Introduction
Esophageal cancer, a serious malignancy with respect
to prognosis and mortality rate, accounts for more than 40,000
deaths worldwide annually (1).
According to the statistical data (2–4),
esophageal carcinoma is the eighth most common cancer and the sixth
most common cause of cancer-related deaths worldwide with more than
80% of total cases and deaths in developing nations, while the
prevalence of esophageal cancer is expected to increase by 140% by
2025. Despite many advances in diagnosis and treatment, the 5-year
survival rate for patients diagnosed with esophageal cancer ranges
only from 15 to 20% (5,6). Therefore, it is necessary to explore
novel therapeutic targets to achieve an improved treatment.
Exploration and understanding of the molecular mechanisms involved
in the development and progression of esophageal cancer provide
possible clues for an improved treatment goal.
Dysregulated expression of G-protein-coupled
receptor (GPCR) and dysregulation of GPCR signaling have been
recognized as a hallmark of cancer (7). Several studies have revealed that GPCR
could affect the multiple biological processes of cancer cells,
including proliferation, migration and invasion (8–10),
while these processes were involved in the development and
progression of cancer. Recently, several GPCRs identified as free
fatty acid receptors have emerged as key players in various
physiological homeostasis mechanisms, and GPR120 is one of the
receptors (11). Oh et al
(12) have demonstrated that GPR120
could function as functional ω-3 PUFA receptor that mediates potent
insulin sensitizing and anti-diabetic effects in vivo by
supressing macrophage-induced adipose tissue inflammation in obese
mice. Since the inflammation effects and macrophage-related
properties are cancer related (13), GPR120 was considered to play a role
in tumorigenesis. However, the role of GPR120 in esophageal cancer
has not yet been explored.
In the present study, we explored the relationship
between GPR120 and esophageal cancer and investigated the function
and mechanisms of GPR120 in esophageal cancer cells in order to
elucidate the role of GPR120 in the development and progression of
esophageal cancer.
Materials and methods
Ethical approval of the study
protocol
All research involving human participants was
approved by the Institutional Review Board of The First Affiliated
Hospital of Bengbu Medical School (Bengbu, China). Written informed
consent was obtained from the participating individuals. The study
protocol on animal research was approved by the Institutional
Animal Care and Use Committee of The First Affiliated Hospital of
Bengbu Medical School which is adherent to the accepted
international guidelines for animal experimentation.
Tissue collection
A total of 100 specimens surgically derived from 100
esophageal cancer patients treated from January 2012 to December
2014 at The First Affiliated Hospital of Bengbu Medical College
were collected. Among them, 50 cases belonged to matched tumor and
normal mucosae, which were taken at least 5–10 cm away from the
edges of a tumor of the same patient. All the enrolled patients did
not receive any neoadjuvant chemotherapy or radiation therapy prior
to esophagectomy. Fresh tumor tissues or corresponding normal
esophageal mucosae were immediately frozen in liquid nitrogen after
dissection, then stored at −80°C until further analysis. Tumor
specimens were carefully microdissected to ensure that at least 90%
of the analyzed tissue contained cancer cells. The clinical
diagnosis, tumor stage, histological differentiation and resection
margin were determined by routine histopathological examination of
hematoxylin and eosin (H&E) stained specimens by an experienced
pathologist.
Immunohistochemistry
The paraffin specimen of each patient was stained by
H&E staining and the pathological type of the tissue was
analyzed by an experienced pathologist. The immunohistochemistry
was performed using streptavidin-peroxidase method. The section was
deparaffinated by the Leica TP1020 tissue processor (Leica
Instruments, Mussloch, Germany) and pre-treated with microwave
antigen retrieval procedure at 100°C for 5 min in 10 mM citrate
buffer (pH 6.0). After incubation in 3% hydrogen peroxide for 6
min, washing with phosphate-buffered saline (PBS) for 3
times, the slide was blocked using 50 µl goat serum at room
temperature (RT) for 30 min. Subsequently, the primary anti-GPR120
antibody (dilution 1:100; cat. no. ab118757; Abcam, Cambridge, MA,
USA) was added and the slide was incubated at 4°C overnight.
Subsequently, the HRP-conjugated sreptavidin was added followed by
washing 3 times with PBS. To visualize the immunostaining, DAB
(Dako, Carpinteria, CA, USA) was used. After the tissue turned
yellow, the sections were washed, re-stained with hematoxylin,
dehydrated and covered.
Assessment of the
immunohistochemistry
The intensity of the immunostaining was evaluated by
two pathologists without knowing the clinical history of the
patients. The cells with dyed membrane or cytoplasm were considered
as positive. Five representative regions of ×200 magnification were
selected to observe and at least 200 cancer cells were presented at
that regions. Frequency and staining intensity of GPR120 by tumor
cells were analyzed, and the expression of GPR120 was quantified
using the modified Histo-score (H-score) (14), with a range of possible scores from
0 to 300. The expression of GPR120 was categorized into two groups
according to the frequency distributions of the H-scores, using a
cut-off score of >100 (H-score, 0–99=negative/low expression and
100–300=positive/high expression).
Cell culture
Human esophageal cancer cell lines Eca-109, TE-1 and
KYSE450 and human colorectal cancer cell line SW480 were obtained
from the American Type Culture Collection (ATCC, Manassas, VA,
USA). The cells were maintained as monolayer cultures in cell
culture flasks with RPMI-1640 medium containing 10% (v/v) fetal
bovine serum (FBS) and 1% antibiotics. Cells were cultured at 37°C
in a humidified atmosphere with 5% CO2. All the cell
culture media and additives were purchased from Invitrogen (Thermo
Fisher Scientific, Inc., Waltham, MA, USA).
Lentiviral shRNA particles
Recombinant lentiviral particles expressing GPR120
or control siRNA were obtained from GenePharma Co. Ltd. (Shanghai,
China). Cells were grown to a certain degree of confluency ~40% and
then infected with lentiviral particles in complete medium for 48
h. To increase infection efficiency, cells were co-treated with the
cationic polymer Polybrene (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany; 8 µg/ml in water). Neither shRNA nor Polybrene affected
cell viability. The siRNA and shRNA had no off-target effects and
at the indicated multiplicity of infection (MOI) and duration,
failed to modulate cell adherence, shape and viability.
Real-time quantitative PCR
Total cellular RNA of human esophageal cancer cell
line Eca-109 was extracted using TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). RT-PCR was performed using a One Step
SYBR® PrimeScript™ RT-PCR kit (Takara Biotechnology,
Co., Ltd., Dalian, China) and an iQ5 real-time PCR Detection system
(Bio-Rad Laboratories, Hercules, CA, USA). The expression of
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene was assessed
simultaneously in all samples as an internal control. Relative gene
expression was determined by the 2−ΔΔCt method (15). Oligonucleotide primers specific for
GPR120, vascular endothelial growth factor (VEGF), interleukin-8
(IL-8), cyclooxygenase-2 (Cox-2) and GAPDH are listed in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Forward primer | Reverse primer |
|---|
| VEGF |
5′-TGCAATGGATCAAGGACCAGAGG-3′ |
5′-TGCAGCCAGCAAGAAGCATCAG-3′ |
| IL-8 |
5′-CCGAGGATCTGATGACGATTA-3′ |
5′-GGCTCCCAGAAATAGCTTCAA-3′ |
| Cox-2 |
5′-CACAGCACAGCCAGGAAGG-3′ |
5′-GTTCCCTGGCTCTGAGTAGTCGA-3′ |
| GAPDH |
5′-GGATTTGGTCGTATTGGG-3′ |
5′-GGAAGATGGTGATGGGATT-3′ |
Western blotting
Cells obtained from the above-mentioned treatment
were lysed in RIPA buffer, followed by high-speed centrifugation
and protein quantification using a bicinchoninic acid assay (Thermo
Fisher Scientific). Cellular proteins were separated by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and transferred
onto polyvinylidenedifluoride membranes. After blocking, the
membranes were incubated with anti-total-(1:1,000; cat. no. 9272)
or -phospho-Akt (1:1,000; cat. no. 5012), phospho-IκB (1:1,000;
cat. no. 2859), E-cadherin (1:1,000; cat. no. 3195), N-cardherin
(1:1,000; cat. no. 13116), vimentin (1:1,000; cat. no. 5741) (Cell
Signaling Technology, Inc., Danvers, MA, USA) and GPR120 monoclonal
primary antibodies (dilution 1:1,000; cat. no. ab118757; Abcam,
Cambridge, MA, USA). GAPDH (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) was used as the loading control. Appropriate
horseradish peroxidase-conjugated secondary antibodies (goat
anti-rabbit HRP conjugate antibody: Dilution 1:2,000; cat. no.
7074; Cell Signaling Technology; and goat anti-mouse HRP-conjugate
antibody: 1:2,000; cat. no. 7076; Cell Signaling Technology) were
applied to detect labeled proteins. The protein bands were
developed with SuperSignal Ultra Chemiluminescent Substrate
(Pierce; Thermo Fisher Scientific) on X-ray films (Kodak Japan
Ltd., Tokyo, Japan).
Cell proliferation
Human esophageal cancer cell line Eca-109
(3×103 cells) were seeded in 96-well plates in complete
medium and infected with GPR120 or control siRNA lentivirus
particles. Two days later, cell proliferation was evaluated by Cell
Counting Kit-8 (CCK-8) method according to the manufacturer's
instructions using a microplate reader (Molecular Devices,
Sunnyvale, CA, USA) to assess the absorbance.
Clone formation
Human esophageal cancer cell line Eca-109 (800
cells) were seeded in 6-well plates in complete medium and infected
with GPR120 or control siRNA lentivirus particles. After medium
replacement at 24 h post-infection, the cells were maintained at
37°C in a humidified atmosphere with 5% CO2 for 7 days,
and then they were stained with crystal violet. The colony survival
with a definition of >50 cells were counted under a light
microscope (DM4000B; Leica Microsystems, Benshein, Germany). The
whole process was performed 3 times to obtain a mean number of
colony formation.
Scratch assay
Human esophageal cancer cell line Eca-109 infected
with GPR120 or control siRNA lentivirus particles were plated at
70,000 cells/well in a 12-well plate. Cells were grown to 90%
confluency and scratched once using a sterile 1-ml pipette tip,
washed twice with complete medium to remove floating cells and cell
components. Images were captured at a ×40 magnification using a
Leica inverted phase contrast microscope (DM IRB; Leica
Microsystems). The area of the gap at 24 h was assessed and
subtracted from that at 0 h to quantify the migrated cells. The
experiments were repeated at least 3 times with similar
results.
Cell invasion
A Transwell system was employed to perform the cell
invasion assay. Briefly, resuspended Eca-109 cells
(2×105 cells) infected with GPR120 or control siRNA
lentivirus particles were seeded into the upper chamber prefilled
with Matrigel and RPMI-1640 medium supplemented with 20% FBS was
added to the lower chamber. After the Transwell plate was
maintained in a routine cell culture incubator for a specific
time-point, the upper chamber was retained and the membranes were
obtained for hematoxylin staining. The cell number of each membrane
was determined in 3 randomly picked fields (magnification, ×200)
under a light microscope. All the experiments were performed in
triplicate.
Nude mice model of ectopic tumor
Athymic nude (nu/nu) 6-weeks old mice were purchased
from Shanghai SLAC Laboratory Animal Co. Ltd. (Shanghai, China).
The tumors were generated by subcutaneous injection of
2×106 GPR120 or control siRNA lentivirus particles
infected Eca-109 cells suspended in 50 µl PBS into the dorsal
region near the thigh. Mice were then weighted and assessed for
tumor size every other day by measuring tumor length and tumor
width. At week 4 post-treatment, all mice were sacrificed by
cervical dislocation and the tumors were excised, weighted and
imaged. For histological analysis, organs from the treated groups
and the control group were fixed in 4% formalin, and then conducted
with paraffin-embedded sections for H&E staining. The slices
were examined by a digital microscope (Leica QWin Plus v3 software;
Leica Microsystems).
Enzyme-linked immunosorbent assay
(ELISA)
Condition medium was obtained from the
above-described cell culture at 1,500 × g centrifugation for 10 min
and was stored at −80°C before further processing. Angiogenesis and
inflammation-related cytokines including VEGF, IL-8 and PGE2, were
determined by ELISA kit (Invitrogen; Thermo Fisher Scientific)
according to the manufacturer's instructions.
Statistical analysis
All statistical analyses were performed using SPSS
version 18 (SPSS, Inc., Chicago, IL, USA). Data are presented as
the mean ± SD. The Student's t-test or one-way analysis of variance
(ANOVA) were used to examine differences between groups. A P-value
of <0.05 was considered to indicate a statistically significant
difference.
Results
Correlation analysis between GPR120
level and clinical parameters in esophageal cancer
In order to investigate the role of GPR120 in
esophageal cancer, we firstly performed the correlation analysis
between the expression level of GPR120 and the clinical parameters
of the esophageal cancer patients. As displayed in Table II, the expression level of GPR120
was significantly elevated in esophageal cancer tissues and
correlated with histological grade (P<0.001), lymph node
metastasis (P=0.003) and metastasis depth (P<0.001). These
results indicated that GPR120 affected the progression of
esophageal cancer.
| Table II.Correlation analysis between the
level of GPR-120 and the clinical parameters. |
Table II.
Correlation analysis between the
level of GPR-120 and the clinical parameters.
|
|
| GPR-120 |
|
|
|---|
|
|
|
|
|
|
|---|
| Parameters | Cases (n=100) | Low | High | χ2 | P-value |
|---|
| Age (years) |
|
|
| 0.219 | 0.640 |
|
≤60 | 43 | 11 | 32 |
|
|
|
>60 | 57 | 17 | 40 |
|
|
| Sex |
|
|
| 0.118 | 0.732 |
|
Female | 42 | 11 | 31 |
|
|
|
Male | 58 | 17 | 41 |
|
|
| Tissue type |
|
|
| 112.5 | <0.001 |
|
Normal | 100 | 100 | 0 |
|
|
|
Tumor | 100 | 28 | 72 |
|
|
| Size (cm) |
|
|
| 0.842 | 0.359 |
| ≤5 | 57 | 18 | 39 |
|
|
|
>5 | 43 | 10 | 33 |
|
|
| Histological
grade |
|
|
| 43.96 | <0.001 |
| I | 16 | 15 | 1 |
|
|
| II | 64 | 13 | 51 |
|
|
|
III | 20 | 0 | 20 |
|
|
| Location |
|
|
| 0.121 | 0.942 |
| Neck
and upperthoracic | 10 | 3 | 7 |
|
|
|
Mid-thoracic | 48 | 14 | 34 |
|
|
|
Lower-thoracic | 42 | 11 | 31 |
|
|
| Lymph node
metastasis |
|
|
| 8.73 | 0.003 |
|
Yes | 45 | 6 | 39 |
|
|
| No | 55 | 22 | 33 |
|
|
| Metastasis
depth |
|
|
| 16.053 | <0.001 |
|
T1+T2 | 31 | 17 | 14 |
|
|
|
T3+T4 | 69 | 11 | 58 |
|
|
Effects of GPR120 on cell
proliferation, clone formation, cell migration and invasion in
esophageal cancer
Since the expression of GPR120 in esophageal cancer
was elevated, we obtained some esophageal cancer cell lines to
investigate the biological function of GPR120. In order to mimic
the clinical status, we first examined the expression level of
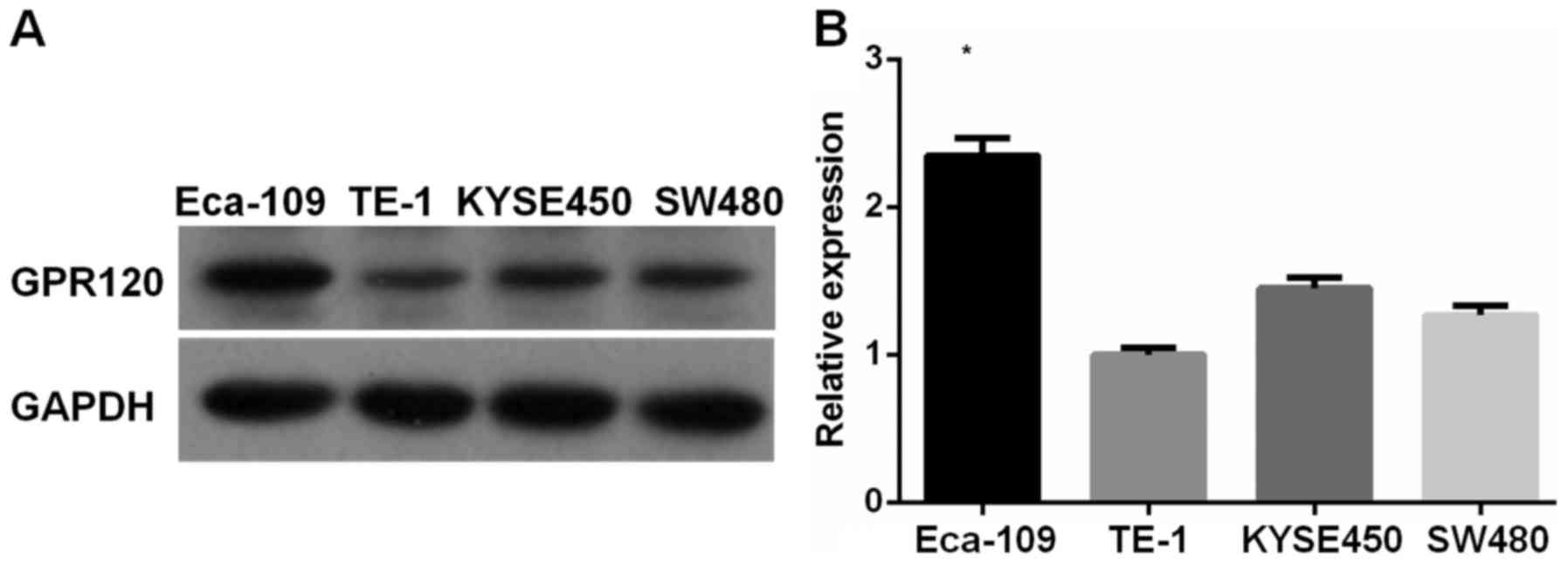
GPR120 in esophageal cancer cell lines. As displayed in Fig. 1, significantly increased level of
GPR120 expression was found in Eca-109 cells compared to TE-1 and
KYSE450 cells. Therefore, Eca-109 cell line was selected as the
cell line for the following experiments. According to the results
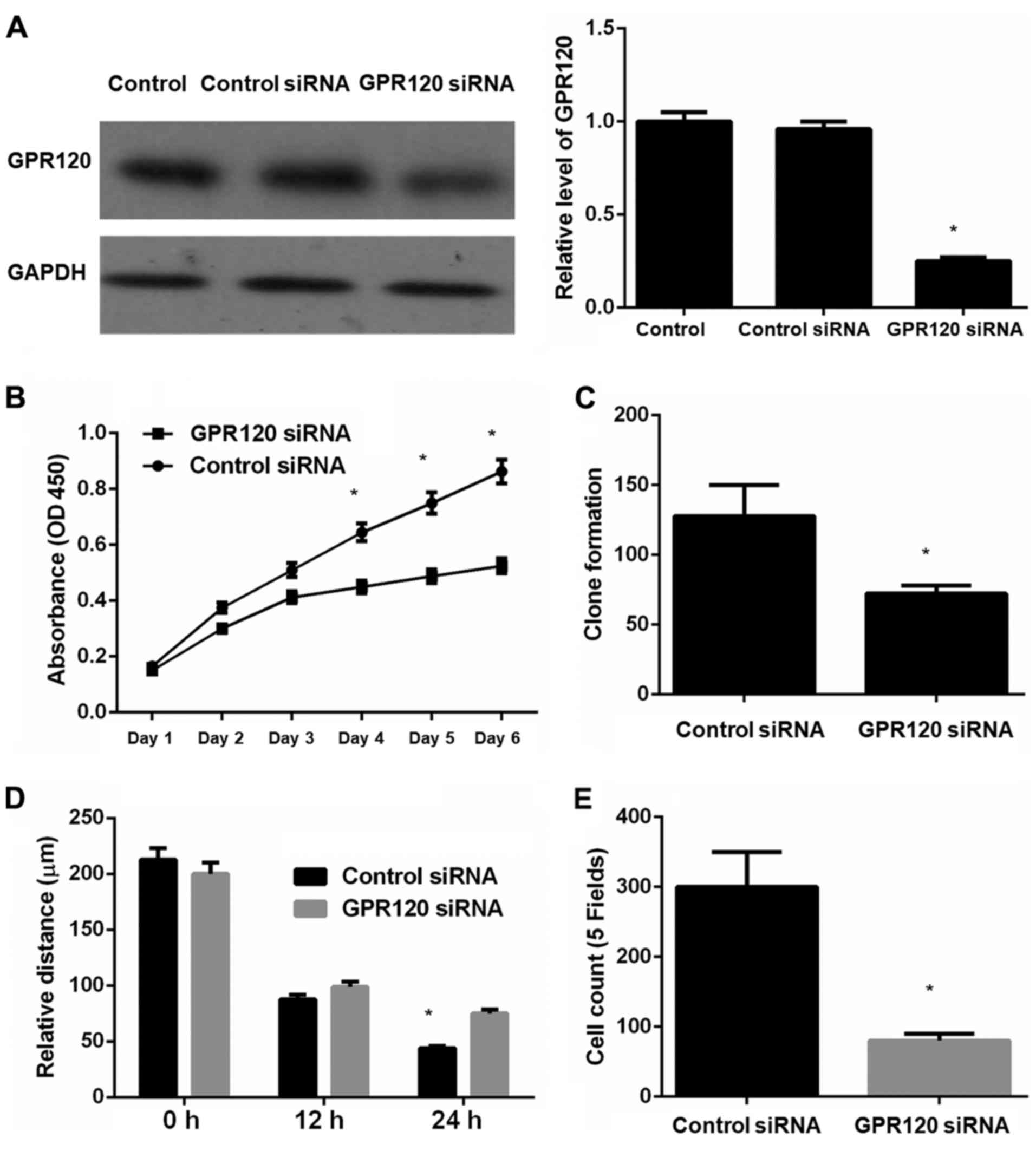
displayed in Fig. 2,
GPR120-knockdown esophageal cancer cell line Eca-109 exhibited a
significantly decreased degree of cell proliferation, clone
formation, cell migration and invasion compared to the control
cells. These results indicated that GPR120 affected the biological
function of esophageal cancer cells via proliferation, clone
formation, migration and invasion.
Effects of GPR120 on tumor growth in
vivo
Due to the tumor-promoting effects of GPR120
observed in vitro, we further established an ectopic tumor
nude mice model to evaluate the effects of GPR120 in vivo.
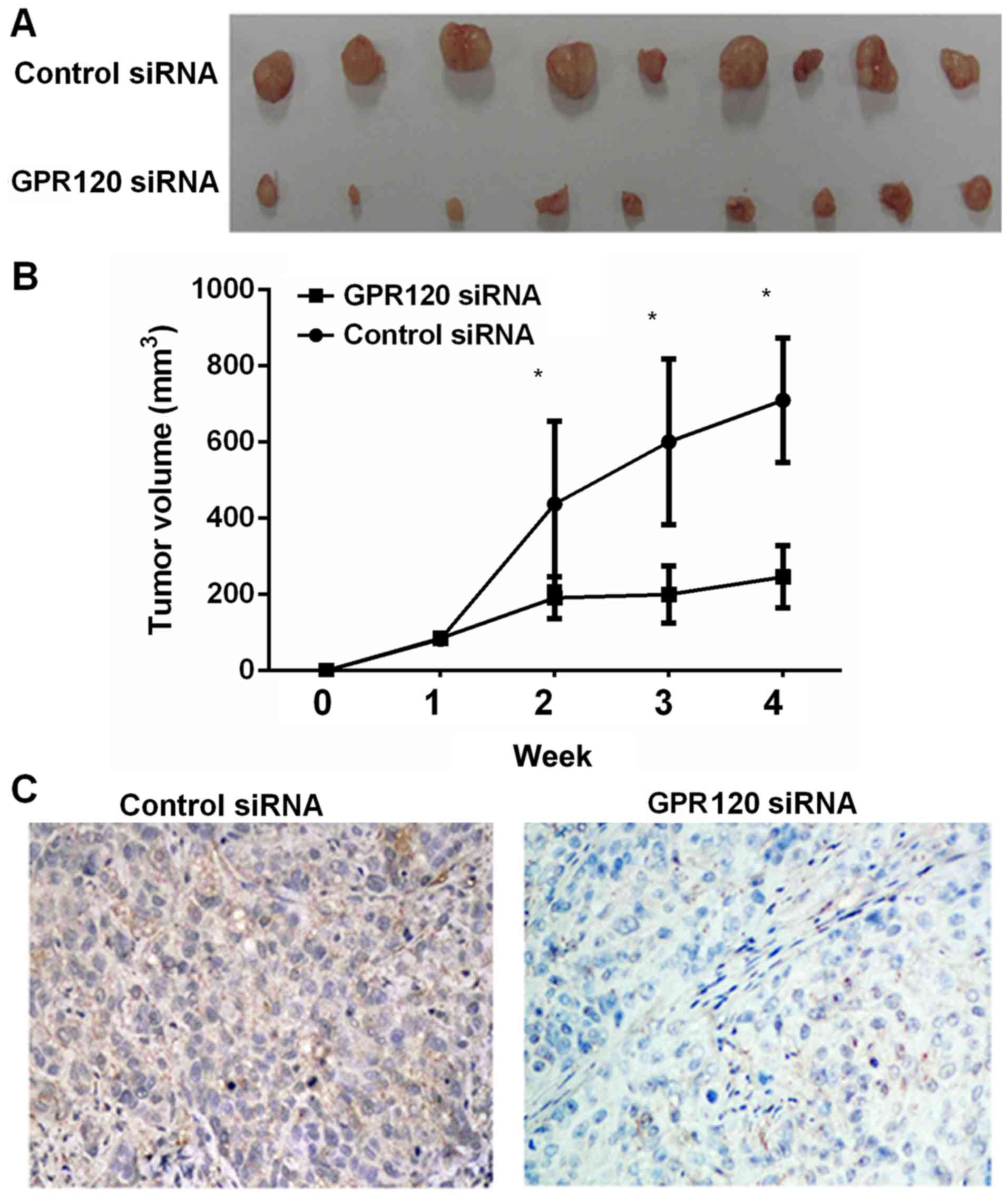
As displayed in Fig. 3A and B,
GPR120-knockdown esophageal cancer cell line Eca-109 exhibited a
decreased level of tumor growth in vivo according to tumor
size and weight. Furthermore, the immunostaining results also
confirmed the effects of GPR120 knockdown in esophageal cancer
cells (Fig. 3C).
Mechanism involved in the effects of
GPR120
We further explored the mechanism involved in the
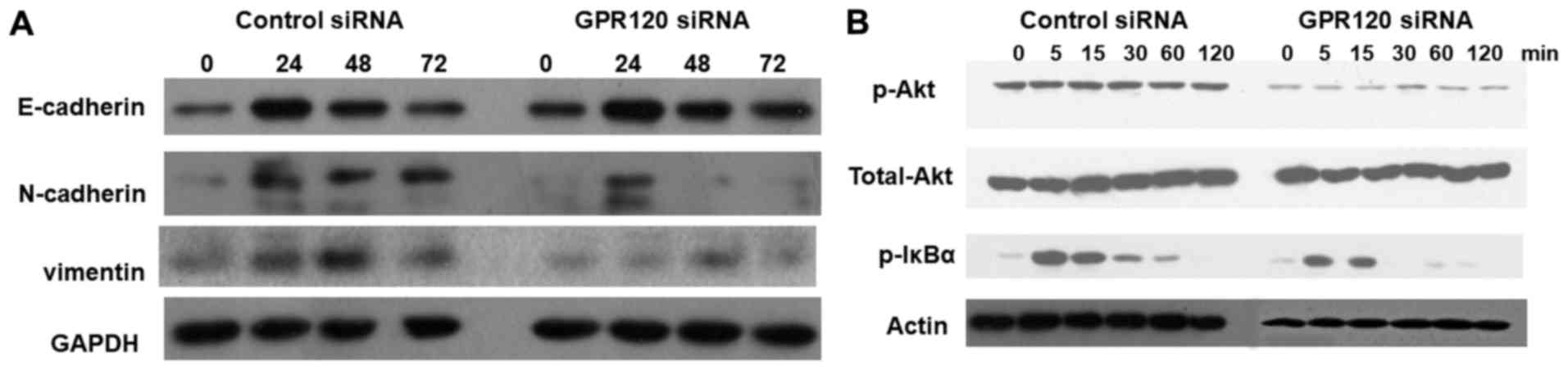
effects of GPR120 in esophageal cancer. Our observations indicated
that GPR120 knockdown in esophageal cancer cell line Eca-109
resulted in increased level of the EMT marker E-cadherin and
decreased level of N-cadherin and vimentin, decreased level of Akt
phosphorylation and I-κB phosphorylation compared to the control
cells (Fig. 4). These results
indicated the possible involvement of EMT process, PI3K/Akt pathway
and NF-κB in the role of GPR120 in esophageal cancer. Furthermore,
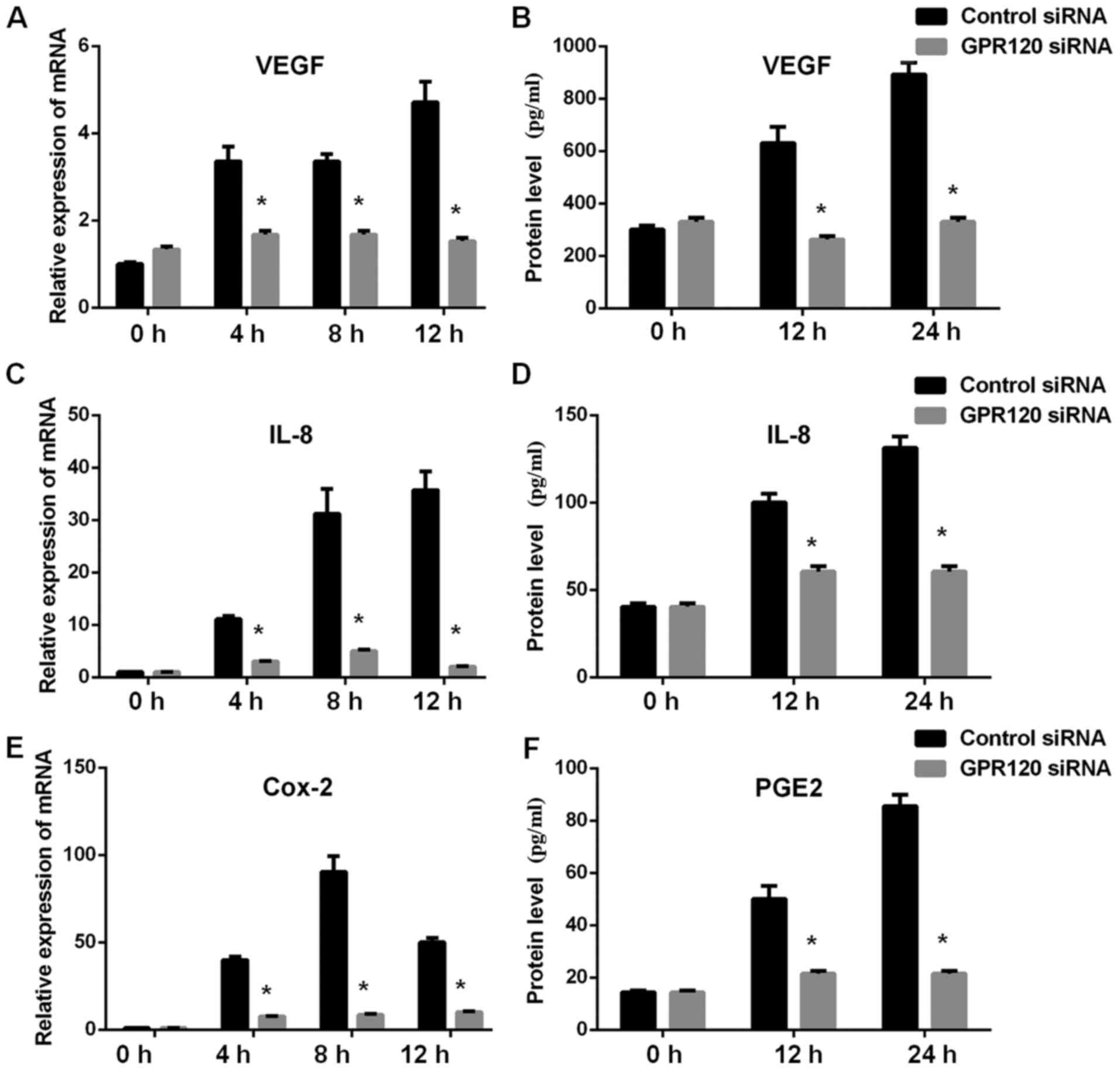
we also examined the possible role of angiogenesis and inflammatory
cytokines on the effects of GPR120 and we observed decreased mRNA
and protein levels of angiogenesis cytokine VEGF, inflammatory
cytokine IL-8 and Cox-2 (protein PGE2) in GPR120-knockdown Eca-109
cells compared to control cells (Fig.
5).
Discussion
Practical strategies have been proposed to prevent
the harmful sequelae of the worldwide obesity epidemic in order to
reduce the future medical burden to society. Research has indicated
the association between obesity and the overall risk for multiple
cancers, including endometrial, colorectal, prostate, pancreatic
and postmenopausal breast cancer (16–20).
However, the exact role of obesity in cancer risk has not been
fully explored. According to a previous study (21), the physiological effects related to
obesity, including increased tissue inflammation, insulin
resistance and/or hyperinsulinemia are considered to play a
critical role in cancer risk. Therefore, dietary intervention is a
potential mean to decrease this type of risk in our daily life.
Altering the balance between dietary ω-3 and ω-6 polyunsaturated
fatty acids (PUFAs) has been considered as an approach for disease
prevention (22,23) and several epidemiological and
preclinical studies have revealed an antitumor effect of ω-3 PUFAs
in cancer patients (24–26). The detailed mechanisms mediated by
which ω-3 PUFAs, particularly eicosapentaenoic acid (EPA) and
docosahexaenoic acid (DHA), exert their anticancer effects are not
well understood despite multiple targets regulating cell
proliferation and survival, inflammation, angiogenesis and
metastasis may be involved (27).
Recently, several GPCRs identified as free fatty acid receptors
have emerged as key players in various physiological homeostasis
mechanisms, and GPR120 has been demonstrated to function as a
receptor for ω-3 PUFAs (11), and
molecular and cellular effects could be generated following the
ligand-receptor interaction.
In the present study, we firstly evaluated the
expression of GPR120 in esophageal cancer tissue and observed
significantly increased GPR120 in esophageal cancer tissues
compared to the normal tissues. Based on this observation, we
performed in vitro and in vivo experiments to
investigate the role of GPR120 in esophageal cancer development and
progression. Our results indicated that GPR120 served as
tumor-promoting regulator in esophageal cancer according to cell
model and nude mice ectopic model. In addition, the investigation
of the underlying mechanism indicated that EMT, PI3K and I-κB
pathway, as well as angiogenesis and inflammation-related cytokines
secretion attributed to the phenotype resulted by GPR120. To the
best of our knowledge, this is the first study to elucidate the
role of GPR120 in esophageal cancer.
Oh et al (12) have demonstrated that GPR120 is a
functional ω-3 PUFA receptor that mediates potent insulin
sensitizing and anti-diabetic effects in vivo by suppressing
macrophage-induced adipose tissue inflammation in obese mice. The
state of chronic, low grade inflammation arising in obesity is
characterized by infiltration of M1-type adipose tissue
macrophages, cells that secrete high levels of proinflammatory
cytokines, including TNF-α, IL1β and IL-6, which are considered to
be major contributors to tissue inflammation and insulin resistance
in obesity (28,29). In cancer patients, increased
inflammation levels are positively correlated with tumor cell
proliferation, tumor stage and lymph node metastasis (30,31).
In the present study, we also demonstrated that GPR120 promoted
tumor cell proliferation, migration and invasion, and its
expression level was associated with tumor stage and lymph node
metastasis.
According to a previous study, the PI3K and NFκB
pathways are involved in the inflammatory signaling pathway and ω-3
PUFAs can inhibit these pathways by sequestering TAB1 in obese mice
(32). In the presents study, we
also demonstrated that GPR120 knockdown resulted in decreased
activity of Akt and I-κB phosphorylation. In a study by Wu et
al (33), they also revealed
that GPR120 exerted its functions via the PI3K and NFκB pathways in
colorectal cancer. However, controversial results also existed. In
a recently published study, Chung et al (34) demonstrated that obesity promoted
mammary tumor progression in a model of postmenopausal breast
cancer and that ω-3 PUFAs inhibited mammary tumor progression in
obese mice, independently of GPR120. We believe that the
differences may be attributed to the type of cancer.
Besides the aforementioned signaling pathways,
GPR120 is considered to enhance cell motility by inducing EMT. In
the present study, we observed significantly increased levels of
E-cadherin and decreased level of N-cadherin and vimentin in GPR120
knockdown esophageal cancer cells compared to control cells, which
is consistent with previous research.
In conclusion, in the present study, we demonstrated
that increased level of GPR120 in esophageal cancer tissues,
functioned as a positive regulator of the development and
progression of esophageal cancer. Furthermore, multiple mechanisms
including EMT, PI3K and I-κB pathway, as well as and angiogenesis
and inflammation-related cytokines secretion were involved.
Acknowledgements
Not applicable.
Funding
This study was supported by the ‘12th Five-year’
Clinical Medical Key Construction Foundation of Anhui province (no.
01Z33).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZC, DL, JL and HJ conceived and designed the
experiments; ZC, DL, JL, YZ, HX, HY, HL, GW, HC, LZ and SY
performed the experiments; ZC, DL, JL, YZ, HX, HY, HL, GW, HC, LZ,
SY and HJ analyzed the data; ZC, DL, JL, YZ, HX, HY, HL, GW, HC,
LZ, SY and HJ contributed reagents/materials/analysis tools; ZC,
DL, JL and HJ contributed to the writing of the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Institutional Review Board of The First Affiliated Hospital of
Bengbu Medical School (Bengbu, China).
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Napier KJ, Scheerer M and Misra S:
Esophageal cancer: A review of epidemiology, pathogenesis, staging
workup and treatment modalities. World J Gastrointest Oncol.
6:112–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Global Burden of Disease Cancer
Collaboration; Fitzmaurice C, Allen C, Barber RM, Barregard L,
Bhutta ZA, Brenner H, Dicker DJ, Chimed-Orchir O, Dandona R,
Dandona L, et al: Global, regional, and national cancer incidence,
mortality, years of life lost, years lived with disability, and
disability-adjusted life-years for 32 cancer groups, 1990 to 2015:
A systematic analysis for the global burden of disease study. JAMA
Oncol. 3:524–548. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liang H, Fan JH and Qiao YL: Epidemiology,
etiology, and prevention of esophageal squamous cell carcinoma in
China. Cancer Biol Med. 14:33–41. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Steury MD, McCabe LR and Parameswaran N: G
protein-coupled receptor kinases in the inflammatory response and
signaling. Adv Immunol. 136:227–277. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nohata N, Goto Y and Gutkind JS: Onco-GPCR
signaling and dysregulated expression of microRNAs in human cancer.
J Hum Genet. 62:87–96. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nogués L, Palacios-García J, Reglero C,
Rivas V, Neves M, Ribas C, Penela P and Mayor F Jr: G
protein-coupled receptor kinases (GRKs) in tumorigenesis and cancer
progression: GPCR regulators and signaling hubs. Semin Cancer Biol.
78–90. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khalil BD, Hsueh C, Cao Y, Abi Saab WF,
Wang Y, Condeelis JS, Bresnick AR and Backer JM: GPCR signaling
mediates tumor metastasis via PI3Kβ. Cancer Res. 76:2944–2953.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hara T, Kashihara D, Ichimura A, Kimura I,
Tsujimoto G and Hirasawa A: Role of free fatty acid receptors in
the regulation of energy metabolism. Biochim Biophys Acta.
1841:1292–1300. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oh DY, Talukdar S, Bae EJ, Imamura T,
Morinaga H, Fan W, Li P, Lu WJ, Watkins SM and Olefsky JM: GPR120
is an omega-3 fatty acid receptor mediating potent
anti-inflammatory and insulin-sensitizing effects. Cell.
142:687–698. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Diakos CI, Charles KA, McMillan DC and
Clarke SJ: Cancer-related inflammation and treatment effectiveness.
Lancet Oncol. 15:e493–e503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
McCarty KS Jr, Miller LS, Cox EB, Konrath
J and McCarty KS Sr: Estrogen receptor analyses. Correlation of
biochemical and immunohistochemical methods using monoclonal
antireceptor antibodies. Arch Pathol Lab Med. 109:716–721.
1985.
|
|
15
|
Ji Y, Strawn TL, Grunz EA, Stevenson MJ,
Lohman AW, Lawrence DA and Fay WP: Multifaceted role of plasminogen
activator inhibitor-1 in regulating early remodeling of vein bypass
grafts. Arterioscler Thromb Vasc Biol. 31:1781–1787. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Golabek T, Bukowczan J, Chłosta P,
Powroźnik J, Dobruch J and Borówka A: Obesity and prostate cancer
incidence and mortality: A systematic review of prospective cohort
studies. Urol Int. 92:7–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kolodecik T, Shugrue C, Ashat M and
Thrower EC: Risk factors for pancreatic cancer: Underlying
mechanisms and potential targets. Front Physiol. 4:4152014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ligibel JA and Strickler HD: Obesity and
its impact on breast cancer: Tumor incidence, recurrence, survival,
and possible interventions. Am Soc Clin Oncol Educ Book.
2013:52–59. 2013. View Article : Google Scholar
|
|
19
|
Patterson RE, Rock CL, Kerr J, Natarajan
L, Marshall SJ, Pakiz B and Cadmus-Bertram LA: Metabolism and
breast cancer risk: Frontiers in research and practice. J Acad Nutr
Diet. 113:288–296. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Van Kruijsdijk RC, van der Wall E and
Visseren FL: Obesity and cancer: The role of dysfunctional adipose
tissue. Cancer Epidemiol Biomarkers Prev. 18:2569–2578. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
James FR, Wootton S, Jackson A, Wiseman M,
Copson ER and Cutress RI: Obesity in breast cancer-what is the risk
factor? Eur J Cancer. 51:705–720. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fares H, Lavie CJ, DiNicolantonio JJ,
O'Keefe JH and Milani RV: Omega-3 fatty acids: A growing ocean of
choices. Curr Atheroscler Rep. 16:3892014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Simopoulos AP: An increase in the
omega-6/omega-3 fatty acid ratio increases the risk for obesity.
Nutrients. 8:1282016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Farvid MS, Eliassen AH, Cho E, Liao X,
Chen WY and Willett WC: Dietary fiber intake in young adults and
breast cancer risk. Pediatrics. 137:e201512262016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fabian CJ, Kimler BF and Hursting SD:
Omega-3 fatty acids for breast cancer prevention and survivorship.
Breast Cancer Res. 17:622015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng JS, Hu XJ, Zhao YM, Yang J and Li D:
Intake of fish and marine n-3 polyunsaturated fatty acids and risk
of breast cancer: Meta-analysis of data from 21 independent
prospective cohort studies. BMJ. 346:f37062013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nabavi SF, Bilotto S, Russo GL, Orhan IE,
Habtemariam S, Daglia M, Devi KP, Loizzo MR, Tundis R and Nabavi
SM: Omega-3 polyunsaturated fatty acids and cancer: Lessons learned
from clinical trials. Cancer Metastasis Rev. 34:359–380. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kraakman MJ, Murphy AJ, Jandeleit-Dahm K
and Kammoun HL: Macrophage polarization in obesity and type 2
diabetes: Weighing down our understanding of macrophage function?
Front Immunol. 5:4702014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ballak DB, Van Diepen JA, Moschen AR,
Jansen HJ, Hijmans A, Groenhof GJ, Leenders F, Bufler P,
Boekschoten MV, Müller M, et al: IL-37 protects against
obesity-induced inflammation and insulin resistance. Nat Commun.
5:47112014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Katanov C, Lerrer S, Liubomirski Y,
Leider-Trejo L, Meshel T, Bar J, Feniger-Barish R, Kamer I,
Soria-Artzi G, Kahani H, et al: Regulation of the inflammatory
profile of stromal cells in human breast cancer: Prominent roles
for TNF-α and the NF-κB pathway. Stem Cell Res Ther. 6:872015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu X, Du L, Feng J, Ling Y and Xu S:
Clinicopathological and prognostic significance of serum cytokine
levels in breast cancer. Clin Lab. 60:1145–1151. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Oh DY, Walenta E, Akiyama TE, Lagakos WS,
Lackey D, Pessentheiner AR, Sasik R, Hah N, Chi TJ, Cox JM, et al:
A Gpr120-selective agonist improves insulin resistance and chronic
inflammation in obese mice. Nat Med. 20:942–947. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu Q, Wang H, Zhao X, Shi Y, Jin M, Wan B,
Xu H, Cheng Y, Ge H and Zhang Y: Identification of
G-protein-coupled receptor 120 as a tumor-promoting receptor that
induces angiogenesis and migration in human colorectal carcinoma.
Oncogene. 32:5541–5550. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chung H, Lee YS, Mayoral R, Oh DY, Siu JT,
Webster NJ, Sears DD, Olefsky JM and Ellies LG: Omega-3 fatty acids
reduce obesity-induced tumor progression independent of GPR120 in a
mouse model of postmenopausal breast cancer. Oncogene.
34:3504–3513. 2015. View Article : Google Scholar : PubMed/NCBI
|