Introduction
Prostate cancer (PCa) is the most common malignant
tumor of the male genital system. It was estimated in 2017, that
161,360 new cancer cases were diagnosed and 26,730 deaths occurred
in the US (1). Androgen receptor
signaling is crucial for the initiation and progression of prostate
cancer (2). Initially, most PCa
cells respond to castration therapy, but eventually tumors relapse
and take on a castration therapy-resistant prostate cancer (CRPC)
phenotype that correlates with poor prognosis and high metastatic
potential. Although targeted drugs and immune checkpoint blockade
drugs have rapidly developed recently, mCRPC has exhibited
resistance to these treatments (3),
thus novel drug combination regimens or therapeutic approaches to
combat PCa are still urgently needed.
Poly (ADP-ribose) polymerases (PARPs) are a family
of related enzymes, and PARP1 is the founding and best
characterized member (4). PARP1 is
activated after binding to DNA strand breaks and uses
NAD+ as a substrate to form long branched polymers of
poly (ADP-ribose) (PAR). PARP1 is involved in many cellular
processes, including various DNA repair pathways and in the
maintenance of genomic stability (5). As a ubiquitously expressed
NAD+-dependent nuclear enzyme, PARP1 has prognostic
value for many human cancers (6).
Homologous recombination (HR) defects renders PCa sensitive to PARP
inhibition (7). However, the use of
PARP inhibitors (PARPi) has been limited to repair-deficient
cancers (8).
PARPi suppresses PARP activation but not its
expression. However, whether the function of small interfering RNA
(siRNA) specific to PARP is identical to PARPi has not been well
defined. Our previous study revealed that PARP1 and PAR expression
were markedly elevated in PCa tissues compared to benign prostate
hyperplasia (BPH) tissues (9), and
that inhibition of PARP1 by siRNA could enhance the activity of
docetaxel against PC3 cells (10).
However, the exact effect of PARP1-siRNA in PCa cell growth and
progression and its mechanism remain unclear. In the present study,
we investigated whether inhibition of PARP1 by siRNA could reduce
the growth and invasion capacity of PCa cell lines regardless of
the BRCA1/2 mutation.
Materials and methods
Cell culture and drugs
Du145, C4-2 and PC3 cells were purchased from the
American Type Culture Collection (ATCC; Manassas, VA, USA). Du145,
C4-2 and PC3 cells were cultured in Dulbecco's modified Eagle's
medium (DMEM), RPMI-1640 and DMEM/F12 medium respectively, with 10%
(v/v) fetal bovine serum (FBS; Gibco/Life Technologies; Thermo
Fisher Scientific, Inc., Grand Island, NY, USA) and 1% (v/v)
penicillin and streptomycin at 37°C in a humidified incubator
supplemented with 5% CO2. Docetaxel was obtained from
Selleck Chemicals (Houston, TX, USA). PC3 monoclonal cells stably
expressing pGPU6-GFP-Neo-PARP1-siRNA-1706 or
pGPU6-GFP-Neo-PARP1-siRNA-NC were generated and cultured in the
presence of G418 (0.5 mg/ml).
RNA interference and transfection
siRNA that targeted the human PARP1 gene was
synthesized by RiboBio Co., Ltd. (Guangzhou, China). The
synthesized siRNA sequences were the following: siRNA-NC,
5′-UUCUCCGAACGUGUCACGUTT-3′; siRNA-1706,
5′-GAGGAAGGUAUCAACAAAUTT-3′; siRNA-2003,
5′-GAGCACUUCAUGAAAUUAUTT-3′; and siRNA-2907,
5′-GAGACCCAAUAGGUUAAUTT-3′.
Transfection of synthesized siRNAs was carried out
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc., Carlsbad, CA, USA) according to the manufacturer's
instructions.
Cell viability assessment
Du145 cells were plated in 96-well plates. After 24
h, the cells were transfected with PARP1-siRNA-NC or PARP1-siRNA-1
using a riboFECT™ CP Transfection Kit from RiboBio Co., Ltd.
(Guangzhou, China). Approximately 72 h later, the cells were
replaced with DMEM without FBS and incubated for another 24 h.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was added to cells for a 3 to 4-h reaction, and dimethyl sulfoxide
(DMSO) was applied to dissolve the sediment. The optical density of
the MTT results was assessed at a wavelength of 490 nm and the cell
survival rate was normalized.
In vitro migration assay
Cells were transfected with the indicated siRNA and
cultured to confluence. Then, an artificial scratch wound was
generated using a 200-µl pipette tip, and cells were washed to
remove the detached cells and debris. Cell migration was captured
using an inverted microscope and evaluated by measuring the
difference in the wound width and counting the migrated number of
cells.
In vitro colony formation assay
Forty-eight hours after transfection, cells were
washed with phosphate-buffered saline (PBS), trypsinized, and
reseeded into 6-pore dishes at a density of 500 cells/well. Under
the same culture conditions, the cells were cultured for another
9–12 days, and then fixed and dyed with 4% crystal violet in
ethanol. The number of cell colonies was calculated.
In vitro invasion assay
Cell invasion was evaluated using a
Transwell® permeable support invasion chamber (Corning
Incorporated, Corning, NY, USA) according to the manufacturer's
protocol. Forty-eight hours after transfection, the cells were
cultured in the insert for 48 h and stained with Mayer's
hematoxylin. Eight fields in each sample were imaged, and invasion
was evaluated by counting the invaded number of cells.
In vivo xenograft model
All animal experimental procedures were carried out
in accordance with a mouse protocol, which was approved by the
Animal Research Committee of Guangzhou Medical University. Male
nude mice (aged 8 weeks; weighing 18–20 g) were fed a standard diet
and water during the study. Fifteen mice were randomly assigned to
3 experimental groups evenly (5 mice in each group):
PC3-siRNA-1706, PC3-siRNA-NC and PC3 C (control). PC3 monoclonal
cells stably expressing pGPU6-GFP-Neo-PARP1-siRNA-1706 or
pGPU6-GFP-Neo-PARP1-siRNA-NC were inoculated subcutaneously in the
flank regions of the mice to establish the tumor model. Following
subcutaneous inoculation, the mice were examined daily for tumor
formation and growth. The sizes of the subcutaneous tumors were
monitored twice weekly by assessing the length and width. The tumor
volumes were calculated using the formula: Volume (mm3)
= length (mm) × width2 (mm2)/2. At 40 days
after inoculation, all experimental mice were euthanized for
further study.
Western blot analysis
For protein extraction, cells or homogenized tissue
were rinsed with PBS and treated in ice-cold lysis buffer. After
scrape, collection and ultra-sonication, the lysate solutions were
centrifuged at 12,000 RCF for 30 min at 4°C. The Bradford protein
assay method was used to assess the protein concentration. Equal
amounts of protein were separated on a 10% SDS polyacrylamide gel
and transferred to a NC membrane (Millipore, Billerica, MA, USA).
The membranes were blocked in 5% non-fat dry milk in PBS-T for 2 h
at room temperature and incubated with primary antibodies
appropriately diluted in PBS-T overnight at 4°C. Primary antibodies
against PARP1 (1:200; mouse, monoclonal; cat. no. sc-8007),
vimentin (1:200; mouse, monoclonal; cat. no. sc-6260) and GAPDH
(1:1,000; mouse, monoclonal; cat. no. sc-47724) were obtained from
Santa Cruz Biotechnology (Santa Cruz, CA, USA). EGFR (1:2,000;
rabbit, monoclonal; cat. no. 1902-1) was obtained from Epitomics
(Cambridge, UK). Anti-tubulin (1:2,000; rabbit, ployclonal; cat.
no. 2148), GSK3-β (1:1,000; rabbit, monoclonal; cat. no. 12456),
pGSK3-β (ser9) (1:1,000; rabbit, monoclonal; cat. no. 5558),
E-cadherin (1:1,000; mouse, monoclonal; cat. no. 14472), Cx43
(1:1,000; rabbit, ployclonal; cat. no. 3512) and cleaved-caspase3
(1:1,000; rabbit, monoclonal; cat. no. 9664) were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA). The membranes
were incubated with secondary antibodies appropriately diluted in
PBS-T for 2 h at room temperature. The secondary antibodies were
anti-mouse or anti-rabbit IgG conjugated to HRP (Santa Cruz
Biotechnology). The membranes were developed for chemiluminescence
detection using an ECL detection kit (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The band intensities were quantified using
ImageJ software, and all western blot analyses were repeated 3
times.
Quantitative real-time PCR
analysis
Total RNA was extracted with a MiniBEST Universal
RNA Extraction kit (Takara, Otsu, Japan). An equal amount of RNA
was reverse transcribed to cDNA and amplified by PCR according to
the manufacturer's protocol (Takara). Quantitative real-time PCR
was performed using the SYBR-Green PCR Master mix (Takara)
according to the manufacturer's protocol. The primers were as
follows: PARP1 forward, 5′-AAGGCGAATGCCAGCGTTAC-3′ and reverse,
5′-GGCACTCTTGGAGACCATGTCA-3′; E-cadherin forward,
5′-TCCTCCCAATACATCTCCCTTCA-3′ and reverse,
5′-TCTCCGCCTCCTTCTTCATCATA-3′; vimentin forward
5′-TTCGCCAACTACATCGACAAGG-3′and reverse,
5′-TTCAAGGTCAAGACGTGCCAG-3′; GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′and reverse,
5′-AUAAUUUCAUGAAGUGC-UCTT-3′. The real-time PCR reaction was
maintained at 95°C for 5 min, followed by 40 cycles at 95°C for 15
sec, 57°C for 30 sec and 72°C for 20 sec. All samples were
processed in triplicate, and all values were normalized to the
expression levels of GAPDH. The relative quantification was
determined by the comparative Ct method.
Hematoxylin and eosin (H&E)
staining
The tumor tissues were washed with PBS, fixed with
4% paraformaldehyde, paraffin-embedded, and sliced to a 3-µm
thickness to generate serial sections. Slices were dewaxed in
xylene for 10 min and placed into a mixture of alcohol and xylene
(1:1) for ~5 min. After the slices were sequentially immersed in
100, 95, 85 and 70% alcohol for 2 min per step, they were placed in
distilled water before staining. Then, the slices were stained with
hematoxylin for ~10 min. Subsequently, the slices were immersed
into water for 1 sec, and then 0.5–1% hydrochloric acid alcohol was
used for a short time to clear the excess hematoxylin. The time was
controlled by microscopy until the nuclei and nuclear chromatin
were clear. After rinsing with water for 15–30 min, the nuclei
became blue, and the slices were rinsed again in distilled water
(short time). Thereafter, 0.5% eosin was applied to stain the
slices for 1 min, and the slices were sequentially immersed in 70,
85, 95, and 100% alcohol for dehydration for 2–3 min per step. The
slices were then cleared with xylene two times (5 min/time). Excess
xylene in the sections was wiped away, and the specimens were not
dried. A small amount of neutral gum was quickly added, and the
glass was covered to seal the specimens.
Immunohistochemistry
All sections were formalin-fixed, paraffin-embedded
and stained according to the standard immunohistochemistry
protocol. Following incubation with primary antibodies for PARP1,
E-cadherin and vimentin, the sections were incubated with
biotinylated secondary antibodies (Maixin Biotech Co., Ltd.,
Fuzhou, China). Subsequently, the avidin-biotin horseradish
peroxidase complex and diaminobenzidine chromogen (Maixin Biotech
Co., Ltd.) were used as the detection system. Each stained slide
was assessed by an experienced laboratory technician independently
in a blinded fashion without any information regarding the
group.
Statistical analysis
Statistical analysis was performed using SPSS 16.0
software (SPSS, Inc., Chicago, IL, USA). All data are expressed as
the mean ± SD. The mean differences (P<0.05) were compared using
a one-way ANOVA (>2 groups) or Student's t-test (2 groups).
Western blot data was analyzed with ImageJ software. In the
figures, an asterisk represents P<0.05 compared to the
corresponding group and indicates a statistically significant
result.
Results
siRNA-mediated PARP1 inhibition
suppresses migration, invasion and proliferation of PC3 cells in
vitro
Two small interfering RNAs (1706 and 2003) used to
knock down the expression of PARP1 in PC3 cells in our previous
study (10), were synthesized and
transfected into PC3 cells. Real-time PCR and western blot analyses
revealed that both PARP1 siRNAs, 1706 and 2003, significantly
decreased PARP1 mRNA and protein expression compared to the
negative control in PC3 cells (Fig. 1A
and B). Then, we investigated the effect of PARP1-siRNA on the
migration and invasion capacity of PC3 cells. The results of the
wound healing assays revealed that both siRNA-1706 and siRNA-2003
significantly suppressed the migration capacities of PC3 cells
in vitro (Fig. 1C). The
results of the Transwell invasion assays revealed that both
siRNA-1706 and siRNA-2003 significantly suppressed the invasion
capacities of PC3 cells in vitro (Fig. 1D).
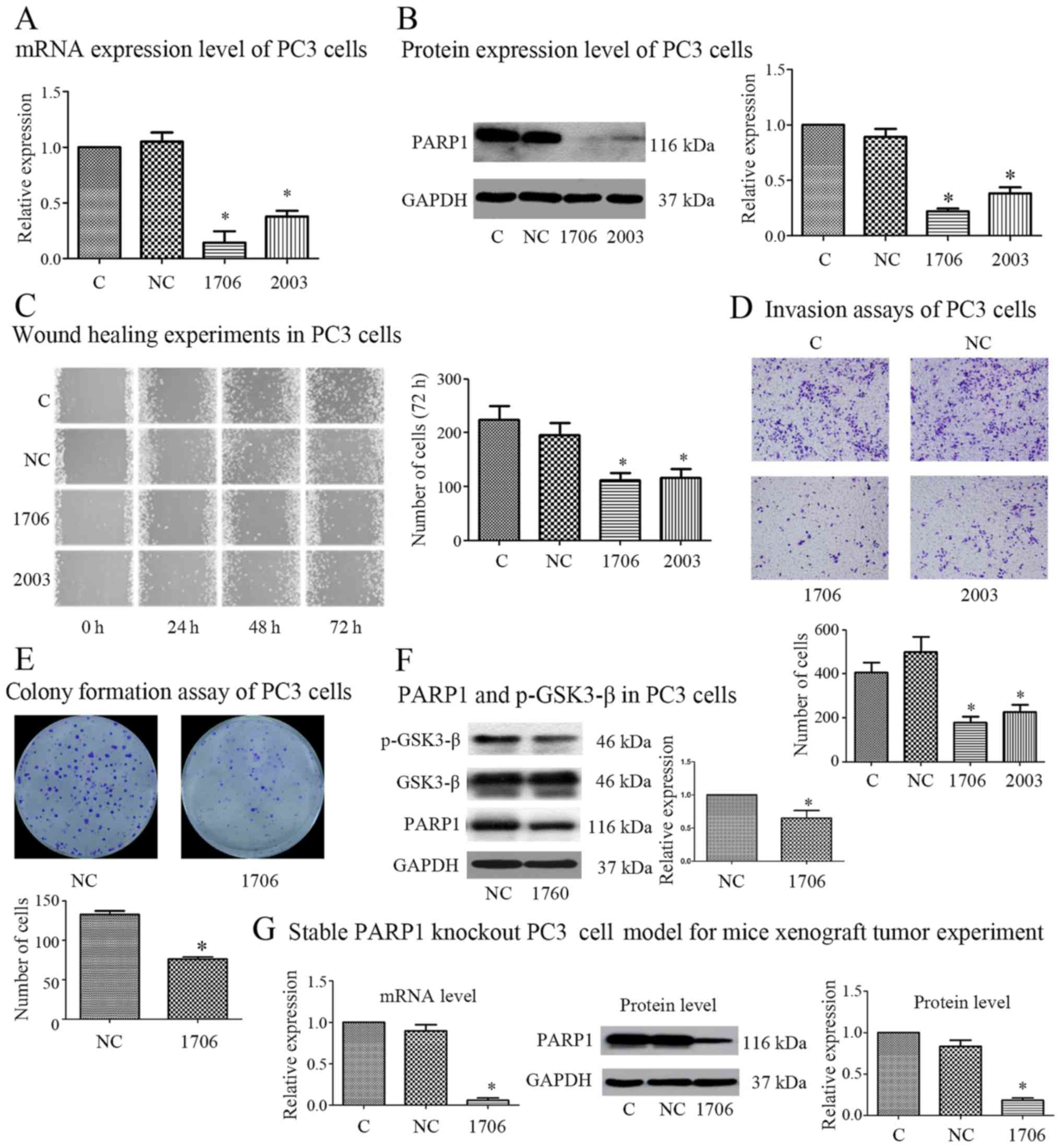 | Figure 1.siRNA-mediated PARP1 inhibition
suppresses the migration, invasion and growth capacities of PC3
cells in vitro. (A and B) Following transfection with the
indicated siRNA, the expression of PARP1 and GAPDH in PC3 cells was
detected by real-time PCR and western blotting. Following
transfection with the indicated siRNA in PC3 cells, (C) cell
migration capacity was detected by wound healing assay, (D) cell
invasion capacity was detected by Transwell invasion assay, (E)
cell proliferation ability was assessed by colony formation assay,
and (F) the expression of p-GSK-3β(Ser9) was detected by western
blotting. (G) A stable model of transfection with the indicated
siRNA in PC3 cells to perform xenograft tumor experiments was
constructed, and the expression of PARP1 and GAPDH was detected by
real-time PCR and western blotting. n=3. *P<0.05 with respect to
the control group. C, control group; NC, negative control group;
1706, siRNA-1706 group; 2003, siRNA-2003 group. |
Since siRNA-1706 was more effective than siRNA-2003
in suppressing the expression of PARP1, siRNA-1706 was further used
in the following experiment. Colony formation assays were analyzed,
and the results revealed that the siRNA-1706 group grew
significantly more slowly than the negative control group (Fig. 1E). In addition, our previous study
confirmed that PARP1 siRNAs could inhibit the EGFR/Akt/FoxO
pathway. In the present study, we found that PARP1-siRNA could
inhibit p-GSK-3β which is also downstream of AKT (Fig. 1F). Therefore, PARP1 knockdown may
suppress PCa progression by inhibiting EGFR/Akt and its downstream
including the FoxO and GSK-3β signaling pathway. To further
investigate whether PARP1 knockdown reduced the tumorigenesis and
metastasis capacity of PCa cells in vivo, we constructed a
stable PARP1 knockout model in PC3 cells by recombinant plasmid.
The PCR and western blot results revealed that stable transfection
of the plasmid could effectively inhibit the expression of PARP1
(Fig. 1G).
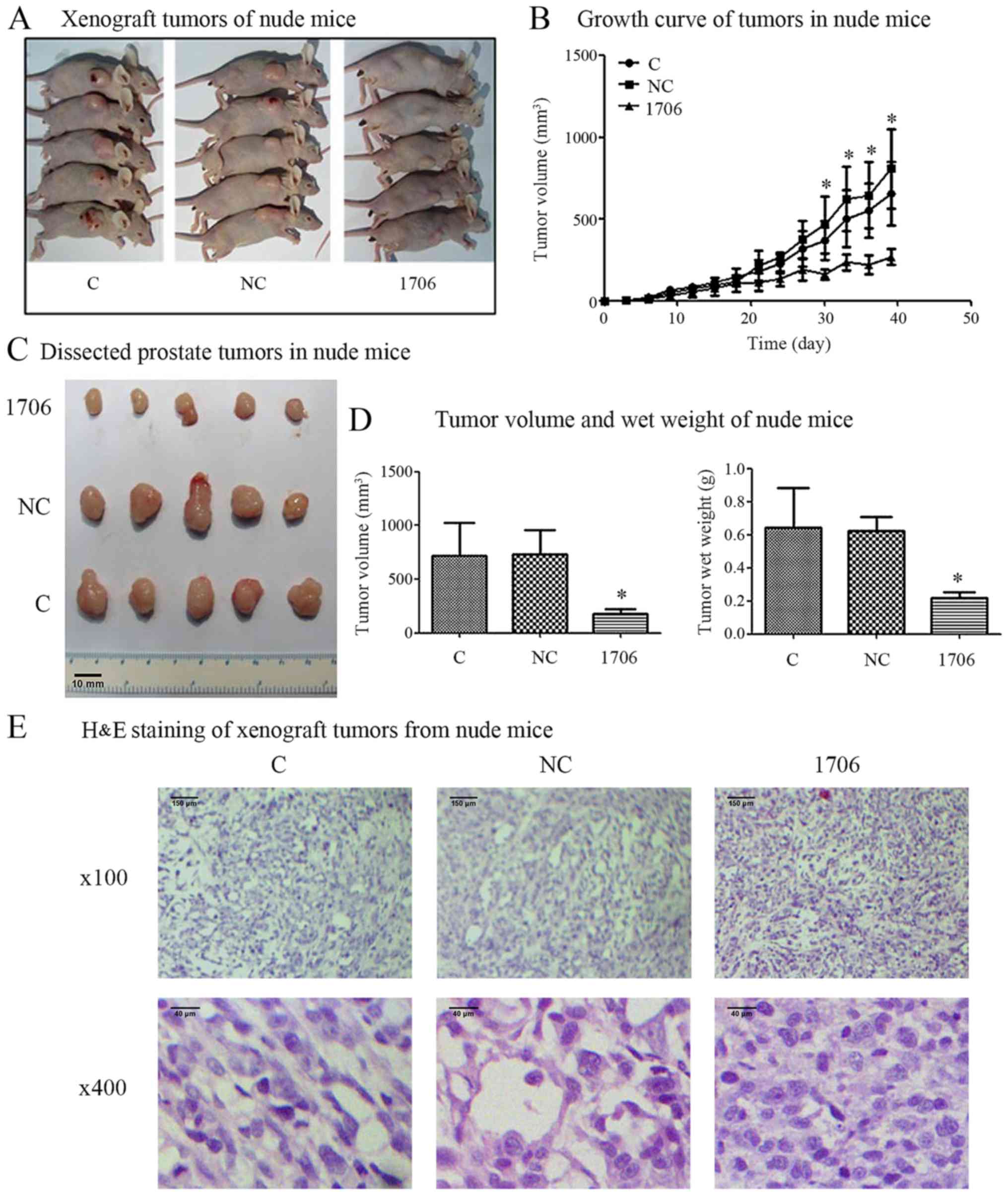
siRNA-mediated PARP1 inhibition
reduces the growth of PC3 cells in vivo
After a stable PARP1 knockout model was constructed
and confirmed, we investigated whether PARP1 knockdown could reduce
the tumor growth and tumorigenesis capacity of PCa cells in
vivo. The results revealed that in comparison with the PC3
control (C) group and the PC3-siRNA-negative control group (NC),
the tumor growth in the PC3-siRNA-1706 group was markedly reduced
(Fig. 2A and B). The mean volumes
and wet weights of the tumors from the PC3-siRNA-1706 group were
significantly decreased in comparison with the PC3 control group
and the PC3-siRNA-NC group (Fig. 2C and
D). The H&E staining results revealed that the control
group and no-load group promoted a larger cell volume, rich in
cytoplasm, in which the nucleus was larger and deeply stained; the
nuclei were round and had prominent nucleoli. In this group,
mitotic activity was more common, the nucleus/cytoplasm ratio was
increased, cells were rich in cytoplasm, and there was a higher
number of interstitial vessels. The PC3-siRNA-1706 group exhibited
smaller PC3 cells in the nude mice with smaller cytoplasms, in
which the nuclei were small, round and lighter in color; the
nucleoli were obvious, the nucleus/cytoplasm ratio was smaller, and
there were fewer interstitial blood vessels (Fig. 2E).
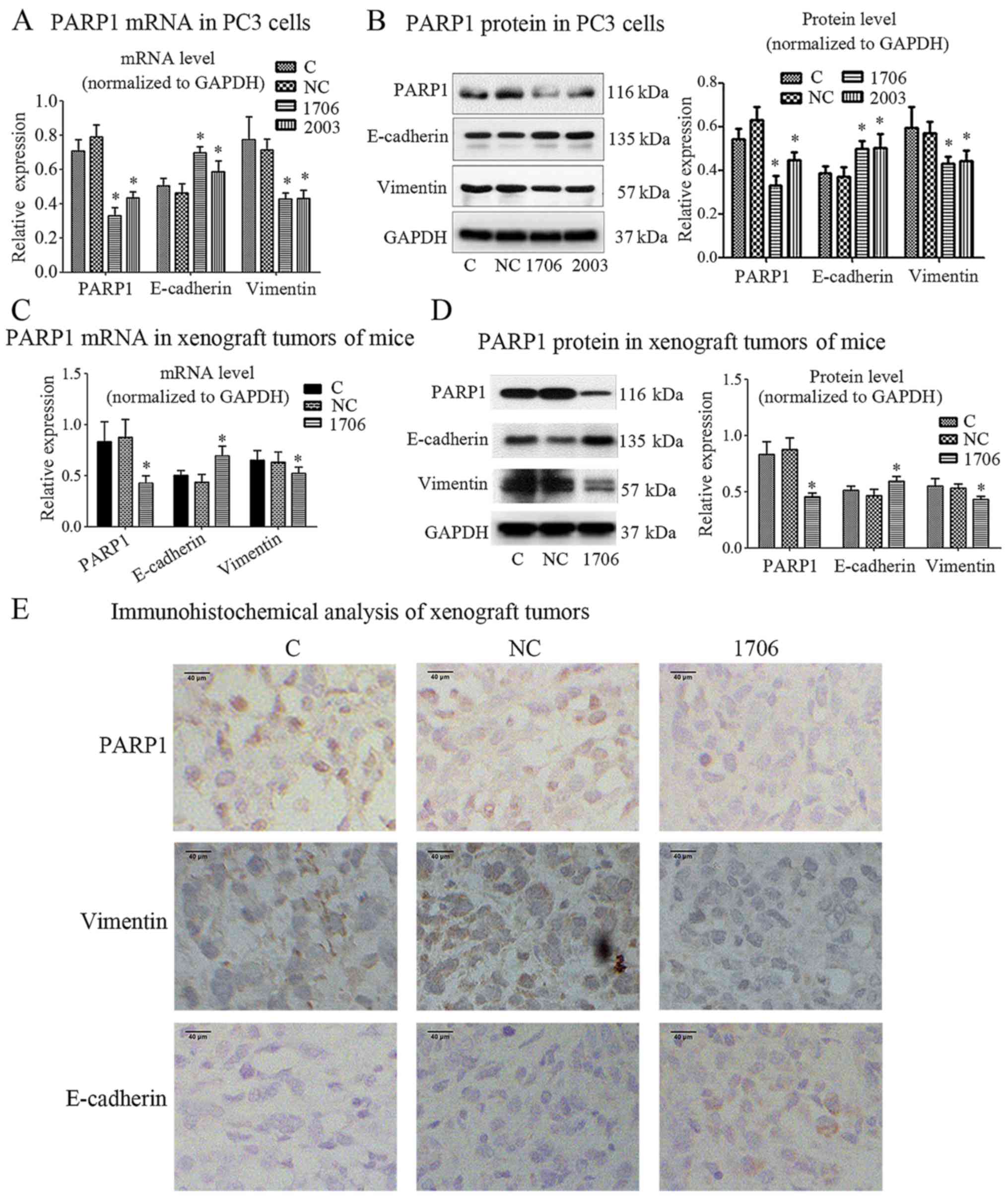
PARP1 silencing and EMT molecular
marker expression
Since EMT is a key step for cancer cells to acquire
invasion and metastasis abilities (11), we analyzed the expression of some
EMT-associated markers in PC3 cells by PARP1 knockdown. Real-time
PCR analysis revealed that PARP1 knockdown significantly
upregulated the mRNA level of E-cadherin, which was accompanied
with significant downregulation of vimentin mRNA (Fig. 3A). The western blot analysis results
further validated the increased E-cadherin expression and decreased
vimentin expression in PC3 cells after knockdown of PARP1 (Fig. 3B). To further explore whether PARP1
knockdown suppressed EMT in PCa cells in vivo, we detected
the expression of EMT-associated markers in xenograft tumors.
Consistent with the in vitro results, real-time PCR and
western blot analyses revealed the same results in vivo
(Fig. 3C and D).
Immunohistochemical analysis revealed that, compared with the PC3
control group and the PC3-siRNA-NC group, the PC3-siRNA-1706 group
had a higher level of the E-cadherin protein and a lower level of
the vimentin protein (Fig. 3E).
siRNA-mediated PARP1 inhibition
suppresses the EGFR signaling pathway in C4-2 and DU145 cells
In order to further confirm the function of PARP1
siRNAs in other PCa cells, siRNA-1706, siRNA-2003, siRNA-2907 and
non-specific siRNA [negative control (NC)] were used to knock down
the expression of PARP1 in C4-2 cells. The results revealed that
siRNA-1706 was the most efficient and was further used in the
following experiment (Fig. 4A).
After PARP1 knockdown by PARP1 siRNA-1706, EGFR was decreased in
C4-2 cells (Fig. 4B). After PARP1
was knocked down by PARP1 siRNA-1706 in C4-2 cells, docetaxel was
used to induce apoptosis and the expression of cleaved-caspase3 was
detected via western blotting (Fig.
4C). Connexin 43 (Cx43) may be associated with apoptosis since
it is a component of gap junctions, therefore its expression was
also detected (Fig. 4C). Compared
with the NC group, EGFR expression in the PARP1 siRNA-1706 group
was significantly decreased in Du145 cells (Fig. 4D). Similar to the C4-2 cells,
cleaved-caspase3 and Cx43 expression in the siRNA-1706 group was
significantly increased with docetaxel (100 nM) treatment in Du145
cells (Fig. 4E). After PARP1 was
knocked down by PARP1 siRNA-1706 in Du145 cells, its proliferation
was decreased (Fig. 4F).
Collectively, these results indicated that PARP1 knockdown reduced
the progression and metastasis capacities of PCa cells both in
vitro and in vivo.
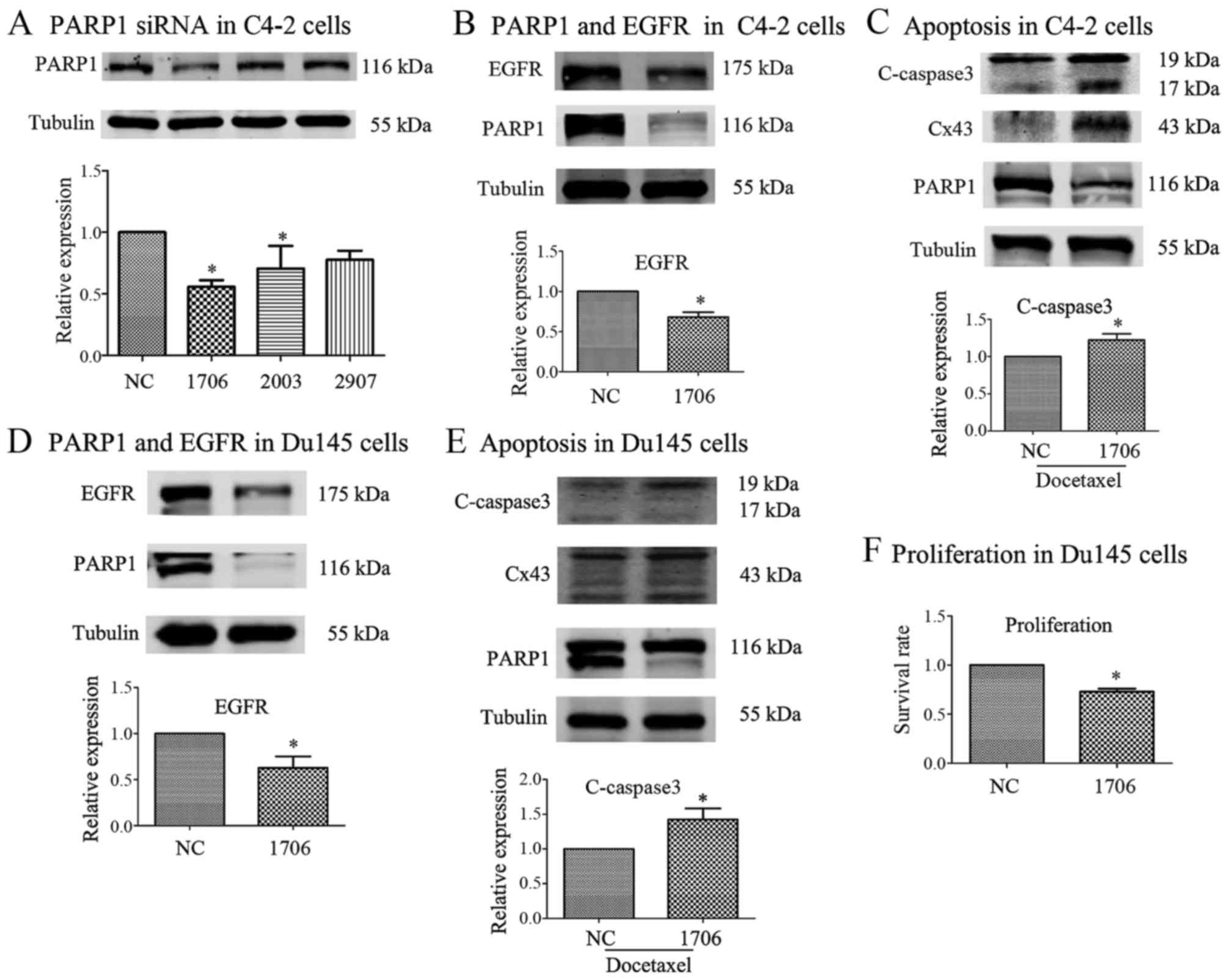 | Figure 4.siRNA-mediated PARP1 inhibition
suppresses the EGFR signaling pathway in C4-2 and DU145 cells. (A)
Non-specific siRNA was used as a negative control (NC), and 3 PARP1
siRNAs (siRNA-1706, −2003 and −2907) were used to knock down PARP1
expression. siRNA-1706 was the most efficient and further used in
the following experiment. (B) Following PARP1 knockdown by PARP1
siRNA-1706, EGFR was decreased in C4-2 cells. (C) Following PARP1
knockdown by PARP1 siRNA-1706 in C4-2 cells, docetaxel was used to
induce apoptosis and expression of cleaved-caspase3 was detected
via western blotting. (D) Compared with the NC group, EGFR
expression in the PARP1 siRNA-1706 group was significantly
decreased in Du145 cells. (E) Similar to the C4-2 cells, c-caspase3
expression in the siRNA-1706 group was significantly increased by
docetaxel treatment in Du145 cells. (F) Following PARP1 knockdown
by PARP1 siRNA-1706 in Du145 cells, its proliferation was
decreased. n=3. *P<0.05 with respect to the negative control
group. C-caspase3, cleaved-caspase3; C, control group; NC, negative
control group; 1706, siRNA-1706 group; 2003, siRNA-2003 group;
2907, siRNA-2907 group. |
Discussion
PCa is a major health problem in older men
worldwide. Progression and metastasis are considered to be major
challenges for PCa treatment. Therefore, studies on the mechanism
of PCa progression and metastasis, and the corresponding
therapeutic methods are urgently required. In the present study,
our results indicated that PARP1-siRNA could suppress the
progression and invasion capacity of PCa cells regardless of the
BRCA1/2 mutation by inhibiting the EGFR/GSK-3β signaling pathway.
Notably, this function of PARP1-siRNA in PCa cells may be different
from PARPi and may provide a potential therapy method for human
CRPC.
PARP1 is an enzyme that catalyzes the covalent
attachment of polymers of ADP-ribose (PAR) moieties on itself and
its target proteins. PARP1 is overexpressed in various tumors and
its overexpression is associated with poor overall survival
(12–15). In fact, targeting DNA repair defects
with PARP1 inhibitors (PARPi) is emerging as a cancer treatment
strategy, especially in BRCA1 mutation cases. However, another
suitable predictive biomarker is still required, since resistance
to PARPi frequently occurs (16–18).
For this reason, we chose PARP1-siRNA instead of PARPi in our
study. We first investigated the effect of PARP1-siRNA on the
progression and invasion capacity of PCa cells. The results
revealed that PARP1-siRNA could act as a tumor progression and
invasion suppressor in PCa cells, suppressing PCa cell
proliferation, migration and invasion capacities in vitro
and tumor growth in vivo.
Initiation of EMT is closely related to drug
resistance, tumor relapse as well as metastasis (19). The expression changes of associated
markers, such as E-cadherin and vimentin, are important
characteristics of EMT (20).
Impaired PARP-1 function promotes prostate tumorigenesis in
vivo via TGF-β-induced EMT (21). However, our results revealed that
PARP1-siRNA significantly altered EMT markers in PC3 cells in
vitro and in vivo, which was visualized by the
upregulation of E-cadherin and the downregulation of vimentin in
PC3 cells with the knockdown of PARP1. These results indicated that
PARP1-siRNA may suppress the progression and metastasis capacities
of prostate cancer cells by suppressing EMT.
EGFR expression was required for primary and
secondary sphere formation of prostate cancer cells (22), and EGFR is a key inducer of EMT in
cancer progression (23,24), since E-cadherin is the downstream of
the EGFR signal (25). EGF binding
to EGFR displays intrinsic tyrosine kinase activity and activates
downstream signals via phosphorylation of key proteins. Among them,
Akt/GSK-3β signaling is a key downstream target of EGFR signaling
(26). In addition, Akt/GSK-3β
signaling is crucial for the EMT of prostate cancer cells (27). PARPi inactivated GSK-3β, which in
turn enhanced PARPi-mediated PD-L1 upregulation (28). Thus, whether PARP1-siRNA could
upregulate PD-L1 the same way as PARPi warrants further
investigation. Our previous study revealed that PARP1-siRNA
enhanced the activity of docetaxel against PC3 cells, and was
associated with an accelerated suppression of the EGF/Akt/FOXO1
signaling pathway and was contrary to ABT88 (a PARPi) (10). However, GSK-3β is also downstream of
the Akt signaling pathway. Therefore, we investigated whether
PARP1-siRNA altered the phosphorylation status of GSK-3β in PC3
cells. Our results revealed that PARP1-siRNA could clearly inhibit
the expression of EGFR and p-GSK-3β. Notably, whether
overexpression of EGFR signaling could abolish the function of
PARP1-siRNA warrants further investigation. Taking our previous and
present study into consideration, PARP1-siRNA, unlike ABT88
(10), may suppress EGF/EGFR/Akt
signaling, with GSK-3β and FoxO1 as down targets, and is highly
correlated with tumor cell growth, EMT and other properties.
Acquired cisplatin-resistant ovarian cancer cells
expressed high levels of PARP-1 proteins, and silencing of PARP-1
increased cisplatin sensitivity in resistant cells (29). Notably, whether PARP1-siRNA can
increase cisplatin sensitivity in resistant PCa cells warrants
further investigation. Besides DNA damage repair function (30), emerging evidence also suggests that
PARP1 has close connections with the transcriptional activities of
the androgen receptor (AR) in PCa. Whether the function of
PARP1-siRNA could regulate AR in PCa cells warrants further
investigation.
In conclusion, the present study indicated that
PARP1-siRNA suppressed the growth, migration and invasion capacity
of PCa cells. Our study also identified a novel role for
PARP1-siRNA in modulating EMT and EGFR/Akt/GSK-3β signaling in PCa
cells. Furthermore, PARP1-siRNA may provide a potential new
therapeutic approach for the treatment of PCa, and we speculate
that the signaling pathway associated with PARP1 knockout may be
different from PARPi in PCa cells.
Acknowledgements
The present study was sustained in part by the
National Natural Science Foundation of China (contract nos.
81402430 and 81602541).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blessing AM, Rajapakshe K, Reddy Bollu L,
Shi Y, White MA, Pham AH, Lin C, Jonsson P, Cortes CJ, Cheung E, et
al: Transcriptional regulation of core autophagy and lysosomal
genes by the androgen receptor promotes prostate cancer
progression. Autophagy. 13:506–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu X, Horner JW, Paul E, Shang X, Troncoso
P, Deng P, Jiang S, Chang Q, Spring DJ, Sharma P, et al: Erratum:
Effective combinatorial immunotherapy for castration-resistant
prostate cancer. Nature. 543:1162017. View Article : Google Scholar
|
|
4
|
Lin KY and Kraus WL: PARP Inhibitors for
cancer therapy. Cell. 169:1832017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ray Chaudhuri A and Nussenzweig A: The
multifaceted roles of PARP1 in DNA repair and chromatin
remodelling. Nat Rev Mol Cell Biol. 18:610–621. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schiewer MJ and Knudsen KE:
Transcriptional roles of PARP1 in cancer. Mol Cancer Res.
12:1069–1080. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Asim M, Tarish F, Zecchini HI, Sanjiv K,
Gelali E, Massie CE, Baridi A, Warren AY, Zhao W, Ogris C, et al:
Synthetic lethality between androgen receptor signalling and the
PARP pathway in prostate cancer. Nat Commun. 8:3742017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ning J, Wakimoto H, Peters C, Martuza RL
and Rabkin SD: Rad51 degradation: Role in oncolytic
virus-poly(ADP-ribose) polymerase inhibitor combination therapy in
glioblastoma. J Natl Cancer Inst. 109:1–13. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu W, Zhu H, Liang Y, Kong Z, Duan X, Li
S, Zhao Z, Yang D and Zeng G: Expression of PARP-1 and its active
polymer PAR in prostate cancer and benign prostatic hyperplasia in
Chinese patients. Int Urol Nephrol. 46:1345–1349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu W, Kong Z, Duan X, Zhu H, Li S, Zeng S,
Liang Y, Iliakis G, Gui Z and Yang D: Inhibition of PARP1 by small
interfering RNA enhances docetaxel activity against human prostate
cancer PC3 cells. Biochem Biophys Res Commun. 442:127–132. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee JY and Kong G: Roles and epigenetic
regulation of epithelial-mesenchymal transition and its
transcription factors in cancer initiation and progression. Cell
Mol Life Sci. 73:4643–4660. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rojo F, Garcia-Parra J, Zazo S, Tusquets
I, Ferrer-Lozano J, Menendez S, Eroles P, Chamizo C, Servitja S,
Ramírez-Merino N, et al: Nuclear PARP-1 protein overexpression is
associated with poor overall survival in early breast cancer. Ann
Oncol. 23:1156–1164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Donizy P, Pietrzyk G, Halon A, Kozyra C,
Gansukh T, Lage H, Surowiak P and Matkowski R: Nuclear-cytoplasmic
PARP-1 expression as an unfavorable prognostic marker in lymph
node-negative early breast cancer: 15-year follow-up. Oncol Rep.
31:1777–1787. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Y, Zhang Y, Zhao Y, Gao D, Xing J and
Liu H: High PARP-1 expression is associated with tumor invasion and
poor prognosis in gastric cancer. Oncol Lett. 12:3825–3835. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galia A, Calogero AE, Condorelli R,
Fraggetta F, La Corte A, Ridolfo F, Bosco P, Castiglione R and
Salemi M: PARP-1 protein expression in glioblastoma multiforme. Eur
J Histochem. 56:e92012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kurfurstova D, Bartkova J, Vrtel R,
Mickova A, Burdova A, Majera D, Mistrik M, Kral M, Santer FR,
Bouchal J and Bartek J: DNA damage signalling barrier, oxidative
stress and treatment-relevant DNA repair factor alterations during
progression of human prostate cancer. Mol Oncol. 10:879–894. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Choi YE, Meghani K, Brault ME, Leclerc L,
He YJ, Day TA, Elias KM, Drapkin R, Weinstock DM, Dao F, et al:
Platinum and PARP inhibitor resistance due to overexpression of
MicroRNA-622 in BRCA1-Mutant ovarian cancer. Cell Rep. 14:429–439.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Y, Krais JJ, Bernhardy AJ, Nicolas E,
Cai KQ, Harrell MI, Kim HH, George E, Swisher EM, Simpkins F and
Johnson N: RING domain-deficient BRCA1 promotes PARP inhibitor and
platinum resistance. J Clin Invest. 126:3145–3157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tong D, Liu Q, Liu G, Xu J, Lan W, Jiang
Y, Xiao H, Zhang D and Jiang J: Metformin inhibits
castration-induced EMT in prostate cancer by repressing
COX2/PGE2/STAT3 axis. Cancer Lett. 389:23–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chaudhry P, Fabi F, Singh M, Parent S,
Leblanc V and Asselin E: Prostate apoptosis response-4 mediates
TGF-β-induced epithelial-to-mesenchymal transition. Cell Death Dis.
5:e10442014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pu H, Horbinski C, Hensley PJ, Matuszak
EA, Atkinson T and Kyprianou N: PARP-1 regulates
epithelial-mesenchymal transition (EMT) in prostate tumorigenesis.
Carcinogenesis. 35:2592–2601. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Day KC, Lorenzatti Hiles GL, Kozminsky M,
Dawsey SJ, Paul A, Broses LJ, Shah R, Kunja LP, Hall C, Palanisamy
N, et al: HER2 and EGFR overexpression support metastatic
progression of prostate cancer to bone. Cancer Res. 77:74–85. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu XL, Zhang XT, Meng J, Zhang HF, Zhao
Y, Li C, Sun Y, Mei QB, Zhang F and Zhang T: ING5 knockdown
enhances migration and invasion of lung cancer cells by inducing
EMT via EGFR/PI3K/Akt and IL-6/STAT3 signaling pathways.
Oncotarget. 8:54265–54276. 2017.PubMed/NCBI
|
|
24
|
El Bezawy R, Cominetti D, Fenderico N,
Zuco V, Beretta GL, Dugo M, Arrighetti N, Stucchi C, Rancati T,
Valdagni R, et al: miR-875-5p counteracts epithelial-to-mesenchymal
transition and enhances radiation response in prostate cancer
through repression of the EGFR-ZEB1 axis. Cancer Lett. 395:53–62.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim GT, Lee SH and Kim YM: Torilis
japonica extract, a new potential EMT suppressor agent by
regulation of EGFR signaling pathways. Int J Oncol. 45:1673–1679.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei A, Fan B, Zhao Y, Zhang H, Wang L, Yu
X, Yuan Q, Yang D and Wang S: ST6Gal-I overexpression facilitates
prostate cancer progression via the PI3K/Akt/GSK-3β/β-catenin
signaling pathway. Oncotarget. 7:65374–65388. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo H, Luo H, Yuan H, Xia Y, Shu P, Huang
X, Lu Y, Liu X, Keller ET, Sun D, et al: Litchi seed extracts
diminish prostate cancer progression via induction of apoptosis and
attenuation of EMT through Akt/GSK-3β signaling. Sci Rep.
7:416562017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiao S, Xia W, Yamaguchi H, Wei Y, Chen
MK, Hsu JM, Hsu JL, Yu WH, Du Y, Lee HH, et al: PARP inhibitor
upregulates PD-L1 expression and enhances cancer-associated
immunosuppression. Clin Cancer Res. 23:3711–3720. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang J, Kho DH, Zhou JY, Davis RJ and Wu
GS: MKP-1 suppresses PARP-1 degradation to mediate cisplatin
resistance. Oncogene. 36:5939–5947. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schiewer MJ, Goodwin JF, Han S, Brenner
JC, Augello MA, Dean JL, Liu F, Planck JL, Ravindranathan P,
Chinnaiyan AM, et al: Dual roles of PARP-1 promote cancer growth
and progression. Cancer Discov. 2:1134–1149. 2012. View Article : Google Scholar : PubMed/NCBI
|