Introduction
The incidence of colorectal cancer (CRC) is
increasing, and CRC currently represents a major cause of
cancer-related morbidity and mortality worldwide, with high
incidence rates in Westernized societies and increasing rates in
developing countries (1,2). As the majority of the patients present
with advanced disease, such as the presence of liver metastases, at
the time of diagnosis, the scope of therapeutic intervention is
significantly limited (3). Various
factors have been confirmed to participate in this progression,
such as the silencing of tumor-suppressor genes, the
hyperactivation or overexpression of proto-oncogenes, and the
dysregulation of genes that are associated with cell growth,
apoptosis or transformation (4–6).
MicroRNAs (miRNAs/miRs) are a recently characterized
class of small non-coding RNA molecules of 20–22 nucleotides.
Mature miRNAs can specifically bind to the 3′-untranslated region
(3′-UTR) of target cell mRNAs, resulting in mRNA degradation or the
inhibition of translation. Post-transcriptional regulation of gene
expression by miRNAs is an important characteristic of the cell
differentiation process, and it has been predicted that there are
numerous as yet undiscovered miRNAs in the genome of humans and
other higher vertebrates (7,8). In
recent decades, studies have validated that miRNAs are key
regulators of diverse cellular processes, including apoptosis,
proliferation, differentiation, metabolism and immunity (9–11).
miR-1 is a muscle-enriched miRNA that inhibits the
proliferation of progenitor cells and promotes myogenesis (12,13).
The downregulated expression of miR-1 has also been identified in
lung, liver, breast, prostate and kidney cancer. The restoration of
miR-1 expression in cancer cell lines was found to markedly
reversed their tumorigenic properties, such as growth, clone
formation, migration, invasion and tumor formation ability in nude
mice (14,15). The decreased expression of miR-1 has
been suggested to be associated with liver metastasis, whereas its
function and the underlying mechanism in colon cancer require
further investigation (16,17). Vascular endothelial growth factor
(VEGF)-A and VEGF-C/D are major factors affecting angiogenesis
and/or lymphangiogenesis (18).
Angiogenesis plays a crucial role in prenatal development, wound
healing, chronic inflammation, angiogenesis and lymphangiogenesis,
and promotes the metastasis and progression of various carcinomas
(19–21). The ectopic expression of VEGF has
been demonstrated to be closely associated with cell proliferation,
invasion and the metastatic potential of colon cancer cells, which
contributes to cancer progression, whereas anti-VEGF-based
antiangiogenic drugs, including bevacizumab, aflibercept,
ramucirumab and tyrosine kinase inhibitors, are routinely used for
the treatment of various types of tumor (22).
In the present study, we examined the expression of
miR-1 in CRC tissues and cell lines using immunohistochemistry and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analyses. The effect of miR-1 on cell growth, apoptosis,
migration and invasion were assessed by several assays.
Furthermore, VEGF was predicted to be a target protein of miR-1 by
bioinformatic analysis, and a negative association of miR-1 and
VEGF expression was observed, which suggests that miR-1
downregulates the expression of VEGF. It is well established that
high expression of VEGF promotes biological behaviors such as cell
proliferation, invasion and metastasis in colon cancer, leading to
cancer progression. Collectively, this study demonstrated that
decreased expression of miR-1 may cause increased expression of
VEGF, which sets a theoretical and practical basis for miR-1 as a
biological marker for CRC diagnosis and targeted therapy.
Materials and methods
Patients and cell lines
A total of 111 samples of cancer and paracancerous
tissues were collected, cut into small sections, immediately frozen
in liquid nitrogen and stored at −80°C. The patients included 45
men and 66 women (median age, 61.3 years; range, 27–85 years). All
tissues were obtained from patients who underwent surgery for CRC
between 2007 and 2012 at the First Hospital of China Medical
University (Shenyang, China). All diagnoses were histologically
confirmed. The study was approved by the Ethics Committee of China
Medical University (Shenyang, China), and written informed consent
was obtained prior to sample collection. In addition, the human
peritoneal mesothelial cell line HMrSV5, and colorectal cancer cell
lines, including HCT-116, CL-187, ClonA1, HT-29 and SW-620, were
cultured in HyClone RPMI-1640 medium (GE Healthcare Life Sciences,
Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS) and
maintained at 37°C in 5% CO2. We failed to identify a
normal colon epithelial cell line with cell identification.
Therefore, the human peritoneal mesothelial cell line (HMrSV5) was
used as a control. The HMrSV5 cell line was derived from normal
peritoneal cells and it did not display malignant biological
behavior. The HCT-116, SW-480, SW-620 and HT-29 cell lines were
obtained from the Cell Repository of the Typical Culture
Preservation Committee of the Chinese Academy of Sciences/The Cell
Resource Center of the Shanghai Academy of Life Sciences, Chinese
Academy of Sciences. The ClonA1, CL-187 and HMrSV5 cell lines were
gifts from the Department of Cell Biology, China Medical University
(Shenyang, China).
RNA extraction and RT-qPCR
Total RNA was isolated from tumor tissues or cells
with Invitrogen™ TRIzol reagent (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), and the miRNAs were then isolated using the
Ambion® mirVana miRNA Isolation kit (Thermo Fisher
Scientific, Inc.). In order to detect the expression of miR-1,
stem-loop RT-qPCR was performed. The primers used in this study
were as follows: RT primer for miR-1,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGATACATAC-3′; RT primer for
U6 snRNA: 5′-AACGCTTCACGAATTTGCGT-3′; miR-1 forward primer,
5′-ACACTCCAGCTGGGTGGAATGTAAAGAAGT-3′; miR-1 reverse primer,
5′-TGGTGTCGTGGAGTCG-3′; U6 forward primer, 5′-CTCGCTTCGGCAGCACA-3′,
U6 reverse primer, 5′-AACGCTTCACGAATTTGCGT-3′. The expression of
VEGF was normalized to GAPDH, and the primers used in RT-qPCR were
as follows: VEGF forward primer, 5′-CAAGGCCAGCACATAGGAGAG3-3′, VEGF
reverse primer, 5′-CCTCGGCTTGTCACATCTTGC-3′; GAPDH forward primer,
5′-GACTGTGGATGGCCCCTCCGG-3′, GAPDH reverse primer,
5′-AGGTGGAGGAGTGGGTGTCGC-3′. The RT-qPCR procedure in this study
was performed as previously described (23).
Cell transfection
miR-1-overexpressing and control cell lines were
constructed by infecting the cells with lentiviral miR-1 and
control vector (Shanghai Genechem Co., Ltd., Shanghai, China),
according to the manufacturer's instructions. The expression of
miR-1 in the stable cell lines was determined with RT-qPCR
analysis.
ELISA
miR-1-overexpressing or control cells
(1×105) were seeded in 6-well plates. After 36 h, the
supernatants were collected and the levels of VEGF were assessed
with a human VEGF ELISA kit (R&D Systems, Inc., Minneapolis,
MN, USA) according to the manufacturer's instructions.
Western blotting
Harvested cells were lysed in RIPA lysis buffer
containing protease inhibitor. Total cellular protein (30 mg/lane)
was separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred to a polyvinylidene
difluoride membrane (EMD Millipore, Billerica, MA, USA). The
membranes were blocked with 5% non-fat milk and incubated with
rabbit anti-VEGF antibody at a dilution of 1:1,000 (cat. no.
ab155944; Abcam, Cambridge, MA, USA), or a rabbit anti-GAPDH
monoclonal antibody at a dilution of 1:3,000 (cat. no. ab181602;
Abcam), or a rabbit anti-VEGF receptor 2 (VEGFR-2) antibody at a
dilution of 1:500 (cat. no. 26415-1-AP; ProteinTech Group, Inc.,
Chicago, IL, USA). A secondary antibody was then incubated with the
membrane for 2 h. Bound proteins were visualized using
electrochemiluminescence (Pierce; Thermo Fisher Scientific, Inc.)
and detected with a bio-imaging system (DNR Bio-Imaging Systems,
Israel).
Immunohistochemistry
Paraffin-embedded sections (4-µm) were
deparaffinized in three xylene washes, and rehydrated through a
graded alcohol series. Following antigen retrieval with 10 mM
sodium citrate buffer, the sections were blocked with goat serum
for 1 h at room temperature. Then sections were incubated with a
VEGF antibody (Abcam) overnight at 4°C or a rabbit anti-VEGFR-2
antibody (1:500) (Proteintech), and the expression of VEGF and
VEGFR-2 were examined with the UltraSensitive™ SP kit (Maixin-Bio,
Fuzhou, Fujian, China) according to the manufacturer's
instructions.
Migration and invasion assays
Cell migration was assessed using 24-well 8-µm-pore
Transwell chambers (Corning Costar, Corning, NY, USA) according to
the manufacturer's instructions. For the invasion assay, the insert
membranes were coated with diluted BD Matrigel™ (1:20) (BD
Biosciences, Franklin Lakes, NJ, USA). A total of 2×105
cells in 100-µl serum-free medium were added to the upper chamber,
and 600 µl 10% FBS medium was added to the corresponding lower
chamber. After 24 h of incubation, the cells were fixed with
methanol and stained with 0.1% crystal violet. Non-invading cells
in the upper surface of the chamber were removed with a cotton
swab, and invading cells on the lower membrane surface were
photographed with an inverted fluorescence microscope (Olympus
DP80; Olympus Corp., Tokyo, Japan) and counted. Six random fields
at a magnification of ×100 for each insert were counted.
Measurements were conducted in triplicate in three separate
experiments. For the migration assay, the procedures were similar,
except that 2×105 cells were added to the inserts
without Matrigel pre-coating.
Wound scratch assay
miR-1-overexpressing or control cells
(1×104) were seeded in 24-well plates. After cells had
grown to a confluence of 80–90%, a scratch was produced in the cell
monolayer using a 200-µl pipette. The medium was discarded and
cells were rinsed with phosphate-buffered saline (PBS) three times
to remove the cell debris, followed by the addition of fresh
culture medium. Wound areas were marked and photographed at
different time points (0, 24 and 48 h) using a phase-contrast
microscope and an Olympus DP74 color camera (Olympus Corp).
CCK-8 assay and cell growth curve
miR-1-overexpressing or control cells
(5×103) were seeded in 96-well plates, and the effect of
miR-1 on cell proliferation was measured by the colorimetric
water-soluble tetrazolium salt (WST) assay from a CCK-8 kit
(Beyotime Institute of Biotechnology, Haimen, China). The
absorbance at 450 nm was measured with a microplate reader. The
extent of proliferation was evaluated every 24 h for 5 days.
Cell cycle distribution analysis
miR-1-overexpressing or control cells
(1×105) were seeded in 6-well plates. After 36 h, the
cells were harvested by trypsinization, collected and washed with
PBS, and fixed with ethanol for 1 h at 4°C. After washing with PBS
three times, the cells were resuspended in 0.2 ml RNase A buffer (1
mg/ml) at 37°C for 30 min and stained with 0.3 ml propidium iodide
(PI) buffer (50 µl/ml). The stained cells were then analyzed for
DNA content by FACS flow cytometry.
Assessment of apoptosis by Annexin
V-FITC
miR-1- overexpressing or control cells
(1×105) were seeded in 6-well plates. After 36 h, the
cells were digested with trypsin and collected by centrifugation.
The cells were washed with cold PBS three times, resuspended in 100
µl binding buffer, and stained with 5 µl FITC-conjugated Annexin V
(10 mg/ml) and 10 µl PI (50 mg/ml) in the dark for 20 min at room
temperature. Then, 300 µl binding buffer was added and apoptotic
cells were analyzed with FACS flow cytometry.
Statistical analysis
The data from three independent experiments are
expressed as the mean ± standard deviation and were processed using
SPSS 17.0 statistical software (SPSS, Inc., Chicago, IL, USA). The
expression levels of miR-1 in CRC and paired adjacent normal tissue
samples were compared by Wilcoxon's paired test. The Student's
t-test was used to evaluate the association between the RT-qPCR
results for miR-1 expression and clinicopathological factors as
documented in Table I. Chi-square
tests were used to determine the association between the expression
of miR-1 and VEGF in primary CRC as shown in Table II. A P-value of <0.05 was
considered to indicate a statistically significant difference.
 | Table I.Relationship between the
clinicopathological parameters and miR-1 expression in primary CRC
cases (N=111). |
Table I.
Relationship between the
clinicopathological parameters and miR-1 expression in primary CRC
cases (N=111).
|
|
| Expression of
miR-1a |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | No. of cases | Mean | 95% CI | P-value |
|---|
| Age (years) |
|
<60 | 48 | −2.441 | −3.42–1.65 | 0.076 |
|
≥60 | 63 | −0.903 |
|
|
| Sex |
|
Male | 45 | −1.303 | −1.09–2.39 | 0.458 |
|
Female | 66 | −1.956 |
|
|
| Tumor size
(cm) |
|
<5 | 78 | −2.594 | −5.21–1.70 | 0.001 |
| ≥5 | 33 | 0.858 |
|
|
|
Differentiation |
| Well +
moderate | 33 | 0.094 | 0.55–4.18 | 0.011 |
|
Poor | 78 | −2.271 |
|
|
| Lymph node
status |
|
Negative | 63 | −3.082 | −5.01–1.91 | 0.001 |
|
Positive | 48 | 0.419 |
|
|
| Metastasis |
|
Negative | 95 | −1.543 | −2.26–2.61 | 0.888 |
|
Positive | 16 | −1.717 |
|
|
| TNM stage |
| I +
II | 55 | −3.122 | −4.69–1.47 | 0.001 |
| III +
IV | 56 | −0.042 |
|
|
| Table II.Association between the expression of
miR-1 and VEGF in the primary CRC cases (N=111). |
Table II.
Association between the expression of
miR-1 and VEGF in the primary CRC cases (N=111).
|
| Expression of
miR-1 |
|
|---|
|
|
|
|
|---|
|
VEGF | High (n=28) | Low (n=83) | P-value |
|---|
|
Positive | 5 | 72 | <0.001 |
|
Negative | 23 | 9 |
|
Results
miR-1 expression in colon cancer is
low
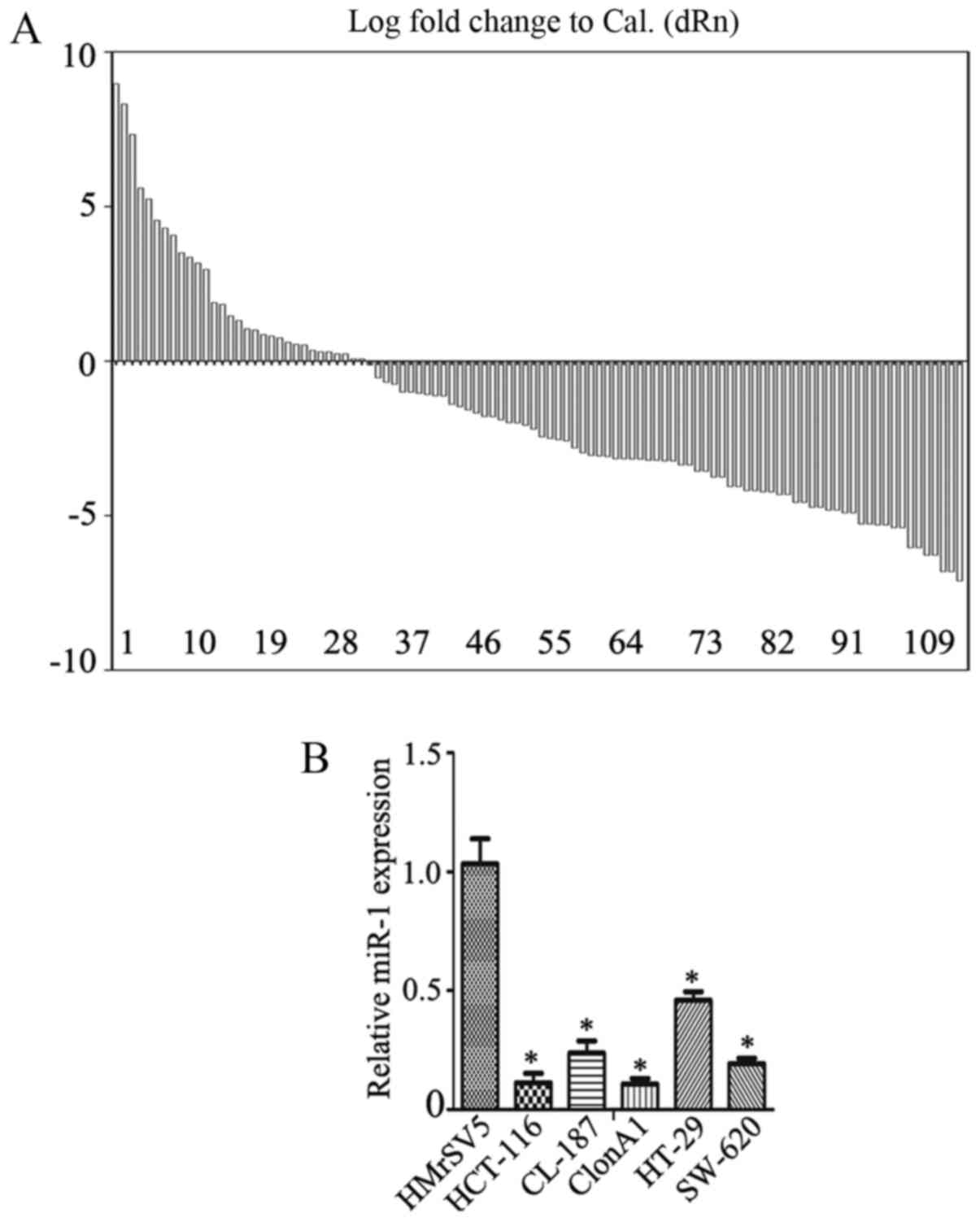
First, the expression of miR-1 in colorectal cancer
tissues and their pair-matched adjacent non-tumor tissues was
examined using RT-qPCR. The data revealed a decrease in miR-1
expression in 70.3% of cases, with a median change of ~1.57-fold
(Fig. 1A; P<0.01), a fold change
relative to the adjacent normal tissue. miR-1 expression was also
measured in several colorectal cancer cell lines, including
HCT-116, CL-187, ClonA1, HT-29 and SW-620, using RT-qPCR. The data
revealed that the expression of miR-1 was significantly
downregulated in HCT-116 (0.07±0.02-fold), CL-187 (0.41±0.03-fold),
ClonA1 (0.04±0.01-fold), HT-29 (0.60±0.01-fold) and SW-620
(0.38±0.04-fold) cells compared with the human peritoneal
mesothelial cell line HMrSV5 (Fig.
1B), one-way ANOVA was used to analyze data in Fig. 1B.
The miR-1 expression and the clinicopathological
characteristics of the CRC specimens are summarized in Table I. The expression of miR-1 was
significantly correlated with tumor size, degree of
differentiation, lymph node metastasis and TNM stage (P=0.001,
P=0.011, P=0.001 and P=0.001, respectively). There were no
significant correlations with the other clinicopathological
characteristics considered, including age and gender. These results
suggest that miR-1 may act as a tumor suppressor in CRC.
Ectopic expression of miR-1 inhibits
the growth of CRC cells
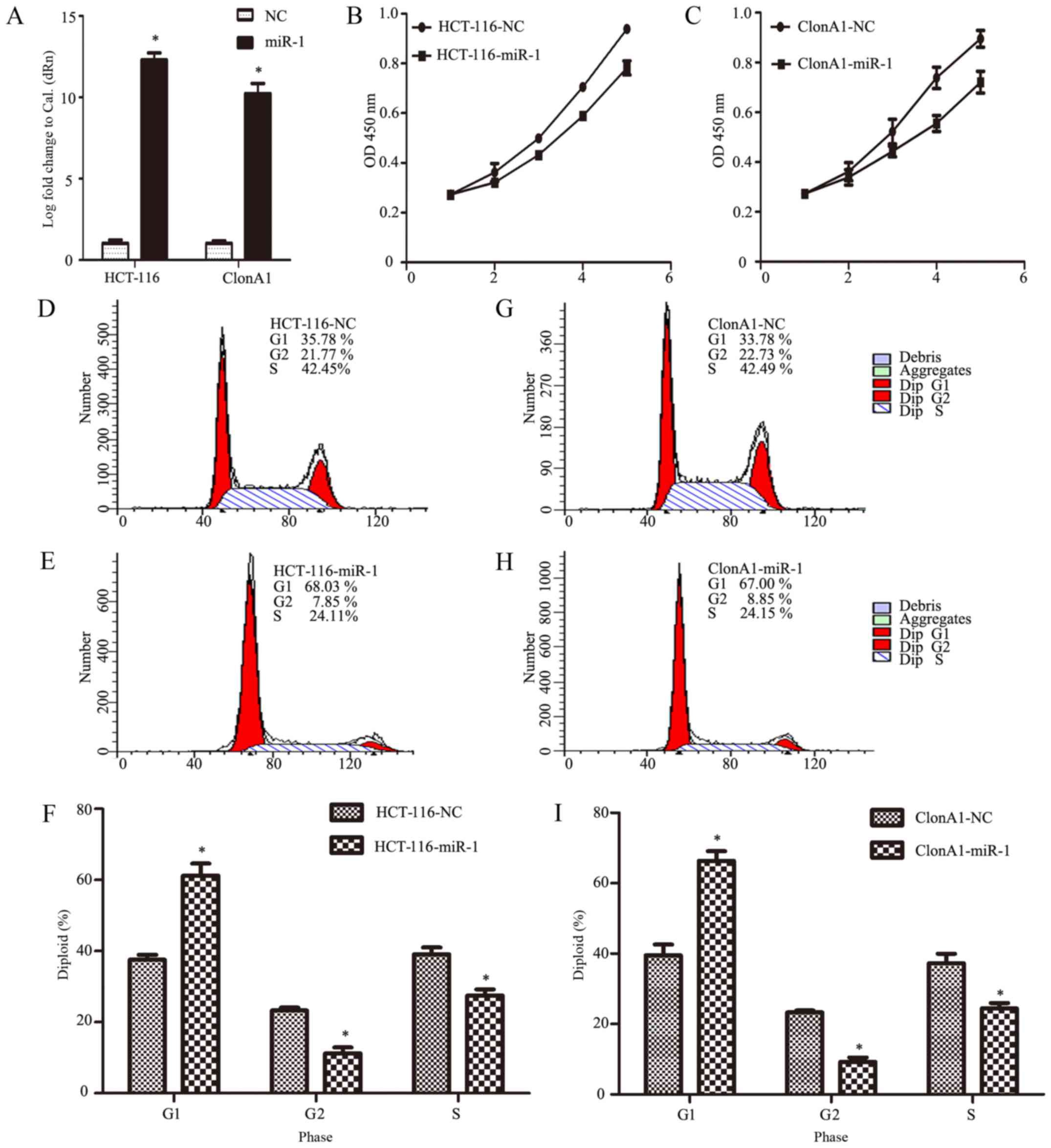
To investigate the biological function of miR-1 in
the development and progression of CRC, HCT-116 and ClonA1 cells
were infected with an miR-1 lentivirus to generate
miR-1-overexpressing cells. Fig. 2A
shows that the expression of miR-1 was effectively altered,
achieving an increase of >10,000-fold (P<0.0001, n=3,
Fig. 2A).
To determine the role of miR-1 in CRC, we
investigated the effect of miR-1 on the proliferation of cells with
CCK-8 and cell count assays at different time points in
vitro. The results demonstrated that the growth of both HCT-116
and ClonA1 miR-1-overexpressing cells was markedly impaired
compared with the negative control cells (Fig. 2B and C). To further confirm the
reason for this change, we assessed the cell cycle phase
distribution with flow cytometric analysis. It was observed that,
when ectopically elevating miR-1 expression in HCT-116 and ClonA1
cells, the transition from the G1 to the S phase was significantly
inhibited. As shown in Fig. 2D-I,
27.51 and 24.41% of miR-1-overexpressing HCT-116 and ClonA1 cells,
respectively, were in the S phase, whereas 39.12 and 37.23% of the
HCT-116 and ClonA1 control cells, respectively, were in the S
phase. Moreover, the cell population in the G1 phase of the cell
cycle was markedly larger in the miR-1-overexpressing cells (61.28
and 66.37%, respectively) compared with that in the negative
control cells (37.58 and 39.48%, respectively). Therefore, these
results indicate that miR-1 arrested the cells at the G1
phase of the cell cycle.
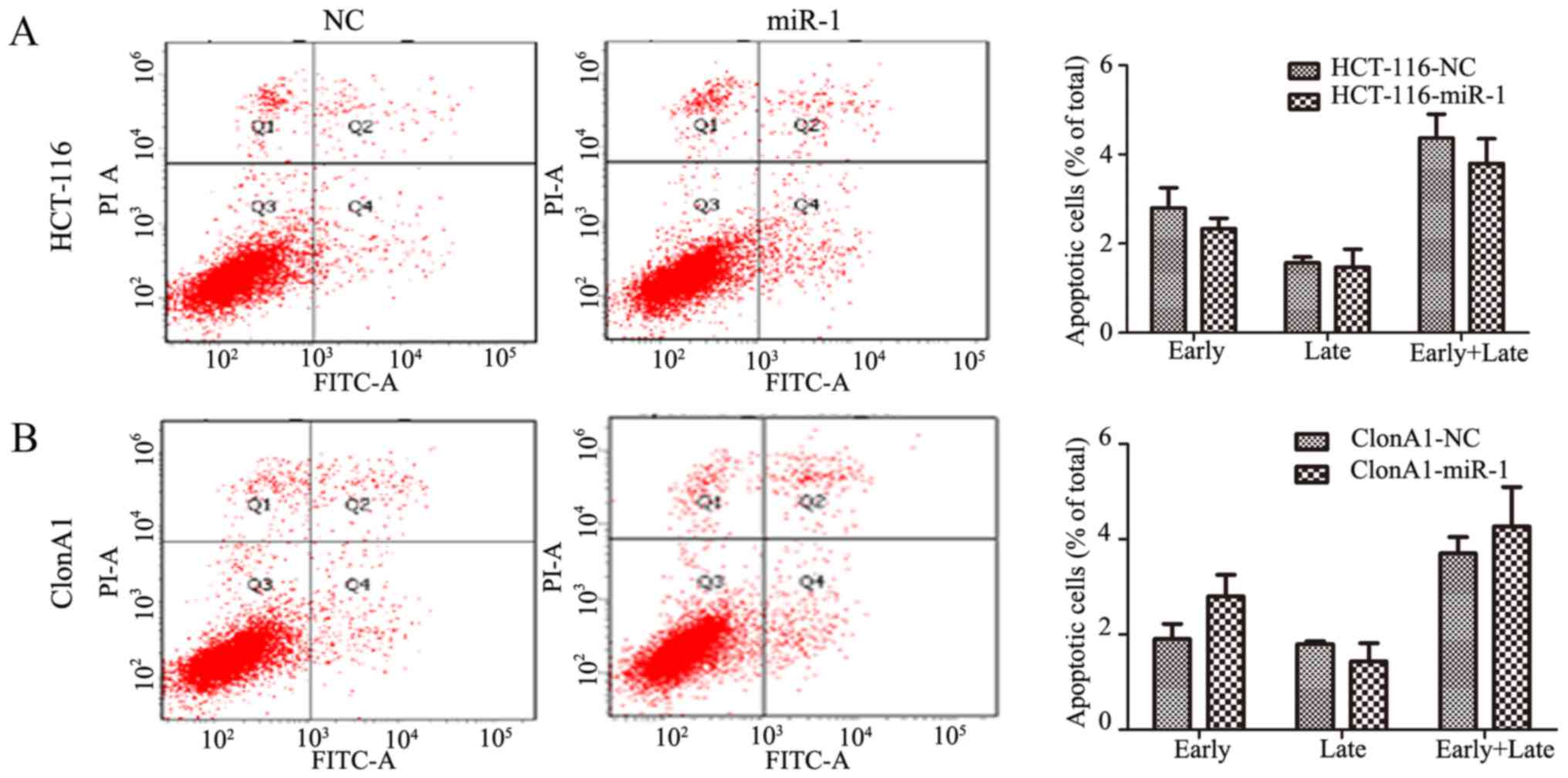
Considering that apoptosis is also a key factor
affecting tumor growth, Annexin V and PI staining was performed to
assess whether miR-1 increased the rate of apoptosis.
Interestingly, no obvious change was observed in
miR-1-overexpressing HCT-116 and ClonA1 cells compared with the
negative control cells (P>0.05, Fig.
3). This finding indicated that the effect of miR-1 on tumor
proliferation is mainly produced through inhibiting cell cycle
transition.
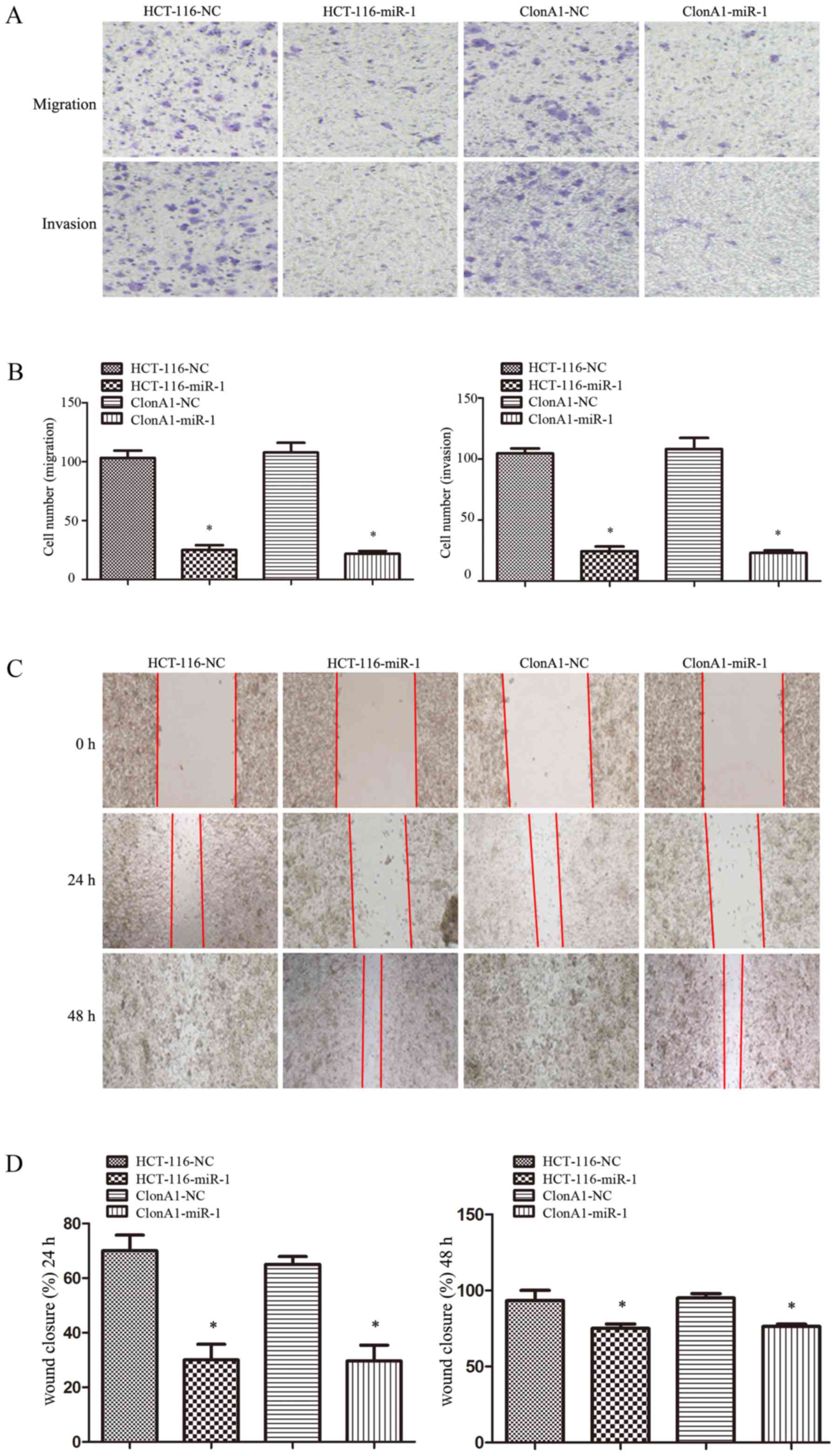
Overexpression of miR-1 suppresses
tumor cell migration and motility in vitro
The ability to invade and migrate through the
basement membrane is the most important property of metastatic
cancer cells. To assess the effect of miR-1 on tumor cell invasion
and motility, Transwell and wound-healing assays were performed.
For the Transwell assay, miR-1-overexpressing or control cells were
seeded on the upper chamber of 24-well Transwell plate. The
migrated cells on the lower chamber of the insert were fixed,
stained, photographed and counted after incubation for 24 h. As
shown in Fig. 4A and B, the
migration rate of miR-1-overexpressing HCT-116 and ClonA1 cells was
inhibited by 76 and 68%, respectively, compared with the control
cells. A corresponding effect on invasion ability was also observed
in a parallel invasion assay.
For the wound-healing assay, the motility of cells
after scratching was monitored under a microscope at different time
points. The results demonstrated that the edges of the wound had
completely fused within 72 h in the HCT-116 and ClonA1 control
cells; however, miR-1-overexpressing cells exhibited a slower wound
closure rate (Fig. 4C and D). All
these results demonstrated that increasing the miR-1 expression in
CRC cells markedly inhibited their migration and motility
abilities.
miR-1 targets and suppresses VEGF
expression in CRC cells
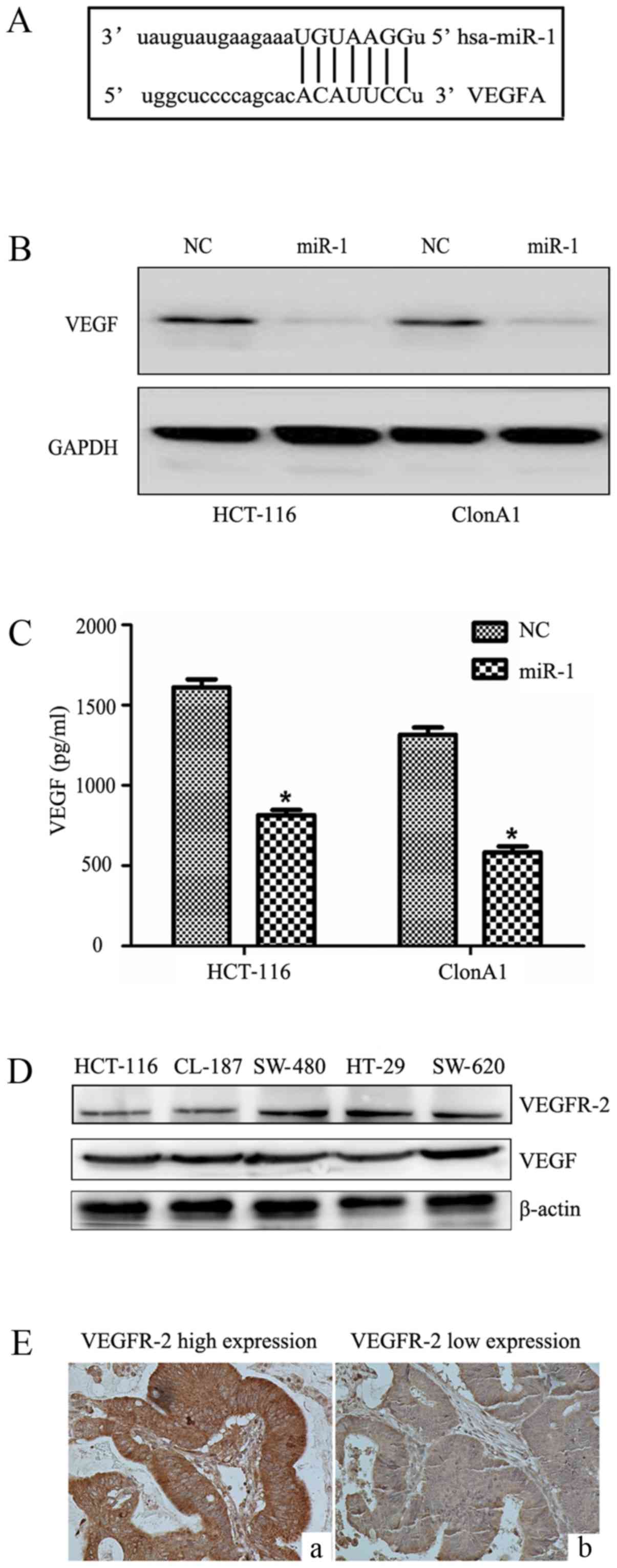
We performed bioinformatic analysis with the
TargetScan, PicTar and Miranda algorithms, and identified a
potential target site in the 3′UTR of VEGF that may interact with
miR-1 (Fig. 5A). To further
determine how miR-1 regulates VEGF, western blotting and ELISA were
performed. We observed that miR-1 overexpression significantly
reduced VEGF protein expression in the HCT-116 and ClonA1 cells
(Fig. 5B), whereas the levels of
VEGF in the culture supernatants of the two cell lines were also
downregulated (Fig. 5C). The
expression of VEGFR-2 and VEGF were examined in five CRC cell lines
(Fig. 5D) by western blotting.
Furthermore, the expression of VEGFR-2 in tumor tissues (Fig. 5E) was assessed by
immunohistochemistry. The results revealed that CRC cell lines and
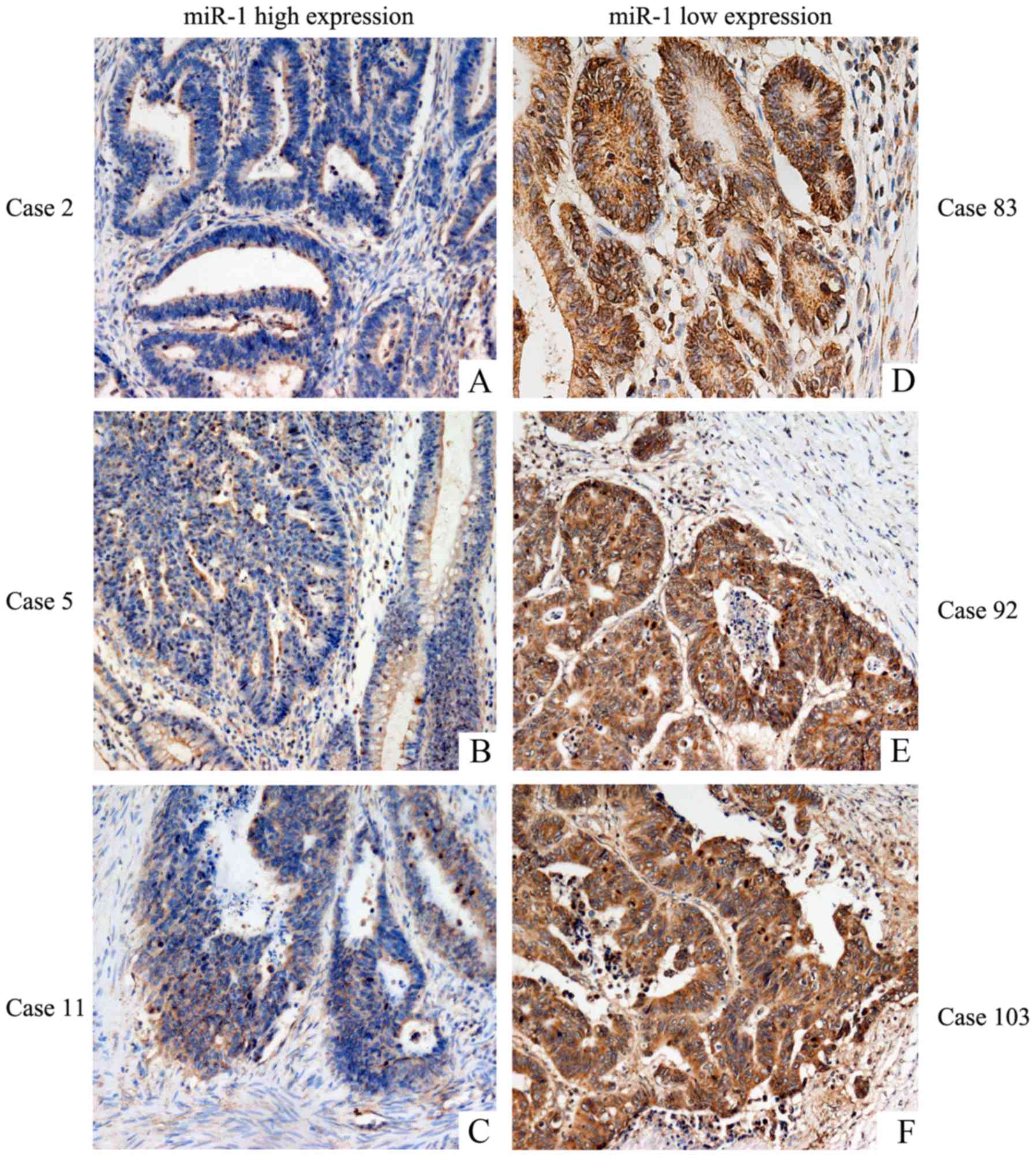
tumor tissues express VEGFR-2. The expression of VEGF was low in
tissues highly expressing miR-1 (Fig.
6A-C), and VEGF was highly expressed in tissues exhibiting low
miR-1 expression (Fig. 6D-F). These
results strongly indicated that miR-1 downregulates the expression
of VEGF at the post-transcriptional level. The results regarding
the negative correlation between the expression of miR-1 and VEGF
are presented in Table II
(P<0.001).
Discussion
Colorectal cancer (CRC) is the third most common
type of cancer worldwide. It is characterized by various hallmarks,
including excessive proliferation, high invasion ability, cellular
heterogeneity, hypoxia, angiogenesis, necrosis and infiltration by
inflammatory cells (2). Despite
major advances in diagnostics and treatment, the prognosis for CRC
patients has not significantly improved over the past decades, and
it remains a major public health concern (1–5).
Therefore, the elucidation of the molecular mechanisms underlying
tumor metastasis and progression is crucial for improving the
treatment and prognosis of CRC patients.
miRNAs are important non-coding RNAs that
participate in the regulation of numerous cellular pathways by
controlling the protein expression of target genes. miR-1, which is
abundant in cardiac tissues and muscles, regulates numerous
functions in the heart, such as cardiac morphogenesis, electrical
conduction and cell cycle control (24). Moreover, miR-1 has also been found
to be a tumor-suppressor miRNA in lung, thyroid and gastric cancer,
as well as head and neck squamous cell carcinoma, by inhibiting
cancer cell proliferation and metastasis (25–28).
In esophageal squamous cell carcinoma and renal cell carcinoma,
miR-1 was found to repress cell proliferation by targeting CDK4
(29). In CRC, miR-1 and miR-374
have been identified as potential biomarkers (16). Recently, miRNA profiling revealed
that miR-1 is downregulated in liver metastases from CRC,
indicating that miR-1 is a potential regulator of angiogenesis
(30,31), although the function of miR-1 and
its underlying mechanism require further investigation. In this
study, we examined the expression of miR-1 in 111 pairs of human
CRC and adjacent tissues and 5 CRC cell lines, and found that the
expression of miR-1 was significantly downregulated. Our data also
revealed that a decreased miR-1 level is negatively associated with
increased malignant behavior of CRC. Further experiments
demonstrated that the restoration of miR-1 expression in CRC cell
lines reduced cell proliferation, and induced
G0/G1 phase arrest, cell invasion and
motility, which provides a theoretical and practical basis for
miR-1 to be used as a biological marker in diagnosis, and a target
for CRC therapy.
Our further experiments revealed that VEGF is a
target gene of miR-1 in CRC cell lines, indicating that miR-1 can
inhibit the protein expression of VEGF-A and its paracrine effects.
There was a negative correlation between miR-1 and VEGF expression
in 111 colon cancer tissues, indicating that the biological effect
of miR-1 may be mediated by inhibiting the VEGF pathway. VEGF-A is
a pivotal angiogenic factor (32,33),
and angiogenesis is involved in a wide variety of physiological and
pathological processes, including organogenesis, development,
inflammation, wound healing and carcinogenesis. Various
transcriptional factors, including HIF1α (34), CREB (35) and NF-κB (36), may promote the expression of VEGF in
tumors. Several studies have reported that knockdown of VEGF-A in
cancer cells suppresses tumor proliferation and invasion. Previous
studies have also demonstrated that the high protein expression of
VEGF in CRC is associated with a relatively poor clinical outcome
and liver metastasis (37,38). In addition, several drugs targeting
the VEGF pathway, including vandetanib, avastin, ranibizumab and
zaltrap, are widely used to treat CRC (39–41).
Furthermore, a recent study demonstrated that miR-1 inhibited the
epithelial-mesenchymal transition and metastasis via regulating the
MAPK and PI3K/AKT pathways (42).
As the MAPK and PI3K/AKT pathways are upregulated and activated by
VEGF, our results suggest that miR-1 may affect the MAPK and
PI3K/AKT pathways via VEGF downregulation. The mechanism underlying
the effects of miR-1 may be complex, and may involve
cross-interactions of additional factors and signaling pathways,
which requires further investigation.
In conclusion, the present study confirmed that
miR-1 acts as a tumor suppressor; miR-1 was found to be
significantly downregulated in CRC cell lines and tissue specimens.
Furthermore, the restoration of miR-1 expression inhibited tumor
proliferation, cell cycle transition, migration and motility by
affecting VEGF expression. These findings indicate a novel
mechanism of tumor suppression by miR-1, and provide a theoretical
and experimental basis for the development of targeted treatments
for CRC through miR-1 and VEGF.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Shenyang
Science and Technology Project of China (grant no. F14-158-9-35)
and the Special-Term Professor from the Educational Department of
Liaoning Province, China (Liao Cai Zhi Jiao no. 2012-512).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
DZ, YS and JZ conceived and supervised the study;
DZ, YS, DZ, GJ and XZ designed experiments; DZ and MD performed
experiments; MD and JZ analyzed data; D Z and DZ wrote the
manuscript; MD and JZ made manuscript revisions. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the China Medical University, and written informed consent was
obtained prior to sample collection.
Consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Clarke SJ, Karapetis CS, Gibbs P, Pavlakis
N, Desai J, Michael M, Tebbutt NC, Price TJ and Tabernero J:
Overview of biomarkers in metastatic colorectal cancer: Tumour,
blood and patient-related factors. Crit Rev Oncol Hematol.
85:121–135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sigurdsson JA, Getz L, Sjönell G,
Vainiomäki P and Brodersen J: Marginal public health gain of
screening for colorectal cancer: Modelling study, based on WHO and
national databases in the Nordic countries. J Eval Clin Pract.
19:400–407. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar R, Jain K, Beeke C, Price TJ,
Townsend AR, Padbury R, Roder D, Young GP, Richards A and Karapetis
CS: A population-based study of metastatic colorectal cancer in
individuals aged ≥80 years: Findings from the South Australian
clinical registry for metastatic colorectal cancer. Cancer.
119:722–728. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Melucci E, Cosimelli M, Carpanese L, Pizzi
G, Izzo F, Fiore F, Golfieri R, Giampalma E, Sperduti I, Ercolani
C, et al: Decrease of survivin, p53 and Bcl-2 expression in
chemorefractory colorectal liver metastases may be predictive of
radiosensivity after radioembolization with yttrium-90 resin
microspheres. J Exp Clin Cancer Res. 32:132013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rawson JB and Bapat B: Epigenetic
biomarkers in colorectal cancer diagnostics. Expert Rev Mol Diagn.
12:499–509. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miller S and Steele S: Novel molecular
screening approaches in colorectal cancer. J Surg Oncol.
105:459–467. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Akbari Moqadam F, Pieters R and den Boer
ML: The hunting of targets: Challenge in miRNA research. Leukemia.
27:16–23. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okayama H, Schetter AJ and Harris CC:
MicroRNAs and inflammation in the pathogenesis and progression of
colon cancer. Dig Dis. 30 (Suppl 2):S9–S15. 2012. View Article : Google Scholar
|
|
9
|
Kanematsu S, Tanimoto K, Suzuki Y and
Sugano S: Screening for possible miRNA-mRNA associations in a colon
cancer cell line. Gene. 533:520–531. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Katoh M: Therapeutics targeting
angiogenesis: Genetics and epigenetics, extracellular miRNAs and
signaling networks (Review). Int J Mol Med. 32:763–767. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pereira DM, Rodrigues PM, Borralho PM and
Rodrigues CM: Delivering the promise of miRNA cancer therapeutics.
Drug Discov Today. 18:282–289. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Roitbak T, Bragina O, Padilla JL and
Pickett GG: The role of microRNAs in neural stem cell-supported
endothelial morphogenesis. Vasc Cell. 3:252011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kusakabe R, Tani S, Nishitsuji K, Shindo
M, Okamura K, Miyamoto Y, Nakai K, Suzuki Y, Kusakabe TG and Inoue
K: Characterization of the compact bicistronic microRNA precursor,
miR-1/miR-133, expressed specifically in Ciona muscle tissues. Gene
Expr Patterns. 13:43–50. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li L, Sarver AL, Alamgir S and Subramanian
S: Downregulation of microRNAs miR-1, −206 and −29 stabilizes PAX3
and CCND2 expression in rhabdomyosarcoma. Lab Invest. 92:571–583.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nasser MW, Datta J, Nuovo G, Kutay H,
Motiwala T, Majumder S, Wang B, Suster S, Jacob ST and Ghoshal K:
Down-regulation of micro-RNA-1 (miR-1) in lung cancer. Suppression
of tumorigenic property of lung cancer cells and their
sensitization to doxorubicin-induced apoptosis by miR-1. J Biol
Chem. 283:33394–333405. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu X, Li S, Xu X, Wu S, Chen R, Jiang Q,
Li Y and Xu Y: The potential value of miR-1 and miR-374b as
biomarkers for colorectal cancer. Int J Clin Exp Pathol.
8:2840–2851. 2015.PubMed/NCBI
|
|
17
|
Sayagués JM, Corchete LA, Gutiérrez ML,
Sarasquete ME, Del Mar Abad M, Bengoechea O, Fermiñán E, Anduaga
MF, Del Carmen S, Iglesias M, et al: Genomic characterization of
liver metastases from colorectal cancer patients. Oncotarget.
7:72908–72922. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang X, Chen X, Fang J and Yang C:
Overexpression of both VEGF-A and VEGF-C in gastric cancer
correlates with prognosis, and silencing of both is effective to
inhibit cancer growth. Int J Clin Exp Pathol. 6:586–597.
2013.PubMed/NCBI
|
|
19
|
Martins SF, Garcia EA, Luz MA, Pardal F,
Rodrigues M and Filho AL: Clinicopathological correlation and
prognostic significance of VEGF-A, VEGF-C, VEGFR-2 and VEGFR-3
expression in colorectal cancer. Cancer Genomics Proteomics.
10:55–67. 2013.PubMed/NCBI
|
|
20
|
Ekinci D, Kargi A, Yalcin AD and Savas B:
The role of VEGF and other parameters in tracking the clinical
course in metronomic chemotherapy. J BUON. 18:245–252.
2013.PubMed/NCBI
|
|
21
|
Zhao SF, Yang XD, Lu MX, Sun GW, Wang YX,
Zhang YK, Pu YM and Tang EY: Prognostic significance of VEGF
immunohistochemical expression in oral cancer: A meta-analysis of
the literature. Tumour Biol. 34:3165–3171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang Y, Zhang Y, Iwamoto H, Hosaka K, Seki
T, Andersson P, Lim S, Fischer C, Nakamura M, Abe M, et al:
Discontinuation of anti-VEGF cancer therapy promotes metastasis
through a liver revascularization mechanism. Nat Commun.
7:126802016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang D, Zhou J and Dong M: Dysregulation
of microRNA-34a expression in colorectal cancer inhibits the
phosphorylation of FAK via VEGF. Dig Dis Sci. 59:958–67. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao Y, Ransom JF, Li A, Vedantham V, von
Drehle M, Muth AN, Tsuchihashi T, McManus MT, Schwartz RJ and
Srivastava D: Dysregulation of cardiogenesis, cardiac conduction,
and cell cycle in mice lacking miRNA-1-2. Cell. 129:303–317. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiao H, Zeng J, Li H, Chen K, Yu G, Hu J,
Tang K, Zhou H, Huang Q, Li A, et al: MiR-1 downregulation
correlates with poor survival in clear cell renal cell carcinoma
where it interferes with cell cycle regulation and metastasis.
Oncotarget. 6:13201–13215. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Leone V, D'Angelo D, Rubio I, de Freitas
PM, Federico A, Colamaio M, Pallante P, Medeiros-Neto G and Fusco
A: MiR-1 is a tumor suppressor in thyroid carcinogenesis targeting
CCND2, CXCR4, and SDF-1alpha. J Clin Endocrinol Metab.
96:E1388–E1398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ueda T, Volinia S, Okumura H, Shimizu M,
Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: A microRNA expression analysis. Lancet Oncol.
11:136–46. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nohata N, Sone Y, Hanazawa T, Fuse M,
Kikkawa N, Yoshino H, Chiyomaru T, Kawakami K, Enokida H, Nakagawa
M, et al: miR-1 as a tumor suppressive microRNA targeting TAGLN2 in
head and neck squamous cell carcinoma. Oncotarget. 2:29–42. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang S, Zhao C, Yang X, Li X, Pan Q,
Huang H, Wen X, Shan H, Li Q, Du Y and Zhao Y: miR-1 suppresses the
growth of esophageal squamous cell carcinoma in vivo and in vitro
through the downregulation of MET, cyclin D1 and CDK4 expression.
Int J Mol Med. 38:113–122. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Slattery ML, Herrick JS, Pellatt DF,
Mullany LE, Stevens JR, Wolff E, Hoffman MD, Wolff RK and Samowitz
W: Site-specific associations between miRNA expression and survival
in colorectal cancer cases. Oncotarget. 7:60193–60205. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Grahnén A and Sjöholm I:
Spectropolarimetric determination of unconjugated bilirubin in
human serum. Z Klin Chem Klin Biochem. 12:2201974.
|
|
32
|
Goel HL and Mercurio AM: VEGF targets the
tumour cell. Nat Rev Cancer. 13:871–882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chekhonin VP, Shein SA, Korchagina AA and
Gurina OI: VEGF in tumor progression and targeted therapy. Curr
Cancer Drug Targets. 13:4232013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weijts BG, Bakker WJ, Cornelissen PW,
Liang KH, Schaftenaar FH, Westendorp B, de Wolf CA, Paciejewska M,
Scheele CL, Kent L, et al: E2F7 and E2F8 promote angiogenesis
through transcriptional activation of VEGFA in cooperation with
HIF1. EMBO J. 31:3871–3884. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kang Z, Zhu H, Luan H, Han F and Jiang W:
Curculigoside A induces angiogenesis through VCAM-1/Egr-3/CREB/VEGF
signaling pathway. Neuroscience. 267:2322014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Anna L, Eugene K, Mayumi J and Matter ML:
Stretch-induced Hypertrophy activates NFkB-mediated VEGF secretion
in adult cardiomyocytes. Plos One. 6:e290552011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Goos JA, de Cuba EM, Coupé VM, Diosdado B,
Delis-Van Diemen PM, Karga C, Beliën JA, Menke-Van der Houven van
Oordt CW, Geldof AA, Meijer GA, et al: Glucose Transporter 1
(SLC2A1) and vascular endothelial growth factor A (VEGFA) predict
survival after resection of colorectal cancer liver metastasis. Ann
Surg. 263:138–145. 2016.PubMed/NCBI
|
|
38
|
Cao D, Hou M, Guan YS, Jiang M, Yang Y and
Gou HF: Expression of HIF-1alpha and VEGF in colorectal cancer:
Association with clinical outcomes and prognostic implications. BMC
Cancer. 9:4322009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhai Z, Yu X, Yang B, Zhang Y, Zhang L, Li
X and Sun H: Colorectal cancer heterogeneity and targeted therapy:
Clinical implications, challenges and solutions for treatment
resistance. Semin Cell Dev Biol. 64:107–115. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Loupakis F, Cremolini C, Yang D, Salvatore
L, Zhang W, Wakatsuki T, Bohanes P, Schirripa M, Benhaim L, Lonardi
S, et al: Prospective validation of candidate SNPs of VEGF/VEGFR
pathway in metastatic colorectal cancer patients treated with
first-line FOLFIRI plus bevacizumab. PLoS One. 8:e667742013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fakih M: The evolving role of
VEGF-targeted therapies in the treatment of metastatic colorectal
cancer. Expert Rev of Anticancer Ther. 13:427–438. 2013. View Article : Google Scholar
|
|
42
|
Yim E, Vivas A, Maderal A and Kirsner RS:
Neuropathy and ankle mobility abnormalities in patients with
chronic venous disease. JAMA Dermatol. 150:385–389. 2014.
View Article : Google Scholar : PubMed/NCBI
|