DCUN1D1 has an
essential role in neddylation in vivo involving nuclear
localization of neddylation components and recruitment and proper
positioning of Ubc12 (31). DCUN1D1
was previously reported to be associated with tumor progression and
development of brain metastasis in patients with NSCLC (32). DCUN1D1 overexpression was associated
with an aggressive clinical course in primary SCC of the lung. It
functions as an oncogene which can transform cells of both
fibroblastic and keratinocytic lineage (33). Luciferase reporter assays suggested
that DCUN1D1 was one of the downstream targets of miR-195. We found
that miR-195 significantly downregulated DCUN1D1 expression at the
post-transcriptional level in LSCC cells. We also found that
DCUN1D1 was overexpressed in human laryngeal carcinoma tissues when
compared with its adjacent normal tissues and negatively correlated
with the levels of miR-195. In addition, DCUN1D1 upregulation was
also frequently found in LSCC with high T stage, N+, and
high clinical stage. Our results indicate that miR-195, as a tumor
suppressor miRNA, is also a negative regulator of the oncogene
DCUN1D1.
More than 90% of carcinoma-associated mortality is
caused by distant metastases (34).
Cancer cell invasion and metastasis is a complicated, multi-step
procedure involving interactions between invading cells, the
extracellular matrix, and other stromal elements (35). Proteolytic enzymes produced by tumor
cells are essential for cancer cells to degrade the extracellular
matrix and penetrate lymphatic or blood vessel walls to extend to
regional or distant sites. Matrix metalloproteinases (MMP), a
family of zinc-dependent endopeptidases, play a distinguished role
in extracellular matrix invasion and angiogenesis (36,37).
The aberrant expression or activation of MMP-2 and MMP-9 has been
linked to promotion of tumor invasion or metastasis in many
different human tumors (38).
DCUN1D1 can induce MMP-2 transcription indirectly through the
upregulation of activator protein 2 (AP2) in a process requiring
p53. It may be a marker for invasion and metastasis in squamous
cell carcinoma (39). Recent study
revealed that miR-218 inhibited invasion in cervical cancer by
targeting DCUN1D1. DCUN1D1 knockdown was confirmed to significantly
decrease the invasiveness of cervical cancer cells (40). However, this effect of DCUN1D1 has
not been studied in LSCC. Our study indicates that DCUN1D1
knockdown can successfully simulate the anti-proliferative,
anti-invasion effects of miR-195 on LSCC cells. Moreover,
introduction of DCUN1D1 can reduce invasion inhibition induced by
miR-195. We also found that both miR-195 and DCUN1D1 siRNA can
reduce invasion involving the post-transcriptional downregulation
of MMP-2 and MMP-9. These effects can be attenuated by introduction
of DCUN1D1.
In conclusion, DCUN1D1 is a potential target of
miR-195 and miR-195 may suppress growth and invasion of LSCC cells
possibly through targeting DCUN1D1, which would provide a candidate
target for cancer therapy.
Acknowledgements
The study was supported by the science and
technology fund of Tianjin Health Bureau (2015KZ106).
References
|
1
|
Chen Z, Jin Y, Yu D, Wang A, Mahjabeen I,
Wang C, Liu X and Zhou X: Down-regulation of the microRNA-99 family
members in head and neck squamous cell carcinoma. Oral Oncol.
48:686–691. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Borel C, Deutsch S, Letourneau A,
Migliavacca E, Montgomery SB, Dimas AS, Vejnar CE, Attar H,
Gagnebin M, Gehrig C, et al: Identification of cis- and
trans-regulatory variation modulating microRNA expression levels in
human fibroblasts. Genome Res. 21:68–73. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Macfarlane LA and Murphy PR: MicroRNA:
Biogenesis, function and role in cancer. Curr Genomics. 11:537–561.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Davidson B, Tropé CG and Reich R: The
clinical and diagnostic role of microRNAs in ovarian carcinoma.
Gynecol Oncol. 133:640–646. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nugent M: MicroRNA function and
dysregulation in bone tumors: The evidence to date. Cancer Manag
Res. 6:15–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li M, Tian L, Ren H, Chen X, Wang Y, Ge J,
Wu S, Sun Y, Liu M and Xiao H: MicroRNA-101 is a potential
prognostic indicator of laryngeal squamous cell carcinoma and
modulates CDK8. J Transl Med. 13:2712015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu JY, Lu JB and Xu Y: MicroRNA-153
inhibits the proliferation and invasion of human laryngeal squamous
cell carcinoma by targeting KLF5. Exp Ther Med. 11:2503–2508. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu S, Jia S and Xu P: MicroRNA-9 as a
novel prognostic biomarker in human laryngeal squamous cell
carcinoma. Int J Clin Exp Med. 7:5523–5528. 2014.PubMed/NCBI
|
|
11
|
Wu X, Cui CL, Chen WL, Fu ZY, Cui XY and
Gong X: miR-144 suppresses the growth and metastasis of laryngeal
squamous cell carcinoma by targeting IRS1. Am J Transl Res. 8:1–11.
2016.PubMed/NCBI
|
|
12
|
Yu WF, Wang HM, Lu BC, Zhang GZ, Ma HM and
Wu ZY: miR-206 inhibits human laryngeal squamous cell carcinoma
cell growth by regulation of cyclinD2. Eur Rev Med Pharmacol Sci.
19:2697–2702. 2015.PubMed/NCBI
|
|
13
|
Zhong G and Xiong X: miR-205 promotes
proliferation and invasion of laryngeal squamous cell carcinoma by
suppressing CDK2AP1 expression. Biol Res. 48:602015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu C, Guan H, Wang Y, Chen M, Xu B, Zhang
L, Lu K, Tao T, Zhang X and Huang Y: miR-195 inhibits EMT by
targeting FGF2 in prostate cancer cells. PLoS One. 10:e01440732015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luo Q, Zhang Z, Dai Z, Basnet S, Li S, Xu
B and Ge H: Tumor-suppressive microRNA-195-5p regulates cell growth
and inhibits cell cycle by targeting cyclin dependent kinase 8 in
colon cancer. Am J Transl Res. 8:2088–2096. 2016.PubMed/NCBI
|
|
16
|
Singh R, Yadav V, Kumar S and Saini N:
MicroRNA-195 inhibits proliferation, invasion and metastasis in
breast cancer cells by targeting FASN, HMGCR, ACACA and CYP27B1.
Sci Rep. 5:174542015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao C, Qi L, Chen M, Liu L, Yan W, Tong S
and Zu X: microRNA-195 inhibits cell proliferation in bladder
cancer via inhibition of cell division control protein 42
homolog/signal transducer and activator of transcription-3
signaling. Exp Ther Med. 10:1103–1108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo J, Wang M and Liu X: MicroRNA-195
suppresses tumor cell proliferation and metastasis by directly
targeting BCOX1 in prostate carcinoma. J Exp Clin Cancer Res.
34:912015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu B, Qu J, Xu F, Guo Y, Wang Y, Yu H and
Qian B: MiR-195 suppresses non-small cell lung cancer by targeting
CHEK1. Oncotarget. 6:9445–9456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tan YG, Zhang YF, Guo CJ, Yang M and Chen
MY: Screening of differentially expressed microRNA in ulcerative
colitis related colorectal cancer. Asian Pac J Trop Med. 6:972–976.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo J, Miao Y, Xiao B, Huan R, Jiang Z,
Meng D and Wang Y: Differential expression of microRNA species in
human gastric cancer versus non-tumorous tissues. J Gastroenterol
Hepatol. 24:652–657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sohn W, Kim J, Kang SH, Yang SR, Cho JY,
Cho HC, Shim SG and Paik YH: Serum exosomal microRNAs as novel
biomarkers for hepatocellular carcinoma. Exp Mol Med. 47:e1842015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu J, Ji A, Wang X, Zhu Y, Yu Y, Lin Y,
Liu Y, Li S, Liang Z, Xu X, et al: MicroRNA-195-5p, a new regulator
of Fra-1, suppresses the migration and invasion of prostate cancer
cells. J Transl Med. 13:2892015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu ZM, Lin YF, Jiang L, Chen LS, Luo XN,
Song XH, Chen SH and Zhang SY: Micro-ribonucleic acid expression
profiling and bioinformatic target gene analyses in laryngeal
carcinoma. Onco Targets Ther. 7:525–533. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu L, Chen L, Xu Y, Li R and Du X:
microRNA-195 promotes apoptosis and suppresses tumorigenicity of
human colorectal cancer cells. Biochem Biophys Res Commun.
400:236–240. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang L, Qian L, Li X and Yan J:
MicroRNA-195 inhibits colorectal cancer cell proliferation,
colony-formation and invasion through targeting CARMA3. Mol Med
Rep. 10:473–478. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo H, Li W, Zheng T and Liu Z: MiR-195
targets HDGF to inhibit proliferation and invasion of NSCLC cells.
Tumour Biol. 35:8861–8866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Singh B, Gogineni SK, Sacks PG, Shaha AR,
Shah JP, Stoffel A and Rao PH: Molecular cytogenetic
characterization of head and neck squamous cell carcinoma and
refinement of 3q amplification. Cancer Res. 61:4506–4513.
2001.PubMed/NCBI
|
|
31
|
Huang G, Kaufman AJ, Ramanathan Y and
Singh B: SCCRO (DCUN1D1) promotes nuclear translocation and
assembly of the neddylation E3 complex. J Biol Chem.
286:10297–10304. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yoo J, Lee SH, Lym KI, Park SY, Yang SH,
Yoo CY, Jung JH, Kang SJ and Kang CS: Immunohistochemical
expression of DCUN1D1 in non-small cell lung carcinoma: Its
relation to brain metastasis. Cancer Res Treat. 44:57–62. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sarkaria I, O-charoenrat P, Talbot SG,
Reddy PG, Ngai I, Maghami E, Patel KN, Lee B, Yonekawa Y, Dudas M,
et al: Squamous cell carcinoma related oncogene/DCUN1D1 is highly
conserved and activated by amplification in squamous cell
carcinomas. Cancer Res. 66:9437–9444. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liotta LA, Steeg PS and Stetler-Stevenson
WG: Cancer metastasis and angiogenesis: An imbalance of positive
and negative regulation. Cell. 64:327–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Folkman J: What is the evidence that
tumors are angiogenesis dependent? J Natl Cancer Inst. 82:4–6.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
MacDougall JR and Matrisian LM:
Contributions of tumor and stromal matrix metalloproteinases to
tumor progression, invasion and metastasis. Cancer Metastasis Rev.
14:351–362. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
O-Charoenrat P, Rhys-Evans PH and Eccles
SA: Expression of matrix metalloproteinases and their inhibitors
correlates with invasion and metastasis in squamous cell carcinoma
of the head and neck. Arch Otolaryngol Head Neck Surg. 127:813–820.
2001.PubMed/NCBI
|
|
39
|
O-Charoenrat P, Sarkaria I, Talbot SG,
Reddy P, Dao S, Ngai I, Shaha A, Kraus D, Shah J, Rusch V, et al:
SCCRO (DCUN1D1) induces extracellular matrix invasion by activating
matrix metalloproteinase 2. Clin Cancer Res. 14:6780–6789. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jiang Z, Song Q, Zeng R, Li J, Li J, Lin
X, Chen X, Zhang J and Zheng Y: MicroRNA-218 inhibits EMT,
migration and invasion by targeting SFMBT1 and DCUN1D1 in cervical
cancer. Oncotarget. 7:45622–45636. 2016. View Article : Google Scholar : PubMed/NCBI
|