Introduction
Traditional Chinese medicine (TCM), including
various natural plant extracts, has shown tremendous potential for
the discovery of alternative drugs for the treatment of cancer
(1,2). Fewer adverse effects and cost
effectiveness endow TCM great advantages over Western
chemotherapeutics (3). Patrinia
scabiosaefolia, a perennial plant mainly distributed in East
Asia, is extensively prescribed for various diseases, such as
oxidative damage (4), inflammation
(5), tumors (6) and edema (7). Of all the above-mentioned effects,
antitumor activity is a prominent characteristic of Patrinia
scabiosaefolia. Peng et al reported the Patrinia
scabiosaefolia inhibited the growth of different cancer cell
models including colorectal cancer mouse tumor tissues, HT-29 and
U266 through G1/S cell cycle arrest, suppression of tumor
angiogenesis and the STAT3 pathway, respectively (6,8,9). A
previous study demonstrated that Patrinia scabiosaefolia
induced the apoptosis of breast carcinoma MCF-7 cells without
caspase-9 activation (10).
However, the mechanisms underlying its antitumor properties emain
elusive, and the antitumor effects of Patrinia
scabiosaefolia deserve increased attention.
Recently, research which has focused on exploiting
metabolic perturbations in the area of cancer therapy has been
extensively investigated (11). ROS
homeostasis serves as a critical factor for the survival of cancer
cells. There is evidence indicating that cancer cells are more
vulnerable to high levels of ROS (12), which further evokes a series of
metabolic dysfunction, such as apoptosis, calcium overload and
bio-macromolecule degradation (13–15).
The calcium ion, as a ubiquitous secondary messenger, induces
intracellular ROS generation. In turn, ROS can regulate the
Ca2+ signaling pathway (14). Excessive accumulation of ROS and
calcium overload can initiate the apoptotic pathway (16). The reciprocal interactions of the
above three alterations intensively promote the death processes of
cancer cells. Thus, manipulation of metabolic disruptions of cancer
cells may be a promising target for cancer treatment.
SIRT-1, as an NAD+-dependent deacetylase,
has been demonstrated to regulate extensive cellular metabolic
processes including cell stress response, apoptosis and lifespan
extension (17–19). A previous study indicated that
excessive expression of SIRT-1 in large B-cell lymphoma was closely
related to poor patient prognosis (20). Recently, SIRT-1 has been
demonstrated to function as an oncoprotein and tumor promoter in
various types of cancer cells (21,22).
Mammalian target of rapamycin (mTOR), a serine/threonine protein
kinase, has been widely accepted to regulate cell growth and
proliferation (23). Various
chronic diseases, such as cancer, ageing and diabetes, are closely
correlated with mTOR (24). There
is evidence indicating that mTOR-dependent mechanisms are
responsible for the tumorigenesis of many types of cancers
(25). Thus, we speculated that
SIRT-1 and mTOR may be involved in the EPS-induced antitumor
effects.
In the present study, we investigated the antitumor
effects of an ethanol extract of Patrinia scabiosaefolia
(EPS) on 786-O cells. MTT assay, colony formation assay and
Hoechst/propidium iodide (PI) staining were performed to detect the
inhibition of proliferation and the pro-apoptotic effects of EPS.
Intracellular ROS and Ca2+ were examined to identify the
metabolic disruptions induced by EPS. However, we conducted western
blotting to explore the expression of SIRT-1 and mTOR after EPS
stimulation. Furthermore, the above antitumor effects were found to
be augmented by the SIRT-1 inhibitor nicotinamide. Thus, we
conclude that EPS induced the death of 786-O cells via SIRT-1 and
mTOR signaling-mediated metabolic disruptions.
Materials and methods
Cell culture and reagents
Human renal cell carcinoma 786-O cells were cultured
in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS),
and human proximal tubular cells (HK-2) were cultured in Dulbeccos
modified Eagles medium (DMEM)/F12 (1:1) supplemented with 10% FBS
(all from HyClone, Logan, UT, USA). Both types of cells were
cultured under the condition of 95% humidity and 5% CO2
at 37°C. After passaging, the cells were cultured for 24 h and then
treated with different concentrations of EPS for 24 h. MTT was
provided by Solarbio (Beijing, China). Hoechst 33342 and PI,
reactive oxygen species assay kit, Fluo-3 AM probe,
N-acetylcysteine (NAC) and LDH cytotoxicity assay kit were
purchased from Beyotime Biotechnology (Shanghai, China). Antibodies
for SIRT-1, p-mTOR, mTOR and GAPDH were purchased from Cell
Signaling Technology (Danvers, MA, USA). All other reagents used
were commercially available.
Preparation of an ethanol extract of
Patrinia scabiosaefolia
Patrinia scabiosaefolia Link (also named
Patrinia hispida Bunge and Patrinia scabiosaefolia
Fisch. ex Trev (Caprifoliaceae family) was provided by Shennong
Bencao Pharmacy (Shandong, China) and authenticated by Professor
Aidong Lang, the analyst of the School of Pharmacy at Shandong
University. Dry herbs of Patrinia scabiosaefolia (the plant
name has already been verified with http://www.theplantlist.org) of 1 kg were drenched in
95% ethanol (10 l) for 3 days, and then filtered. The filtrates
were evaporated in a vacuum evaporator. The relative density was
determined as 1.05. The EPS powder was obtained by a freeze-dryer.
Subsequently, EPS was dissolved in dimethyl sulfoxide (DMSO) to
produce a stock solution with the concentration of 300 mg/ml. The
final concentrations of DMSO in all the culture medium were
<0.5%. Patrinia scabiosaefolia specimen was deposited at
the Urology Laboratory of Cardiovascular Research Center in Qilu
Hospital of Shandong University.
MTT assay
MTT assay was performed to evaluate the cell
viability of 786-O and HK-2 cells. 786-O and HK-2 cells were seeded
into 96-well plates at a density of 5×103/well, and
incubated with different concentrations of EPS for 24 h.
Subsequently, MTT (0.5 mg/ml) was added into each well and the
cells were incubated for 4 h under conditions of 37°C and 5%
CO2. Then, the supernatant was removed and the formazan
crystals were dissolved in 100 µl DMSO. The absorption value was
measured at 570 nm by an ELISA reader (Thermo Multiskan MK3; Thermo
Fisher Scientific, Waltham, MA, USA).
Colony formation assay
786-O cells were seeded in 6-well plates at the
density of 1×103/well, and were then cultured for 10–14
days after stimulations. When colonies were visible, the incubation
was terminated. Subsequently, the cells were rinsed with
phosphate-buffered saline (PBS) and stained with crystal violet
after being fixed with methanol. Colonies >50 cells were counted
under a microscope with low magnification.
Hoechst 33342 and PI double
staining
Hoechst/PI double staining was conducted to examine
the apoptotic rate of 786-O cells. In brief, 786-O cells were
cultured with 10 µg/ml Hoechst 33342 and PI at 37°C for 15 min and
then rinsed with PBS three times after stimulation. Morphological
alterations of 786-O cells were observed by fluorescence microscope
(Carl Zeiss SAS, Jena, Germany). Hoechst 33342 can penetrate the
normal cell membrane, whereas PI merely penetrates the damaged cell
membrane. Cell staining with Hoechst 33342 manifests blue
fluorescence, whereas cells stained with PI manifest red
fluorescence. The apoptotic rate and necrotic rate of 786-O cells
were identified as the percentages of apoptotic cell nuclei and
necrotic cell nuclei in five random fields.
LDH release assay
To further verify the necrosis promoting effect of
EPS, LDH release assay was conducted. 786-O cells were plated in a
96-well at the density of 5×103/well. After a 24-h
treatment with EPS, 100 µl of the supernatant was transferred to a
clear 96-well plate, and then 50 µl LDH reaction working solution
was added to each well for 30 min at room temperature.
Subsequently, the absorption value was measured at 490 nm by an
ELISA reader (Thermo Multiskan MK3). The relative LDH activity was
represented by the following equation: (ODsample -
ODblank)/(ODmax - ODblank).
ROS assay
ROS assay was performed to investigate the level of
intracellular ROS. 786-O cells were seeded into a 24-well at the
density of 3×104/well. The cells were co-incubated with
250 µl DCFH-DA probe (10 µM, 1:1,000 diluted with RPMI-1640 medium)
at 37°C for 20 min after stimulation. Then, the cells were rinsed
with medium for three times and 200 µl of the medium was added into
every well. All the images were captured by fluorescence microscopy
and the fluorescent intensity was analyzed by ImageJ software
[National Institutes of Health (NIH), Bethesda, MD, USA].
[Ca2+]i
measurement
786-O cells were plated into a 24-well plate at the
density of 3×104/well. After EPS stimulation, the cells
were washed by PBS, and then incubated with 5 µM Fluo-3 AM probe
for 30 min at 37°C. Subsequently, the cells were washed with PBS
and then observed by fluorescence microscopy. The fluorescence
intensity analysis was performed by ImageJ software.
Western blotting
786-O cells were harvested by RIPA buffer plus
phenylmethylsulfonyl fluoride (PMSF), and then total proteins were
collected after stimulation. The concentrations of proteins were
measured using the BCA method. Protein samples were subjected to
10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE). The protein bands of interest were subsequently
electrotransferred to a nitrocellulose (NC) membrane and blocked
with 5% skim milk for 1 h at room temperature. Incubations were
conducted with primary antibodies (1:1,000) against GAPDH, p-mTOR,
mTOR and SIRT-1 at 4°C overnight and subsequently secondary
antibodies (1:5000) under room temperature for 1 h. After that, the
gray scale values of the bands were detected using western
immunoblotting detection (ECL system) reagents and then
semi-quantitative analyzed by ImageJ software.
Statistical analysis
All data are presented as mean ± standard error of
the mean (SEM). Statistical analyses were conducted with one-way
analysis of variance (ANOVA) followed by Tukey's post hoc analysis
and unpaired t-test. The value of P<0.05 was considered as
statistically significant. In addition, GraphPad Prism software
(GraphPad Software, Inc., La Jolla, CA, USA) was used in all
processes of the statistical analysis.
Results
EPS significantly inhibits the
proliferation and growth of 786-O and HK-2 cells
The ethanol extract of Patrinia
scabiosaefolia (EPS) exhibited significant antiproliferation
effects on the 786-O cells. Under the stimulation of EPS, cell
viability of the cells was markedly decreased in both
concentration-dependent and time-dependent manners compared to the
control group. At the concentration of 0.6 mg/ml, 786-O cells
exposed to EPS had a cell viability of 0.49±0.12 (P<0.05),
whereas, there was only 0.14±0.01 (P<0.001) viability when the
concentration reached 1.0 mg/ml (Fig.
1B). Morphological alterations were consistent with the above
results. The cells shrunk into a bright-round shape and less number
of cells were noted with shorter processes following treatment with
EPS. These damages were enhanced with the increasing EPS
concentration (Fig. 1A). The
time-dependent manner of the antiproliferation effects was also
determined by MTT assay (Fig. 1C).
Furthermore, as shown in Fig. 1D and
E, the colony count significantly decreased from 100.0±1.5 to
20.3±0.9 when the cells were co-incubated with EPS at 0.2 mg/ml for
24 h (P<0.001). Whereas, almost no colonies could be observed
under the concentration of 0.6 mg/ml EPS (P<0.001).
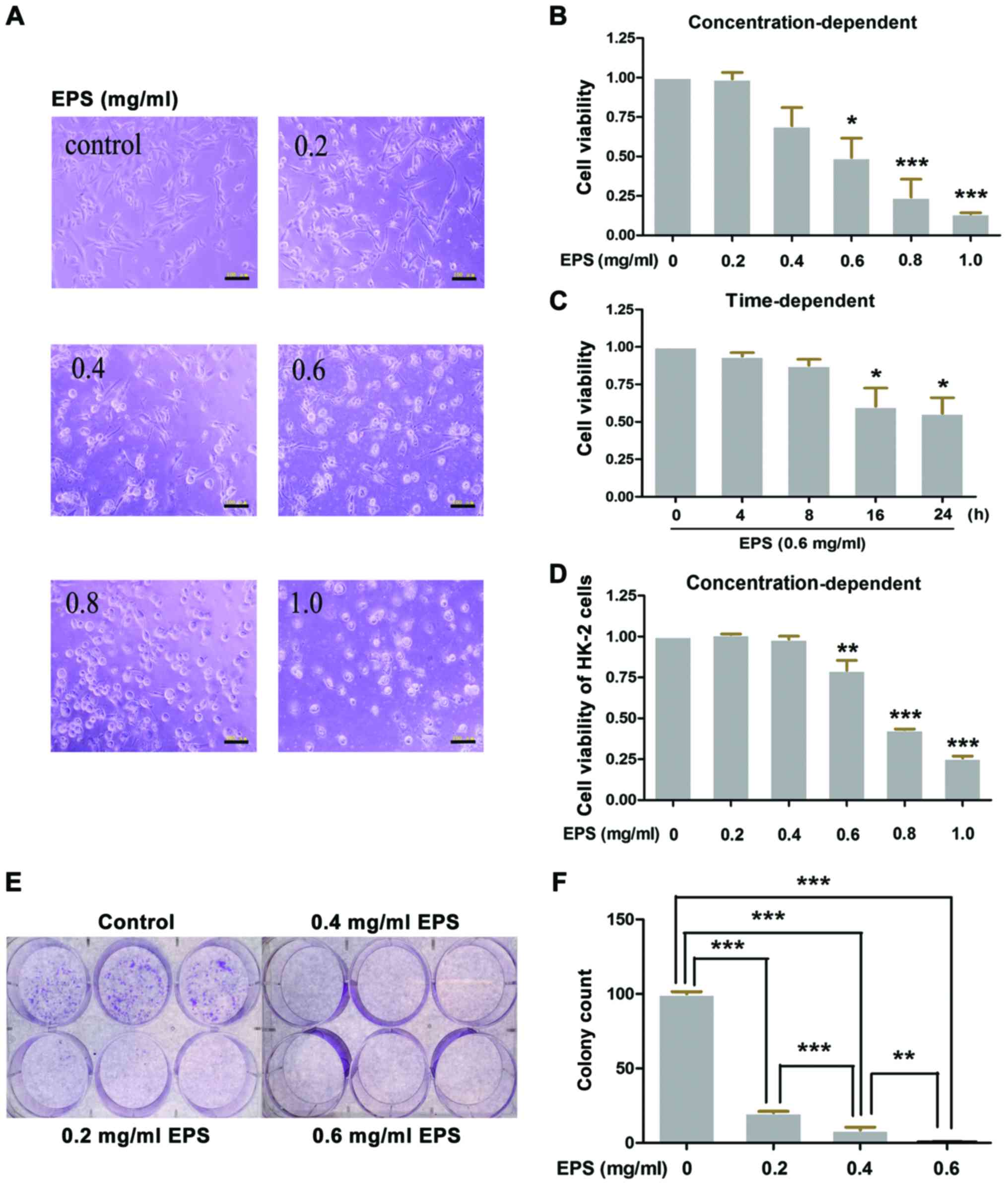 | Figure 1.Ethanol extract of Patrinia
scabiosaefolia (EPS) significantly inhibits the proliferation
and growth of 786-O and HK-2 cells. (A and B) 786-O cells were
cultured with different concentrations of EPS (0, 0.2, 0.4, 0.6,
0.8 and 1.0 mg/ml) for 24 h. (B) Then, cell viability was measured
by MTT assay and (A) images of morphological alterations were
captured by microscopy; *P<0.05, ***p<0.001, vs. 0 mg/ml. (C)
786-O cells were incubated with EPS at 0.6 mg/ml, and then cell
viability was measured at 0, 4, 8, 16 and 24 h, respectively;
*P<0.05, vs. 0 h. (D) HK-2 cells were cultured with different
concentrations of EPS (0, 0.2, 0.4, 0.6, 0.8 and 1.0 mg/ml) for 24
h and then cell viability was measured by MTT assay; **P<0.01,
***p<0.001, vs. 0 mg/ml. (E and F) Colony formation assay was
performed. After incubation with different concentrations of EPS
(0, 0.2, 0.4 and 0.6 mg/ml) for 24 h, 786-O cells were cultured for
10–14 days, and then the colony formation was determined. (E) The
difference in the colony count was analyzed; **P<0.01,
***p<0.001. All data were analyzed by one-way ANOVA with post
hoc test (n=3). Scale bars in A, 100 µm. |
Human proximal tubular cells (HK-2) were also used
to investigate the effects of EPS on normal cells. As shown in
Fig. 1D, the cell viability of the
HK-2 cells was significantly decreased in a concentration-dependent
manner. Cell viabilities of 0.793±0.062 (P<001) and 0.257±0.013
(P<0.001), respectively were noted at concentrations of 0.6 and
1.0 mg/ml.
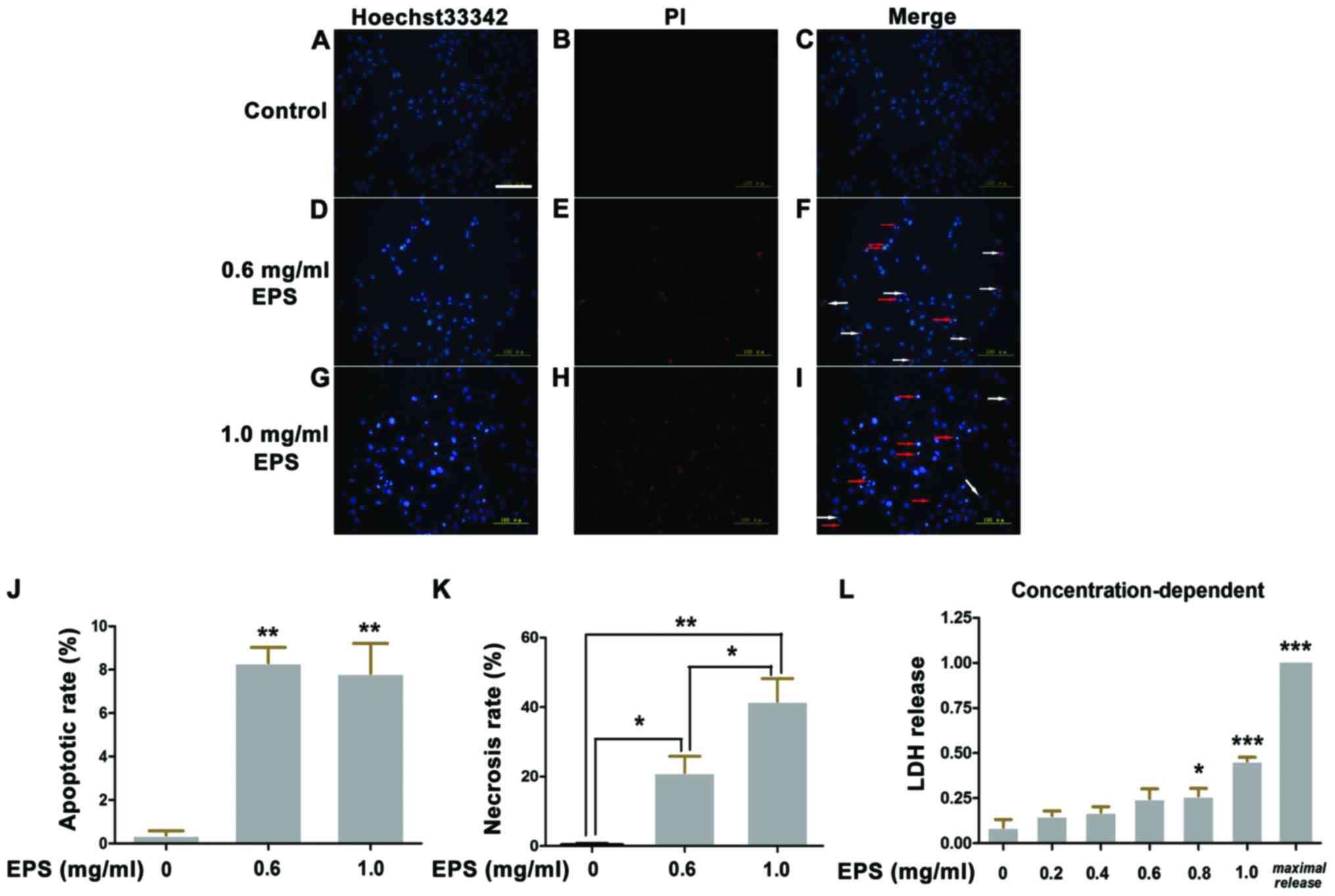
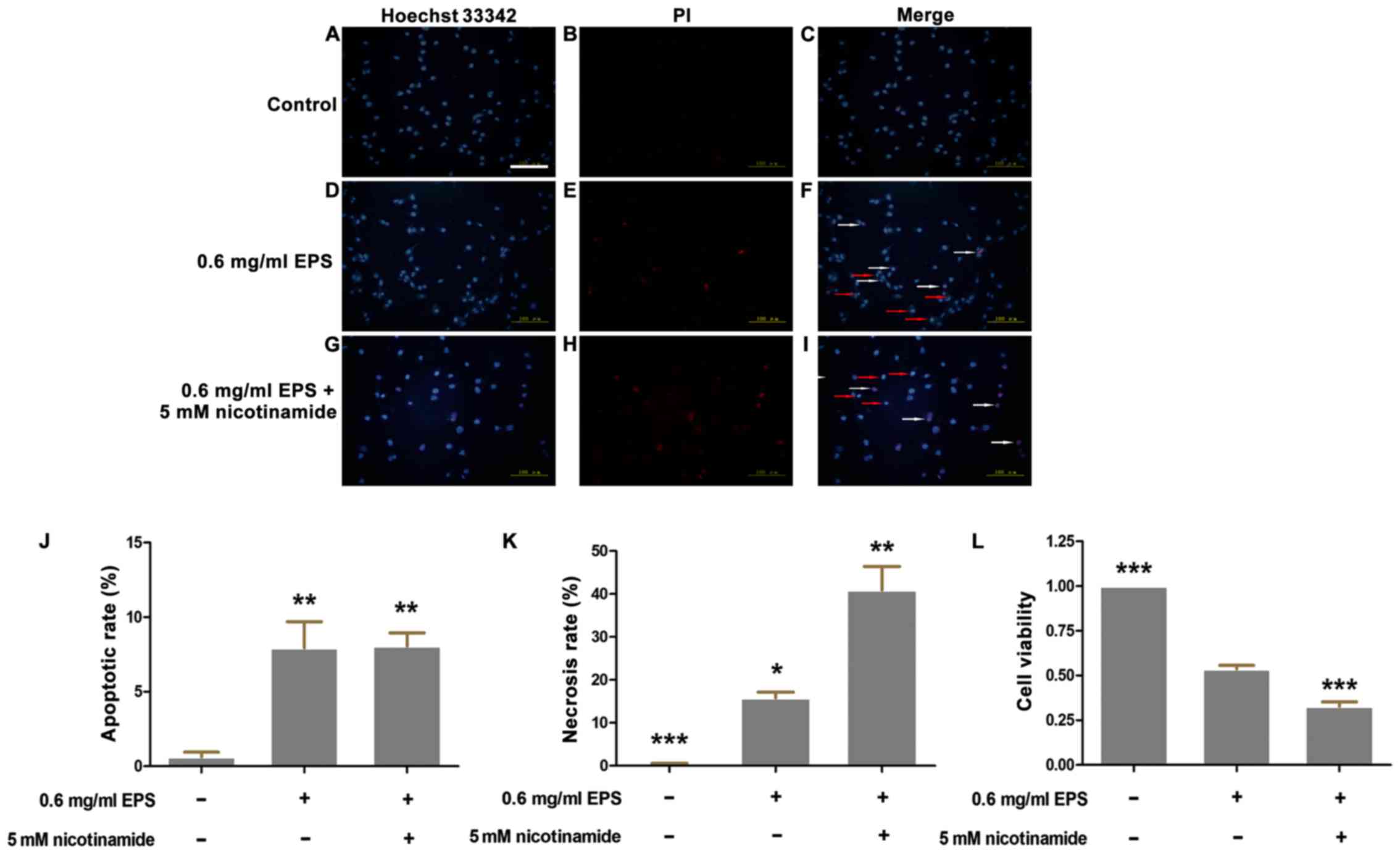
EPS significantly promotes apoptosis
and necrosis of 786-O cells
Apoptosis, which is characterized by chromatin
condensation and nuclear shrinkage, is identified by bright blue
fluorescence following staining with Hoechst 33342/PI. In contrast,
red fluorescence indicates necrotic cells due to the penetration of
PI and viable cells show dull blue fluorescence. As shown in
Fig. 2, more necrotic and apoptotic
cells were detected in the EPS stimulation group as compared to the
control group. At the concentration of 0.6 mg/ml, the apoptotic
rate was 8.3±0.7% and the necrotic rate was 21.2±4.6% compared to
0.4±0.2% for apoptosis and 0.4±0.4% for necrosis in the control
group. When the concentration of EPS increased to 1.0 mg/ml, the
necrotic rate of cells was markedly increased from 21.2±4.6 to
41.7±6.6% (P<0.05). However, there was no significant difference
regarding the apoptotic rate between treatment with EPS
concentrations of 0.6 and 1.0 mg/ml (P>0.05).
To further verify the promotion effect of EPS on
necrosis, LDH release assay was performed. There is a positive
linear relationship between the level of LDH release and the
necrotic rate. As shown in Fig. 2L,
as the EPS concentration increased, the level of LDH release was
significantly elevated in a concentration-dependent manner.
At the concentration of 1.0 mg/ml, the LDH release
rate was 0.455±0.021 (P<0.001), which was consistent with the
necrotic rate determined by Hoechst 33342/PI staining.
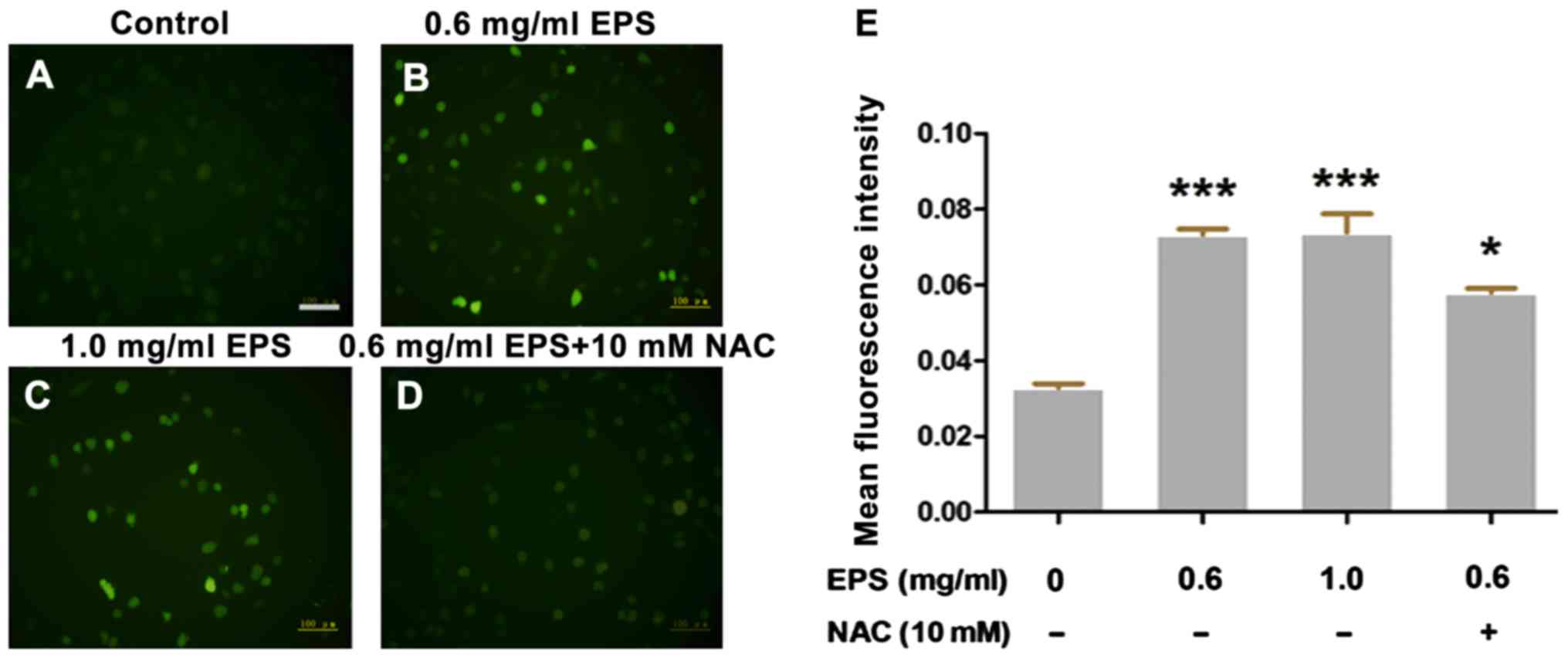
Effects of EPS on the intracellular
ROS level
To investigate the oxidative damage effects of EPS
on 786-O cells, ROS assay was performed to measure the
intracellular ROS level. H2DCFDA can be hydrolyzed by
intracellular esterases, whereas ROS oxidates non-fluorescent
H2DCFDA to convert to the highly fluorescent
2′,7′-dichlorofluorescein (DCF). Thus the level of green
fluorescence can indicate the intracellular ROS level. As shown in
Fig. 3, the intracellular ROS level
was significantly elevated compared to that noted in the control
group after EPS exposure (P<0.01). However, in the presence of
10 mM NAC, the level of intracellular ROS was significantly
decreased compared to that following treatment with 0.6 mg/ml EPS
alone (P<0.05). However, there was no obvious
concentration-dependent trend in the EPS-induced ROS increase
(P>0.05).
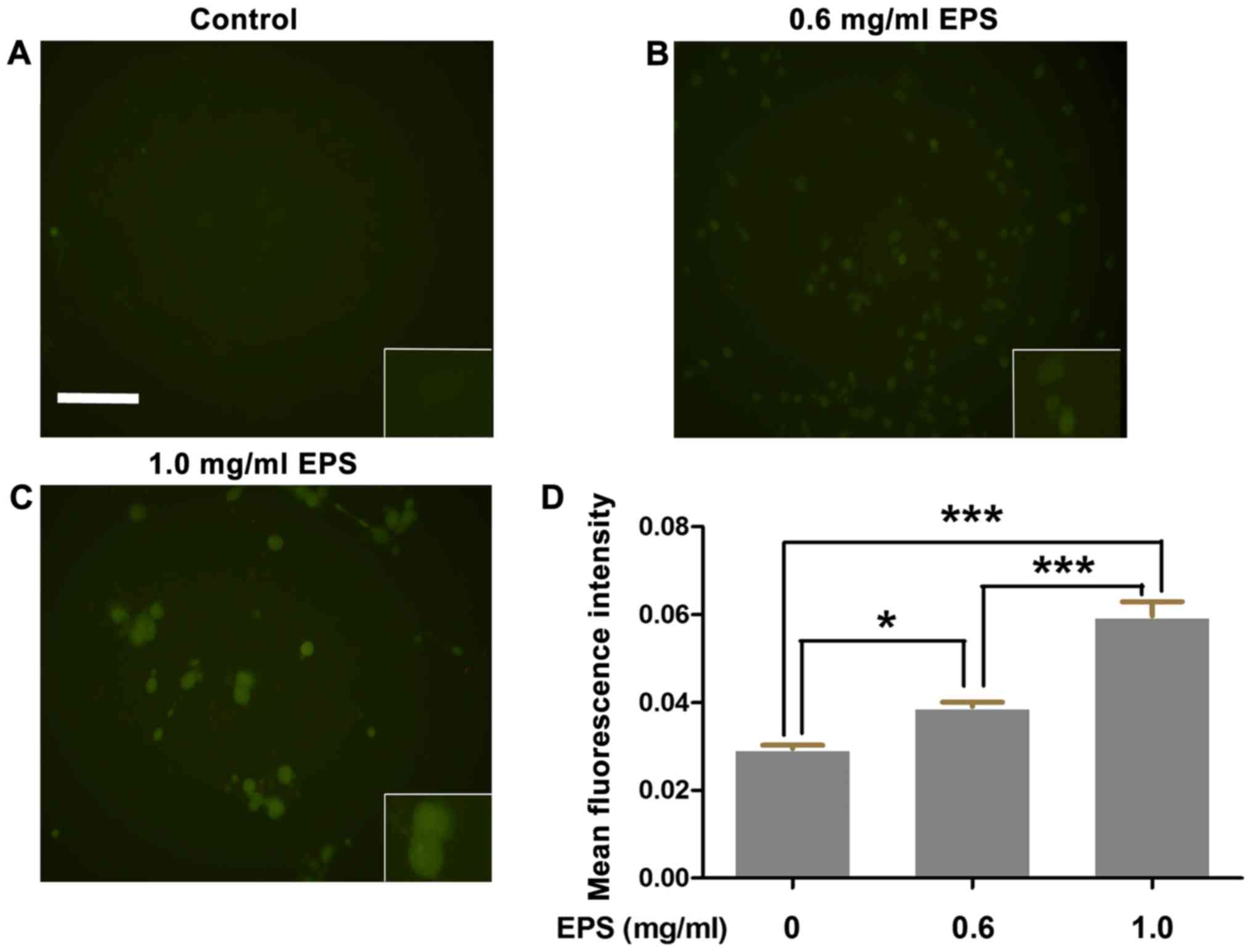
Effects of EPS on the intracellular
calcium concentration
To explore the metabolic perturbation effects of EPS
on 786-O cells, we detected the intracellular calcium concentration
by Fluo-3 AM probe. These probes specifically bind to
[Ca2+]i and emit green fluorescence. As shown
in Fig. 4, the mean fluorescence
intensity of the EPS exposure group was significantly higher than
that of the control group (P<0.01). In addition, the above
effects were significantly enhanced with the increase in EPS
concentration (P<0.001).
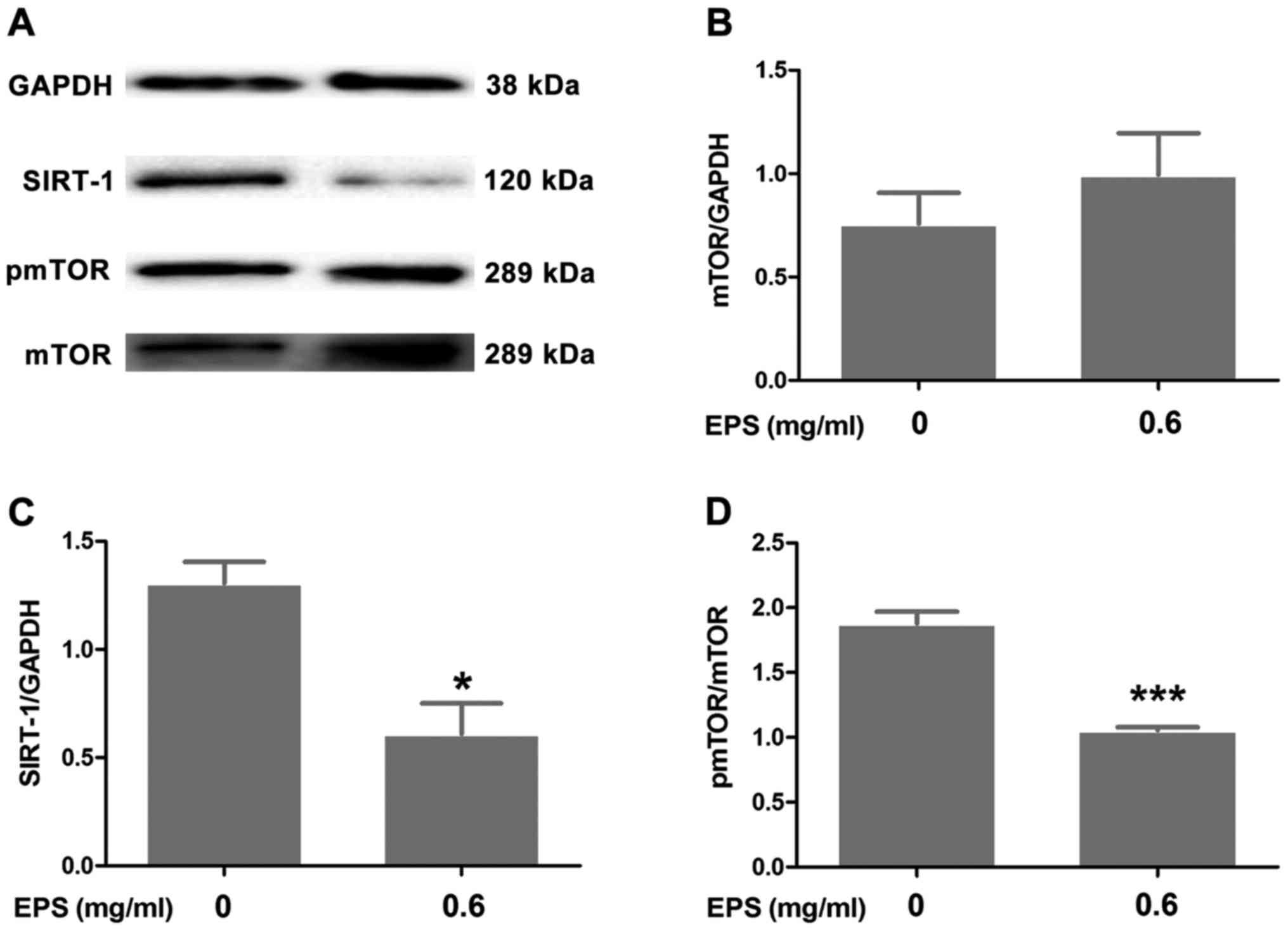
EPS downregulates the expression of
SIRT-1 and induces the dephosphorylation of mTOR
Western blotting was performed to investigate the
involvement of SIRT-1 and mTOR signaling in the antitumor effects
of EPS. As shown in Fig. 5, EPS
significantly downregulated the expression of SIRT-1 (P<0.05)
and decreased the ratio of p-mTOR/mTOR (P<0.001) in the 786-O
cells. Nevertheless, there was no statistically significant
difference regarding the ratio of mTOR/GAPDH between the EPS
exposure group and the control group (P>0.05).
SIRT-1 inhibitor nicotinamide partly
enhances the antitumor effects of EPS
To further corroborate the involvement of SIRT-1
signaling in the antitumor effects of EPS, nicotinamide, a specific
SIRT-1 inhibitor, was adopted for co-incubation with EPS. Cell
viability assay and Hoechst 33342/PI double staining were carried
out to explore the synergetic effects of nicotinamide. As shown in
Fig. 6, the cells co-cultured with
EPS plus nicotinamide had a significantly higher necrotic rate of
40.9±5.5% than that of 15.8±1.3% in the EPS group (without
nicotinamide, P<0.01). The results of the MTT assay verified the
above phenomena. The cell viability of the nicotinamide and EPS
exposure group was significantly lower (0.33±0.02)compared to the
EPS exposure group (0.54±0.02) (P<0.001). However, there was no
significant difference regarding the apoptotic rate between the
nicotinamide and EPS exposure group and the EPS exposure only group
(P>0.05).
Discussion
In the present study, we initially investigated the
antitumor effects of EPS on 786-O cells in vitro. Our
results demonstrated that EPS showed vigorous antiproliferation and
growth inhibitory effects even at a low concentration of 0.6 mg/ml,
which needed a ×500 dilution of the stock solution. Certain
fluorescence staining assays were performed to identify the manner
of cell death and the involvement of metabolic disruption. The
results indicated that an increase in ROS and calcium overload may
be responsible for the processes of metabolic perturbations.
Furthermore, the underlying molecular mechanism was explored and we
found that SIRT-1 and mTOR signaling were involved in the antitumor
effects of EPS.
Previous studies have reported that Patrinia
scabiosaefolia showed marked antiproliferation effects on
various types of tumor cells, including HT-29, MCF-7 and HeLa cells
(4,10,26).
Our results demonstrated that EPS significantly decreased the cell
viability of 786-O cells in concentration- and time-dependent
manners. As shown in Fig. 1A, the
effects of EPS on cells involved not only antiproliferation, but
also death-promotion based on morphological observation. Moreover,
EPS markedly impeded the formation of cell colonies. However, the
concentration-dependent tendency in the colony formation assay was
not in accordance with the MTT results. We speculated that the
lower initial cell seeding number may be responsible for this
inconsistency. The interactions of cells are vital to their
survival. Thus, a lower cell number made the cells more vulnerable
to EPS. As shown in Fig. 1D, EPS
showed significant antiproliferation effects on HK-2 cells.
However, at the concentration of 0.6 and 1.0 mg/ml, the cell
viabilities of HK-2 cells were 0.793±0.062 and 0.257±0.013, while
cell viabilities of 786-O were 0.493±0.124 and 0.136±0.008. This
result indicated that EPS exhibited, at least partly, specificity
in regards to its antitumor effects.
Metabolic reprogramming, which refers to a series of
metabolic alterations that stem from the molecular level, is a
distinguishing characteristic of cancer cells (11). Changes in metabolism are critical
for malignant proliferation and metastasis. However, these
alterations also make the cells more susceptible to disruptions due
to its metabolic dependency (27).
As an essential role of metabolic regulation, ROS can affect the
metabolism of cancer cells (12),
and further cause cascaded reactions of metabolic perturbations
including Ca2+ overload and apoptosis (14,16).
Our results showed that EPS significantly induced apoptosis and
necrosis of 786-O cells. However, intracellular Ca2+ and
ROS levels were markedly elevated after the exposure of EPS, while
the antioxidant NAC partly reversed the increase in ROS induced by
EPS. Therefore, metabolic disruptions may mediate the antitumor
effects of EPS, which indicate a new target for cancer
intervention.
SIRT-1, as an NAD+-dependent deacetylase,
has been demonstrated to mediate extensive cellular metabolic
processes including cell stress response, apoptosis and cell cycle
arrest (17–19). SIRT-1 deacetylates a series of
downstream molecules and induces extensive alterations, including
an increase in ROS and apoptosis (28,29).
Previous research has reported that MHY2256, an SIRT inhibitor,
showed significant anticancer effects on MCF-7 and SKOV-3 cells
through p53 acetylation (30).
Accumulated evidence has demonstrated that overexpression of SIRT-1
is found in many malignant tumors including gastric cancer,
hepatocellular carcinoma tissues, ovarian epithelial tumors and is
closely linked with poor survival outcomes (21,31,32).
Our results showed that the expression of SIRT-1 was significantly
downregulated after EPS exposure, which was consistent with the
above literature. Mammalian target of rapamycin (mTOR), formed by
mTOR complex 1 (mTORC1) and mTOR complex 2 (mTORC2), has been
implicated in metabolism, tumorigenesis and aging (24,33,34).
The mTOR signaling pathway is involved in energy conservation,
growth and division based on its protein kinase characteristics.
PI3K/Akt/mTOR and mTOR/P53/P21 pathways have been demonstrated to
be involved in antitumor effects by induction of autophagy and
inhibition of proliferation (35,36).
Wu et al reported that the Akt/GSK3β/mTOR pathway executes
the effects of neuroprotection through an antioxidative mechanism
(37). However, previous studies
and clinical practice have also demonstrated that rapamycin, a
specific inhibitor of mTOR, shows marked antitumor effects
(24). Our results indicated that
EPS obviously decreased the ratio of pmTOR/mTOR compared to the
control group, which suggests that dephosphorylation of mTOR may
induce the antitumor effects of EPS.
SIRT-1 and mTOR both mediate crucial molecular
pathways in the signaling transduction network. However, there is
no direct and explicit link between the two regulators to date
(17). Previous study has reported
that SIRT-1 negatively regulated mTOR signaling potentially
mediated by sclerosis complex 2 (TSC2) (17). Guo et al reported that SIRT-1
mediated neuron regeneration by suppressing mTOR signaling
(38). Nevertheless, there is
evidence indicating that plumbagin induced the apoptosis and
autophagy in prostate cancer cells by inhibition of both SIRT-1 and
mTOR signaling (39). Therefore, to
further substantiate the involvement of SIRT-1 signaling, we
employed nicotinamide, an inhibitor of SIRT-1, to block expression
of SIRT-1 signaling. The results demonstrated that nicotinamide
significantly enhanced the antitumor effects of EPS, which matched
our original inference.
However, the role of SIRT-1 and mTOR regarding
tumorigenesis is still controversial and there must be several
intermediate molecules between SIRT-1 and mTOR. Zhang et al
reported that the behaviors of SIRT-1 in tumorigenesis depend on
p53 mutations (18). Thus, the
alterations of P53 in cancer tissue, along with SIRT-1 and mTOR
signaling, deserve much attention in future experiments. Otherwise,
given to the involvement of intracellular ROS, the role of MAPKs
needs to be further investigated. The MAPK cascade can be activated
by ROS via various mechanisms (40). Thus, elucidation of MAPK alterations
may provide new insight into the mechanism underlying the antitumor
effects of EPS.
Based on previous literature, 10 bioactive agents
have been identified (41),
including iridoids (42), saponins
(41,43), triterpenes (44,45)
and lactones (46). Detailed
chemical fingerprint of NMR can be reviewed in the above
literature. Forty-four essential oils of Patrinia
scabiosaefolia have been identified by chemical fingerprinting
of GC (Gas Chromatography) (4).
Thus, identification of these bioactive components can provide
valuable directions for further research.
In summary, the present study investigated the
effects of EPS on 786-O cells and partly validated the involvement
of ROS and Ca2+-mediated SIRT-1 and mTOR signaling. We
highlighted the markedly antitumor effects of EPS and provide novel
insight in drug discovery for cancer treatments.
Acknowledgements
The present study was supported by grants of the
National Natural Science Foundation of China (nos. 81372335,
81400696 and 81502213), a grant of the Shandong Provincial Natural
Science Foundation (ZR2014HQ026), a grant of the Bureau of Science
and Technology of Jinan (no. 201303040), and the Science and
Technology Development Project of Shandong (no. 2011GSF11807).
Glossary
Abbreviations
Abbreviations:
|
EPS
|
ethanol extract of Patrinia
scabiosaefolia
|
|
ROS
|
reactive oxygen species
|
|
SIRT-1
|
sirtuin-1
|
|
mTOR
|
mammalian target of rapamycin
|
References
|
1
|
Kan XX, Li Q, Chen X, Wang YJ, Li YJ, Yang
Q, Xiao HB, Wang ZX, Chen Y, Weng XG, et al: A novel cell cycle
blocker extracted from Stellera chamaejasme L. inhibits the
proliferation of hepatocarcinoma cells. Oncol Rep. 35:3480–3488.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brglez Mojzer E, Knez Hrnčič M, Škerget M,
Knez Ž and Bren U: Polyphenols: Extraction methods, antioxidative
action, bioavailability and anticarcinogenic effects. Molecules.
21:212016. View Article : Google Scholar
|
|
4
|
Lin J, Cai QY, Xu W, Lin JM and Peng J:
Chemical composition, anticancer, anti-neuroinflammatory, and
antioxidant activities of the essential oil of Patrinia
scabiosaefolia. Chin J Integr Med. Sep 1–2016.(Epub ahead of
print). View Article : Google Scholar
|
|
5
|
Cho EJ, Shin JS, Noh YS, Cho YW, Hong SJ,
Park JH, Lee JY, Lee JY and Lee KT: Anti-inflammatory effects of
methanol extract of Patrinia scabiosaefolia in mice with ulcerative
colitis. J Ethnopharmacol. 136:428–435. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang M, Sun G, Shen A, Liu L, Ding J and
Peng J: Patrinia scabiosaefolia inhibits the proliferation of
colorectal cancer in vitro and in vivo via G1/S cell cycle arrest.
Oncol Rep. 33:856–860. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ju HK, Baek SH, An RB, Bae K, Son KH, Kim
HP, Kang SS, Lee SH, Son JK and Chang HW: Inhibitory effects of
nardostachin on nitric oxide, prostaglandin E2, and tumor necrosis
factor-α production in lipopolysaccharide activated macrophages.
Biol Pharm Bull. 26:1375–1378. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen L, Liu L, Ye L, Shen A, Chen Y,
Sferra TJ and Peng J: Patrinia scabiosaefolia inhibits colorectal
cancer growth through suppression of tumor angiogenesis. Oncol Rep.
30:1439–1443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng J, Chen Y, Lin J, Zhuang Q, Xu W,
Hong Z and Sferra TJ: Patrinia scabiosaefolia extract suppresses
proliferation and promotes apoptosis by inhibiting the STAT3
pathway in human multiple myeloma cells. Mol Med Rep. 4:313–318.
2011.PubMed/NCBI
|
|
10
|
Chiu LC, Ho TS, Wong EY and Ooi VE: Ethyl
acetate extract of Patrinia scabiosaefolia downregulates
anti-apoptotic Bcl-2/Bcl-XL expression, and induces apoptosis in
human breast carcinoma MCF-7 cells independent of caspase-9
activation. J Ethnopharmacol. 105:263–268. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schulze A and Harris AL: How cancer
metabolism is tuned for proliferation and vulnerable to disruption.
Nature. 491:364–373. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Trachootham D, Alexandre J and Huang P:
Targeting cancer cells by ROS-mediated mechanisms: A radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Panieri E and Santoro MM: ROS homeostasis
and metabolism: A dangerous liason in cancer cells. Cell Death Dis.
7:e22532016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan Y, Wei CL, Zhang WR, Cheng HP and Liu
J: Cross-talk between calcium and reactive oxygen species
signaling. Acta Pharmacol Sin. 27:821–826. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ganie SA, Dar TA, Bhat AH, Dar KB, Anees
S, Zargar MA and Masood A: Melatonin: A potential anti-oxidant
therapeutic agent for mitochondrial dysfunctions and related
disorders. Rejuvenation Res. 19:21–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Z, Wang H, Wang Q and Sun J: Buyang
Huanwu decoction vigorously rescues PC12 cells against
6-OHDA-induced neurotoxicity via Akt/GSK3β pathway based on serum
pharmacology methodology. Rejuvenation Res. 19:467–477. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghosh HS, McBurney M and Robbins PD: SIRT1
negatively regulates the mammalian target of rapamycin. PLoS One.
5:e91992010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang ZY, Hong D, Nam SH, Kim JM, Paik YH,
Joh JW, Kwon CH, Park JB, Choi GS, Jang KY, et al: SIRT1 regulates
oncogenesis via a mutant p53-dependent pathway in hepatocellular
carcinoma. J Hepatol. 62:121–130. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Z, Gu H, Gan L, Xu Y, Feng F, Saeed M
and Sun C: Reducing Smad3/ATF4 was essential for Sirt1 inhibiting
ER stress-induced apoptosis in mice brown adipose tissue.
Oncotarget. 8:9267–9279. 2017.PubMed/NCBI
|
|
20
|
Jang KY, Hwang SH, Kwon KS, Kim KR, Choi
HN, Lee NR, Kwak JY, Park BH, Park HS, Chung MJ, et al: SIRT1
expression is associated with poor prognosis of diffuse large
B-cell lymphoma. Am J Surg Pathol. 32:1523–1531. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen HC, Jeng YM, Yuan RH, Hsu HC and Chen
YL: SIRT1 promotes tumorigenesis and resistance to chemotherapy in
hepatocellular carcinoma and its expression predicts poor
prognosis. Ann Surg Oncol. 19:2011–2019. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee H, Kim KR, Noh SJ, Park HS, Kwon KS,
Park BH, Jung SH, Youn HJ, Lee BK, Chung MJ, et al: Expression of
DBC1 and SIRT1 is associated with poor prognosis for breast
carcinoma. Hum Pathol. 42:204–213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dibble CC and Cantley LC: Regulation of
mTORC1 by PI3K signaling. Trends Cell Biol. 25:545–555. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zoncu R, Efeyan A and Sabatini DM: mTOR:
From growth signal integration to cancer, diabetes and ageing. Nat
Rev Mol Cell Biol. 12:21–35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Menendez JA and Lupu R: Fatty acid
synthase and the lipogenic phenotype in cancer pathogenesis. Nat
Rev Cancer. 7:763–777. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu L, Shen A, Chen Y, Wei L, Lin J,
Sferra TJ, Hong Z and Peng J: Patrinia scabiosaefolia induces
mitochondrial-dependent apoptosis in a mouse model of colorectal
cancer. Oncol Rep. 30:897–903. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cairns RA, Harris IS and Mak TW:
Regulation of cancer cell metabolism. Nat Rev Cancer. 11:85–95.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu X, Yang T, Sun T and Shao K:
SIRT1mediated regulation of oxidative stress induced by Pseudomonas
aeruginosa lipopolysaccharides in human alveolar epithelial cells.
Mol Med Rep. 15:813–818. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li B, He X, Zhuang M, Niu B, Wu C, Mu H,
Tang F, Cui Y, Liu W, Zhao B, et al: Melatonin ameliorates
busulfan-induced spermatogonial stem cell oxidative apoptosis in
mouse testes. Antioxid Redox Signal. Jan. 27–2017.(Epub ahead of
print). View Article : Google Scholar :
|
|
30
|
Park EY, Woo Y, Kim SJ, Kim DH, Lee EK, De
U, Kim KS, Lee J, Jung JH, Ha KT, et al: Anticancer effects of a
new SIRT inhibitor, MHY2256, against human breast cancer MCF-7
cells via regulation of MDM2-p53 binding. Int J Biol Sci.
12:1555–1567. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang S, Huang S, Deng C, Cao Y, Yang J,
Chen G, Zhang B, Duan C, Shi J, Kong B, et al: Co-ordinated
overexpression of SIRT1 and STAT3 is associated with poor survival
outcome in gastric cancer patients. Oncotarget. 8:18848–18860.
2017.PubMed/NCBI
|
|
32
|
Jang KY, Kim KS, Hwang SH, Kwon KS, Kim
KR, Park HS, Park BH, Chung MJ, Kang MJ, Lee DG, et al: Expression
and prognostic significance of SIRT1 in ovarian epithelial tumours.
Pathology. 41:366–371. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim I, Rodriguez-Enriquez S and Lemasters
JJ: Selective degradation of mitochondria by mitophagy. Arch
Biochem Biophys. 462:245–253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Brachmann SM, Hofmann I, Schnell C,
Fritsch C, Wee S, Lane H, Wang S, Garcia-Echeverria C and Maira SM:
Specific apoptosis induction by the dual PI3K/mTor inhibitor
NVP-BEZ235 in HER2 amplified and PIK3CA mutant breast cancer cells.
Proc Natl Acad Sci USA. 106:pp. 22299–22304. 2009; View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen J, Yuan J, Zhou L, Zhu M, Shi Z, Song
J, Xu Q, Yin G, Lv Y, Luo Y, et al: Regulation of different
components from Ophiopogon japonicus on autophagy in human lung
adenocarcinoma A549 cells through PI3K/Akt/mTOR signaling pathway.
Biomed Pharmacother. 87:118–126. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin F, Lei S, Ma J, et al: [Inhibitory
effect of jianpi-jiedu prescription-contained serum on colorectal
cancer SW48 cell proliferation by mTOR-P53-P21 signalling pathway].
Zhong Nan Da Xue Xue Bao Yi Xue Ban. 41:1128–1136. 2016.(abstract
in English). PubMed/NCBI
|
|
37
|
Wu J, Zhu D, Zhang J, Li G, Liu Z and Sun
J: Lithium protects against methamphetamine-induced neurotoxicity
in PC12 cells via Akt/GSK3β/mTOR pathway. Biochem Biophys Res
Commun. 465:368–373. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guo W, Qian L, Zhang J, Zhang W, Morrison
A, Hayes P, Wilson S, Chen T and Zhao J: Sirt1 overexpression in
neurons promotes neurite outgrowth and cell survival through
inhibition of the mTOR signaling. J Neurosci Res. 89:1723–1736.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou ZW, Li XX, He ZX, Pan ST, Yang Y,
Zhang X, Chow K, Yang T, Qiu JX, Zhou Q, et al: Induction of
apoptosis and autophagy via sirtuin1- and PI3K/Akt/mTOR-mediated
pathways by plumbagin in human prostate cancer cells. Drug Des
Devel Ther. 9:1511–1554. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Son Y, Cheong YK, Kim NH, Chung HT, Kang
DG and Pae HO: Mitogen-activated protein kinases and reactive
oxygen species: How can ROS activate MAPK pathways? J Signal
Transduct 2011. 7926392011.
|
|
41
|
Gao L, Zhang L, Li N, Liu JY, Cai PL and
Yang SL: New triterpenoid saponins from Patrinia scabiosaefolia.
Carbohydr Res. 346:2881–2885. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Taguchi H and Endo T: Letter: Patrinoside,
a new iridoid glycoside from Patrinia scabiosaefolia. Chem Pharm
Bull. 22:1935–1937. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nakanishi T, Tanaka K, Murata H, Somekawa
M and Inada A: Phytochemical studies of seeds of medicinal plants.
III. Ursolic acid and oleanolic acid glycosides from seeds of
Patrinia scabiosaefolia Fischer. Chem Pharm Bull. 41:183–186. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Choi JS and Woo WS: Triterpenoid
glycosides from the roots of Patrinia scabiosaefolia. Planta Med.
53:62–65. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Inada A, Yamada M, Murata H, Kobayashi M,
Toya H, Kato Y and Nakanishi T: Phytochemical studies of seeds of
medicinal plants. I. Two sulfated triterpenoid glycosides,
sulfapatrinosides I and II, from seeds of Patrinia scabiosaefolia
FISCHER. Chem Pharm Bull. 36:4269–4274. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang MY, Choi YH, Yeo H and Kim J: A new
triterpene lactone from the roots of Patrinia scabiosaefolia. Arch
Pharm Res. 24:416–417. 2001. View Article : Google Scholar : PubMed/NCBI
|