Introduction
Eighty percent of malignant tumors that develop in
the central nervous system (CNS) are malignant gliomas, which
display histological similarities to glial cells, including
astrocytes and oligodendrocytes (1). Glioblastoma multiforme (GBM) is the
most common primary malignant adult brain tumor. GBM patients have
a median survival of 13–16 months even following therapy (2). The current standard of care consists
of maximal safe surgical resection with a combination radiotherapy
and adjuvant temozolomide chemotherapy. However, the patient 5-year
survival rate is <10% (3).
Malignant gliomas are aggressive brain tumors with limited
therapeutic options, possibly because of the highly tumorigenic
subpopulations of glioma stem cells (GSCs) (4). GSCs have been proven to be resistant
to various chemotherapeutic agents, such as temozolomide, the
first-line treatment for glioblastoma, allowing these cells to
survive therapy, leading to radioresistance and tumor relapse
(5,6). As the current treatment modalities of
GBM are not specifically designed to kill GSCs and the
effectiveness is limited, new agents targeting GSCs for potential
use in GBM clinical therapy have attracted attention.
Isoliquiritigenin (ISL), extracted from
Glycyrrhiza spp., is a promising candidate as a natural,
alternative method of cancer therapy (7). It is a flavonoid compound with
efficient antitumor activity (8).
In particular, many studies both in vitro and in
vivo, have demonstrated the antitumor efficacy of ISL against
various types of cancer (9–12). It has been suggested that ISL can
inhibit proliferation and induce apoptotic and necrotic cell death
of glioma cells (13). In the
present study, the antitumor activity of ISL on human GSCs was
examined.
Notch signaling has been reported to be involved in
fetal development and tissue homeostasis by directing many cellular
functions, including cell growth and differentiation, cell fate
determination and regulation of stem cell maintenance (14). Notch signaling pathway is an
attractive target for cancer therapy as targeting Notch signaling
could overcome multi-drug resistance (MDR) (15). It was found that a Notch/γ-secretase
inhibitor (GSI) in combination with radiotherapy and temozolomide
attenuated proliferation, decreased glioma stem cell markers and
resulted in a marked reduction in clonogenic survival in primary
and established glioma cell lines (16). Here, we hypothesized that the
anticancer effects of ISL are related to the Notch1 signaling
pathway. The present study proposes the possibility of the
therapeutic application and an intracellular antitumor mechanism of
ISL in GSC-targeted treatment for human brain glioma.
Materials and methods
Isolation of GSCs
SHG44 human glioma cells from the American Type
Culture Collection (ATCC, Manassas, VA, USA) were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS), 100 U/ml penicillin and 100 µg/ml
streptomycin at 37°C in a humidified incubator with 5%
CO2. Tumor cells grown to 80% confluence were rinsed
with sterile phosphate-buffered saline (PBS) (0.01 M, pH 7.4) and
digested at 37°C with 0.25% trypsin for 1 min, then resuspended in
a serum-free neural stem cell medium consisting of D-MEM/F-12, 2%
B27 (Gibco BRL, Gaithersburg, MD, USA), 20 ng/ml EGF (PeproTech,
Inc., Rocky Hill, NJ, USA), 20 ng/ml bFGF (PeproTech, Inc.), and
then seeded into 6-well suspension cell culture plates (Beaver,
China). After 7 days of culture at 37°C in 5% humidified
CO2 atmosphere, the medium was removed by centrifugation
and GSC-forming spheres were digested with 0.25% trypsin into a
single-cell suspension. GSCs were subcultured and enriched three
times before being used for assays.
Identification of surface phenotypes
of GSCs
GSCs grown on polylysine-coated coverslips were
fixed using 4% paraformaldehyde for 15 min and permeabilized with
0.5% Triton X-100 (PBST) for 10 min. The coverslips were blocked
with 5% bovine serum albumin (BSA) in PBS (0.01 M, pH 7.4) for 30
min at room temperature, followed by incubation with primary
antibodies targeted against rabbit anti-CD133 (#PAB12663; 1:200;
Abnova), mouse anti-nestin (#ab18102; 1:200; Abcam) and rabbit
anti-Bcl-2 (#ab32124; 1:100; Abcam) overnight at 4°C. After three
washes with PBS, the coverslips were incubated with PE-labeled
rabbit anti-mouse IgG (#ab7000; 1:100; Abcam) and FITC-labeled goat
anti-rabbit IgG (#ab6717; 1:1,000; Abcam) for 120 min at 37°C.
Nuclear DNA was labeled in blue with DAPI. The staining results
were imaged on a Nikon C-1 laser-scanning confocal microscope
(Nikon, Tokyo, Japan).
Treatments
After the cells were grown to sub-confluence, they
were supplemented with various concentrations (10, 20, 40, 80 and
160 µM, respectively) of ISL (purity ≥98%; Shanghai Yuanye
Bio-Technology Corp., Shanghai, China), 2.5 µM DAPT (a
Notch/γ-secretase inhibitor) (purity >98.5%; Gene Operation,
USA) or 10% FBS (as positive control) for different times.
Treatment effects on proliferation and differentiation were
examined by further assays described below.
Cell proliferation assay
Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto,
Japan) assay was used to measure cell proliferation of the GSCs.
Briefly, 3×103 cells per well were seeded into 96-well
suspension cell culture plates (Beaver) in triplicate. At the
appropriate time (12, 24, 48 and 72 h), 10 µl CCK-8 solution was
added to each well, and the plates were incubated for 4 h at 37°C
under a moist atmosphere with 5% CO2. The absorbance at
450 nm was measured using the Gen5 microplate reader (BioTek,
Winooski, VT, USA).
The formation of GSC spheres was observed and imaged
after 9 days of treatment (ISL was supplemented every other day)
using an Olympus IX71 Inverted Microscope (Olympus, Tokyo, Japan).
The diameters and numbers of GSC-forming spheres were calculated
from 5 random fields of view by Image-Pro Plus 6.0 software.
Immunofluorescence staining
After 48 h of treatment, GSCs grown on
polylysine-coated coverslips were fixed in 4% paraformaldehyde for
15 min and permeabilized with 0.5% Triton X-100 (PBST) for 10 min.
The coverslips were blocked with 5% BSA in PBS (0.01 M, pH 7.4) for
30 min at room temperature, followed by incubation with primary
antibodies targeted against mouse anti-GFAP (#ab4648; 1:50; Abcam),
mouse anti-β III Tubulin (#ab78078; 1:1,000; Abcam) and rabbit
anti-Hes1 (#ab119776; 1:100; Abcam) overnight at 4°C. After three
washes with PBS, the coverslips were incubated with PE-labeled
rabbit anti-mouse IgG (#ab7000; 1:100; Abcam) and FITC-labeled goat
anti-rabbit IgG (#ab6717; 1:1000; Abcam) for 120 min at 37°C.
Nuclear DNA was labeled in blue with DAPI. The fluorescent sections
were observed and photographed with a Nikon C-1 laser-scanning
confocal microscope (Nikon). The number of GFAP+ cells
and β III tubulin+ cells were determined in 5 random
fields of view by Image-Pro Plus 6.0.
Quantitative real-time PCR (qPCR)
The cultured cells were harvested and lysed at 48 h
after ISL and DAPT treatment. Total cellular RNA was extracted
using TRIzol reagent (Takara Biotechnology Co., Ltd., Dalian,
China). To remove any DNA contamination, RNA samples were treated
with Recombinant DNase I (Takara Biotechnology). Single-stranded
cDNA was synthesized from total RNA with the PrimeScript™ RT Master
Mix (Takara Biotechnology). qPCR was performed for HES1 and
internal control GAPDH using the SYBR Premix Ex Taq™ II (Takara,
Shiga, Japan) on a 7300 real-time PCR system (Applied Biosystems,
Singapore, Singapore). All reactions were carried out in triplicate
and expression data (after being calibrated with GAPDH levels) were
analyzed using the 2−∆∆Cq method. Optimal
oligonucleotide primers used in the above PCR assays were designed
and synthesized by Takara Biotechnology based on published human
cDNA sequences. The primer sequences were as follows: HES1 sense
5′-GTGTCAACACGACACCGGATAAAC-3′ and antisense
5′-CAGAATGTCCGCCTTCTCCAG-3′; GAPDH sense 5′-GCACCGTCAAGGCTGAGAAC-3′
and antisense 5′-TGGTGAAGACGCCAGTGGA-3′. All reactions were run in
triplicate.
Western blot analysis
The protein expression levels of differentiation
markers (GFAP and β III tubulin) and Notch1 signaling pathway
markers (Notch1 and Hes1) were examined by western blotting.
β-actin levels were evaluated as a loading control. Briefly, cells
were lysed and homogenized in RIPA lysis buffer (Solarbio, Beijing,
China). The supernatants were obtained by centrifugation at 4°C for
10 min at 12,000 rpm and the total protein contents were quantified
using a BCA kit (Solarbio). For electrophoresis, equal amounts of
protein (15 µg/lane) were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis on
8, 10 or 12% polyacrylamide gel depending on the target protein and
transferred onto PVDF membranes (Millipore, Billerica, MA, USA)
using a wet electroblotting system (Mini Trans-Blot; Bio-Rad). For
immunoblotting, the membranes were blocked in 5% non-fat dry milk
for 2 h at room temperature and incubated overnight at 4°C with one
of the following primary antibodies: GFAP (#ab4648; 1:400; Abcam),
β III tubulin (#ab78078; 1:800; Abcam), Notch1 (#ab128076; 1:800;
Abcam), Hes1 (#ab119776; 1:800; Abcam) and β-actin (#AP0060;
1:3,000; Bioworld Technology Inc.). After three washes in
Tris-buffered saline containing 0.5% Tween-20 (TBST), the samples
were treated with HRP-conjugated goat anti-rabbit (#E030120-01;
1:10,000; EarthOx Life Science, Millbrae, CA, USA) or goat
anti-mouse secondary antibodies (#ab6728; 1:10,000; Abcam) for 1 h
at room temperature. The antibody labeling was visualized using
enhanced chemiluminescence reagent (Millipore) and exposed on X-ray
film (Kodak, China). For the immunoblot analyses, densitometry was
performed with Image-Pro Plus 6.0 software. The optical densities
(ODs) of the protein bands were calculated relative to the ODs of
the reference protein β-actin.
Statistical analysis
Data are expressed as the mean ± SD. All statistical
analyses were performed using the SPSS software program (version
21.0; IBM Corp., Armonk, NY, USA). The differences between the
experimental groups were analyzed by one-way analysis of variance
(ANOVA). P<0.05 was considered statistically significant.
Results
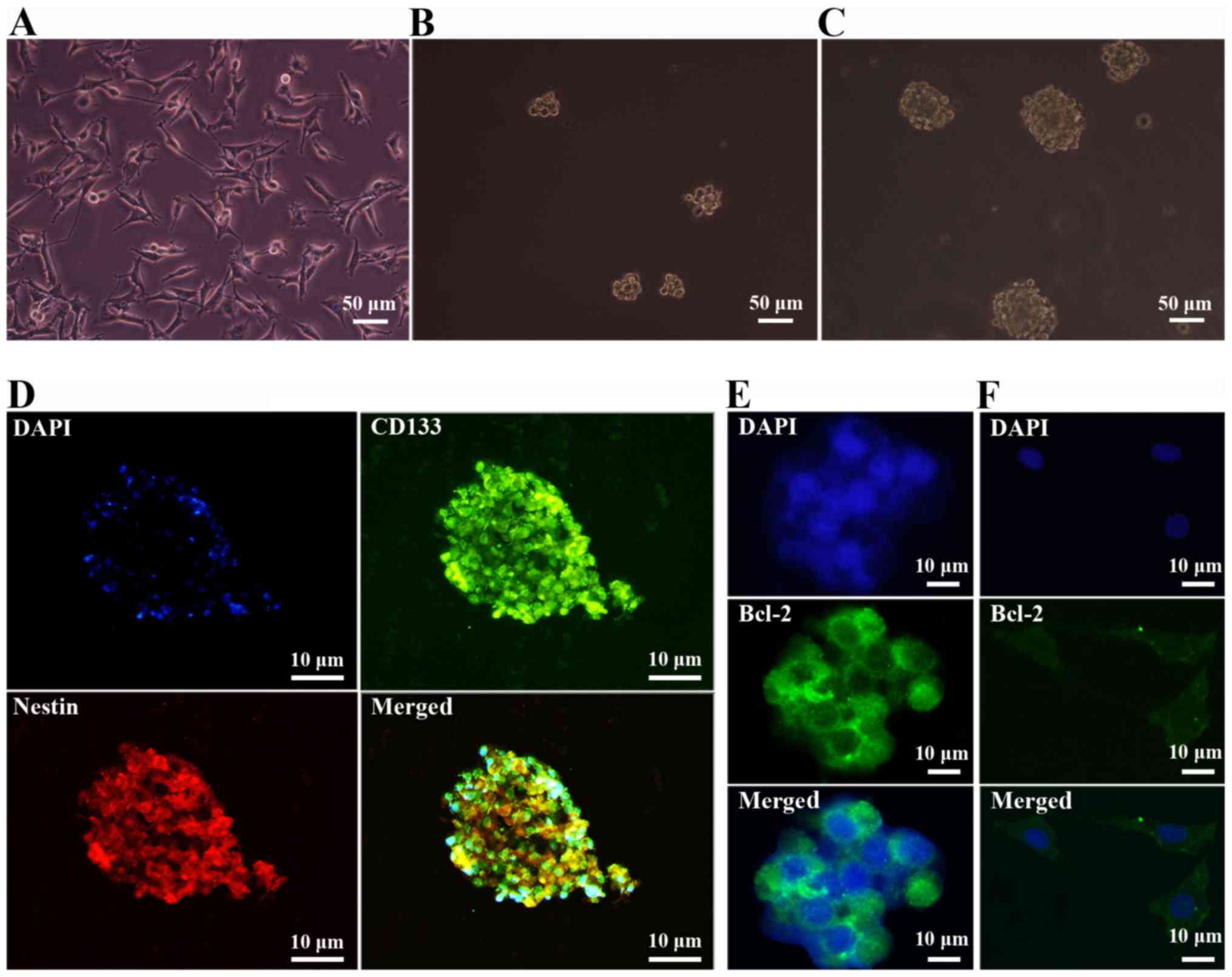
Glioma cells and GSC-forming
spheres
Within 48–72 h of serum-free culture SHG44 human
glioma cells yielded a minority fraction of cells that possessed
extensive proliferative and self-renewal potential, and generated
free-floating neurosphere-like brain tumor spheres. These spheres
gradually enlarged as the time of the culture increased (Fig. 1B and C). In comparison, glioma cells
cultured in DMEM with 10% FBS growed adherently (Fig. 1A).
Characterization of surface phenotypes
of GSCs
Expression of GSC surface markers were examined in
human GSC-forming spheres. As shown in Fig. 1D, the cells expressed characteristic
neural stem cell (NSC) markers CD133 and Nestin. SHG44 human glioma
cells and tumor neurospheres were postive for the proto-oncogene
Bcl-2 which distinguished them from normal neurons and NSCs
(Fig. 1E and F). The
immunofluorescence staining data indicated that the tumor spheres
expressed typical surface markers of GSCs and thus were used for
experiments as described below.
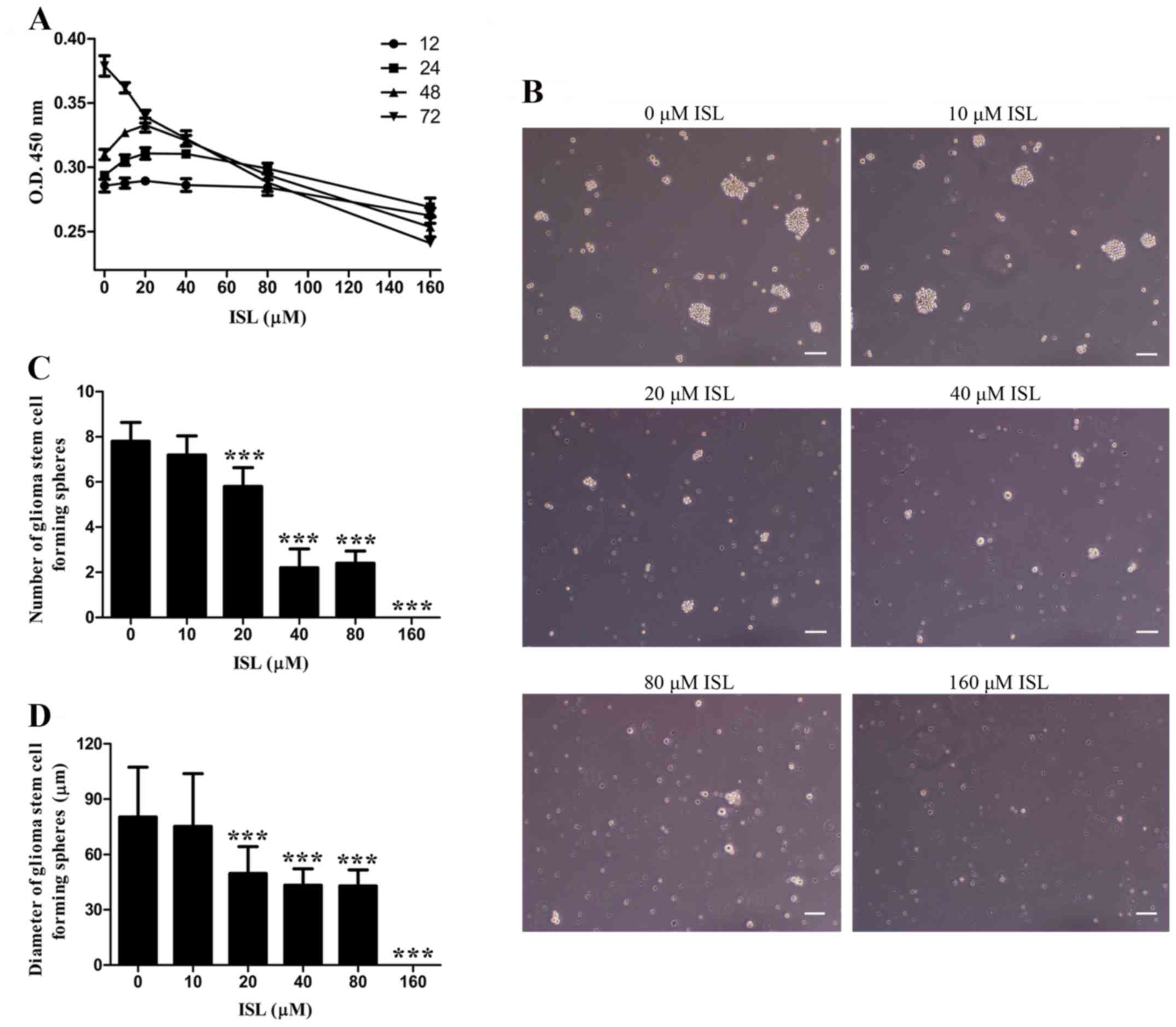
ISL inhibits cell proliferation in a
dose-dependent manner
To investigate the influence of ISL on GSC
proliferation, we firstly examined the activities of cells treated
with various concentrations of ISL for different times. As shown in
Fig. 2A, within 24 and 48 h of
treatment the proliferative activity of the cells was significantly
higher in the 10, 20 and 40 µM ISL groups than that noted in the 0
µM ISL group. There was no significant difference between the 0 and
80 µM ISL group within 24 h, but the activity of cells began to
decline after 48 h. After 72 h, the cell proliferation of the ISL
treatment groups were obviously inhibited in a dose-dependent
manner, with a half inhibitory concentration IC50 of
102.744 µmol/l.
Formation of neurospheres is a morphological feature
of GSCs. Then we examined the influence of ISL on GSC-forming
spheres on the 9th day of treatment. It was found that the 0 µM ISL
group had the largest diameter and highest number of tumor spheres
formed from the GSCs (Fig. 2B-D).
The diameters and the numbers were both decreased by ISL in a
dose-dependent manner, and 160 µM ISL completely prevented
neurophere formation, displaying a similar tendency with cell
activity at 72 h.
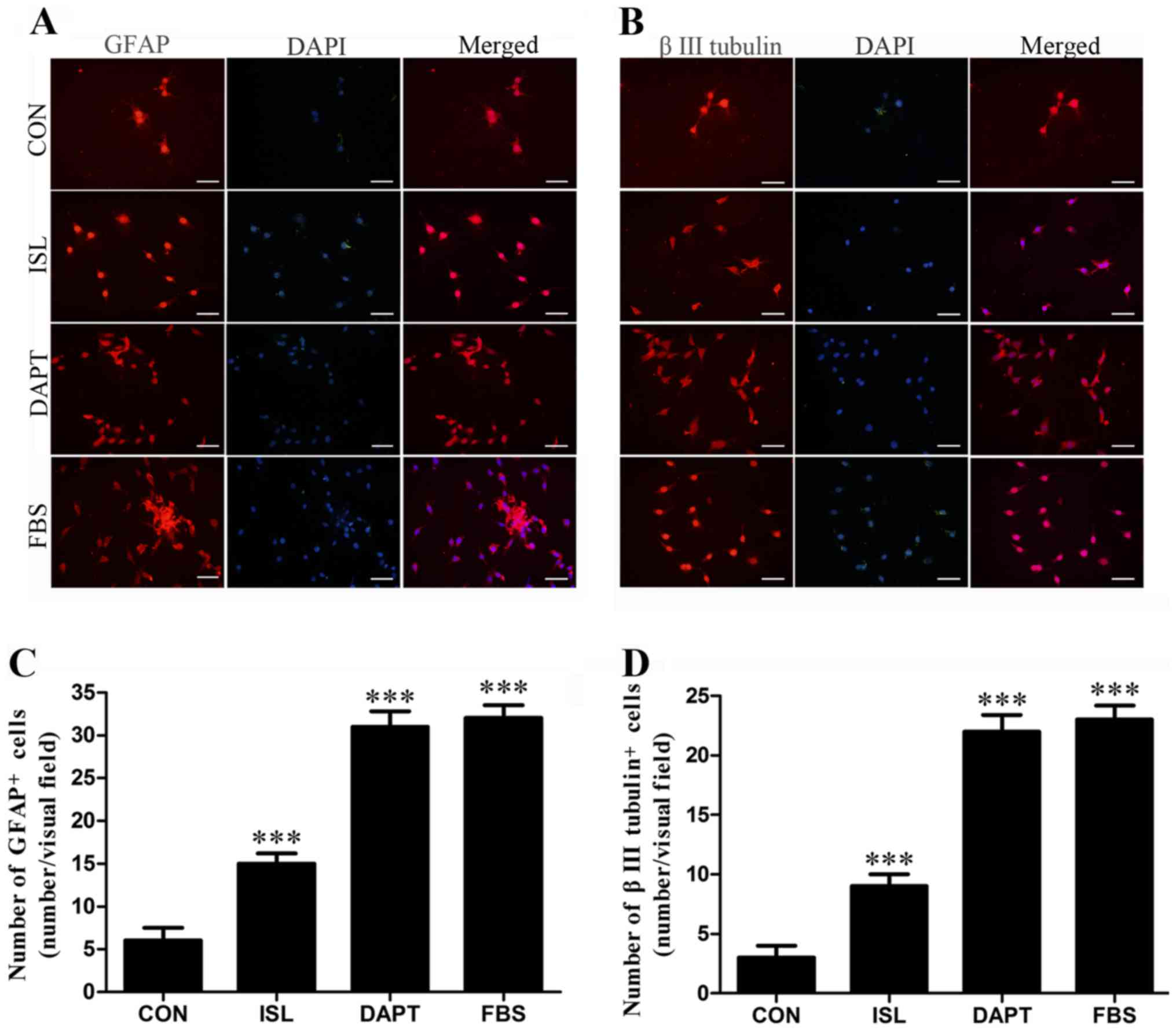
Differentiation-promoting action of
ISL on GSCs
To determine whether ISL promotes the
differentiation of GSCs, intracellular differentiation markers GFAP
and β III tubulin were examined after 48 h of ISL treatment by
immunofluorescence staining. As shown in Fig. 3, the numbers of differentiation
marker-positive cells were higher in the ISL group and DAPT group
than that in the control group after 48 h. As a comparison, FBS
effectively induced the differentiation of GSCs.
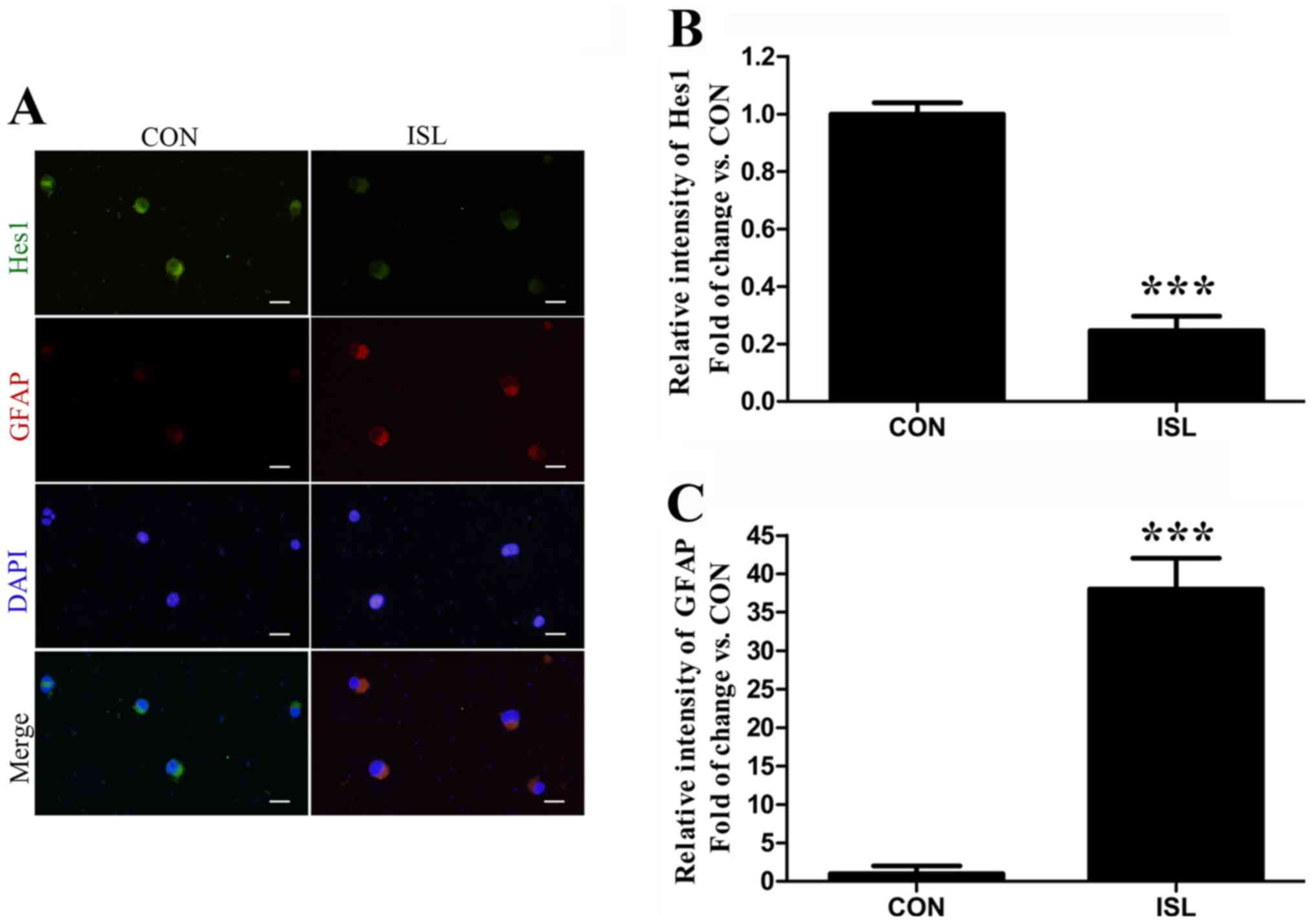
To determine whether ISL inhibits Hes1 (Notch1
target gene) in the differentiated cells, GFAP and Hes1 were
assessed together after the cells were treated for 48 h. The
relative intensity of Hes1 was decreased in the GFAP+
cells induced by ISL (Fig. 4).
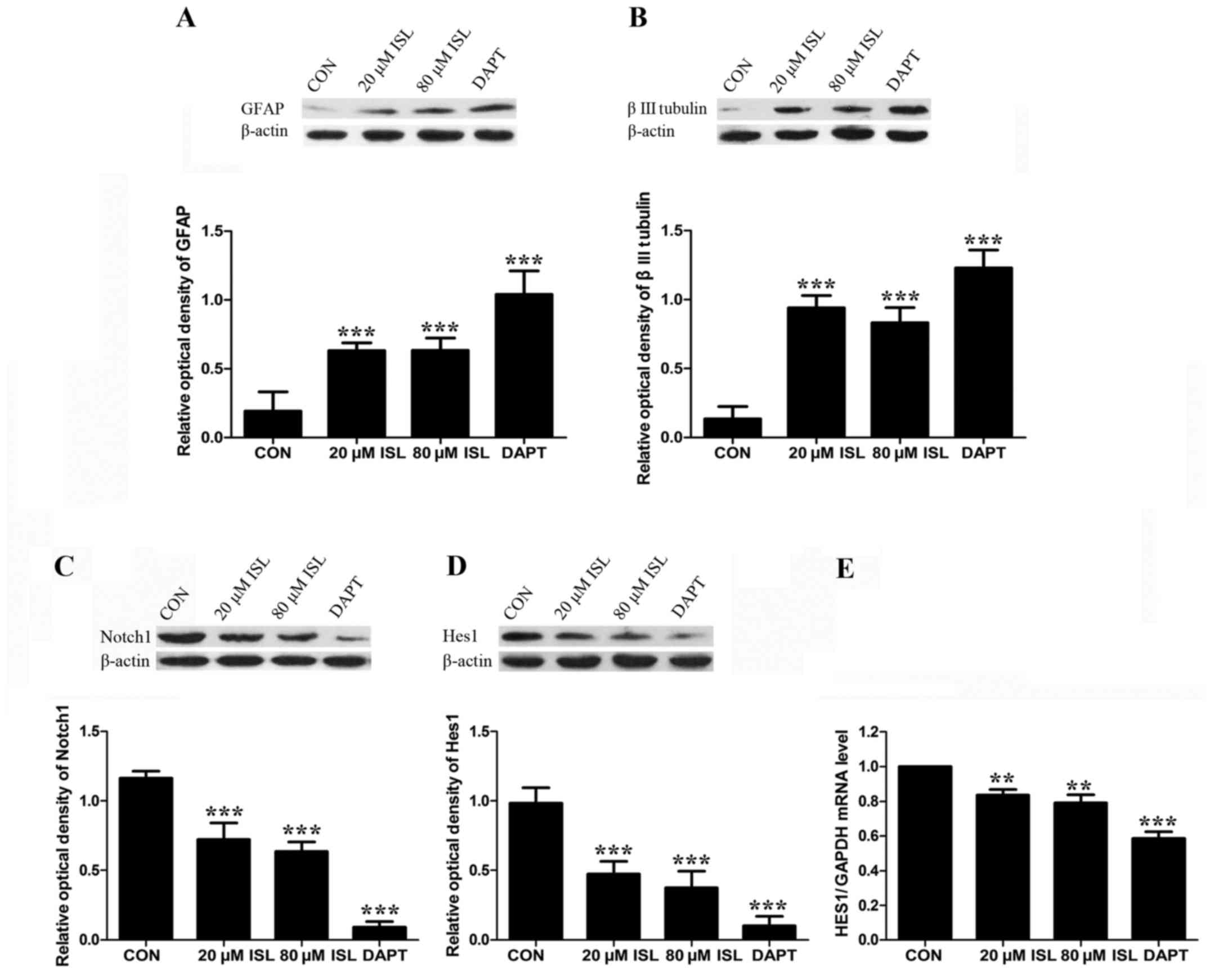
The protein expression levels of differentiation
markers were also examined after 48 h of ISL treatment. As
expected, the protein expression levels of GFAP and β III tubulin
in the DAPT group and ISL groups were higher than those of the
control, and DAPT induced higher GFAP and β III tubulin protein
expression levels compared to the ISL groups. However, there was no
obvious difference between the low dose (20 µM) and high dose (80
µM) ISL groups (Fig. 5A and B).
Ability of ISL to downregulate the
Notch signaling pathway
To examine whether ISL inhibits the proliferation
and induces the differentiation of human GSCs through Notch1
signaling, Notch1 and Hes1 were analyzed 48 h after GSCs were
respectively treated with ISL and DAPT. The predicted molecular
weight of Notch1 is 272 kDa. In the present study, the band
observed at 120 kDa could potentially be a truncated form of
Notch1, following cleavage by furin-like protease. The protein
expression levels of Notch1 in the DAPT group and ISL groups were
lower than those of the control (Fig.
5C). The mRNA and protein expression levels of Hes1 were also
significantly decreased compared to the control (Fig. 5D and E). DAPT induced lower Notch1
and Hes1 expression levels compared to the ISL groups.
Discussion
It has been reported that supplementation with ISL
may be an effective approach to inhibit the growth of tumors and
counteract the adverse effects of existing drugs for cancer
patients (17). Despite the studies
demonstrating the efficacy of ISL in tumor therapy, its anticancer
activities against GSCs remain unclear. In the present study, we
identified that ISL inhibited proliferation and induced
differentiation of human GSCs, and inhibited the activation of the
Notch1 pathway.
Serum-free culture is a simple and convenient method
for neural stem cell (NSC) enrichment and has been applied for
cancer stem cell (CSC) isolation from brain tumors and many other
types of cancers (18). Using
serum-free medium with supplemental factors B27, EGF and bFGF, the
present study isolated tumor spheres from SHG44 human glioma cells
on suspension cell culture plates. CD133 and Nestin are important
cell surface markers present on NSCs. A high level of CD133
expression may be an independent risk factor for glioma patient
prognosis and high expression of Nestin tends to correlate with a
worse outcome for glioma patients (19). As previously described, GSCs develop
high chemoresistance and radioresistance. In this respect, a high
level of CD133 or/and Nestin expression in glioma cells may imply a
high proportion of GSCs which contribute to adverse prognosis. In
this study, CD133 and Nestin were used together to examine the stem
cell property of tumor spheres and the expression of Bcl-2 provided
further evidence that these neurospheres were GCSs derived from the
SHG44 cell line.
It has been reported that the inhibition rate of U87
human glioma cells can be ~80% after a 72-h treatment with 40–60 µM
ISL (13,20). Our data indicated that ISL can
dose-dependently inhibit the proliferation of human SHG44 GSCs 72 h
after treatment. GSC proliferation was obviously inhibited after a
72-h incubation with 10 µM ISL. However, the IC50 of
102.744 µmol/l seemed much higher than that of other studies
mentioned above. This might be because GSCs are proven to be more
resistant to various chemotherapeutic agents. Some recent studies
have suggested the use of ISL as a natural candidate to reduce
CSC-like populations and colony formation, accompanied by enhanced
CSC chemosensitivity (21,22). Here, we investigated the inhibitory
effect of ISL on the formation of glioma neurospheres for the first
time. In terms of long-term efficacy, the diameters and the numbers
of GSC-forming spheres on the 9th day of 20 µM ISL treatment were
significantly decreased. Thus, we believe that 20 µM ISL can be
sufficient to inhibit GSCs. Unfortunately, there are few
toxicological evaluations reported for ISL treatment. Our findings
will provide the theoretical and experimental bases for ISL
clinical use in the treatment of glioma. Further studies are still
required to investigate the clinical side-effects in
vivo.
Here, we also showed the stimulating effect of ISL
on GSC differentiation into downstream neural lineages. It has been
reported that ISL facilitated the differentiation of human
promyelocytic leukemia and mouse melanoma cells (23,24).
The present study first showed that ISL induced GSCs to
differentiate into astrocytes (GFAP-positive) (25) and neurons (β III tubulin-positive)
(26). As a result, after
differentiation induced by ISL, the majority of GSCs were able to
be killed by ISL, leading to decreased cell numbers. Neurons are
highly differentiated cells and rich in tubulin isotypes (27). Another study found that β-tubulin is
required for cell viability and neurite outgrowth, and the
suppression of β І isotype resulted in significant loss of cell
viability during cell differentiation (28). In the present study, it is perhaps
not coincidental that ISL increased the viability of GSCs at 24 and
48 h when a proliferation inhibitory effect was not dominant.
Therefore, GSC proliferation was activated following 20 µM ISL at
48 h when GSC viability was mainly increased in the differentiating
cells. On the contrary, GSC proliferation was inhibited by 80 µM
ISL at 48 h as a high dose of ISL exhibited a strong growth
inhibitory effect which abrogated the increase in viability of the
differentiated cells. GSCs and NSCs express similar signatures,
such as infinite self-renewal ability, neurosphere formation, and
multipotential differentiation capacity (6,29).
GSCs in the control group treated without ISL still generated a
small population of differentiated cells, but were much less than
those in the ISL groups. Further studies are required to
investigate the precise mechanism of how ISL induces GSCs to
differentiate into each phenotype.
Recently, an accumulating body of evidence has
emerged to suggest that the Notch1 signaling pathway plays an
important role in cell fate decisions, and functions as a key
regulator of cell growth, differentiation and proliferation
(30,31). Fan et al (32) observed that inhibition of the Notch1
signaling pathway by a γ-secretase inhibitor resulted in the
apoptosis and differentiation of medulloblastoma cell lines, and
reduced tumor progression. In the present study, the Notch1 pathway
was both inhibited by DAPT and ISL. The mRNA and protein expression
levels of Notch1 were altered in a similar trend with cell
proliferation and in a reverse trend with GFAP and β III tubulin,
indicating that Notch1 signaling was blocked directly or indirectly
by ISL and therefore changed the proliferation and differentiation
of GSCs. However, there was no obvious difference in Notch1 pathway
expression between low dose (20 µM) and high dose (80 µM) ISL
groups. This is possible as other pathways may be involved. In
terms of differentiation, cells treated with low dose and high dose
ISL had the same protein level of differentiation markers. The
possible reason may be that cell differentiation-induced action
reached a plateau. DAPT induced lower Notch1 and Hes1 expression
levels compared to the ISL groups. It is proposed that DAPT and ISL
might have different target sites of downregulation of Notch1.
Further studies are still required to investigate the crosstalk
between Notch and other signaling pathways, including the NF-κB,
Wnt and PI3K/Akt/mTOR pathways in GSCs.
ISL is a flavonoid compound with efficient antitumor
activity. However, the drug clearance has not been measured in
recent studies. Yang et al (33) firstly reported the transmembrane
transport of ISL and other cardio-cerebral vascular protection
flavonoids using the CaCo-2 and blood-brain barrier (BBB) cell
models in vitro. An increasing number of studies have shown
that flavonoids are capable of crossing the blood-brain barrier via
different BBB models such as RBE-4 cells, hCMEC/D3 cells, and
ECV304/C6 coculture (34–36). In situ (rat) models have
demonstrated that flavonoids are able to traverse the BBB in
vivo. In recent years, new strategies such as the use of
carrier-mediated transport systems and nanotechnology-based
approaches enable drug transport into the brain (37). These will hopefully reduce the
excretion of ISL by the body and lead toward the successful
treatment of glioma patients. Nevertheless, further studies are
still required to reveal the underlying mechanisms and comfirm the
benefit of ISL treatment in vivo.
In conclusion, the present study confirmed our
hypothesis that ISL can effectively inhibit the proliferation and
induce the differentiation of human GSCs. The therapeutic effect
may be related to downregulation of the Notch1 signaling pathway.
However, further studies are needed to identify the target/receptor
molecules of ISL on the cell surface, and to comfirm the
therapeutic effect in vivo. These studies will help to
screen for the optimal parameters and elucidate the mechanism of
ISL as a promising agent for the treatment of human glioma.
References
|
1
|
Chen J, McKay RM and Parada LF: Malignant
glioma: Lessons from genomics, mouse models, and stem cells. Cell.
149:36–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lau D, Magill ST and Aghi MK: Molecularly
targeted therapies for recurrent glioblastoma: Current and future
targets. Neurosurg Focus. 37:E152014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Clarke J, Penas C, Pastori C, Komotar RJ,
Bregy A, Shah AH, Wahlestedt C and Ayad NG: Epigenetic pathways and
glioblastoma treatment. Epigenetics. 8:785–795. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Codrici E, Enciu A-M, Popescu I-D, Mihai S
and Tanase C: Glioma stem cells and their microenvironments:
Providers of challenging therapeutic targets. Stem Cells Int.
2016:57284382016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen J, Li Y, Yu T-S, McKay RM, Burns DK,
Kernie SG and Parada LF: A restricted cell population propagates
glioblastoma growth after chemotherapy. Nature. 488:522–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schonberg DL, Lubelski D, Miller TE and
Rich JN: Brain tumor stem cells: Molecular characteristics and
their impact on therapy. Mol Aspects Med. 39:82–101. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng F, Du Q, Peng C, Wang N, Tang H, Xie
X, Shen J and Chen J: A Review: The pharmacology of
isoliquiritigenin. Phytother Res. 29:969–977. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Weng CJ and Yen GC: Flavonoids, a
ubiquitous dietary phenolic subclass, exert extensive in vitro
anti-invasive and in vivo anti-metastatic activities. Cancer
Metastasis Rev. 31:323–351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jung SK, Lee M-H, Lim DY, Kim JE, Singh P,
Lee SY, Jeong CH, Lim TG, Chen H, Chi YI, et al: Isoliquiritigenin
induces apoptosis and inhibits xenograft tumor growth of human lung
cancer cells by targeting both wild type and L858R/T790M mutant
EGFR. J Biol Chem. 289:35839–35848. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee SK, Park K-K, Kim KR, Kim H-J and
Chung W-Y: Isoliquiritigenin inhibits metastatic breast cancer
cell-induced receptor activator of nuclear factor kappa-B
ligand/osteoprotegerin ratio in human osteoblastic cells. J Cancer
Prev. 20:281–286. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu CH, Chen HY, Wang CW, Shieh TM, Huang
TC, Lin LC, Wang KL and Hsia SM: Isoliquiritigenin induces
apoptosis and autophagy and inhibits endometrial cancer growth in
mice. Oncotarget. 7:73432–73447. 2016.PubMed/NCBI
|
|
12
|
Zhang X, Yeung ED, Wang J, Panzhinskiy EE,
Tong C, Li W and Li J: Isoliquiritigenin, a natural anti-oxidant,
selectively inhibits the proliferation of prostate cancer cells.
Clin Exp Pharmacol Physiol. 37:841–847. 2010.PubMed/NCBI
|
|
13
|
Zhou GS, Song LJ and Yang B:
Isoliquiritigenin inhibits proliferation and induces apoptosis of
U87 human glioma cells in vitro. Mol Med Rep. 7:531–536. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Platonova N, Lesma E, Basile A, Bignotto
M, Garavelli S, Palano MT, Moschini A, Neri A, Colombo M and
Chiaramonte R: Targeting Notch as a therapeutic approach for human
malignancies. Curr Pharm Des. 23:108–134. 2017.PubMed/NCBI
|
|
15
|
Majidinia M, Alizadeh E, Yousefi B,
Akbarzadeh M and Zarghami N: Downregulation of Notch signaling
pathway as an effective chemosensitizer for cancer treatment. Drug
Res (Stuttg). 66:571–579. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yahyanejad S, King H, Iglesias VS, Granton
PV, Barbeau LM, van Hoof SJ, Groot AJ, Habets R, Prickaerts J,
Chalmers AJ, et al: NOTCH blockade combined with radiation therapy
and temozolomide prolongs survival of orthotopic glioblastoma.
Oncotarget. 7:41251–41264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee CK, Son SH, Park KK, Park JH, Lim SS
and Chung WY: Isoliquiritigenin inhibits tumor growth and protects
the kidney and liver against chemotherapy-induced toxicity in a
mouse xenograft model of colon carcinoma. J Pharmacol Sci.
106:444–451. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duan JJ, Qiu W, Xu SL, Wang B, Ye XZ, Ping
YF, Zhang X, Bian XW and Yu SC: Strategies for isolating and
enriching cancer stem cells: Well begun is half done. Stem Cells
Dev. 22:2221–2239. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu B, Sun C, Feng F, Ge M and Xia L: Do
relevant markers of cancer stem cells CD133 and Nestin indicate a
poor prognosis in glioma patients? A systematic review and
meta-analysis. J Exp Clin Cancer Res. 34:442015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao S, Chang H, Ma P, Gao G, Jin C, Zhao
X, Zhou W and Jin B: Inhibitory effect of DNA topoisomerase
inhibitor isoliquiritigenin on the growth of glioma cells. Int J
Clin Exp Pathol. 8:12577–12582. 2015.PubMed/NCBI
|
|
21
|
Wang N, Wang Z, Peng C, You J, Shen J, Han
S and Chen J: Dietary compound isoliquiritigenin targets GRP78 to
chemosensitize breast cancer stem cells via β-catenin/ABCG2
signaling. Carcinogenesis. 35:2544–2554. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang N, Wang Z, Wang Y, Xie X, Shen J,
Peng C, You J, Peng F, Tang H, Guan X, et al: Dietary compound
isoliquiritigenin prevents mammary carcinogenesis by inhibiting
breast cancer stem cells through WIF1 demethylation. Oncotarget.
6:9854–9876. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen H, Zhang B, Yuan X, Yao Y, Zhao H,
Sun X and Zheng Q: Isoliquiritigenin-induced effects on Nrf2
mediated antioxidant defence in the HL-60 cell monocytic
differentiation. Cell Biol Int. 37:1215–1224. 2013.PubMed/NCBI
|
|
24
|
Chen X, Zhang B, Yuan X, Yang F, Liu J,
Zhao H, Liu L, Wang Y, Wang Z and Zheng Q:
Isoliquiritigenin-induced differentiation in mouse melanoma B16F0
cell line. Oxid Med Cell Longev. 2012:5349342012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Venere M, Fine HA, Dirks PB and Rich JN:
Cancer stem cells in gliomas: Identifying and understanding the
apex cell in cancer's hierarchy. Glia. 59:1148–1154. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Singh SK, Clarke ID, Terasaki M, Bonn VE,
Hawkins C, Squire J and Dirks PB: Identification of a cancer stem
cell in human brain tumors. Cancer Res. 63:5821–5828.
2003.PubMed/NCBI
|
|
27
|
Guo J, Qiang M and Ludueña RF: The
distribution of β-tubulin isotypes in cultured neurons from
embryonic, newborn, and adult mouse brains. Brain Res. 1420:8–18.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo J, Walss-Bass C and Ludueña RF: The
beta isotypes of tubulin in neuronal differentiation. Cytoskeleton
(Hoboken). 67:431–441. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gage FH and Temple S: Neural stem cells:
Generating and regenerating the brain. Neuron. 80:588–601. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim MY, Mo JS, Ann EJ, Yoon JH and Park
HS: Dual regulation of notch1 signaling pathway by adaptor protein
fe65. J Biol Chem. 287:4690–4701. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Purow BW, Haque RM, Noel MW, Su Q, Burdick
MJ, Lee J, Sundaresan T, Pastorino S, Park JK, Mikolaenko I, et al:
Expression of Notch-1 and its ligands, Delta-like-1 and Jagged-1,
is critical for glioma cell survival and proliferation. Cancer Res.
65:2353–2363. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fan X, Matsui W, Khaki L, Stearns D, Chun
J, Li YM and Eberhart CG: Notch pathway inhibition depletes
stem-like cells and blocks engraftment in embryonal brain tumors.
Cancer Res. 66:7445–7452. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang Y, Bai L, Li X, Xiong J, Xu P, Guo C
and Xue M: Transport of active flavonoids, based on cytotoxicity
and lipophilicity: An evaluation using the blood-brain barrier cell
and Caco-2 cell models. Toxicol In Vitro. 28:388–396. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Faria A, Pestana D, Teixeira D, Azevedo J,
De Freitas V, Mateus N and Calhau C: Flavonoid transport across
RBE4 cells: A blood-brain barrier model. Cell Mol Biol Lett.
15:234–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Youdim KA, Qaiser MZ, Begley DJ,
Rice-Evans CA and Abbott NJ: Flavonoid permeability across an in
situ model of the blood-brain barrier. Free Radic Biol Med.
36:592–604. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Faria A, Meireles M, Fernandes I,
Santos-Buelga C, Gonzalez-Manzano S, Dueñas M, de Freitas V, Mateus
N and Calhau C: Flavonoid metabolites transport across a human BBB
model. Food Chem. 149:190–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fonseca-Santos B, Gremião MP and Chorilli
M: Nanotechnology-based drug delivery systems for the treatment of
Alzheimer's disease. Int J Nanomedicine. 10:4981–5003. 2015.
View Article : Google Scholar : PubMed/NCBI
|