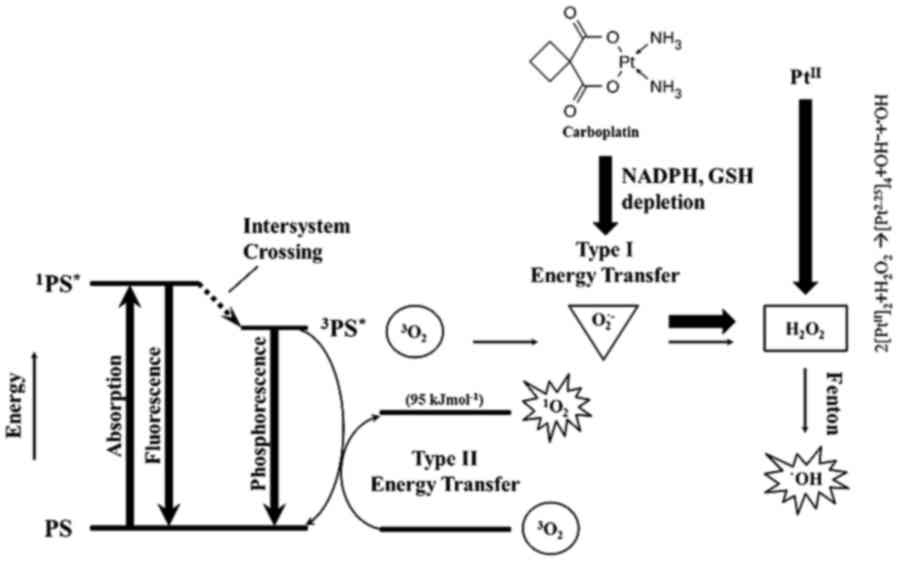
Introduction
Photodynamic therapy (PDT) is a local therapy; thus,
in principle, PDT can be safely combined with any systemic therapy
by exploiting non-overlapping cellular targets. Previous studies
have demonstrated the enhancement of the therapeutic efficacy of
combination treatment with carboplatin (CBP) and PDT (1–7)
through synergism (2) and the
subsequent ROS-mediated downregulation of biological functions
(3,4) or modulation of EGRF/PARP protein
expression (5). In particular,
low-dose CBP combined with Photofrin®-PDT (ccPDT)
resulted in a relapse-free period of more than 3 years when used to
treat cervical or endometrial cancer patients, while also
preserving fertility and enabling the successful delivery of
babies, as determined in our previous clinical efficacy study
(1). However, the involvement of
specific reactive oxygen species (ROS) and related cell death
pathways in the therapeutic enhancement by ccPDT remains to be
elucidated despite ROS-mediated synergistic therapeutic enhancement
becoming increasingly evident (2–4).
Anticancer metal complexes (platinum, gold, arsenic, ruthenium,
rhodium, copper, vanadium, cobalt, manganese, gadolinium and
molybdenum) have been shown to strongly interact with or even
disturb cellular redox homeostasis, and ROS generation via the
Fenton reaction is known (8).
The destructive power of large-scale ROS production
is highlighted by the fact that PDT uses photoactivation of
chemicals that produce ROS; primarily, but not exclusively, singlet
oxygen (1O2), to kill cancer cells and to
treat local infections (9). The
predominant type of ROS generated by the photosensitizer depends on
the type of assembly of photosensitizer molecules (monomer, dimer),
the reaction and local oxygen concentrations, such that Type I
reactions produce O2•– while type II produce
1O2 (10,11).
It was suggested that 1O2 produced at
plasma/ER/Golgi membranes mediates necrosis, whereas ROS other than
1O2 likely produced by the mitochondria,
activate the intrinsic apoptosis pathway (4,9,11–13).
While most clinically approved photosensitizers (porphyrins,
chlorins, or chemically related species) have high efficiency to
produce 1O2 (9,11),
enhanced ROS production via either electron or energy transfer or a
new cytotoxic pathway may improve the therapeutic efficacy of
combined treatment with different biological consequences compared
to PDT alone. In this study, ccPDT-induced modulation of ROS
production and cellular death were examined to elucidate the
mechanism of the therapeutic efficacy in ccPDT.
Materials and methods
Materials
Photofrin® was kindly provided by
LightpharmTech (Seoul, Korea). Carboplatin was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Fetal bovine serum
(FBS) was purchased from MP Biomedicals, LLC (Solon, OH, USA).
RPMI-1640, PBS and penicillin-streptomycin (P/S) were obtained from
Gibco BRL (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Aminophenyl fluorescein (APF), dihydroethidium (hydroethidine,
DHE), and H2DCFDA were obtained from Molecular Probes
(Thermo Fisher Scientific, Inc., Waltham, MA, USA). ROSstar™ 650
was purchased from LI-COR Biosciences (Lincoln, NE, USA). The
Apoptosis/Necrosis Detection Kit (blue, red, green; ab176750) was
obtained from Abcam (Cambridge, UK). The HeLa cell line was
purchased from the Cell Bank of the Committee of Type Culture
Collection (Seoul, Korea). The 96-well plates (clear/black) and
4-well cell culture slides were supplied by SPL Life Sciences
(Pocheon, Korea).
Carboplatin cytotoxicity test to
determine the non-toxic dose
To establish the low-dose response rate of HeLa
cells to carboplatin (CBP), cells were treated with 0–1,000 µM CBP
in standard media. HeLa cells were cultured at 37°C in a 5%
CO2 humidified atmosphere in RPMI-1640 medium
supplemented with 10% (v/v) fetal bovine serum (FBS) and 1% (v/v)
P/S. Four batches of cells for each CBP concentration were prepared
by seeding at a density of 1×105 cells/ml onto clear
96-well plates and incubated overnight. After changing to a medium
containing 0, 5, 20, 100 and 1,000 µM CBP, the cells were incubated
in the dark for 24 h, and the cytotoxicity was measured using the
MTT (3(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)
assay.
Concurrent Photofrin and CBP
treatment
HeLa cells at a density of 1×105 cells/ml
were seeded in 96-well plates (clear/black) or 4-well cell culture
slides, and incubated overnight. After washing with PBS twice, the
cells were incubated with 20 µM Photofrin-containing medium for 3
h, or/and washed twice with PBS, and re-incubation with 100 µM
CBP-containing medium. Medium was replaced with PBS prior to light
irradiation.
Assessment of ROS levels
Five wells of cells per group were seeded and
treated as described above with concurrent Photofrin and CBP
treatment in 44 black 96-well plates. After twice washing, 4 ROS
probe groups of each 11 plates were incubated with medium
containing 1 µM of APF, 5 µM of DHE, 25 µM of ROSstar™ 650, and 1
µM of H2DCFDA in the dark separately for 30 min. The
cells were then washed twice and placed in PBS, and light
irradiation was performed using a 630-nm laser with an intensity of
2.5 mW/cm2 for 0, 100, 200, 300, 400, 500, 600, 700,
800, 900 and 1,000 sec. The generation of each type of ROS was
measured by fluorescence intensity using a plate reader set at the
appropriate wavelength: APF, λex=465
nm/λem=510 nm; DHE, λex=510
nm/λem=590 nm; ROSstar™, λex=635
nm/λem=670 nm, and H2DCFDA,
λex=510 nm/λem=535 nm.
Apoptosis/necrosis assay
Cells prepared in 4-well cell culture slides were
treated according to the above concurrent Photofrin and CBP
treatment. Cells were treated by light irradiation at: λ=630 nm;
fluence rate, 2.5 mW/cm2; light dose, 660 mJ. After
treatment, the medium was replaced, and cells were incubated for 24
h. The apoptosis/necrosis assay was performed according to the
manufacturer's instructions. Cells were treated with a mixture of
staining solutions: 20 µl of Apopxin™ Deep Red Indicator (100X) for
apoptotic cells, 10 µl of Nuclear Green DCS1 (200X) for necrotic
cells, 10 µl of CytoCalcein Violet 450 (200X) for viable cells, and
2 ml of assay buffer. The cells were then incubated for 30 min at
room temperature, washed two times with assay buffer, and finally
analyzed by confocal microscopy with the following wavelengths:
Apopxin Deep Red Indicator; λex=640
nm/λem=663–738 nm; Nuclear Green DCS1,
λex=488 nm/λem=500–550 nm; and CytoCalcein
violet 450, λex=405 nm/λem=425–475 nm.
Cell viability assay
Each group contained 4 (wells) ×11 slide samples for
multiple irradiation doses. HeLa cells were prepared for each
experimental group by seeding at a density of 1×105
cells/ml in 96-well plates (clear/black) or 4-well chamber cell
culture slides, and incubated overnight. After washing twice with
PBS, the cells were incubated with 20 µM Photofrin-containing
medium for 3 h, washed twice with PBS and re-incubated in 100 µM
CBP-containing medium. Four different experimental groups were
prepared; cells incubated with no drugs (control), Photofrin (PF),
carboplatin (CBP), or Photofrin and carboplatin (PF+CBP; ccPDT).
The medium was replaced with PBS prior to light irradiation. Light
irradiation was performed with a fluence rate of 2.5
mW/cm2 at various light doses: 0, 330, 660, 990, 1,320,
1,650, 1,980, 2,310, 2,640, 2,970, 3,300 mJ for which the total
area irradiated was 1.32 cm2. Cell viability was
determined using the MTT assay 4 h after light irradiation.
Statistical analysis
ROS-fluorescence intensity and OD540 data
on the viability levels are presented as the mean ± standard
deviation (SD). One-way analysis of variance (ANOVA) was used for
data analyses. The Levene's test was used to demonstrate the equal
variances of the variables. Post hoc analysis using Bonferroni's
multiple comparison was used to determine significant differences.
All testing was performed using IBM SPSS statistical software v23
(IBM Corp., Armonk, NY, USA).
Results
Cytotoxicity of carboplatin- and
PF-mediated PDT
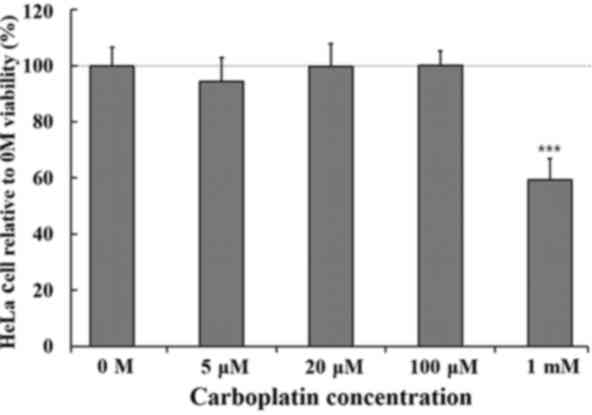
Significant CBP-mediated cytotoxicity was not
induced in HeLa cells treated with up to 100 µM CBP, as shown in
Fig. 1. When treated with 1 mM of
CBP, cell viability was decreased by 30%. Thereafter, 100 µM CBP
was used in further ROS measurement studies or cytotoxicity assays
for the combined treatment.
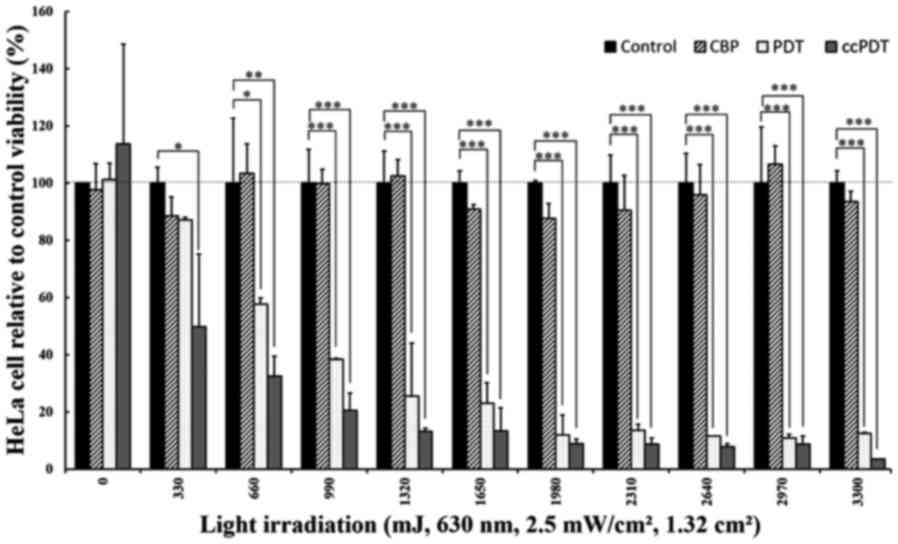
When the cells were treated with 100 µM CBP in
combination with varying light doses and 20 µM Photofrin (PF+CBP),
the percentage of cell viability was decreased significantly
(P<0.001) compared to each individual treatment alone (Fig. 2). When treated with 330 mJ of light,
Photofrin (PF)-PDT did not produce significant cytotoxicity, while
ccPDT (PF+CBP) treatment showed a 50.2±25.4 or 42.8±28.2% reduction
in viable cells compared to the control or PF-PDT alone,
respectively. The enhanced reduction in cell viability reached
67.5±6.9 or 43.7±3.1% compared to the control or PF-PDT alone,
respectively, when treated with the light dose of 660 mJ. The
relative cytotoxicity ratio of combined treatment to PF-PDT alone
gradually increased from 1.74 to 3.54 with a light dose ranging
from 330 to 3,300 mJ.
Increased apoptotic effect following
combined treatment
Triple staining with Apopxin Deep Red Indicator,
Nuclear Green DCS1, and CytoCalcein Violet 450 was performed to
determine the cellular damage pathway for individual and combined
treatments. As shown in Fig. 3, the
dominate pathway for PF-PDT alone was necrotic death, while
enhanced apoptosis was observed for ccPDT. Light irradiation in the
presence of CBP or PF alone demonstrated slight apoptosis, 6.0±2.1
or 3.3±1.8% in average for multiple sample batches, respectively,
based on counting cells in fluorescence images, but this apoptotic
effect increased notably (35.2±9.4%) following PDT treatment under
the coexistence of CBP and Photofrin (ccPDT).
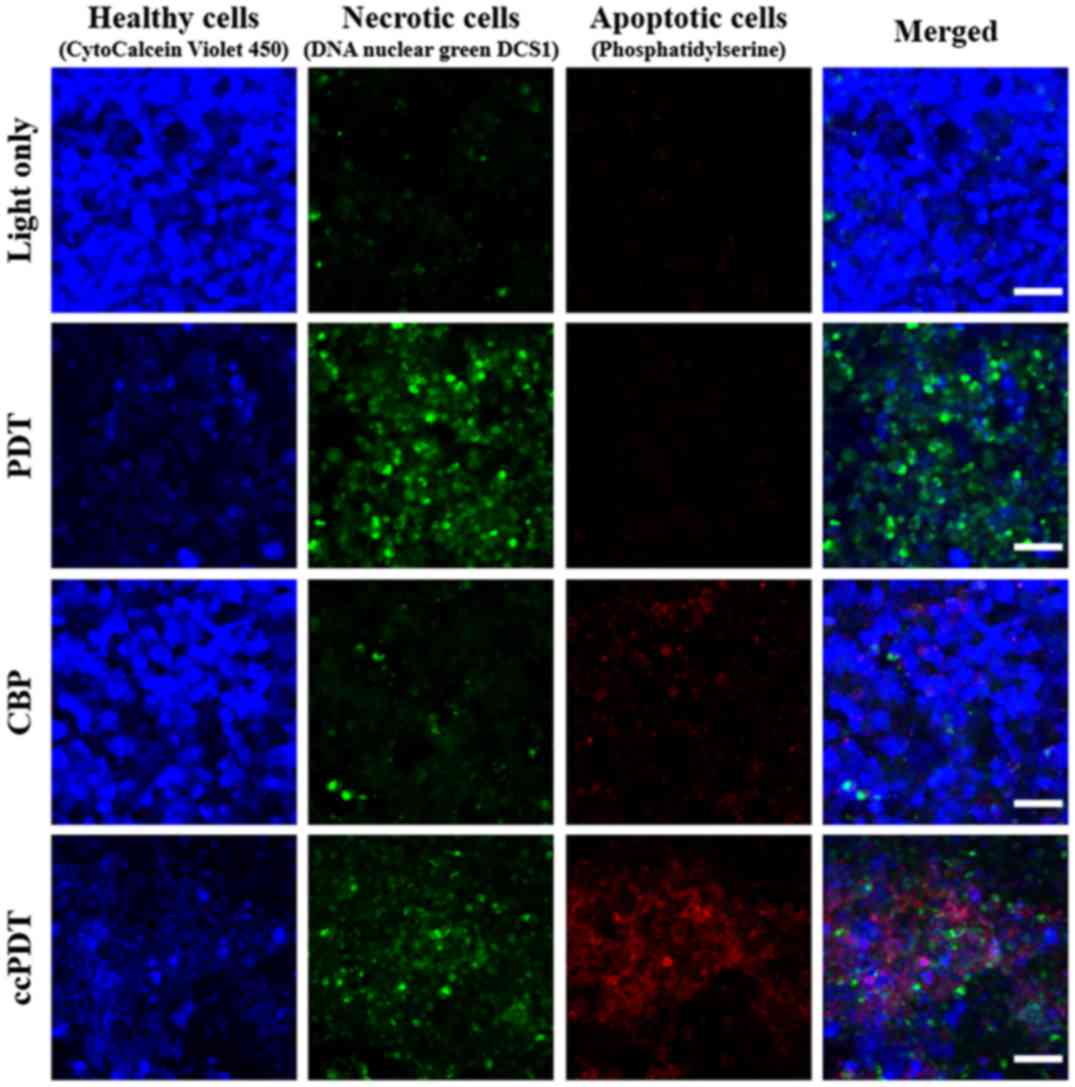 | Figure 3.Necrosis and apoptosis assays of HeLa
cells treated with light only, 20 µM Photofrin-PDT (PF-PDT), light
irradiation in the presence of 100 µM carboplatin only (CBP), and
ccPDT. Healthy viable cells were stained with CytoCalcein Violet
450 (blue), necrotic cells with DNA nuclear green DCS1 (green), and
apoptotic cells with phosphatidylserine (red). (Nikon A1+,
objective: CFI Plan Apo Lambda 10X, NA: 0.45, scale bar: 50 µm).
Apoptotic cell death was only 6.0±2.1% following CBP treatment,
while 7.3±3.5% cells exhibited necrotic death. In contrast, the
apoptotic effect was increased by 35.2±9.4% following ccPDT. |
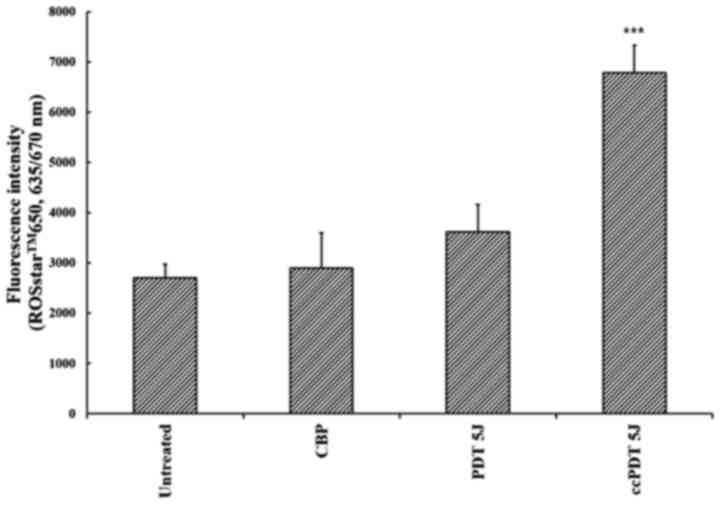
ROS production following ccPDT
treatment
In PF-PDT alone with a relatively high fluence rate
(35 mW/cm2) and light dose (6.6 J), ROS production was
increased by 33.7±19.9% compared with the untreated control, while
ccPDT demonstrated enhanced ROS production by a factor of 2.4 or
1.9 compared with the untreated or PF-PDT alone, respectively
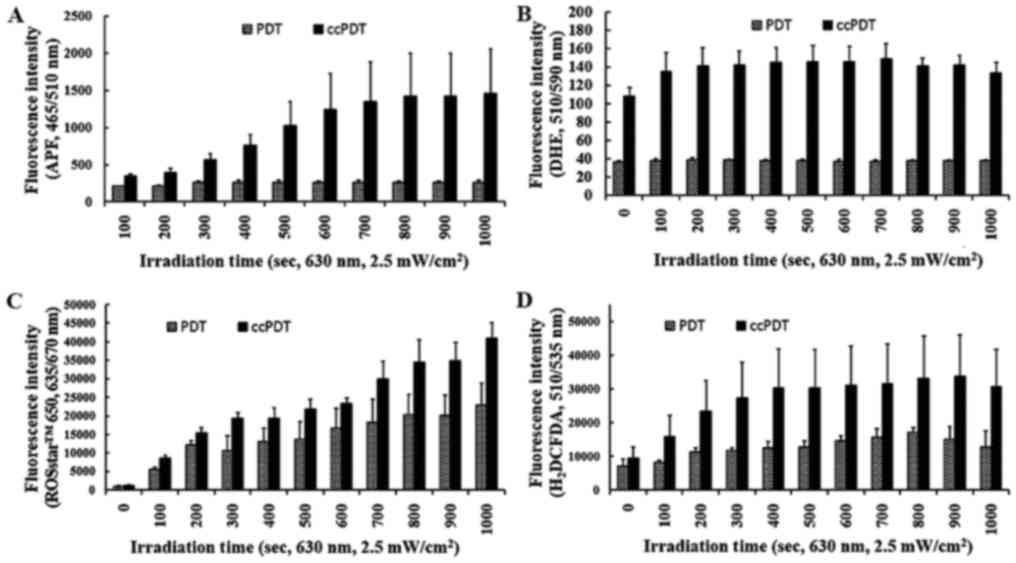
(Fig. 4). In PF-PDT alone under
irradiation with a relatively lower fluence rate (2.5
mW/cm2), yields of OH• and
O2▪– were residual and did not increase with
higher light doses, and were much smaller than those in ccPDT
(Fig. 5). However, ROS detection
based on ROSstar™ 650 increased with increasing light doses even in
the PF-PDT alone treatment group, indicating enhancement of
H2O2 with increasing light doses in the
PF-PDT alone group (Fig. 5C and D).
In contrast, ccPDT showed enhanced production of
O2▪– by a factor of 3 on average over the
range of irradiation doses and a gradual increment of the
OH• yield over the range of light doses. In the Type I
reaction of PDT, electron transfer to triplet oxygen can produce
O2▪–. In ccPDT, the residual
O2▪– in PDT alone increased sharply, but
saturated quickly with increasing light doses. In contrast, the
yield of OH• in the combined treatment group was highly
enhanced compared to PDT alone, and this enhancement increased with
higher light doses, but remained residual in PDT alone. The 50%
enhanced production of H2O2 by the
dismutase-mediated conversion of O2▪– was
observed to increase gradually with increasing light doses in the
combined treatment group (ccPDT) (Fig.
5D).
Discussion
Concurrent ccPDT was found to produce hydroxyl
radicals as the major additional ROS compared to the prevalent
singlet oxygen following PF-PDT alone. The most common oxidation
states of platinum are +2 (d8) and +4 (d6)
(8). Cisplatin and carboplatin are
typical PtII drugs when the preferred oxidation state of
the Pt center is +2. CBP-stimulated production of
O2▪– can be converted to
H2O2 by cellular dismutase (14–16),
which can be further transformed into OH· by means of
either the CBP-mediated Fenton-like reaction via a light-mediated
redox active platinum ion (17–19) or
the Fe+2-mediated Fenton reaction from oxidized [4Fe-4S]
clusters of proteins (20,21) under the combined regime, as shown in
the following equation:
2[PtII]2+H2O2➔[Pt2.25]4+OH-+•OH
This synergistic reaction may result in enhanced
reactive conversion of CBP in the presence of PDT-mediated oxygen
radicals under the ccPDT regime (18,19).
The notable increase in the OH• yield and relatively
constant O2▪– yield with a light dose
supports the hypothesis for a Fenton-like reaction via
photoactivated CBP. On the other hand, O2▪–
contributes to the iron release by oxidizing the [4Fe-4S] clusters
(22), which acts as a catalyst for
H2O2 decomposition in the Fenton reaction,
producing more OH• (20,21).
In normal untreated cells, H2O2 as a
metabolic byproduct can then be removed through enzymatic reactions
(e.g., catalase) and thiol-systems (e.g. glutathione) to avoid the
Fenton reaction formation of OH•. In PF-PDT-treated
cells, cellular removal of H2O2 may be
partially impaired as a result of damage to the removal pathway or
because the Fenton reaction-mediated conversion to OH·
is limited by a much lower electron transfer rate (k ≤
1×107 M−1sec−1) compared to a
higher rate of energy transfer (k≤1-3×109
M−1sec−1) (23). These results led to the residual
accumulation of dose-dependent intracellular
H2O2 (Fig.
5D), and lipid peroxidation contributing to necrotic damage
together with the primary damaging species,
1O2. In the presence of CBP under ccPDT, the
PtII oxidation status enables CBP to easily react with
soft bases such as sulfur-containing glutathione (GSH) and other
cysteine-rich molecules (8).
CBP-stimulated production of O2▪– may lead to
the decrease in protective antioxidant systems, and effectively
sensitizing the cell to oxidative stress via GSH and NADPH
depletion (6,16,24).
The generation of OH▪ can result in a cascade of
different ROS, each with unique properties and preferred biological
targets. Necrotic death was a main pathway of cellular damage by
Photofrin-mediated PDT under the present regime, as observed in
Fig. 3, whereas the apoptotic
effects were superimposed in ccPDT as shown also in another report
(7). However, in general, the
pathway for PF-PDT-mediated cell damage depended on the
concentration of PF and the light dose as revealed in previous
research (25). In the present
study we aimed to demonstrate enhanced production of hydroxyl
radicals and related apoptotic effects under only current ccPDT
(concurrent low-dose CBP and conventional PF-PDT) regime. General
dependency of cellular death on the PF-concentration was not
pursued in this study. This enhanced apoptosis can be initiated by
the CBP-induced mitochondrial-ROS response (14–16)
with depletion of the antioxidant enzyme; however, apoptosis did
not occur significantly under CBP alone, as shown in the individual
CBP only group (Fig. 3). This
result was largely enhanced by accumulated OH• (12,13),
suggesting a synergistic effect by ccPDT. A similar effect was
reported following cisplatin treatment showing NADPH depletion,
which resulted in an altered mitochondrial redox status that then
caused the generation of OH▪ and induction of apoptosis (26–28).
Overall, determining the outcome of PDT on a cellular level is
complex. Nevertheless, some general themes can be observed
(29). With high doses of PDT or PS
localization to the plasma membrane, necrosis is the dominant form
of cell death. With mild PDT and damage to the mitochondria or
anti-apoptotic components, apoptosis is triggered. With low
PDT-induced damage to organelles, autophagy is initiated to repair
the damage (30). Therefore, in
PF-PDT with conventional light dose where PF is localized mainly to
the plasma membrane, autophagy may not be the major mechanism of
cellular damage in this ccPDT despite application of variable light
dose.
ROS-mediated therapeutic efficacy is depicted in
Fig. 6. In summary, these results
suggest that in Photofrin-PDT alone, necrotic damage is obtained by
a 1O2-mediated Type II reaction, while
O2▪– is converted into
H2O2, which is not removed efficiently due to
partially impaired antioxidant systems under PDT. Conversely, in
ccPDT, production of O2▪– is enhanced by CBP,
and H2O2 can be further converted into more
toxic OH· via both the potent CBP-mediated Fenton-like
reaction and enhancement of the oxidized [4Fe-4S]-mediated Fenton
reaction. Therefore, the therapeutic enhancement in low-dose
CBP-based ccPDT may be due to the exploitation of the synergistic
enhancement of OH· that led to superimposed
apoptosis-based cellular death, while avoiding side effects by
reducing the effective dosage of carboplatin.
Acknowledgements
The authors thank Ahn Jin-Cheol of the Medical Laser
Research Center, Dankook University for the laser equipment.
Funding
The present study was supported in part by a grant
from the Catholic University of Daegu (no. 20175003) and the
Industrial Materials Fundamental Technology Development Program
(no. 10062194).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
CYS and CJE performed experiment. CYS and KJK
designed the study. JSH reviewed and edited the manuscript. KJK
wrote the manuscript. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors report no conflicts of interest.
References
|
1
|
Ahn TG, Lee BR, Kim JK, Choi BC and Han
SJ: Successful full term pregnancy and delivery after concurrent
chemo-photodynamic therapy (CCPDT) for the uterine cervical cancer
staged 1B1 and 1B2: Preserving 4 fertility in young women. Gynecol
Oncol Case Rep. 2:54–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rizvi I, Celli JP, Evans CL, Abu-Yousif
AO, Muzikansky A, Pogue BW, Finkelstein D and Hasan T: Synergistic
enhancement of carboplatin efficacy with photodynamic therapy in a
three-dimensional model for micrometastatic ovarian cancer. Cancer
Res. 70:9319–9328. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Biswas R, Mondal A and Ahn JC:
Deregulation of EGFR/PI3K and activation of PTEN by photodynamic
therapy combined with carboplatin in human anaplastic thyroid
cancer cells and xenograft tumors in nude mice. J Photochem
Photobiol B. 148:118–127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mao W, Sun Y, Zhang H, Cao L, Wang J and
He P: A combined modality of carboplatin and photodynamic therapy
suppresses epithelial-mesenchymal transition and matrix
metalloproteinase-2 (MMP-2)/MMP-9 expression in HEp-2 human
laryngeal cancer cells via ROS-mediated inhibition of MEK/ERK
signalling pathway. Lasers Med Sci. 31:1697–1705. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hwang H, Biswas R, Chung PS and Ahn JC:
Modulation of EGFR and ROS induced cytochrome c release by
combination of photodynamic therapy and carboplatin in human
cultured head and neck cancer cells and tumor xenograft in nude
mice. J Photochem Photobiol B. 128:70–77. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Peterson K, Harsh G IV, Fisher PG, Adler J
and Le Q: Daily Low-dose carboplatin as a radiation sensitizer for
newly diagnosed malignant glioma. J Neurooncol. 53:27–32. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
He P, Ahn JC, Shin JI, Hwang HJ, Kang JW,
Lee SJ and Chung PS: Enhanced apoptotic effect of combined modality
of 9-hydroxypheophorbide alpha-mediated photodynamic therapy and
carboplatin on AMC-HN-3 human head and neck cancer cells. Oncol
Rep. 21:329–334. 2009.PubMed/NCBI
|
|
8
|
Jungwirth U, Kowol CR, Keppler BK,
Hartinger CG, Berger W and Heffeter P: Anticancer activity of metal
complexes: Involvement of redox processes. Antioxid Redox Signal.
15:1085–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Agostinis P, Berg K, Cengel KA, Foster TH,
Girotti AW, Gollnick SO, Hahn SM, Hamblin MR, Juzeniene A, Kessel
D, et al: Photodynamic therapy of cancer: An update. CA Cancer J
Clin. 61:250–281. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ogilby PR: Singlet oxygen: There is indeed
something new under the sun. Chem Soc Rev. 39:3181–3209. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Buytaert E, Dewaele M and Agostinis P:
Molecular effectors of multiple cell death pathways initiated by
photodynamic therapy. Biochim Biophys Acta. 1776:86–107.
2007.PubMed/NCBI
|
|
12
|
Matroule JY, Carthy CM, Granville DJ,
Jolois O, Hunt DW and Piette J: Mechanism of colon cancer cell
apoptosis mediated by pyropheophorbide-A methylester
photosensitization. Oncogene. 20:4070–4084. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li D, Li L, Li P, Li Y and Chen X:
Apoptosis of HeLa cells induced by a new targeting
photosensitizer-based PDT via a mitochondrial pathway and ER
stress. Onco Targets The. 8:703–711. 2015. View Article : Google Scholar
|
|
14
|
Husain K, Jagannathan R, Hasan Z, Trammell
GL, Rybak LP, Hazelrigg SR and Somani SM: Dose response of
carboplatin-induced nephrotoxicity in rats. Pharmacol Toxicol.
91:83–89. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Husain K, Whitworth C, Somani SM and Rybak
LP: Carboplatin-induced oxidative stress in rat cochlea. Hear Res.
159:14–22. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Reddy Kishore YV, Reddy Sreenivasula P and
Shivalingam MR: Cisplatin or carboplatin caused suppression in
anti-oxidant enzyme defense system in liver, kidney and testis of
male albino rats. J Biomed Sci and Res. 2:23–28. 2010.
|
|
17
|
Bernhard L: Cisplatin: Chemistry and
biochemistry of leading anticancer drug. Wiley-VCH; 1st edition.
Zurich, Switzerland: pp. 4641999
|
|
18
|
Tonetti M, Giovine M, Gasparini A, Benatti
U and De Flora A: Enhanced formation of reactive species from
cis-diammine-(1,1-cyclobutanedicarboxylato)-platinum(II)
(carboplatin) in the presence of oxygen free radicals. Biochem
Pharmacol. 46:1377–1383. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deavall DG, Martin EA, Horner JM and
Roberts R: Drug-induced oxidative stress and toxicity. J Toxicol.
2012:6454602012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim JH, Bothe JR, Alderson TR and Markley
JL: Tangled web of interactions among proteins involved in
iron-sulfur cluster assembly as unraveled by NMR, SAXS, chemical
crosslinking, and functional studies. Biochim Biophys Acta.
1853:1416–1428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tracey Rouault: Iron-Sulfur Clusters in
Chemistry and Biology. Walter de Gruyter GmbH & Co KG.
3002014.
|
|
22
|
Benov L: How superoxide radical damages
the cell. Protoplasma. 217:33–36. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Davies MJ: Reactive species formed on
proteins exposed to singlet oxygen. Photochem Photobiol Sci.
3:17–25. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Afanas'ev IB: Superoxide Ion: Chemistry
and Biological Implications Vol II Oxygen radicals in Biology. CRC
Press; pp. 541991
|
|
25
|
Gao S, Zhang M, Zhu X, Qu Z, Shan T, Xie
X, Wang Y and Feng X: Apoptotic effects of Photofrin-Diomed 630-PDT
on SHEEC human esophageal squamous cancer cells. Int J Clin Exp
Med. 8:15098–15107. 2015.PubMed/NCBI
|
|
26
|
Marullo R, Werner E, Degtyareva N, Moore
B, Altaville G, Ramalingam SS and Doetsch PW: Cisplatin induces a
mitochondrial-ROS response that contributes to cytotoxicity
depending on mitochondrial redox status and bioenergetics
fucntions. PLoS One. 8:E811622013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martins NM, Santos NA, Curti C, Bianchi ML
and Santos AC: Cisplatin induces mitochondrial oxidative stress
with resultant energetic metabolism impairment, membrane
rigidification and apoptosis in rat liver. J Appl Toxicol.
28:337–344. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mandic A, Hansson J, Linder S and Shoshan
MC: Cisplatin induces endoplasmic reticulum stress and
nucleus-independent apoptotic signaling. J Biol Chem.
278:9100–9106. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mroz P, Yaroslavsky A, Kharkwal GB and
Hamblin MR: Cell death pathways in photodynamic therapy of cancer.
Cancers. 3:2516–2539. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
van Straten D, Mashayekhi V, de Bruijn HS,
Oliveira S and Robinson DJ: Oncologic photodynamic therapy: Basic
principles, current clinical status and future Directions. Cancers.
9:pii: E19. 2017. View Article : Google Scholar : PubMed/NCBI
|