Introduction
The clinical outcome of muscle-invasive bladder
cancer is poor: approximately half the patients treated by radical
cystectomy develop metastases within 2 years, and without other
treatment the 5-year survival rate after surgery is ~50% (1,2).
Metastatic bladder cancer is a chemosensitive disease and systemic
cisplatin-based combination chemotherapy is the first-line
treatment modality for it. Despite response rates of 40–60%
achieved by cisplatin-based chemotherapy, in most patients the
disease may progress after a median time of ~8 months (3). Targeted molecular therapy interfering
with cellular processes essential for cancer growth has been
introduced as a novel treatment strategy during the last 2 decades,
but no targeted agent approved for bladder cancer treatment has yet
emerged (4).
The signal transducer and activator of transcription
(STAT) proteins are involved in cytokine- and growth factor-induced
signal transduction (5). These
latent cytoplasmic transcription factors become activated through
tyrosine phosphorylation typically catalyzed by
cytokine-receptor-associated kinases, Janus kinases (Jak), or
growth factor receptor tyrosine kinases and are translocated to the
nucleus to activate target genes (5). One of the STAT proteins, STAT3, was
originally identified as an acute-phase response factor activated
by the interleukin-6 (IL-6) family of cytokines (6). Further studies have shown that it can
be activated by a variety of cytokines and growth factors (7). It regulates the transcription of key
components of cell cycle control and modulates the activity of
proteins regulating apoptosis, and constitutively activated STAT3
due to aberrantly produced growth cytokines or growth factors which
have been reported in a variety of malignant tumors, including
bladder cancer (4,8–14).
Moreover, activation of STAT3 leads to increased invasiveness of
cancer and constitutive activation of STAT3 is associated with
advanced cancer stage and metastatic disease (9,14–16).
Given these findings, we postulated that STAT3 could be a novel
therapeutic target for bladder cancer and in the present study
examined whether the STAT3 inhibitor WP1066 exhibited antitumor
activity against bladder cancer.
Material and methods
Cell culture and reagents
Human bladder cancer cell lines T24 and UMUC-3
[American Type Culture Collection (ATCC) Manassas, VA, USA] were
maintained in RPMI-1640 supplemented with 10% fetal bovine serum at
37°C in a 5% CO2 atmosphere. WP1066 (Calbiochem, La
Jolla, CA, USA) used in the present study was dissolved in 50 mM of
dimethyl sulfoxide (DMSO) and stored at −20°C. The antibodies used
in the present study were antibodies against phospho-STAT3
(p-STAT3, 1:1,000), STAT3 (1:1,000), phospho-ERK (p-ERK, 1:1,000),
ERK (1:1,000), cleaved caspase-3 (1:1,000), cleaved PARP (1:1,000),
Bcl-2 (1:1,000), Bcl-xL (1:1,000) (all from Cell Signaling
Technology, Inc., Danvers, MA, USA), and β-actin (1:3,000; Chemicon
International, Inc., Temecula, CA, USA).
Horseradish-peroxidase-conjugated secondary antibodies and an
enhanced chemiluminescence system were obtained from Amersham
Pharmacia Biotech (Piscataway, NJ, USA).
Western blot analysis
Cells that had been treated with the indicated
concentrations of WP1066 or the corresponding amount of DMSO for
the indicated time-points were lysed in RIPA buffer (10 mM
Tris-HCl, 150 mM NaCl, 1% Triton X-100, 5 mM EDTA, 1% sodium
deoxycholate, 0.1% SDS, 1.2% aprotinin, 5 µM leupeptin, 4 µM
antipain, 1 mM phenylmethylsulfonyl fluoride and 0.1 mM
Na3VO4). Equal amounts of the resulting
lysates were separated using 10% SDS-PAGE and transferred to
nitrocellulose membranes. The membranes were blocked with a
solution containing 5.0% skim milk, and then incubated with a
primary antibody overnight at 4°C. They were then incubated with a
secondary antibody coupled to horseradish peroxidase (Amersham,
Arlington Heights, IL, USA). The reactive proteins were visualized
using enhanced chemiluminescence according to the manufacturer's
recommendations. The relative intensities of protein expression
were calculated using the ImageJ program.
Cell counts and cell viability
assays
Following incubation overnight in 6-well plates
(5×104 cells/well, in triplicate), the cells were
treated with WP1066 at the indicated concentrations. The total
number of cells in 3 independent wells in each group were counted
at the indicated time-points after incubation using a
hemocytometer, and the mean value of 4 fields was recorded. The
results were expressed as the mean ± SE of the total number of
cells in each well in each group. Cell viability was assessed by
MTS assay using the CellTiter 96 AQueous Non-Radioactive
Cell Proliferation Assay (Promega, Madison, WI, USA) according to
the manufacturer's instructions. Briefly, the cells were incubated
overnight in 96-well plates (1.5×104 cells/well), and
were then treated for 24 h with the indicated concentrations of
WP1066 or with the corresponding amount of WP1066-free DMSO. Two
hours after adding MTS the plates were read at 490 nm on a
microplate autoreader. The results were expressed as the mean
optical density (OD) of the 6-well set for each group, and the
plates were assessed twice with similar results.
Cell cycle analysis and determination
of apoptosis
Cells were incubated for 24 h with the indicated
concentrations of WP1066 or the corresponding amount of DMSO.
Propidium-iodide stained nuclear fractions were obtained and the
cell cycle data were acquired using a flow cytometer with CellQuest
software (Becton-Dickinson, Heidelberg, Germany). Apoptotic cells
were assessed by double-staining with FITC-conjugated Annexin V and
propidium-iodide. This staining was carried out using the Annexin V
apoptosis detection kit (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA).
Wound healing assay
Cells were grown to confluence on 6-well tissue
culture plates and a wound was made by scraping the middle of the
cell monolayer with a P200 pipette tip. After floating cells were
removed by extensive washing with ice-cold phosphate-buffered
saline (PBS), fresh complete medium containing WP1066 or the
corresponding amount of DMSO was added. Migration and cell movement
throughout the wound area were examined after 12 h. The ImageJ
program was used to estimate and compare the covered areas
(17).
Matrigel invasion assay
An invasion assay was performed using
Matrigel-coated Transwell inserts (BD Biosciences, San Diego, CA,
USA) with 8-µm pores in 24-well plates. Briefly, a suspension of
1×105 cells in 500 µl serum-free medium was added to the
insert, and 750 µl serum-free medium supplemented with WP1066 or
the corresponding amount of DMSO was added to the bottom of the
well. After the plates were incubated for 12 h at 37°C, the inserts
were fixed in methanol, the filters were stained with 1% toluidine
blue in 1% borax, and in 3 independent experiments the number of
cells that invaded through the Matrigel-coated Transwell inserts
was counted in at least 10 fields/well.
Gelatin zymography
Gelatinolytic activity of MMP-2 and MMP-9 was
analyzed by gelatin gel zymography using a commercially available
kit (code no. AK45; Cosmo Bio Co., Ltd., Tokyo, Japan). In brief,
following incubation overnight in 6-well plates (5×104
cells/well), the cells were treated with WP1066 at the indicated
concentrations in serum-free medium for 12 h, and then the
conditioned medium was collected. Equal amounts of protein from the
medium were separated on pre-casted gels containing gelatin. The
gels were washed in denaturing buffer for 60 min at room
temperature and then incubated at 37°C overnight in developing
buffer. They were then stained with 0.25% Coomassie brilliant blue
R-250 for 4 h at room temperature and destained in distilled water
containing 30% methanol and 10% glacial acetic acid. The areas
where the gelatin substrate had been degraded by gelatinases
developed into white lines on a dark background. The relative
intensities of gelatinolytic activity were calculated using the
ImageJ program.
Statistical analysis
Results are expressed as the mean ± SE for 3
independent experiments. Variables for different groups were
compared using Student's t-test or analysis of variance (ANOVA),
and P<0.05 was considered to indicate a statistically
significant result.
Results
WP1066 inactivates STAT3 in bladder
cancer cells
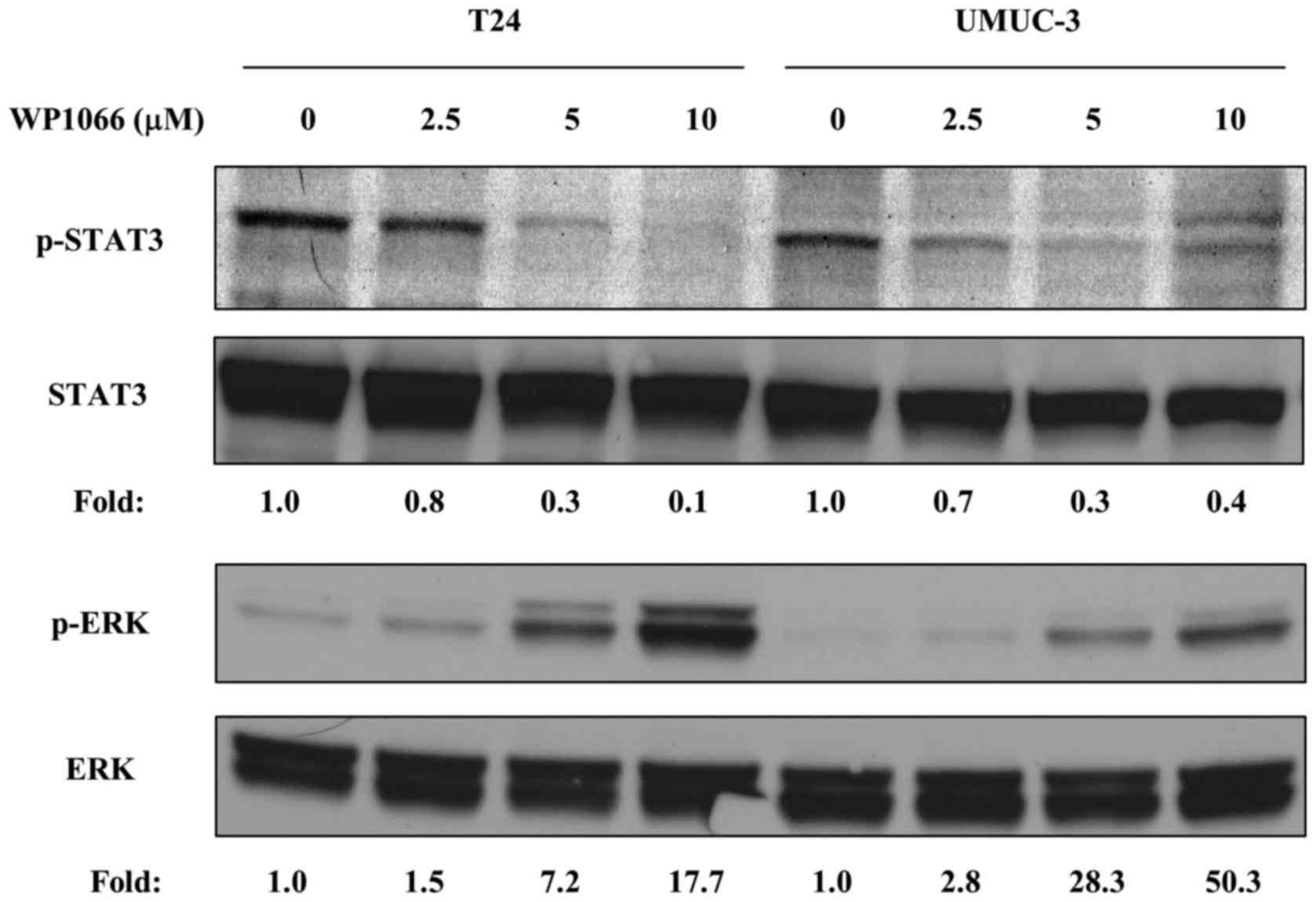
We first tested whether WP1066 inactivated STAT3 in
bladder cancer cells. It has been reported that 5–10 µM WP1066
inhibited STAT3 phosphorylation effectively in other cancer cell
lines (18,19), thus, we incubated T24 and UMUC-3
cells with 2.5, 5 and 10 µM WP1066 for 24 h and examined the
phosphorylation of STAT3 at Tyr705. WP1066 inhibited STAT3
phosphorylation in both cell lines in a dose-dependent manner, and
10 µM WP1066 completely abolished it (Fig. 1). Conversely, phosphorylation of
ERK, which often acts contrary to STAT3 (18,20,21),
was instead increased by WP1066 treatment in a dose-dependent
manner (Fig. 1).
WP1066 suppresses bladder cell
viability and proliferation by inducing apoptosis
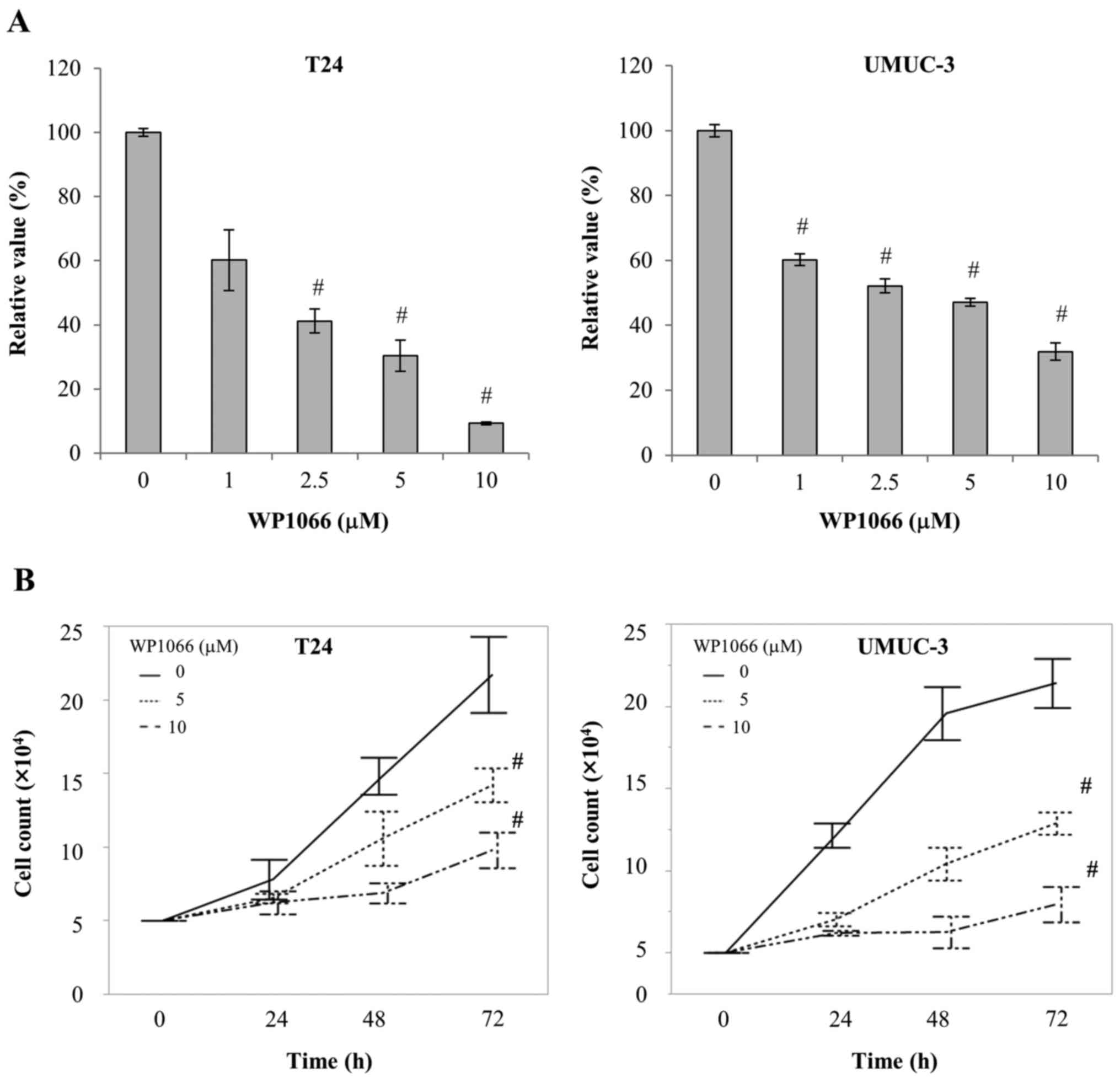
We next examined the ways in which the viability of
bladder cancer cells was affected when STAT3 was inhibited by
WP1066. We treated T24 and UMUC-3 cells with 1–10 µM WP1066 for 24
h, and examined their viability by MTS assay. WP1066 significantly
suppressed the viability of both T24 and UMUC-3 cells, the former
at concentrations >2.5 µM and the latter at concentrations >1
µM (P<0.001; Fig. 2A). In
addition, the number of cells present after treatment with 5 or 10
µM WP1066 for 72 h was significantly smaller than that present
after incubation without WP1066 (P<0.001; Fig. 2B). We further examined the effect of
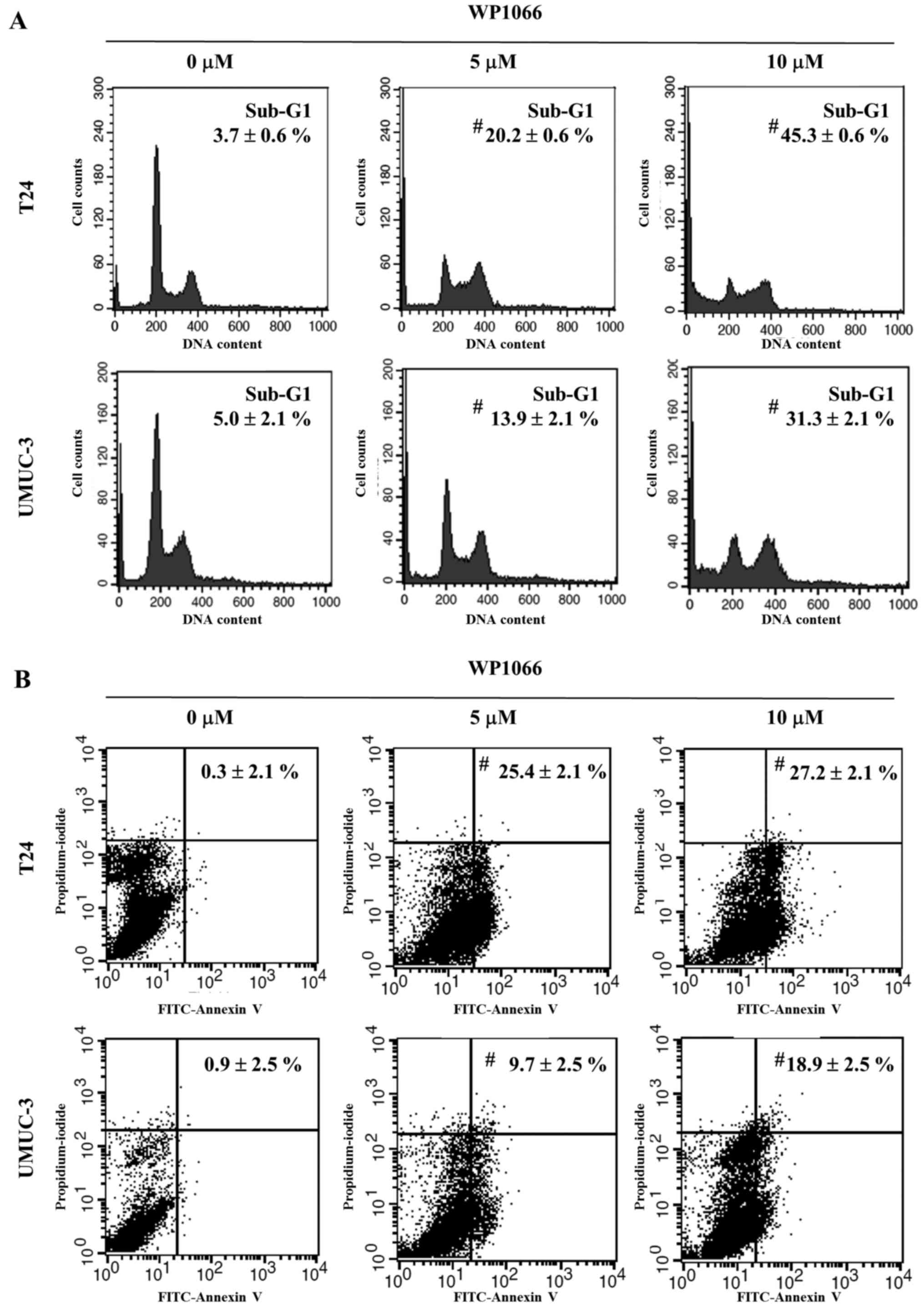
WP1066 on cell cycle progression by flow cytometry. T24 and UMUC-3
cells were treated with 5 or 10 µM WP1066 for 24 h, and their cell
cycles were analyzed. In both cell lines the sub-G1 population,
which indicates the percentage of apoptotic cells, was
significantly increased by WP1066 (P<0.001 in both cell lines;
Fig. 3A). To confirm the induction
of apoptosis by WP1066 treatment, T24 and UMUC-3 cells incubated
for 24 h with 5 or 10 µM WP1066 were double-stained with Annexin V
and propidium iodide and analyzed by flow cytometry. The percentage
of the population that was Annexin-positive and propidium
iodide-negative, which are apoptotic cells, was significantly
greater for cells treated with WP1066 than for control cells
(P<0.001 in both cell lines; Fig.
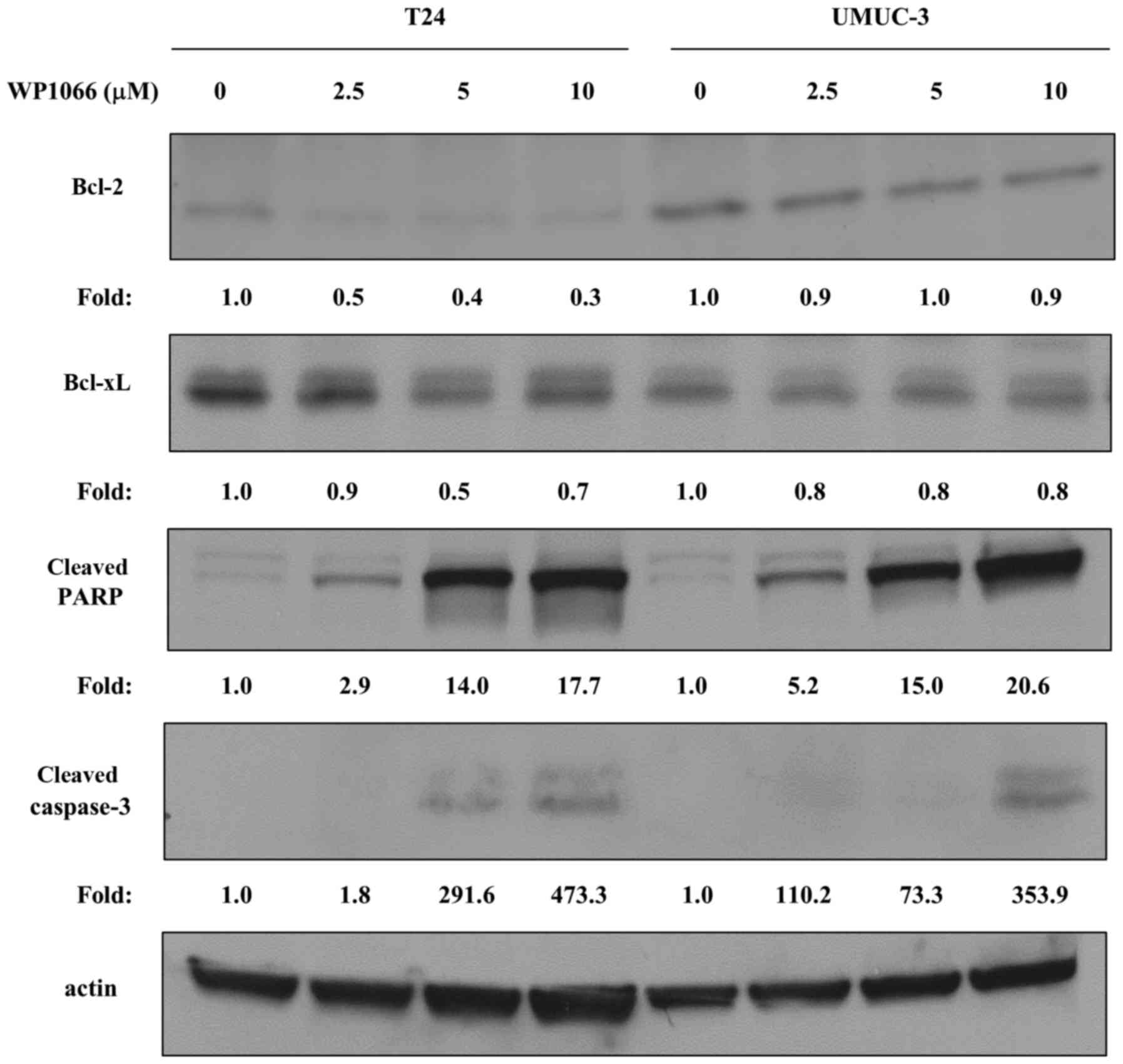
3B). STAT3 has been shown to regulate the expression of
apoptosis-related proteins such as Bcl-2 and Bcl-xL (22,23).
We further examined the effects of WP1066 on these
apoptosis-related molecules and found that the Bcl-2 and Bcl-xL in
T24 cells were decreased with treatment of WP1066, however they
were not altered in UMUC-3 cells (Fig.
4). In contrast, cleaved caspase-3 and cleaved PARP were
evident in both T24 and UMUC-3 cells treated with WP1066, which is
also evidence that WP1066 induced apoptosis (Fig. 4).
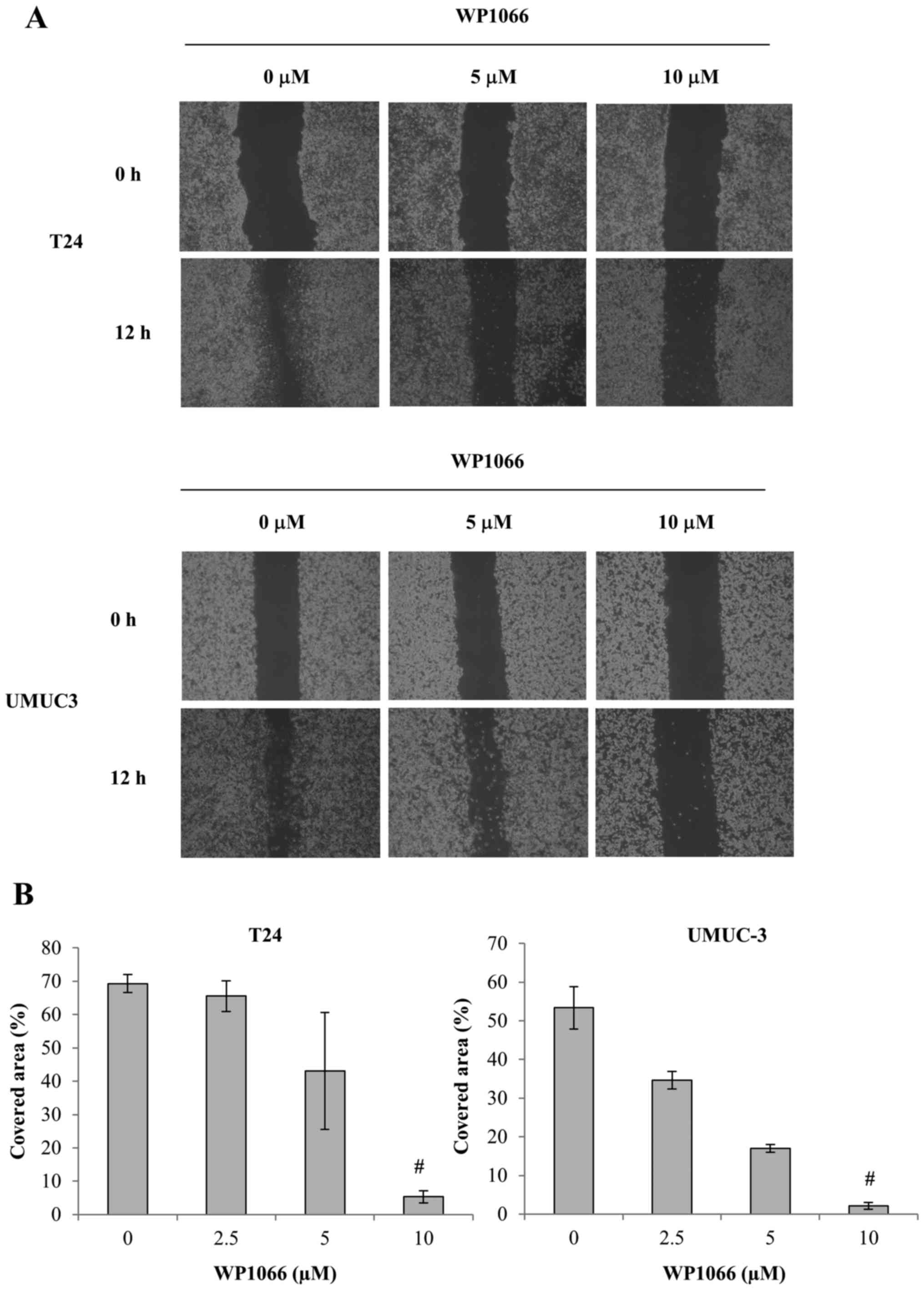
WP1066 decreases the motility and
invasiveness of bladder cancer cells
Previous studies have shown that STAT3 promotes cell
motility and invasiveness by increasing the activation of matrix
metalloproteinases (MMPs), including MMP-2 and MMP-9 (24,25).
To assess the effect of WP1066 on the motility and invasiveness of
bladder cancer cells, we examined wound healing and used Matrigel
invasion assays. Monolayers of T24 and UMUC-3 cells were scraped
with the tip of a sterile pipette to create a uniform wound, and
then treated with DMSO or 5 or 10 µM WP1066 for 12 h before the
covered area was assessed. We chose this treatment time since no
significant decrease in the total number of cells was found for
either T24 or UMUC-3 cells until 12 h after WP1066 treatment. In
both cell lines, however, the covered area was significantly
smaller for WP1066-treated cells than control cells (Fig. 5). In addition, the Matrigel invasion
assay demonstrated that the number of T24 and UMUC-3 cells invading
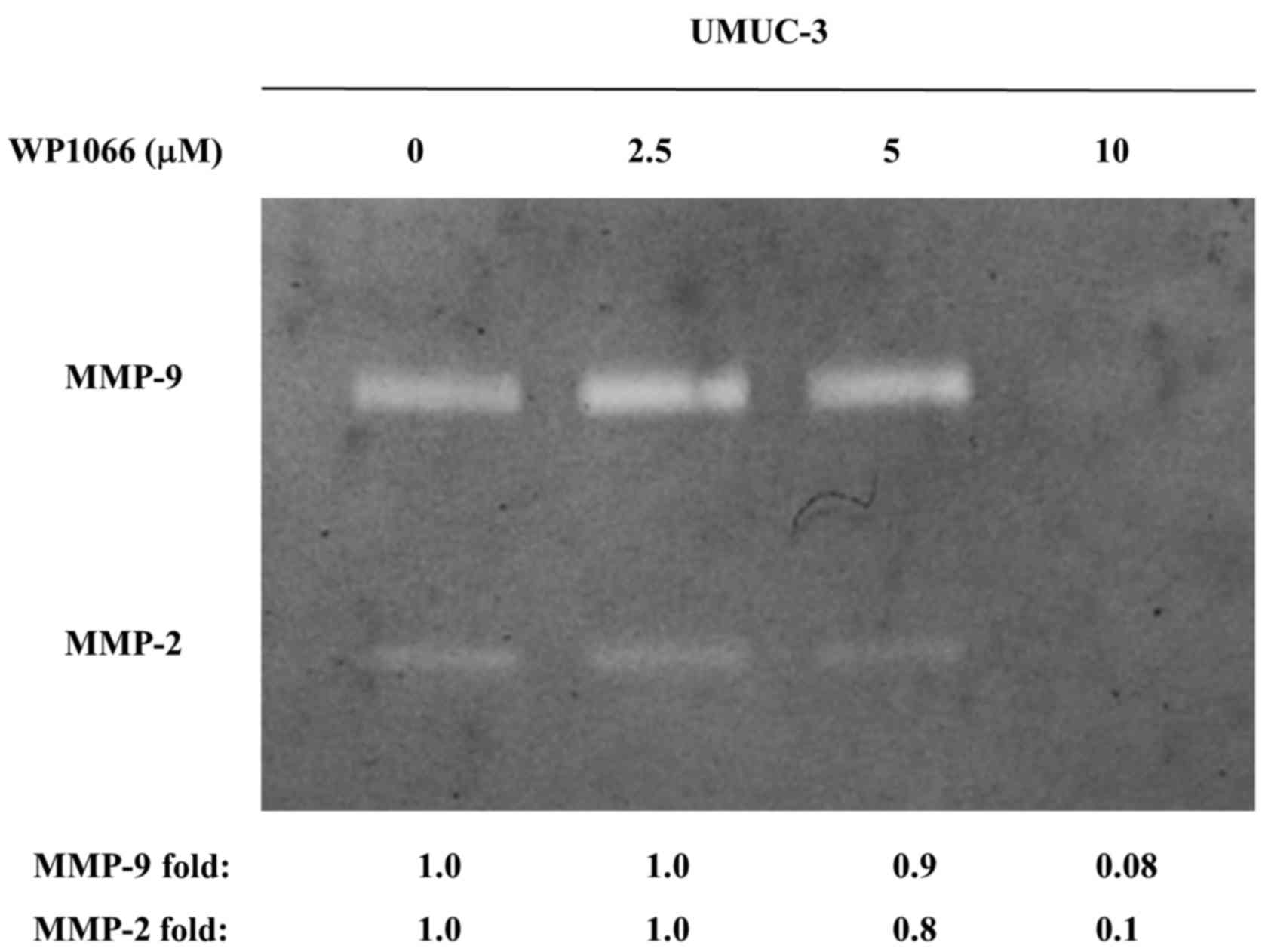
through the chamber was significantly decreased by WP1066 (Fig. 6). We further examined the effect of
WP1088 treatment on the activities of MMP-2 and MMP-9 by gelatin
zymography. Treatment with 10 µM WP1066 inhibited the activity of
MMP-2 and MMP-9 completely in UMUC-3 cells (Fig. 7). These results revealed that WP1066
decreased the motility and invasiveness of bladder cancer cells by
inhibiting MMP activity.
Discussion
In various types of malignant tumors, including
bladder cancer, activation of STAT3 is aberrantly increased and has
been reported to be associated with tumor aggressiveness and poor
clinical outcome (7,26,27).
Degoricija et al examined STAT3 gene expression in 36
bladder cancer tissues and found that it was upregulated in 27
(75%) (26). Chen et al
examined STAT3 activation in 100 bladder cancer tissues by
immunohistochemistry using a phospho-specific STAT3 antibody that
recognizes only activated STAT3 and found increased staining in 19
of them (13). In addition, STAT3
has recently been shown to be an important factor for development
of invasive bladder cancer (14).
STAT3 transgenic mice exposed to nitrosamine developed invasive
cancer from carcinoma in situ (CIS) directly, circumventing
the non-invasive papillary stage (14). These studies indicated that STAT3
was deeply implicated in the aggressiveness and progression of
bladder cancer. Thus inhibitors of the STAT3 signaling pathway
should have enormous potential in the treatment of bladder cancer.
In the study reported herein, we investigated the pharmacological
antitumor efficacy of the STAT3 inhibitor WP1066 against bladder
cancer cells in terms of their proliferation and invasiveness.
WP1066 is one of the commercially available STAT3
inhibitors and has been shown to display a potent inhibitory
activity against the upstream protein tyrosine kinase JAK2
(19). Iwamaru et al
synthesized WP1066 by modifying the structure of AG490, a compound
selected by screening a group of tyrphostins screened for their
ability to block STAT3 activity (19). We previously demonstrated that
WP1066 inhibited STAT3 activity in renal cancer cells grown both
in vitro and in vivo, with a low concentration (2.5
µM) of WP1066, significantly inhibiting STAT3 activity and cell
viability (18). The present study
revealed that the same concentration of WP1066 also significantly
inhibited STAT3 activity and cell viability significantly in both
T24 and UMUC3 bladder cancer cells. In contrast, WP1066 application
resulted in a reciprocal increase in the activities of ERKs 1 and
2, the kinases responsible for activating the ras/MAP kinase
signaling pathway. A similar observation with respect to ERK
activity has been made in cell lines derived from renal cell
carcinoma, gastric cancer and glioma (18,28,29).
ERK activation induced by WP1066 may compromise the antitumor
activity of STAT3 inhibition in bladder cancer, as it is a key
mitotic signal and promotes proliferation and invasiveness
(30).
STAT3 promotes the proliferation of cancer cells by
upregulating the expression of the antiapoptotic proteins Bcl-2 and
Bcl-xL, and inhibiting it inhibits cancer cell proliferation by
downregulating their expression (11,21).
Chen et al reported that interrupting the STAT3 signaling
pathway in UMUC3 bladder cancer cells using an adenovirus-mediated
dominant-negative STAT3 resulted in cell growth inhibition and
apoptosis with downregulation of Bcl-2 and Bcl-xL (13). Our results revealing growth
inhibition and induction of apoptosis by pharmacological
attenuation of STAT3 activity with downregulation of Bcl-2 and
Bcl-xL expression in T24 cells was consistent with previous studies
(22,23). Another important role of STAT3 in
cancer biology is regulating migration and invasive activities
(16,31). Ito et al reported that STAT3
regulates the migration and invasion of T24 bladder cancer cells by
modulating the expression of matrix metalloproteinase-1 (31). In consistency with their study, the
migration and invasiveness of T24 and UMUC-3 bladder cancer cells
were decreased with treatment of WP1066. Although WP1066 decreased
the proliferation and invasiveness of bladder cancer cells, we
could not demonstrate the antitumor efficacy of WP1066 in
vivo. In a preliminary study we tried using a xenograft model
of bladder cancer cells to examine the antitumor activity of WP1066
in the same way as in our previous study with renal cancer cells
(18) but found no significant
tumor reduction (data not shown). This is one of the limitations of
the present study.
Although cisplatin-based chemotherapy is the
first-line treatment for metastatic bladder cancer, chemoresistance
remains a major clinical challenge. Activated STAT has been
suggested to confer resistance to cisplatin-based chemotherapy by
preventing apoptosis, and pharmacological inhibition of STAT3
activity using WP1066 or adenovirus-mediated STAT3 depletion has
been shown to potentiate the efficacy of cisplatin (32,33).
In addition, recent studies demonstrated the efficacy of molecular
targeted therapy using receptor tyrosine kinase inhibitors of the
epidermal growth factor receptor (EGFR) family against bladder
cancer (34). STAT3 activity has
been suggested to be responsible for the resistance to EGFR
inhibitors, and targeting STAT3 with a STAT3 decoy decreased
cellular viability in models of EGFR inhibitor resistance (35). These results revealed that WP1066
could be a potent agent combined with conventional or targeted
therapy and that efficacy of such combinations should be
investigated in the near future.
In conclusion, we revealed that the STAT inhibitor
WP1066 induces apoptosis of bladder cancer cells and inhibited
their viability and invasive activity. These results indicate that
using WP1066 to inhibit the STAT3 signaling pathway could be an
effective therapeutic strategy against bladder cancer.
References
|
1
|
Ploussard G, Shariat SF, Dragomir A, Kluth
LA, Xylinas E, Masson-Lecomte A, Rieken M, Rink M, Matsumoto K,
Kikuchi E, et al: Conditional survival after radical cystectomy for
bladder cancer: Evidence for a patient changing risk profile over
time. Eur Urol. 66:361–370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rosenberg JE, Carroll PR and Small EJ:
Update on chemotherapy for advanced bladder cancer. J Urol.
174:14–20. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
von der Maase H, Sengelov L, Roberts JT,
Ricci S, Dogliotti L, Oliver T, Moore MJ, Zimmermann A and Arning
M: Long-term survival results of a randomized trial comparing
gemcitabine plus cisplatin, with methotrexate, vinblastine,
doxorubicin, plus cisplatin in patients with bladder cancer. J Clin
Oncol. 23:4602–4608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van Kessel KE, Zuiverloon TC, Alberts AR,
Boormans JL and Zwarthoff EC: Targeted therapies in bladder cancer:
An overview of in vivo research. Nat Rev Urol. 12:681–694. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schindler C and Darnell JE Jr:
Transcriptional responses to polypeptide ligands: The JAK-STAT
pathway. Annu Rev Biochem. 64:621–651. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Akira S, Nishio Y, Inoue M, Wang XJ, Wei
S, Matsusaka T, Yoshida K, Sudo T, Naruto M and Kishimoto T:
Molecular cloning of APRF, a novel IFN-stimulated gene factor 3
p91-related transcription factor involved in the gp130-mediated
signaling pathway. Cell. 77:63–71. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu H, Lee H, Herrmann A, Buettner R and
Jove R: Revisiting STAT3 signalling in cancer: New and unexpected
biological functions. Nat Rev Cancer. 14:736–746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bruserud Ø, Nepstad I, Hauge M, Hatfield
KJ and Reikvam H: STAT3 as a possible therapeutic target in human
malignancies: Lessons from acute myeloid leukemia. Expert Rev
Hematol. 8:29–41. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Horiguchi A, Oya M, Shimada T, Uchida A,
Marumo K and Murai M: Activation of signal transducer and activator
of transcription 3 in renal cell carcinoma: A study of incidence
and its association with pathological features and clinical
outcome. J Urol. 168:762–765. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim HS, Park YH, Lee J, Ahn JS, Kim J,
Shim YM, Kim JH, Park K, Han J and Ahn MJ: Clinical impact of
phosphorylated signal transducer and activator of transcription 3,
epidermal growth factor receptor, p53, and vascular endothelial
growth factor receptor 1 expression in resected adenocarcinoma of
lung using tissue microarray. Cancer. 116:676–685. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Masuda M, Suzui M, Yasumatu R, Nakashima
T, Kuratomi Y, Azuma K, Tomita K, Komiyama S and Weinstein IB:
Constitutive activation of signal transducers and activators of
transcription 3 correlates with cyclin D1 overexpression and may
provide a novel prognostic marker in head and neck squamous cell
carcinoma. Cancer Res. 62:3351–3355. 2002.PubMed/NCBI
|
|
12
|
Takemoto S, Ushijima K, Kawano K,
Yamaguchi T, Terada A, Fujiyoshi N, Nishio S, Tsuda N, Ijichi M,
Kakuma T, et al: Expression of activated signal transducer and
activator of transcription-3 predicts poor prognosis in cervical
squamous-cell carcinoma. Br J Cancer. 101:967–972. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen CL, Cen L, Kohout J, Hutzen B, Chan
C, Hsieh FC, Loy A, Huang V, Cheng G and Lin J: Signal transducer
and activator of transcription 3 activation is associated with
bladder cancer cell growth and survival. Mol Cancer. 7:782008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ho PL, Lay EJ, Jian W, Parra D and Chan
KS: Stat3 activation in urothelial stem cells leads to direct
progression to invasive bladder cancer. Cancer Res. 72:3135–3142.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun Y, Cheng MK, Griffiths TR, Mellon JK,
Kai B, Kriajevska M and Manson MM: Inhibition of STAT signalling in
bladder cancer by diindolylmethane: Relevance to cell adhesion,
migration and proliferation. Curr Cancer Drug Targets. 13:57–68.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang N, Duan WD, Leng JJ, Zhou L, Wang X,
Xu YZ, Wang XD, Zhang AQ and Dong JH: STAT3 regulates the migration
and invasion of a stem-like subpopulation through microRNA-21 and
multiple targets in hepatocellular carcinoma. Oncol Rep.
33:1493–1498. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schneider CA, Rasband WS and Eliceiri KW:
NIH Image to ImageJ: 25 years of image analysis. Nat Methods.
9:671–675. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Horiguchi A and Asano T, Kuroda K, Sato A,
Asakuma J, Ito K, Hayakawa M, Sumitomo M and Asano T: STAT3
inhibitor WP1066 as a novel therapeutic agent for renal cell
carcinoma. Br J Cancer. 102:1592–1599. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Iwamaru A, Szymanski S, Iwado E, Aoki H,
Yokoyama T, Fokt I, Hess K, Conrad C, Madden T, Sawaya R, et al: A
novel inhibitor of the STAT3 pathway induces apoptosis in malignant
glioma cells both in vitro and in vivo. Oncogene. 26:2435–2444.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Horiguchi A, Asano T, Asano T, Ito K,
Sumitomo M and Hayakawa M: Pharmacological inhibitor of fatty acid
synthase suppresses growth and invasiveness of renal cancer cells.
J Urol. 180:729–736. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Horiguchi A, Oya M, Marumo K and Murai M:
STAT3, but not ERKs, mediates the IL-6-induced proliferation of
renal cancer cells, ACHN and 769P. Kidney Int. 61:926–938. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bhattacharya S, Ray RM and Johnson LR:
STAT3-mediated transcription of Bcl-2, Mcl-1 and c-IAP2 prevents
apoptosis in polyamine-depleted cells. Biochem J. 392:335–344.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chang Q, Bournazou E, Sansone P, Berishaj
M, Gao SP, Daly L, Wels J, Theilen T, Granitto S, Zhang X, et al:
The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and
metastasis. Neoplasia. 15:848–862. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou X, Ren Y, Liu A, Han L, Zhang K, Li
S, Li P, Li P, Kang C, Wang X, et al: STAT3 inhibitor WP1066
attenuates miRNA-21 to suppress human oral squamous cell carcinoma
growth in vitro and in vivo. Oncol Rep. 31:2173–2180.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang X, Sun Y, Pireddu R, Yang H, Urlam
MK, Lawrence HR, Guida WC, Lawrence NJ and Sebti SM: A novel
inhibitor of STAT3 homodimerization selectively suppresses STAT3
activity and malignant transformation. Cancer Res. 73:1922–1933.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Degoricija M, Situm M, Korać J, Miljković
A, Matić K, Paradžik M, Marinović Terzić I, Jerončić A, Tomić S and
Terzić J: High NF-κB and STAT3 activity in human urothelial
carcinoma: A pilot study. World J Urol. 32:1469–1475. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen RJ, Ho YS, Guo HR and Wang YJ: Rapid
activation of Stat3 and ERK1/2 by nicotine modulates cell
proliferation in human bladder cancer cells. Toxicol Sci.
104:283–293. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sai K, Wang S, Balasubramaniyan V, Conrad
C, Lang FF, Aldape K, Szymanski S, Fokt I, Dasgupta A, Madden T, et
al: Induction of cell-cycle arrest and apoptosis in glioblastoma
stem-like cells by WP1193, a novel small molecule inhibitor of the
JAK2/STAT3 pathway. J Neurooncol. 107:487–501. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Judd LM, Menheniott TR, Ling H, Jackson
CB, Howlett M, Kalantzis A, Priebe W and Giraud AS: Inhibition of
the JAK2/STAT3 pathway reduces gastric cancer growth in vitro and
in vivo. PLoS One. 9:e959932014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Monami G, Gonzalez EM, Hellman M, Gomella
LG, Baffa R, Iozzo RV and Morrione A: Proepithelin promotes
migration and invasion of 5637 bladder cancer cells through the
activation of ERK1/2 and the formation of a paxillin/FAK/ERK
complex. Cancer Res. 66:7103–7110. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Itoh M, Murata T, Suzuki T, Shindoh M,
Nakajima K, Imai K and Yoshida K: Requirement of STAT3 activation
for maximal collagenase-1c (MMP-1) induction by epidermal growth
factor and malignant characteristics in T24 bladder cancer cells.
Oncogene. 25:1195–1204. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han Z, Hong Z, Gao Q, Chen C, Hao Z, Ji T,
Hu W, Yan Y, Feng J, Liao S, et al: A potent oncolytic adenovirus
selectively blocks the STAT3 signaling pathway and potentiates
cisplatin antitumor activity in ovarian cancer. Hum Gene Ther.
23:32–45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou X, Ren Y, Liu A, Jin R, Jiang Q,
Huang Y, Kong L, Wang X and Zhang L: WP1066 sensitizes oral
squamous cell carcinoma cells to cisplatin by targeting
STAT3/miR-21 axis. Sci Rep. 4:74612014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mooso BA, Vinall RL, Mudryj M, Yap SA,
deVere White RW and Ghosh PM: The role of EGFR family inhibitors in
muscle invasive bladder cancer: A review of clinical data and
molecular evidence. J Urol. 193:19–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sen M, Joyce S, Panahandeh M, Li C, Thomas
SM, Maxwell J, Wang L, Gooding WE, Johnson DE and Grandis JR:
Targeting Stat3 abrogates EGFR inhibitor resistance in cancer. Clin
Cancer Res. 18:4986–4996. 2012. View Article : Google Scholar : PubMed/NCBI
|