Introduction
Lung cancer is the leading cause of death among
males worldwide from all cancers (1), and non-small cell lung cancer (NSCLC)
accounts for more than 85% of lung cancer cases; of which
adenocarcinoma (40%) is the most common subtype, followed by
squamous cell (25%) and large cell carcinoma (10%) (2). Chemotherapy is the most common
treatment for advanced-stage lung cancer patients. Such therapy is
generally performed with platinum-based chemotherapy. However, this
treatment provides only a modest benefit to survival.
Unsatisfactorily, the overall 5-year survival rate for all stages
of NSCLC is only 17% (3). The
unavoidable development of drug resistance is the most important
cause of treatment failure in patients subjected to chemotherapy.
When tumors become resistant to chemotherapeutic drugs, they
commonly develop cross-resistance to other anticancer drugs.
Therefore, choosing an ideal non-cross-resistant drug that improves
drug responsiveness and increases the overall survival rate has
become a critical issue in cancer management.
The taxanes, paclitaxel (PAX) and docetaxel (DOC),
are microtubule-stabilizing agents that function primarily by
interfering with spindle microtubule dynamics causing cell cycle
arrest and apoptosis (4). The
chemical statuses of the two taxanes are almost identical. DOC is
regarded as a second-generation taxane. The effects of DOC are
correlated with mitotic arrest and cellular toxicity (5). Currently, DOC is indicated for
first-line therapy in combination with a platinum compound and as
monotherapy in patients with NSCLC who experience failure during a
platinum-based regimen.
P-glycoprotein (P-gp) is the product of the MDR1
gene, which is also recognized as ABCB1. P-gp is a pump that
presents in normal tissues such as the gastrointestinal tract and
brain to prevent the accumulation of toxic substances (6). Overexpression of P-gp is thought to be
one of the most common mechanisms underlying resistance to taxanes
in cancer models (7).
Pemetrexed (PEM; LY231514) is a novel antifolate
drug that has been approved for first-line treatment of patients
with advanced non-squamous NSCLC in combination with cisplatin. PEM
is also used as a single agent for relapsed or chemotherapy
refractory NSCLC after platinum-containing chemotherapy (8). PEM is a unique folate antagonist that
inhibits thymidylate synthase (TS), dihydrofolate reductase (DHFR)
and the purine synthetic enzyme glycinamide ribonucleotide
formyltransferase (GARFT) as multitargeted antifolate (9,10).
Two adenocarcinoma cell lines (PC-9 and A549) with
PEM resistance were established and recently analyzed. TS and DHFR
were significantly increased in the 4 PEM-resistant A549 sublines
(11). Data obtained from patients
have also suggested that TS expression, rather than DHFR, may be an
important predictive factor of the treatment efficacy of PEM in
NSCLC (12). Furthermore, it has
been concluded by meta-analysis (13) that better responses usually appeared
in patients with a lower expression of TS with a significant
association between TS expression and outcomes of PEM-based
chemotherapy for NSCLC. Therefore, it is believed that upregulation
of TS gene expression may play an important role in PEM resistance
(14) and downregulation of TS may
increase PEM response.
Previously, we reported that DOC-selected A549
sublines (A549/D16 and A549/D32) expressed high levels of P-gp and
were cross-resistant to VCR and DXR (15). We further defined the
characteristics of these sublines with antifolate drugs (PEM and
MTX) in the present study. We found that only PEM overcame DOC
resistance and that TS in the de novo nucleic acid synthesis
pathway may be downregulated by wild-type TP53 in DOC-selected A549
sublines. We further demonstrated that TS was downregulated by
DOC-activated wild-type TP53 in two human lung cancer cell lines
(A549 and H460), however it was not downregulated in cells with
mutated TP53 and null-TP53. Therefore, according to our data, the
application sequence of DOC followed by PEM in patients with
wild-type TP53 status is suggested. This approach may lead to
better cancer control in NSCLC.
Materials and methods
Drugs and chemicals
PEM was provided by Eli Lilly Corporation
(Indianapolis, IN, USA). Doxorubicin (DXR; 44584), methotrexate
(MTX) hydrate (M8407) and vincristine sulfate salt (VCR; V8879)
were purchased from Sigma-Aldrich (St. Louis, MO, USA). The drugs
were dissolved in ddH2O (PEM, DXR and VCR), MTX was
dissolved in 10% dimethyl sulfoxide (DMSO) stored at −20°C, and was
diluted in culture medium immediately before use. DOC (10 mg) was
obtained from Aventis Pharmaceuticals Inc. (Bridgewater, NJ, USA)
and was dissolved in 11.2 ml DMSO and 20 ml absolute ethanol as a
375 µM stock.
Lung cancer cell lines
Human A549, H460 and H1355 cells were cultured at
37°C in Dulbecco's modified Eagle's medium (DMEM) supplemented with
10% fetal bovine serum (FBS), 1% NEAA, 1% sodium pyruvate, 1%
L-glutamine, 100 IU/ml penicillin and 100 mg/ml streptomycin. H1299
and CL1-0 cells were cultured in RPMI-1640 medium. The DOC
resistant sublines were established from parental cells in a
stepwise manner by exposure to increasing concentrations of DOC as
previously described (15). The
DOC-resistant sublines maintained at 16 and 32 nM of DOC are
denoted as A549/D16 and A549/D32, respectively.
Cytotoxicity assay (MTT assay)
Chemosensitivity to PEM was determined using an MTT
assay. The detailed steps of MTT assay have been previously
described (15). The cells were
exposed to various concentrations of PEM in fresh medium for 96 h.
Mean values were calculated from 3 independent experiments.
Protein extraction and western blot
analysis
Protein extracts were prepared as previously
described (15). Proteins (10–30
µg) transferred onto polyvinylidene fluoride (PVDF) membranes were
reacted with polyclonal anti-TS (Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA), anti-DHFR, FPGS, GGH and TK1 (GeneTex,
Irvine, CA, USA), anti-β-actin (NeoMarker, Fremont, CA, USA) and
anti-TP53 (Dako, Carpinteria, CA, USA) separately, followed by
conjugation of anti-rabbit (Santa Cruz Biotechnology, Inc.) or
anti-mouse (Calbiochem, La Jolla, CA, USA) IgG to horseradish
peroxidase. A chemiluminescence detection kit (ECL; GE Healthcare
Bioscience, Amersham Place, UK) was used to determine the levels of
protein expression. The Image J quantity software was downloaded
from the National Institutes of Health to measure the intensity of
each blot.
Clonogenic cell survival assay
Cells were seeded in 6-well plates (150
cells/plate). After 24 h of incubation, the cells were treated with
respective doses of DOC (48 h) or PEM (96 h) and eventually
cultured for 10 days. The colonies formed were fixed with ice-cold
methanol for 30 min and then stained with 20% Giemsa. Survival
fractions were calculated by normalization to the appropriate
control groups.
Cell cycle assay by flow
cytometry
For the cell cycle analysis, cells
(3×105) were seeded in 10-cm plates and incubated for 24
h. The cells were exposed to PEM (0–110 nM) with fresh medium for
96 h at 37°C. Cells were trypsinized and washed, then resuspended
in 75% ice-cold ethanol (75%) overnight at 4°C. This was followed
by washing with 1X PBS and centrifugation; the cell pellets were
gently dispersed with 300 µl of 1X PBS with 30 µl of RNase A (10
mg/ml) for 30 min. Propidium iodide (PI) (10 µl; 500 µg/ml) and 300
µl of 1X PBS were added followed by another 30 min incubation at
room temperature. Subsequently the cells were filtered by a nylon
mesh (40 µm), and then analyzed with a FACSCalibur flow cytometer
(BD Biosciences, San Jose, CA, USA).
TS, GGH cloning and expression
To clone the human TS ORF for expression, the total
RNA of H1299 cells was reverse-transcribed into cDNA by the
MultiScribe reverse transcription system (Applied Biosystems,
Foster City, CA, USA). The forward primer
(5′-TCTCGCTAGCATGCCTGTGGCCGGCTCGGA) and the reverse primer
(5′-TCTCGTTTAAACCTAAACAGCCATTTCCATTT) were applied to amplify the
ORF of TS by PCR. The obtained PCR product was digested with
NheI and PmeI, and then purified. The ORF fragment
was ligated into the lentiviral vector pLAS3w.Ppuro, which was
provided by the National RNAi Core Facility Platform (Academia
Sinica, Taiwan). The forward primer
(5′-TCTCGCTAGCATGGCCAGTCCGGGCTGCCT) and the reverse primer
(5′-TCTCCCTGCAGGTCAATCAAATATGTAACATT) for GGH were also used to
clone the gene into the same vector, followed by NheI and
SbfI digestion and ligation. A549 and A549/D16 cells that
were seeded in 6-cm plates were transduced with the lentivirus
carrying TS, GGH or empty vector as control to express TS or GGH
for 48 h. The transduced cells were harvested and seeded into
24-well plates for MTT assay as previously described.
Mice xenograft models
Experimental procedures and handling were conducted
in accordance with the international guidelines for laboratory
animals and all efforts were made to minimize suffering. The
present study was approved by the Chung Shan Medical University
Animal Care Committee (permit no. 1238). To establish A549/D16
tumor xenografts, 4-week-old male (NOD.CB17-Prkdcscid/IcrCrlBltw)
mice purchased from BioLASCO Taiwan Co., Ltd. (Taipei, Taiwan) were
subcutaneously injected with 5×106 cells (100 µl in
serum-free DMEM) plus 100 µl Matrigel (354234; BD Biosciences).
Animals were fed a low folic acid diet [AIN-93M folate deficient
diet w/1% succinylsulfathiazole from TestDiet (St. Louis, MO,
USA)], and then randomly divided into 3 groups consisting of 4
animals each (PBS, DOC and PEM groups). Thirty days after tumors
has been established, the mice were injected with DOC (7.5 mg/kg),
PEM (100 mg/kg) and PBS at day-30 and −37 intraperitoneally. The
tumor sizes were assessed every 3 days following drug injection and
tumor volume was calculated by the following formula: 0.5 × larger
diameter (mm) × small diameter2 (mm).
Statistical analysis
All values are presented as the mean ± SD. Data were
compared among groups using a t-test and p<0.05 was considered
to indicate a statistically significant result.
Results
DOC-selected multi-drug resistant A549
cells have high PEM sensitivity
According to an MTT sensitivity assay, the
established A549/D16 and A549/D32 sublines revealed their drug
sensitivities (Table I) in terms of
IC50 (inhibition concentration) values compared with the
parental A549 cells. The A549/D16 and A549/D32 sublines revealed a
109.1- and 286.59-fold resistance to DOC, respectively, when
compared with the parental cells. Both sublines also exhibited
cross-resistance to VCR and DXR. To investigate whether DOC
resistance may result in cross-resistance to antimetabolic
chemotherapy, the sensitivities to PEM and methotrexate (MTX) were
also determined. Notably, only minimal resistances were detected
with MTX treatment. For example; the A549/D16 and A549/D32 had a
1.8- and 2.0-fold resistance to MTX, respectively. Moreover, the
A549/D16 and A549/D32 sublines had a 0.31- and 0.29-fold resistance
to PEM, respectively, indicating DOC-resistant A549 sublines have
higher PEM sensitivity.
 | Table I.Characterization of drug
sensitivities with DOC-resistant A549 sublines by MTT assay. |
Table I.
Characterization of drug
sensitivities with DOC-resistant A549 sublines by MTT assay.
|
| IC50 ±
SDa (nmol/l) |
|---|
|
|
|
|---|
| Drug (nM) | A549 | A549/D16 | A549/D32 |
|---|
| Docetaxel |
7.6±1.8
(1.0) |
829.2±28.6
(109.1) |
2177.9±21.2
(286.5) |
| Vincristine |
20.0±2.2
(1.0) |
690.1±25.7
(34.5) |
811.0±27.4
(40.5) |
| Doxorubicin |
316.7±14.0
(1.0) |
3,468.3±29.1 (10.9) |
8,028.0±24.1 (25.3) |
| Pemetrexed |
1304.7±94.7
(1.0) |
187.7±2.5
(0.1) |
120.3±8.6
(0.1) |
| Methotrexate |
48.7±0.6
(1.0) |
87.0±1.7
(1.8) |
95.0±1.0
(2.0) |
PEM treatment decreases the growth of
DOC-resistant A549 sublines according to a clonogenic survival
assay
The DOC-resistant A549 sublines exhibited a
significant sensitivity to PEM. To further characterize the
cytotoxic effects of DOC and PEM to A549 and DOC-resistant
sublines, we applied a clonogenic assay to determine the ability of
these cells to clonally expand and create colonies. After 48 h of
DOC exposure, the cells were cultured for 10 days and the colonies
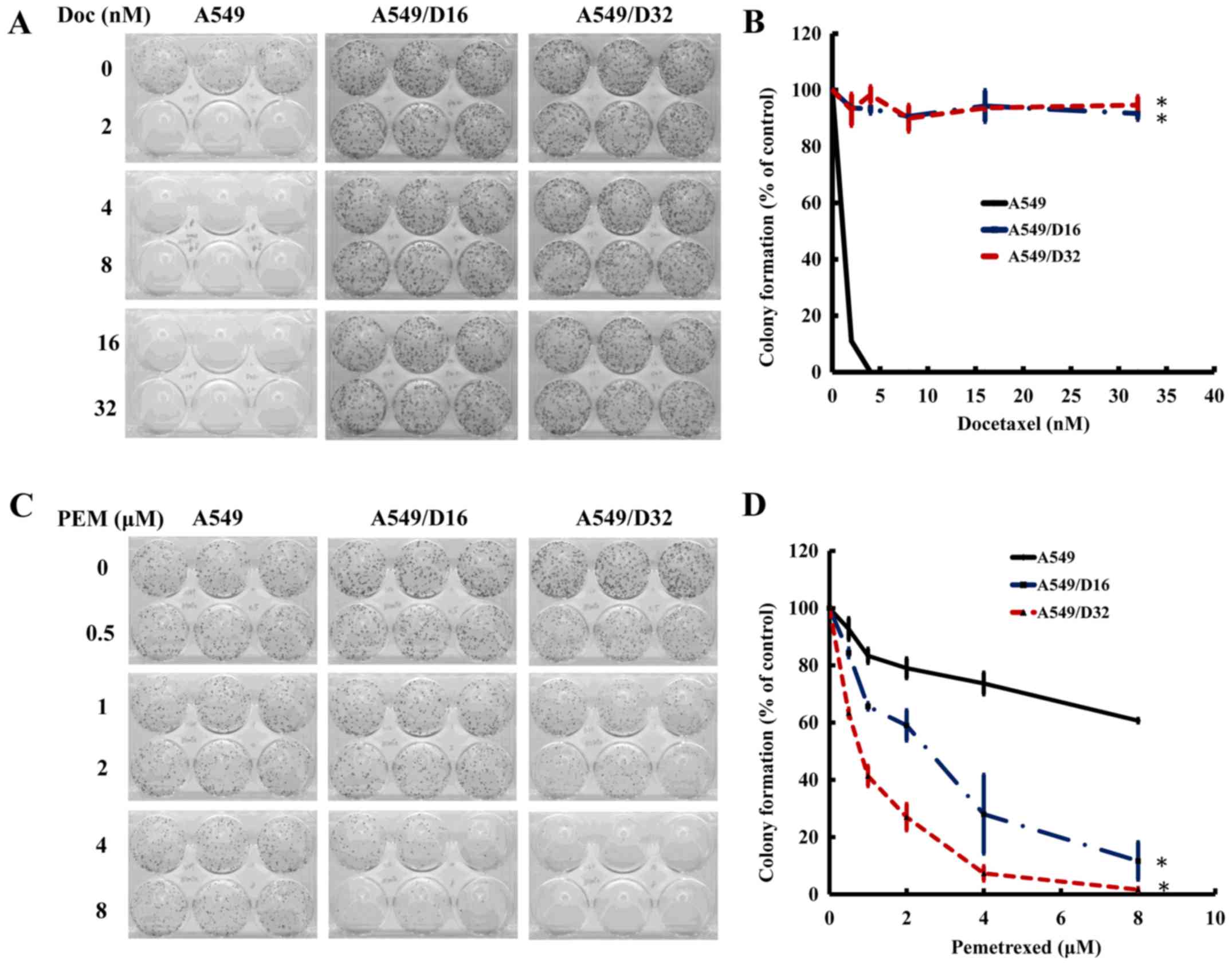
were photographed (Fig. 1A). The
survival percentage was calculated as shown in Fig. 1B. None of the A549 cells survived
under 16 nM of DOC treatment, but >80% of A549/D16 and A549/D32
cells were still able to proliferate; this confirms the DOC
resistance of these sublines. When the cells were exposed to PEM
for 96 h and maintained in fresh medium for 10 days (Fig. 1C), 33% of A549 cells survived at the
PEM concentration of 600 nM. In contrast, only 5 and 4% of A549/D16
and A549/D32 cells remained, respectively (Fig. 1D). According to these data, we
concluded that DOC-resistant A549 cells have a high sensitivity to
PEM toxicity in vitro.
Downregulation of TS in the de novo
DNA synthesis pathway in DOC-resistant A549 cells contributes to
high PEM sensitivity
PEM is a multi-target folate synthesis inhibitor
(9) and analysis has shown that
high TS expression results in PEM resistance (13,16).
Therefore, it would be logical to assume that high sensitivity to
PEM may result from folate synthesis deficiency in DOC-resistant
A549 cells. The enzymes of TS and DHFR are associated with de
novo synthesis and TK1 is associated with the salvage pathway
of DNA synthesis (17). The results
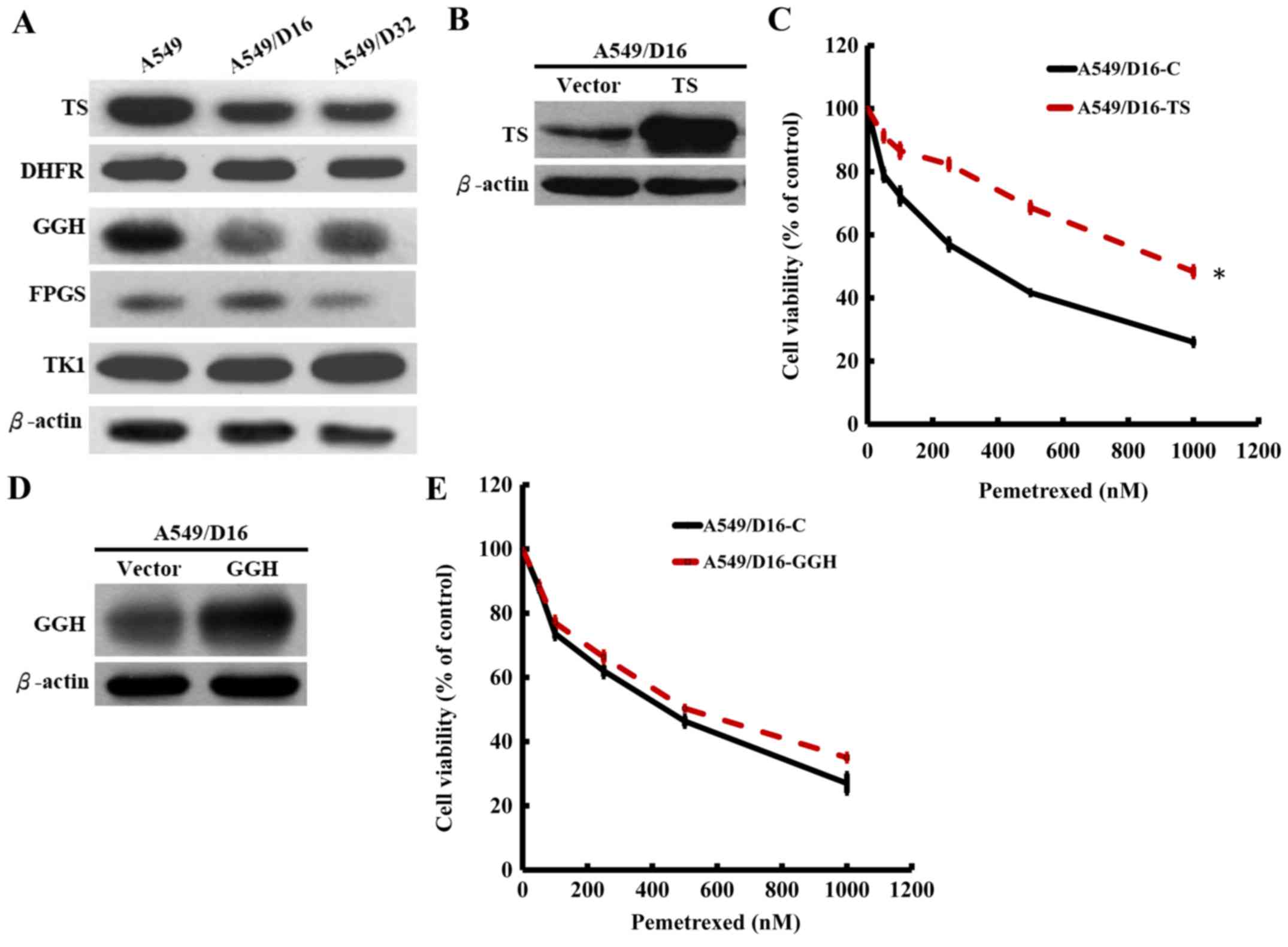
from the western blot analysis revealed that only the TS protein
level was significantly decreased in the DOC-resistant A549
sublines, but not the DHFR and TK1 protein levels when compared
with the parental A549 cells (Fig.
2A). PEM is polyglutamated to active pentaglutamide
intracellularly by a reaction catalyzed by folylpolyglutamate
synthase (FPGS). The enzyme counteracting antifolate
polyglutamylation is γ-glutamyl hydrolase (GGH) (9). The levels of GGH were also decreased
in both sublines with slightly decreased FPGS levels observed. The
data indicated that only TS in de novo DNA synthesis was
downregulated in DOC-resistant A549 cells and that the low level of
GGH may also contribute to PEM sensitivity by insufficiently
removing polyglutamylated PEM. Therefore, we subcloned TS and GGH
genes and constructed a lentiviral expression system for
overexpression. To compare the PEM sensitivities, the
TS-overexpressed A549/D16 and vector-control A549/D16 cells were
treated with respective doses of PEM and analyzed by MTT assay.
When TS was overexpressed in A549/D16 cells (Fig. 2B), the PEM sensitivity was
significantly decreased (Fig. 2C).
We also investigated the overexpression effect of GGH (Fig 2D) on PEM sensitivity, but
insignificant results were obtained (Fig. 2E). These data demonstrated that
downregulated TS is the major marker related to DOC resistance in
A549 cells.
Lower concentration of PEM induces
apoptotic death in vitro and upregulation of TS, TP53, p21 and
phosphorylated ERK in DOC-resistant A549 cells
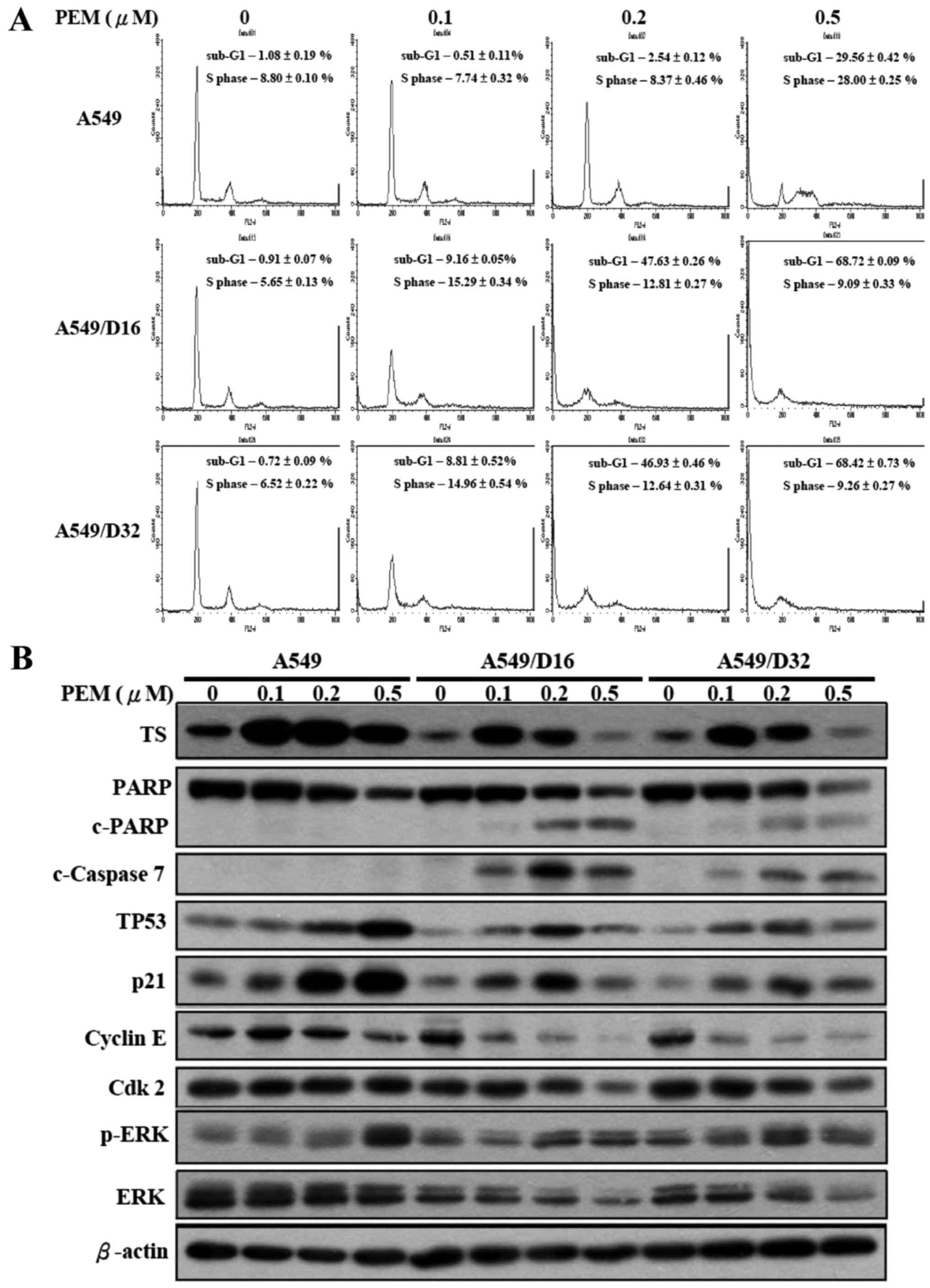
We further investigated the cell cycle distribution
of A549 and A549/D16 cells under low dosages of PEM treatment by
flow cytometry. No significant effects were observed with A549
cells. Conversely, the subG1 fraction of A549/D16 cells increased
with PEM concentration (0.1 µM, 9%), reaching a maximum level of
47% at 0.2 µM of PEM treatment. Similar results were also found in
A549/D32 cells (Fig. 3A). In order
to delineate the molecules that were affected by PEM treatment in
A549 and DOC-resistant sublines, we analyzed TS, TP53, p21 and
ERK/p-ERK as previously reported (16), as well as cleaved PARP (c-PARP),
caspase-7, cyclin E and Cdk2 by western blotting (Fig. 3B). Low expression levels of TS in
DOC-resistant sublines were increased at 0.1 µM of PEM treatment;
in addition, the c-PARP and c-caspase-7 levels were also increased
in correlation with apoptosis induction. Cyclin E and Cdk2 levels
gradually decreased indicating that the S phase was arrested by
PEM. TP53 and p21 were correspondingly induced, which also
demonstrated that DOC-selected A549 sublines maintain TP53 and p21
activities but can be activated by lower doses of PEM when compared
with parental cells. The levels of phosphorylated ERK (p-ERK) were
increased in DOC-resistant sublines, suggesting that ERK
phosphorylation still plays a role in PEM-induced cell death. This
result is similar to the findings of previous studies (16,18).
PEM inhibits the tumor growth of
DOC-resistant A549 cells in vivo
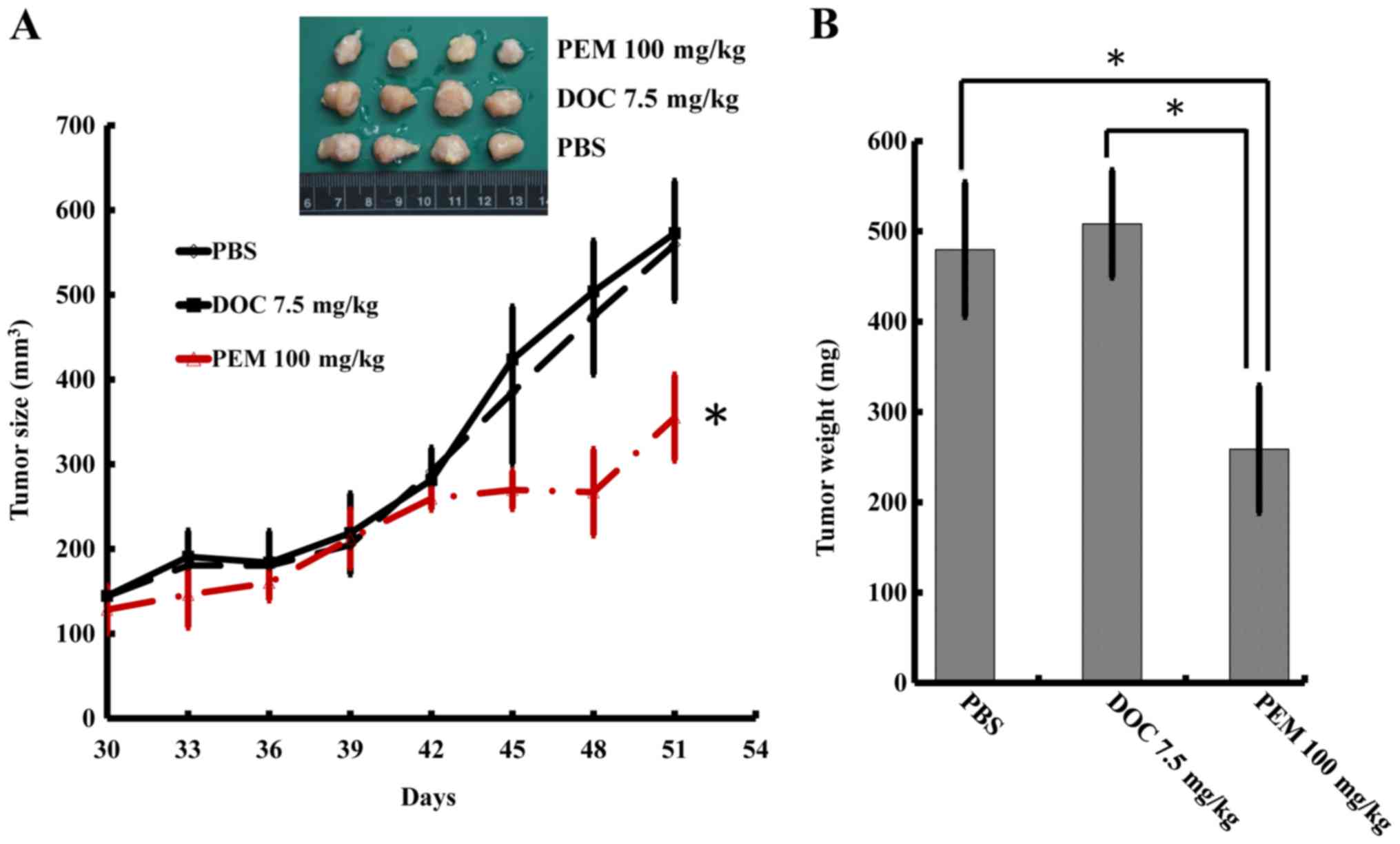
To determine whether DOC resistance may be overcome
by PEM in vivo, we inoculated the A549/D16 cells into mice
subcutaneously. The mice were fed an AIN-93M folate deficient diet
w/1% succinylsulfathiazole (catalog no. 1812813; TestDiet) to
increase PEM sensitivity (19). The
PBS group served as tumor growth controls and the DOC group served
as evidence for DOC resistance. Since DOC presents significant
toxicity to animals, the experiments were terminated after 51 days.
The tumors treated with PEM exhibited significant growth
inhibition, according to size measurements (Fig. 4A). The total weights of the tumors
were assessed and are listed in Fig.
4B, which revealed that the tumors in the group of PEM-treated
mice significantly weighed less than the tumors in the groups
treated with PBS or DOC.
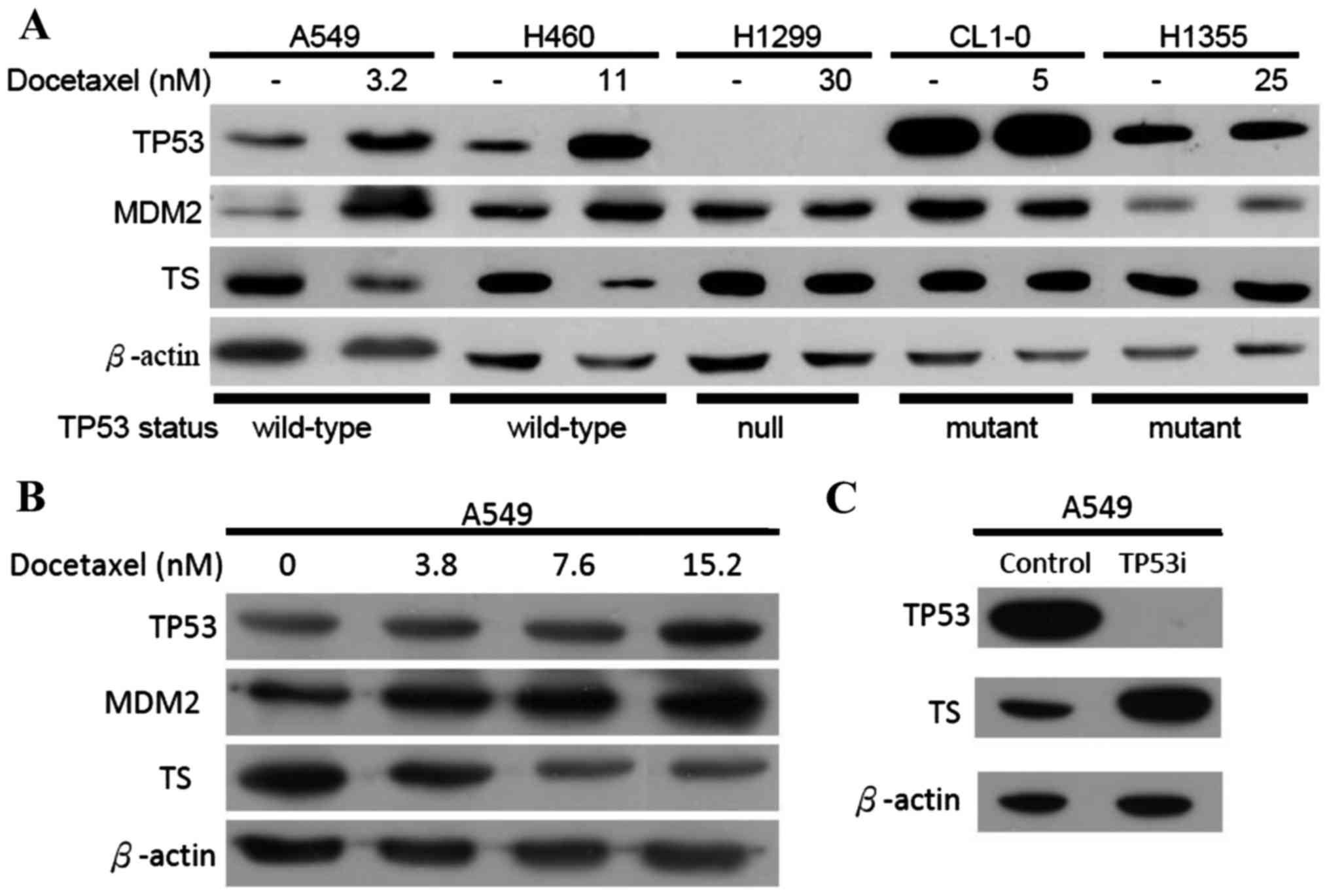
DOC induces wild-type TP53 and
downregulates TS
Previously, we reported that DOC induces TP53
transcription in A549 cells (20).
Therefore, we proposed that long-term exposure to DOC which
downregulates the folate synthesis enzymes in A549 cells may be
associated with TP53 activity. To investigate this hypothesis,
several lung cancer cell lines were selected; the H460 cells had
wild-type TP53, the H1299 cells had null TP53, and the H1355 and
CL1-0 cells had mutated TP53. All cancer cell lines were treated
with sub-lethal doses of DOC as indicated and analyzed by western
blotting. The levels of MDM2 and TP53 proteins should be increased
when wild-type TP53 is activated (21,22).
This was demonstrated in the present study when A549 and H460 cells
were exposed to DOC (Fig. 5A).
Neither MDM2 nor TP53 protein levels were altered in H1299, H1355
and CL1-0 cells treated with DOC. These data support our hypothesis
that inducing TP53 inhibits TS expression. To determine whether the
association between TP53 and TS depends on drug concentration,
various doses of DOC were added to cells and analyzed by western
blot analyses (Fig. 5B).
Downregulated TS expressions was correlated with increased DOC
concentrations. We previously established an RNAi-p53-A549 cell
line clone in which TP53 was inhibited (23). The levels of TS were significantly
increased (Fig. 5C) in these
TP53-inhibited A549 cells. According to our data, the expression of
the TS protein was regulated by wild-type TP53.
Discussion
In 2005 in the UK, the National Institute for Health
and Care Excellence (NICE) produced comprehensive guidelines on the
management of patients with lung cancer. These guidelines
recommended chemotherapy for NSCLC patients; DOC, gemcitabine
(GEM), paclitaxel (PAX) or vinorelbine (VNB) in combination with
either cisplatin (CIS) or carboplatin (CARB) were recommended as
standard first-line treatments for patients with advanced locally
or metastatic disease. Further guidance has been published which
recommends PEM in combination with CIS as a first-line treatment
for patients with non-squamous locally advanced or metastatic
disease (24). In the same study,
the effectiveness of first-line chemotherapy was evaluated using
electronic databases (MEDLINE, EMBASE and Cochrane Library). In
patients with non-squamous disease, PEM + platinum (PLAT) increased
overall survival (OS) statistically and significantly compared with
GEM + PLAT and DOC + PLAT increased OS statistically and
significantly compared with PAX + PLAT (24). DOC is an anti-microtubule agent and
PEM is an antifolate agent; therefore, they target different
cellular proteins for cancer treatment (25). In addition to the enormous efforts
being made to assist clinicians in making decisions regarding how
to treat patients of advanced NSCLC with various anticancer drugs,
we endeavored to define the influencing factors that may relieve
DOC-resistance using another anticancer drug as second-line
chemotherapy.
We established DOC-resistant A549 sublines and
defined their sensitivities with various anticancer drugs (DOC,
VCR, DXR, MTX and PEM). Notably, the DOC-resistant A549 sublines
not only avoided cross-resistance to PEM, in contrast, those
sublines were also more sensitive to PEM than parental A549 cells.
According to our previous study (15), A549/D16 and A549/D32 cells were
resistant to DOC due to overexpression of P-gp. Therefore, we
concluded that P-gp overexpression does not affect PEM sensitivity.
Our data also demonstrated that only TS, neither DHFR nor TK1, was
downregulated in DOC-resistant sublines. Overexpressing TS
decreased PEM cytotoxicity in DOC-resistant cells. Furthermore,
although a low GGH expression level indicates less ability to
counteract the multi-targeted action of PEM generated by
FPGS-mediated polyglutamylation, overexpression of GGH in
DOC-resistant A549 cells has little effect on decreasing PEM
cytotoxicity. Higher PEM sensitivity in DOC-resistant sublines can
be attenuated by overexpressing the TS gene. This indicates that
low TS expression is a major factor in PEM sensitivity. Therefore,
finding a way to decrease TS expression in NSCLC before PEM
application may be worthy of more investigation to improve the
efficacy of PEM.
It has been demonstrated that wild-type TP53
inhibits mouse TS promoter activity by >95%. In contrast, an
expression plasmid that encodes a mutant form of TP53 has little
effect on mouse TS promoter activity (26). To determine whether human TS
expression is associated with TP53 status, we examined several
human lung cancer cell lines under DOC treatment and tested to see
whether TS expression could be regulated. Our data demonstrated
that the level of TS protein was significantly decreased only in
wild-type TP53 containing human lung cells (A549 and H460) that had
been treated by DOC in a dose-dependent manner. Conversely, TS and
TP53 levels were reported in 84 breast tumor samples that were
examined using immunohistochemistry (27). Another study revealed that wild-type
TP53 status was associated with low protein expression of TS in
human colon carcinoma cell lines LoVo 92 (wild-type TP53) and WiDr
T (mutant TP53) (28). Previously,
we reported that DOC significantly induced human wild-type TP53
transcription (20,23), however induced wild-type TP53
associated with a TS decrease in human cancer cells has yet to be
demonstrated. Another interesting finding was that mutant TP53
could not be activated by DOC, which was demonstrated by the
constant levels of MDM2 expression in H1355 and CL1-0 cancer cells
containing mutant TP53 (Fig. 5A).
Therefore, the status of TP53 which altered the expression of TS
was not only found in our lung cancer model, it was also found in
breast and colon cancers, as reported by other researchers.
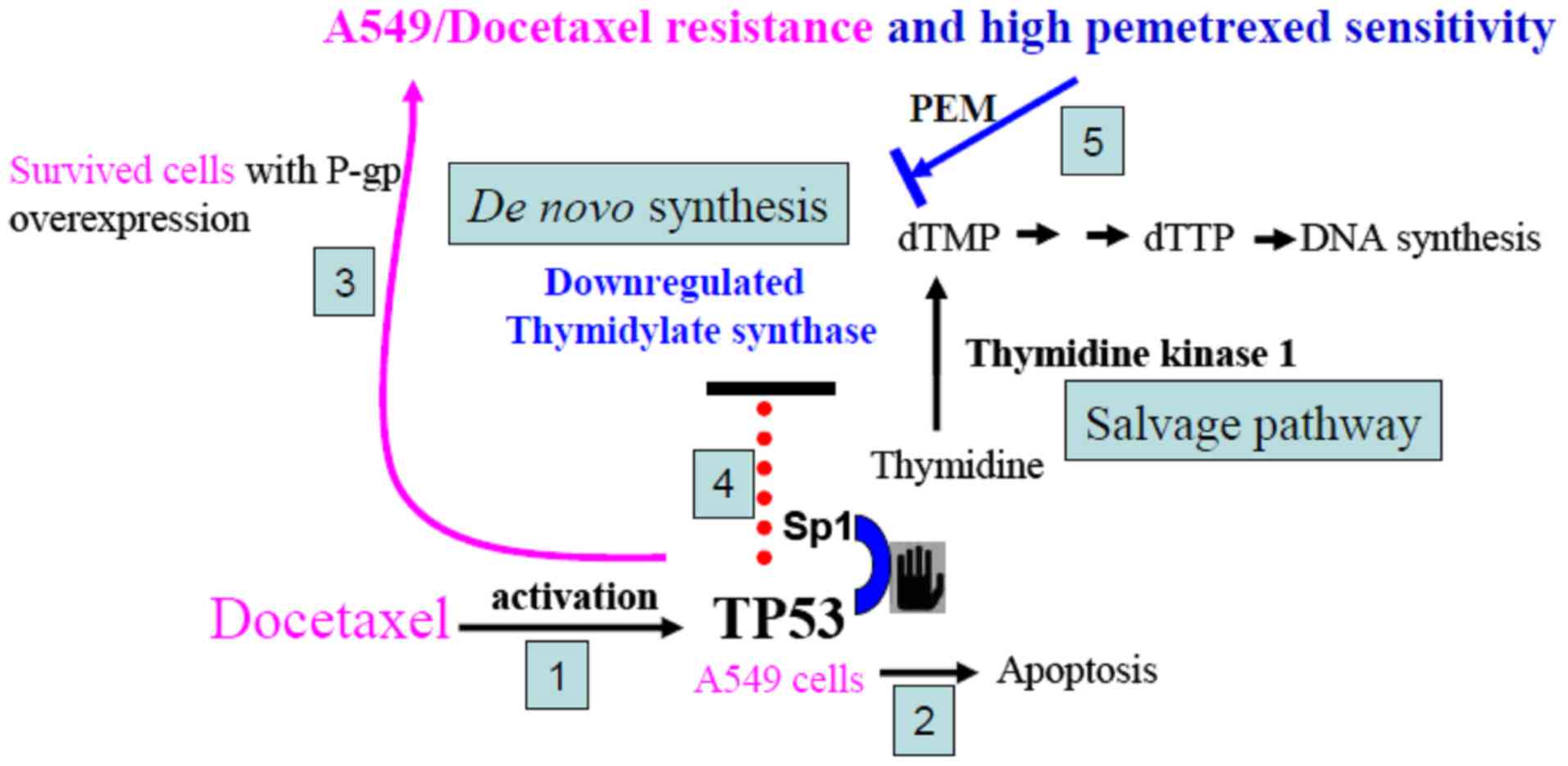
Although the mechanism involving DOC-induced TP53 suppression of TS
expression was not examined in the present study, we proposed a
possible link to TS inhibition. Previously, the upstream of the
essential promoter region of human TS was isolated. The promoter
region of the human TS gene does not contain any DNA motifs that
are typical of eukaryotic promoters, such as a TATA box, a CAAT
box, or a typical GC box. Notably, a CACCC box and an Sp1-binding
motif were identified revealing that the Sp1-binding motif acts as
the core sequence of the promoter to maintain the basic promoter
activity of genes for human TS (29). Notably, it has been demonstrated
that oxaliplatin treatment induces enrichment of TP53 at the
deoxyuridine triphosphate nucleotidohydrolase (dUTPase) promoter
with a concomitant decrease in Sp1 and a transcriptional
suppression of dUTPase (30). The
5′ region of the human dUTPase gene has been identified as a
binding site for Sp1 and it has been suggested that TP53 acts on
the region of the promoter containing the Sp1 sites, which may
prevent activation of the dUTPase gene by Sp1. Therefore, we
provide a plausible mechanism to explain why TS expression was
inhibited in DOC-resistant A549 cells (Fig. 6). Our previously reported data
revealed that the initial response of cancer cells to DOC treatment
involved wild-type TP53 (23). DOC
treatment activated TP53 and induced apoptotic death for most
cells. The surviving cells developed high P-gp expression. In these
DOC-resistant cells, activated TP53 occupied the Sp1 region and
downregulated TS expression. The salvage pathway of TK1 may provide
essential dTMP for TS-suppressed cells to continuously proliferate
as these cells are gradually selected by higher doses of DOC in
vitro. Therefore, by DOC selection, we demonstrated that A549
cells with downregulated TS were more sensitive to PEM than
parental A549 cells.
Our data revealed that application of DOC in
wild-type TP53 NSCLC as first-line chemotherapy may be a better
choice than VNB or VCR, which have similar microtubule
destabilization mechanisms. Indeed, mutations of the TP53 gene
occur in ~50% of NSCLC (31,32).
Determining TP53 status in NSCLC may be of great value in the
choice of first-line chemotherapy and management of resistance.
Recent advances in chemotherapy are welcome, but
their effects remain small for patients with NSCLC. In the present
study, we revealed that the chemotherapeutic drug DOC induced
wild-type, but not mutant TP53 and inhibited TS expression. The
de novo nucleic acid synthesis pathway was downregulated in
DOC-resistant lung cancer cells, resulting in high PEM
sensitivity.
We wondered what the optimal chemotherapy treatment
was for recurrent NSCLC patients who have previously received
taxane-based chemotherapy. Ideally, we proposed that for lung
cancer with wild-type TP53, DOC should be used as the first-line
anticancer drug. When DOC-resistance develops, PEM could be used as
the second-line treatment when the TS expression level is examined
accordingly. Our findings revealed that the sequence of
chemotherapy treatment can be optimized with DOC followed by PEM in
wild-type TP53 NSCLC. This sequence is well suited for extended
treatment intervals due to such properties as absence of cumulative
toxicity, non-cross-resistance, positive benefits on the quality of
life, and convenient scheduling. Undoubtedly, PEM may continue to
be used extensively in patients with NSCLC.
Acknowledgements
The present study was supported by the Ditmanson
Medical Foundation Chia-Yi Christian Hospital (grant no. R105-023)
and a grant to M.-F.W. (no. CSH-2014-C-007).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roengvoraphoj M, Tsongalis GJ, Dragnev KH
and Rigas JR: Epidermal growth factor receptor tyrosine kinase
inhibitors as initial therapy for non-small cell lung cancer: Focus
on epidermal growth factor receptor mutation testing and
mutation-positive patients. Cancer Treat Rev. 39:839–850. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alamgeer M, Ganju V and Watkins DN: Novel
therapeutic targets in non-small cell lung cancer. Curr Opin
Pharmacol. 13:394–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fitzpatrick FA and Wheeler R: The
immunopharmacology of paclitaxel (Taxol), docetaxel (Taxotere), and
related agents. Int Immunopharmacol. 3:1699–1714. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Horwitz SB: Mechanism of action of taxol.
Trends Pharmacol Sci. 13:134–136. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leonard GD, Fojo T and Bates SE: The role
of ABC transporters in clinical practice. Oncologist. 8:411–424.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fojo AT and Menefee M: Microtubule
targeting agents: Basic mechanisms of multidrug resistance (MDR).
Semin Oncol. 32 Suppl 7:S3–S8. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baldwin CM and Perry CM: Pemetrexed: A
review of its use in the management of advanced non-squamous
non-small cell lung cancer. Drugs. 69:2279–2302. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shih C, Habeck LL, Mendelsohn LG, Chen VJ
and Schultz RM: Multiple folate enzyme inhibition: Mechanism of a
novel pyrrolopyrimidine-based antifolate LY231514 (MTA). Adv Enzyme
Regul. 38:135–152. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goldman ID and Zhao R: Molecular,
biochemical, and cellular pharmacology of pemetrexed. Semin Oncol.
29 Suppl 18:S3–S17. 2002. View Article : Google Scholar
|
|
11
|
Zhang D, Ochi N, Takigawa N, Tanimoto Y,
Chen Y, Ichihara E, Hotta K, Tabata M, Tanimoto M and Kiura K:
Establishment of pemetrexed-resistant non-small cell lung cancer
cell lines. Cancer Lett. 309:228–235. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen CY, Chang YL, Shih JY, Lin JW, Chen
KY, Yang CH, Yu CJ and Yang PC: Thymidylate synthase and
dihydrofolate reductase expression in non-small cell lung
carcinoma: The association with treatment efficacy of pemetrexed.
Lung Cancer. 74:132–138. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Y, Yin TJ, Zhou R, Zhou S, Fan L and
Zhang RG: Expression of thymidylate synthase predicts clinical
outcomes of pemetrexed-containing chemotherapy for non-small-cell
lung cancer: A systemic review and meta-analysis. Cancer Chemother
Pharmacol. 72:1125–1132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sigmond J, Backus HH, Wouters D, Temmink
OH, Jansen G and Peters GJ: Induction of resistance to the
multitargeted antifolate Pemetrexed (ALIMTA) in WiDr human colon
cancer cells is associated with thymidylate synthase
overexpression. Biochem Pharmacol. 66:431–438. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiu LY, Ko JL, Lee YJ, Yang TY, Tee YT
and Sheu GT: L-type calcium channel blockers reverse docetaxel and
vincristine-induced multidrug resistance independent of ABCB1
expression in human lung cancer cell lines. Toxicol Lett.
192:408–418. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chiu LY, Hsin IL, Yang TY, Sung WW, Chi
JY, Chang JT, Ko JL and Sheu GT: The ERK-ZEB1 pathway mediates
epithelial-mesenchymal transition in pemetrexed resistant lung
cancer cells with suppression by vinca alkaloids. Oncogene.
36:242–253. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shintani M, Urano M, Takakuwa Y, Kuroda M
and Kamoshida S: Immunohistochemical characterization of pyrimidine
synthetic enzymes, thymidine kinase-1 and thymidylate synthase, in
various types of cancer. Oncol Rep. 23:1345–1350. 2010.PubMed/NCBI
|
|
18
|
Yang TY, Chang GC, Chen KC, Hung HW, Hsu
KH, Sheu GT and Hsu SL: Sustained activation of ERK and
Cdk2/cyclin-A signaling pathway by pemetrexed leading to S-phase
arrest and apoptosis in human non-small cell lung cancer A549
cells. Eur J Pharmacol. 663:17–26. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
van der Wilt CL, Backus HH, Smid K, Comijn
L, Veerman G, Wouters D, Voorn DA, Priest DG, Bunni MA, Mitchell F,
et al: Modulation of both endogenous folates and thymidine enhance
the therapeutic efficacy of thymidylate synthase inhibitors. Cancer
Res. 61:3675–3681. 2001.PubMed/NCBI
|
|
20
|
Chuang JC, Sheu GT, Wang PC, Liao FT, Liu
WS, Huang CF, Tseng MH and Wu MF: Docetaxel and 5-fluorouracil
induce human p53 tumor suppressor gene transcription via a short
sequence at core promoter element. Toxicol In Vitro. 26:678–685.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barak Y, Juven T, Haffner R and Oren M:
mdm2 expression is induced by wild type p53 activity. EMBO J.
12:461–468. 1993.PubMed/NCBI
|
|
22
|
Wu X, Bayle JH, Olson D and Levine AJ: The
p53-mdm-2 autoregulatory feedback loop. Genes Dev. 7:1126–1132.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chang JT, Chang GC, Ko JL, Liao HY, Liu
HJ, Chen CC, Su JM, Lee H and Sheu GT: Induction of tubulin by
docetaxel is associated with p53 status in human non small cell
lung cancer cell lines. Int J Cancer. 118:317–325. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brown T, Pilkington G, Bagust A, Boland A,
Oyee J, Tudur-Smith C, Blundell M, Lai M, Saborido C Martin,
Greenhalgh J, et al: Clinical effectiveness and cost-effectiveness
of first-line chemotherapy for adult patients with locally advanced
or metastatic non-small cell lung cancer: A systematic review and
economic evaluation. Health Technol Assess. 17:1–278. 2013.
View Article : Google Scholar
|
|
25
|
Yang TY, Chang GC, Chen KC, Hung HW, Hsu
KH, Wu CH, Sheu GT and Hsu SL: Pemetrexed induces both intrinsic
and extrinsic apoptosis through ataxia telangiectasia
mutated/p53-dependent and -independent signaling pathways. Mol
Carcinog. 52:183–194. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee Y, Chen Y, Chang LS and Johnson LF:
Inhibition of mouse thymidylate synthase promoter activity by the
wild-type p53 tumor suppressor protein. Exp Cell Res. 234:270–276.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Calascibetta A, Martorana A, Cabibi D,
Aragona F and Sanguedolce R: Relationship between thymidylate
synthase and p53 and response to FEC versus taxane adjuvant
chemotherapy for breast carcinoma. J Chemother. 23:354–357. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Giovannetti E, Backus HH, Wouters D,
Ferreira CG, van Houten VM, Brakenhoff RH, Poupon MF, Azzarello A,
Pinedo HM and Peters GJ: Changes in the status of p53 affect drug
sensitivity to thymidylate synthase (TS) inhibitors by altering TS
levels. Br J Cancer. 96:769–775. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Horie N and Takeishi K: Identification of
functional elements in the promoter region of the human gene for
thymidylate synthase and nuclear factors that regulate the
expression of the gene. J Biol Chem. 272:18375–18381. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wilson PM, Fazzone W, LaBonte MJ, Lenz HJ
and Ladner RD: Regulation of human dUTPase gene expression and
p53-mediated transcriptional repression in response to
oxaliplatin-induced DNA damage. Nucleic Acids Res. 37:78–95. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bodner SM, Minna JD, Jensen SM, D'Amico D,
Carbone D, Mitsudomi T, Fedorko J, Buchhagen DL, Nau MM, Gazdar AF,
et al: Expression of mutant p53 proteins in lung cancer correlates
with the class of p53 gene mutation. Oncogene. 7:743–749.
1992.PubMed/NCBI
|
|
32
|
Takahashi T, Nau MM, Chiba I, Birrer MJ,
Rosenberg RK, Vinocour M, Levitt M, Pass H, Gazdar AF and Minna JD:
p53: A frequent target for genetic abnormalities in lung cancer.
Science. 246:491–494. 1989. View Article : Google Scholar : PubMed/NCBI
|