Introduction
Colorectal cancer (CRC) is one of the leading
contributors to cancer-related mortality and morbidity in China
(1). The morphological features and
molecular subtypes of colorectal tumors have been studied
extensively and include microsatellite instability, the CpG island
methylation phenotype and chromosomal instability. Furthermore,
typical molecular patterns have been identified to distinguish
different CRC subtypes derived from distinctive locations. For
example, BRAFV600E occurs more frequently in serrated
CRC, while tubular CRC has an increased rate of the KRAS mutation
(2,3). Moreover, several molecular markers are
also used in targeted therapy. For example, the epidermal growth
factor receptor (EGFR) is a target of panitumumab, a fully human
monoclonal antibody specific to EGFR used to treat EGFR-expressing
metastatic CRC with disease progression (4). Similarly, bevacizumab, which targets
VEGF, is used in metastatic CRC with standard chemotherapy
treatment (5). However, a more
detailed network of the molecular pathways deregulated in CRC is
needed. Studying these molecular alterations can help further
elucidate the mechanisms underlying CRC and improve clinical
care.
The ubiquitin-proteasome system controls protein
homeostasis of almost the entire proteome. Ubiquitin-mediated
proteolysis employs an enzymatic cascade that primarily involves E1
activating enzymes, E2 conjugating enzymes and E3 ubiquitin
ligases. Deregulation of ubiquitin ligase has been shown to play
crucial roles in many pathological activities. Tripartite motif
proteins (TRIMs) represent one of the largest families of E3
ubiquitin ligases. TRIM58, a TRIM protein containing a RING motif,
a B box type 1 and 2, and a coiled-coil domain in its N-terminal
region, was identified as an E3 ubiquitin ligase and was first
shown to regulate terminal erythropoiesis (6). TRIM58 was reported to be involved in a
variety of cancers, including hepatocellular carcinoma and lung
cancer. Hypermethylation of TRIM58 and downregulation of its mRNA
expression were associated with tumorigenesis of hepatocellular
carcinoma and worse prognosis of patients after hepatectomy
(7). Similarly, aberrant
inactivation of TRIM58 caused by CpG island hypermethylation
promoted early carcinogenesis of early-stage lung adenocarcinoma
regardless of smoking status (8).
However, the association between TRIM58 expression and CRC
prognosis and the role of TRIM58 in CRC have not yet been
determined.
Herein, we studied the potential role of TRIM58 in
colorectal carcinogenesis. Instead of measuring gene promoter
methylation, we directly evaluated the expression of TRIM58 and its
clinical significance in a cohort of patients with CRC.
Furthermore, we investigated the effects of TRIM58 on CRC
progression and the underlying molecular mechanisms. Our data
suggest that both mRNA and protein levels of TRIM58 are commonly
suppressed in CRC tissues and cell lines. In vitro studies
showed that ectopic TRIM58 expression specifically influenced cell
invasion, while it had minimal effects on cell proliferation and
migration. In addition, TRIM58 expression modulated the activation
of epithelial-to-mesenchymal transition (EMT) and the matrix
metalloproteinase (MMP) genes. This study indicates that TRIM58 is
a potential biomarker of CRC prognosis; it acts as a tumor
suppressor and has a specific role in the regulation of cancer cell
invasion in CRC.
Materials and methods
Patients and sample collection
This study was approved by the Institutional Review
Board of the Sixth Affiliated Hospital, Sun Yat-sen University. All
samples were collected with the patients' written informed consent
and approval from the Institutional Review Board.
Fresh frozen paired samples (n=48 for mRNA assay
with 30 males vs. 18 females, and n=30 for protein assay containing
25 males vs. 5 females) of primary CRC and adjacent normal colon
tissue (2 cm from the tumor border), ranging from stage I to stage
IV, were collected from the Department of Surgery at the Sixth
Affiliated Hospital of Sun Yat-sen University during the period of
October 2010 to July 2015. Age of all patients with TRIM58 mRNA
detected ranged from 30 to 71 years old, while patients whose
samples were used for TRIM58 protein detection varied from 32 to 88
years old.
Clinical tissue samples were all confirmed
histopathologically and stored in Invitrogen™ RNAlater solution
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) and RIPA lysis
buffer (Thermo Fisher Scientific, Inc.) containing PMSF (Beijing
Dingguo Biotechnology, Co., Ltd., Beijing, China) and PPI (Beijing
Dingguo Biotechnology) at −80°C until extraction.
Cell culture
Eleven human colorectal cell lines, HCT8, KM12,
Caco-2, DLD-1, HCT116, LoVo, HT-29, SW480, SW620, RKO and HCT15,
and 293 cells with SV40-T antigen (293T) were used. Caco-2, DLD-1,
HCT15, HCT116, HT-29, HCT8 and KM12 cells were cultured in
RPMI-1640 medium with 10% fetal bovine serum (FBS). SW480, SW620
and 293T cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) with 10% FBS. RKO cells were cultured in EMEM with 10% FBS,
and LoVo cells were cultured in F-16 with 10% FBS. All cells were
incubated at 37°C with 5% CO2.
Transient transfection and
establishment of stable clone cells
The TRIM58 construct vectors (TRIM58,
pTRIM58-IRES2-EGFP; Empty, H316) or TRIM58 siRNA (siT58#1, GGAG
GGAGCTCTTAAGGAA; siT58#2, AAAUUUCAUUCUA CAAUGUCA) were used to
transiently transfect HCT8 and KM12 cells using Invitrogen
Lipofectamine 3000 transfection reagent (Thermo Fisher Scientific,
Inc.). Briefly, 4 µg of vectors or 5 µl of siRNA was mixed with 4
µl of transfection reagent. The transfection protocol was performed
according to the manufacturer's instructions, and transfection was
confirmed by RT-qPCR and western blot analysis. For stable
transfected cell selection, after 48 h of transfection, the cells
were seeded onto fresh media with 10% FBS (Gibco; Thermo Fisher
Scientific) and 1 mg/ml G418 (Geneticin; Thermo Fisher Scientific,
Inc.). Resistant clones were selected for 7 days and passaged.
Tissue microarray (TMA) construction
and immunohistochemistry (IHC)
A paraffin-embedded tissue microarray and related
clinicopathological parameters were obtained from the Sixth
Affiliated Hospital of Sun Yat-sen University. Paraffin-embedded
tissues were cut into 4-µm thick sections, deparaffinized in
xylene, rehydrated through a graded alcohol series, and heat
treated for 30 min in citrate buffer (pH 6.0; Dako; Agilent
Technologies, Inc., Santa Clara, CA, USA) for antigen retrieval.
Endogenous peroxidase activity was blocked for 10 min with reagent
from an IHC kit (cat. no. SP9000; ZSGB-Bio, Beijing, China).
Samples were then blocked with 5% blocking buffer for 1 h, followed
by incubation overnight with primary anti-TRIM58 antibody
(HPA023637, rabbit anti-human, 1:200; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at 4°C. The next day, the sections were
incubated for 1 h with secondary antibody provided by the kit (cat.
no. SP9000; ZSGB-Bio) after 3 washes with PBST, 10 min per wash,
and then were stained by DAB kit (cat. no. ZLT-9017; ZSGB-Bio).
Finally, the sections were observed under a light microscope
(Olympus BX53; Olympus Corp., Tokyo, Japan). The immunostaining
results were evaluated by two independent pathologists blinded to
the patient status to avoid possible bias.
The staining intensity was scored as: 0 points,
negative; 4 points, weak intensity; 8 points, moderate intensity;
or 12 points, strong intensity. Staining density was scored
separately based on the percentage of cells stained as follows: 0
points, 0%; 4 points, 1–25%; 8 points, 26–50%; 10 points, 51–75%;
or 12 points, >75%. The final score was calculated as the sum of
the intensity score and the density score. Final scores ≥8 points
were considered high expression, and final scores <8 points were
considered low expression.
RNA isolation and RT-qPCR
RNA was extracted from CRC cell lines, human CRC
tissues and normal tissues with Invitrogen TRIzol reagent (Thermo
Fisher Scientific, Inc.). cDNAs were synthesized from extracted RNA
(1 µg/20 µl) with a ReverTra Ace-α RT-PCR kit (Toyobo Co., Ltd.,
Osaka, Japan), and real-time quantitative PCR amplification of
TRIM58 and β-actin was performed with 1 µl of reverse cDNA from CRC
cell lines, human CRC tissues and normal tissues per well in a
LightCycler 488 system (Roche Diagnostics, Indianapolis, IN, USA).
The gene encoding β-actin was used as the reference gene, and the
∆∆Cq method was used to analyze the results (9). The final results are presented as fold
changes in the charts/graphs. [TRIM58 (forward)
5′-GCGGGATCCAGCTTTACAT-3′ and (reverse) 5′-GGCTGGAAGCAGAGAACATC-3′;
β-actin (forward) 5′-TTGTTACAGGAAGTCCCTTGCC-3′ and (reverse) 5′-
ATGCTATCACCTCCCCTGTGTG-3′. The thermocycling conditions were as
follows: precubation at 95°C for 10 min; amplification at 95°C for
10 sec, 60°C for 10 sec, 72°C for 10 sec, 45 cycles; melting at
95°C for 10 sec, 65°C for 60 sec and 97°C for 1 sec.
Western blotting
Cells were washed twice with PBS and lysed with RIPA
lysis buffer (Thermo Fisher Scientific, Inc.) containing PMSF
(Beijing Dingguo Biotechnology) and PPI (Beijing Dingguo
Biotechnology). The protein concentration was determined with a
bicinchoninic acid (BCA) protein assay kit (Beijing Dingguo
Biotechnology). Equal amounts of protein (~20 µg) were separated
using 8, 10 or 12% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gels, depending on the molecular weight
of the protein, and were then transferred to 0.22-µm polyvinylidene
difluoride filter (PVDF) membranes. After that, the transferred
membranes were blocked in TBST buffer containing 5% non-fat milk or
bovine serum albumin (BSA) for 1 h and then incubated overnight at
4°C with primary antibodies. Next, the membranes were washed three
times with TBST for at least 30 min and probed with HRP-linked
secondary antibodies for 1.5 h at room temperature. Protein bands
were visualized with a chemiluminescence detection kit (Thermo
Fisher Scientific, Inc.) after three washes with TBST buffer and
semi-quantified analysis was carried out using ImageJ software
(National Institutes of Health, Bethesda, MD, USA).
The primary antibodies and concentrations used were
as follows: anti-TRIM58 (dilution 1:1,000; cat. no. H6-NBP1-88608;
Novus Biologicals LLC, Littleton, CO, USA) and anti-GAPDH (dilution
1:1,000; cat. no. 60004–1-Ig; Proteintech Group, Rosemont, IL,
USA). The following primary rabbit anti-human polyclonal antibodies
were purchased from Cell Signaling Technology (Danvers, MA, USA):
anti-β-catenin (dilution 1:1,000; cat. no. 8480), anti-E-cadherin
(dilution 1:1,000; cat. no. 3195), anti-N-cadherin (dilution 1:500;
cat. no. 13116), anti-Snail (dilution 1:500; cat. no. 3879),
anti-Slug (dilution 1:500; cat. no. 9585), anti-MMP-2 (dilution
1:250; cat. no. 13132S) and anti-MMP-9 (dilution 1:250; cat. no.
13667S). The secondary antibodies were anti-rabbit-IgG-HRP and
anti-mouse-IgG-HRP (dilution 1:10,000; cat. nos. 7074S and
7076).
Proliferation assay using an IncuCyte
imaging system
First, 2×104 cells were seeded in 96-well
plates with RPMI-1640 plus 10% FBS. The plate was then placed into
and incubated in an IncuCyte Essens Bioscience incubator (Essens
Bioscience, Birmingham, UK). Live cell images were collected every
4 h. Proliferation rates based on cell confluence were determined
by live cell imaging using IncuCyte software (Essens
Bioscience).
Colony formation assay
First, 2,000 cells were seeded in 6-well plates with
RPMI-1640 plus 10% FBS and incubated at 37°C with 5% CO2
for 7 to 10 days. After colony formation, cells were fixed with 4%
PFA at room temperature for 30 min and stained with 0.01% crystal
violet solution for 1 min. Data were collected after the plate was
completely dry.
Wound-healing assays with the IncuCyte
time-lapse imaging system
After transfection with the TRIM58 vector or TRIM58
siRNA, cells were seeded in 6-well plates and incubated until 100%
confluence was reached. The cell layer was scratched with a 10-µl
pipette tip (Rainin; Mettler-Toledo AG, Greifensee, Switzerland)
and washed with PBS three times. The plate was placed in the
IncuCyte Essens Bioscience incubator (Essens Bioscience) and
cultured in fresh serum-free DMEM containing the TRIM58 vector or
TRIM58 siRNA transfection mix for 24–72 h. Live cell images were
collected every 4 h. Migration distance was calculated using ImageJ
software (National Institutes of Health, Bethesda, MD, USA).
Invasion assay
Cell invasion was evaluated with 8-µm Transwell
filters (Corning Inc., Corning, NY, USA). The upper chambers of the
Transwell filters were coated with Matrigel (Corning Inc.) and
incubated at 37°C for at least 30 min. Then, 4×104 cells
were seeded on the Matrigel-coated filter with RPMI-1640 plus 1%
FBS, and RPMI-1640 containing 10% FBS was added to the lower
chamber. After 24 or 36 h, non-migrating cells were carefully
removed from the upper chamber with a cotton swab, and the filters
were fixed and stained with 0.01% crystal violet solution for 1
min. Invading cells were photographed, and the invaded cell
proportion was determined by measuring the OD570 nm value of the
crystal violet eluted with ethanol.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 6.0 (GraphPad Software, Inc., La Jolla, CA, USA). All data
are presented as the mean ± SEM. Comparisons to determine
significant differences between two experimental groups were made
using two-tailed t-tests or Chi-square test, and one-way ANOVA with
Dunnett's post hoc test or Bonferroni post hoc test was conducted
to determine significant differences among three or more groups.
Potential risk factors of OS were evaluated by univariate analysis
and multivariate analysis using Cox proportional hazards model.
Kaplan-Meier survival curves were determined by log-rank test.
Significance was set at P<0.05.
Validation of TRIM58 expression based
on the Oncomine database
We utilized the Oncomine database (http://www.oncomine.org) to validate the expression of
TRIM58 in patients. The differential expression analysis was
directly performed using Oncomine online analysis tools (10).
Results
Trim58 expression is suppressed in CRC
tissues and cell lines
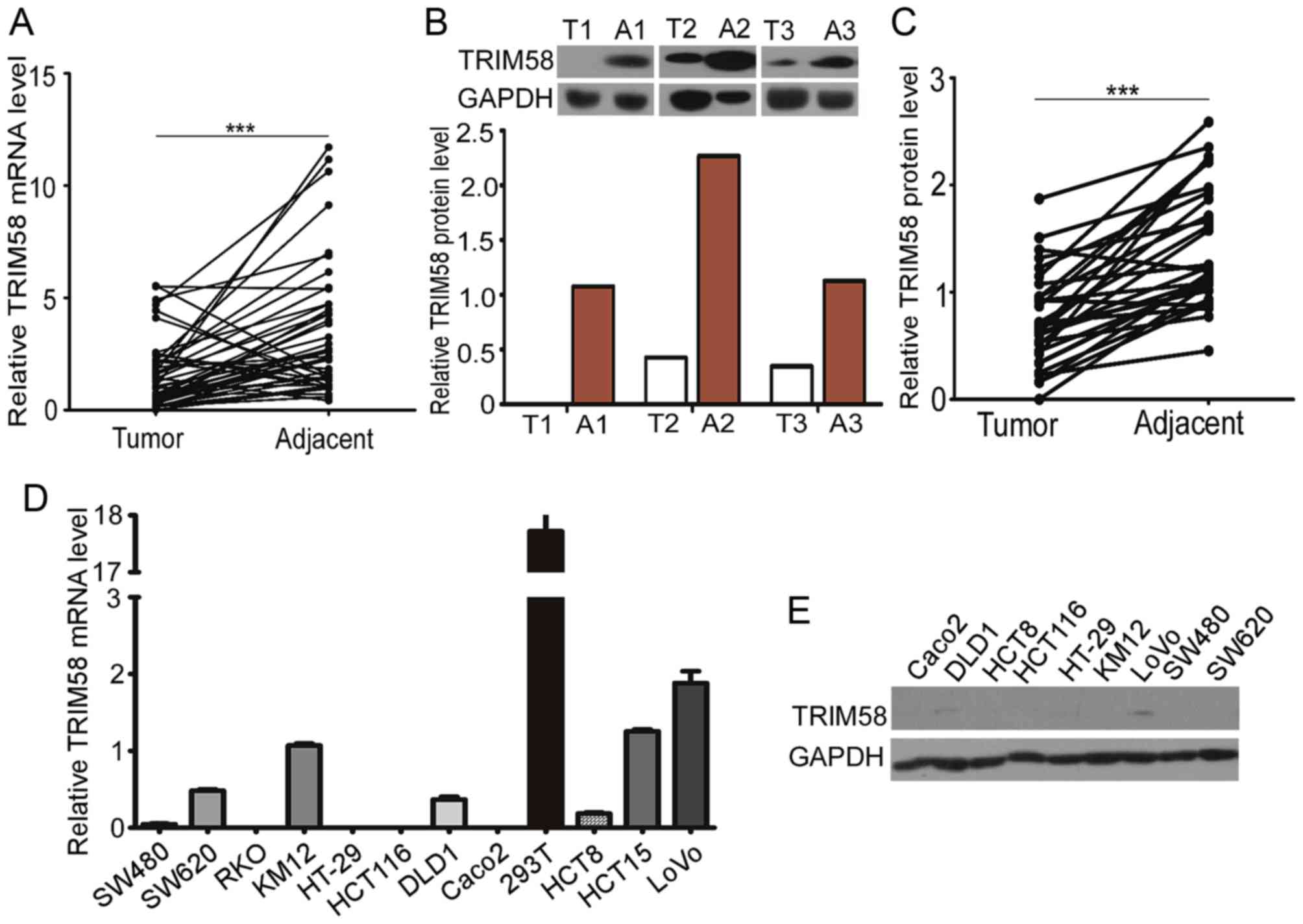
To examine TRIM58 expression in clinical CRC
samples, we evaluated the TRIM58 mRNA and protein levels in 48
tumor tissues with matched adjacent normal samples from CRC
patients. Trim58 expression was significantly lower in human CRC
than in non-tumorous tissues (Fig.
1A). Western blot analysis further confirmed that expression of
TRIM58 protein was lower in 30 pairs of human CRC tissues than the
level in the matched adjacent normal tissues (Fig. 1B and C).
Having shown downregulation of TRIM58 in human CRC
tissues, we further investigated the expression of TRIM58 in human
CRC cell lines. We examined the mRNA levels of TRIM58 in 11 human
CRC cell lines and protein levels in 9 of the cell lines (Fig. 1D and E). The TRIM58 mRNA levels in
CRC cell lines were generally low compared with that in the 293T
cell line, and TRIM58 protein was expressed at low levels in most
of the CRC cell lines examined. Taken together, these results
indicate that TRIM58 expression was suppressed in both CRC tissues
and cell lines.
Low expression of TRIM58 is correlated
with poor prognosis in CRC patients
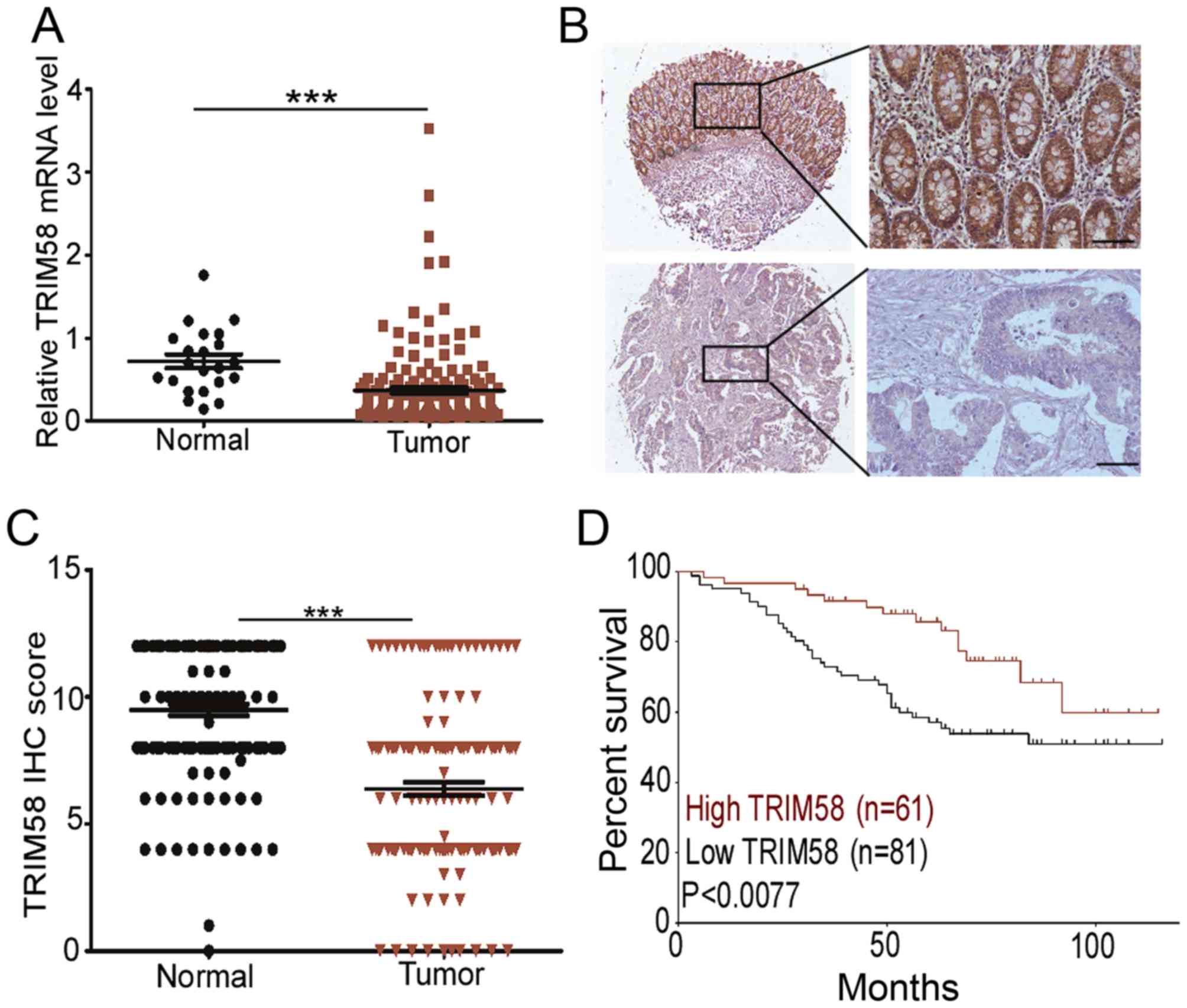
To determine the clinical implications of TRIM58 in
CRC, we evaluated the relationship between TRIM58 expression level
and pathological features of the disease. First, we analyzed TRIM58
mRNA expression with the Oncomine database using RNA sequencing
data of CRC patients from The Cancer Genome Atlas (TCGA). The
results revealed that the TRIM58 mRNA level was lower in cancer
tissues than in normal tissues (Fig.
2A). To further validate the suppressed expression of TRIM58 in
CRC, we recruited a cohort of 152 patients for TMA analysis. The
IHC staining confirmed that TRIM58 was primarily localized in the
cytoplasm (Fig 2B) in the tissues.
IHC scores of TRIM58 revealed that in 68.57% (96/140) of the paired
samples, TRIM58 expression was downregulated in CRC compared to
normal tissues (Fig. 2C)
Since TRIM58 expression was significantly decreased
in CRC, we next investigated whether TRIM58 expression was
associated with the clinical characteristics of CRC patients.
Kaplan-Meier analysis showed that a low level of TRIM58 was
correlated with the poor overall survival in the cohorts
(P<0.0077, Fig. 2D).
Clinicopathological characteristic analysis revealed that low
expression was positively correlated with various clinical stages
in the cohort (P<0.05, Table I),
in particular T status (P<0.004, Table I). Furthermore, univariate Cox
regression analysis revealed that TRIM58 expression was an
independent prognostic factor for poor survival (P<0.05,
Table II).
 | Table I.Association between expression of
TRIM58 and clinicopathological characteristics of the CRC
patients. |
Table I.
Association between expression of
TRIM58 and clinicopathological characteristics of the CRC
patients.
|
| TRIM58 |
|
|---|
| Variables | Low expression n
(%) | High expression n
(%) | P-value |
|---|
| Sex |
|
| 0.803 |
| Male | 44 (54.3) | 40 (56.3) |
|
|
Female | 37 (45.7) | 31 (43.7) |
|
| Median age
(years) |
|
| 0.324 |
|
<60 | 43 (53.1) | 32 (45.1) |
|
| ≥60 | 38 (46.9) | 39 (54.9) |
|
| Histological
grade |
|
| 0.428 |
| G1 | 5 (8.2) | 5 (11.6) |
|
| G2 | 55 (90.2) | 38 (88.4) |
|
| G3 | 1 (1.6) | 0 (0.00) |
|
| pT status |
|
| 0.004b |
| T1 | 2 (2.5) | 3 (4.2) |
|
| T2 | 9 (11.1) | 18 (25.4) |
|
| T3 | 63 (77.8) | 49 (69.0) |
|
| T4 | 7 (8.6) | 1 (1.4) |
|
| pN status |
|
| 0.055 |
| N0 | 41 (51.2) | 50 (70.4) |
|
| N1 | 31 (38.8) | 17 (23.9) |
|
| N2 | 8 (10%) | 4 (5.6%) |
|
| pM status |
|
| 0.556 |
| M0 | 77 (96.3) | 69 (97.2) |
|
| M1 | 3 (3.8) | 2 (2.8) |
|
| Clinical stage
(Duke's) |
|
| 0.025a |
| I | 10 (12.8) | 16 (22.5) |
|
| II | 30 (38.5) | 33 (46.5) |
|
|
III | 38 (48.7) | 21 (29.6) |
|
| IV | 0 (0.0) | 1 (1.4) |
|
| Neo chemotherapy
(%) |
|
| 0.17 |
| No | 6 (7.4) | 1(1.4) |
|
|
Yes | 75 (92.6) | 70 (98.6) |
|
|
Recurrence-metastasis |
|
| 0.025a |
| No | 40 (58.0) | 46 (76.7) |
|
|
Yes | 29 (42.0) | 14 (23.3) |
|
| Table II.Univariate and multivariate analyses
of different prognostic parameters of the CRC patients in the
testing cohort (n=152). |
Table II.
Univariate and multivariate analyses
of different prognostic parameters of the CRC patients in the
testing cohort (n=152).
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Parameters | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Sex (male vs.
female) | 1.1 (0.6–1.8) | 0.781 | 1.1 (0.6–2.0) | 0.689 |
| Age (<60 vs. ≥60
years) | 1.1 (0.6–1.8) | 0.759 | 1.2 (0.7–2.1) | 0.421 |
| Neo-chemotherapy
(no vs. yes) | 2.4 (1.0–6.0) | 0.06 | 1.1 (0.3–3.8) | 0.831 |
| Histological grade
(G1 or G2 vs. G3 or G4) | 0.4 (0.2–0.6) | <0.001 | 0.5 (0.9–2.2) | 0.335 |
| pT status (T1 or T2
vs. T3 or T4) | 0.2 (0.05–0.5) | 0.002 | 0.2 (0.6–0.7) | 0.012 |
| pN status (N0 vs.
N1or N2) | 0.3 (0.2–0.6) | <0.001 | 1.0 (0.2–4.7) | 0.951 |
| pM status (M0 vs.
M1) | 0.2 (0.09–0.6) | 0.001 | 0.3 (0.1–0.9) | 0.024 |
| TRIM58 expression
(low vs. high) | 1.8 (1.0–3.0) | 0.044a | 1.2 (0.7–2.2) | 0.562 |
In conclusion, these results indicated that low
expression of TRIM58 is correlated with the poor prognosis of CRC
patients, indicating that TRIM58 expression may play an important
role in the tumorigenesis of CRC.
TRIM58 overexpression has no
significant influence on the proliferation, colonization or
migration of human CRC cell lines
Our results clearly indicated that high TRIM58
expression was associated with the TNM stage, suggesting the role
of TRIM58 in cell proliferation, cell colonization and cell
motility. Therefore, to identify the specific roles in which TRIM58
modulates tumorigenesis, we randomly selected two CRC cell lines
(HCT8 and KM12) with low expression of TRIM58 (Fig. 1D and E) and transfected the cells
with a TRIM58 overexpression vector (TRIM58, pTRIM58-IRES2-EGFP) or
an empty vector as a negative control (Empty, H316). The efficacy
of TRIM58 overexpression was examined by RT-PCR and western blot
analyses. The results showed that HCT8 and KM12 cells efficiently
overexpressed TRIM58 (Fig.
3A-C).
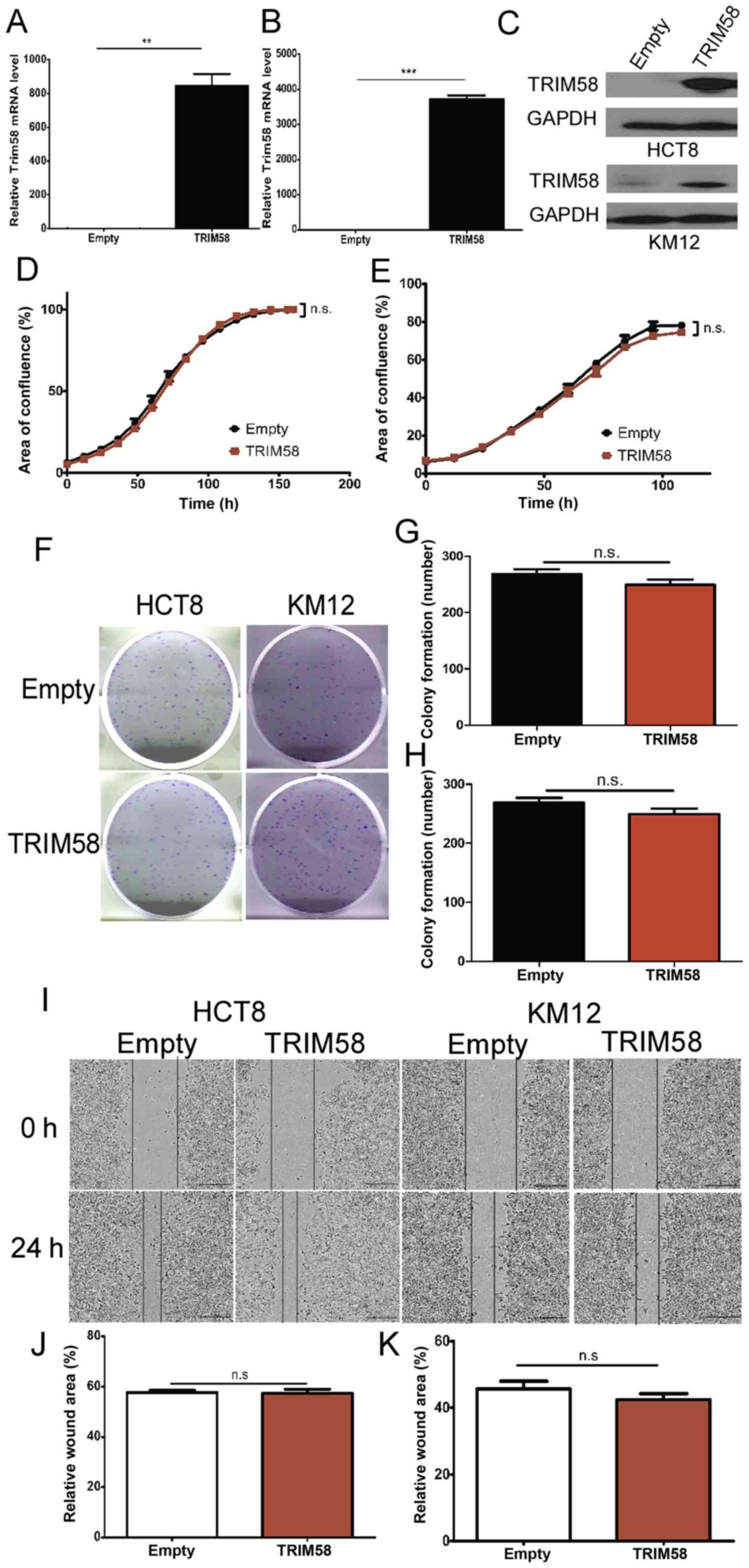 | Figure 3.TRIM58 overexpression has no
significant influence on the proliferation, colonization or
migration of human colorectal cancer cell lines. (A-C) The
overexpression of TRIM58 mRNA (A: HCT8 cells and B: KM12 cells) and
(C) protein in CRC cell lines was confirmed by RT-PCR and western
blot analysis.**: n=3, P<0.001 and ***: n=3, P<0.0001,
two-tailed Student's t-test. (D and E) Proliferation assay (D: HCT8
cells and E: KM12 cells) conducted using IncuCyte. Cells were
transfected with the TRIM58 overexpression vector on day 1 and day
3. n.s. indicates not significant, n=3, P>0.05, two-tailed
Student's t-test. (F-H) Images (F) and statistical analysis (G:
HCT8 cells and H: KM12 cells) of colony formation assays indicated
that TRIM58 overexpression had no significant influence on cell
colonization. n.s. indicates not significant, n=3, P>0.05,
two-tailed Student's t-test. (I-K) Images of wound-healing assays
(I) and statistical analysis (J: HCT8 cells and K: KM12 cells) were
performed to show that TRIM58 did not inhibit cell migration. n.s.
indicates not significant, n=3, P>0.05, two-tailed Student's
t-test. Scale bar, 100 µm. TRIM, tripartite motif protein; CRC,
colorectal cancer. |
Next, we determined whether TRIM58 overexpression
inhibits CRC cell proliferation. To this end, cellular
proliferation was assessed using IncuCyte time-lapse imaging. We
observed that TRIM58 overexpression barely inhibited the
proliferation of HCT8 and KM12 cells during the observation time
(Fig. 3D and E). To further
investigate the role of TRIM58 in CRC progression, we analyzed the
cell colony formation ability of HCT8 and KM12 cells overexpressing
TRIM58. Our results demonstrated that TRIM58 overexpression had
little influence on CRC cell colonization (Fig. 3F-H). Similarly, wound-healing assays
showed that TRIM58 overexpression did not affect the migration of
the HCT8 and KM12 cells (Fig.
3I-K). These data suggested that TRIM58 inhibits tumor
progression via other processes.
Overexpression of TRIM58 specifically
reduces invasion of CRC cells and inhibits the expression of
related genes
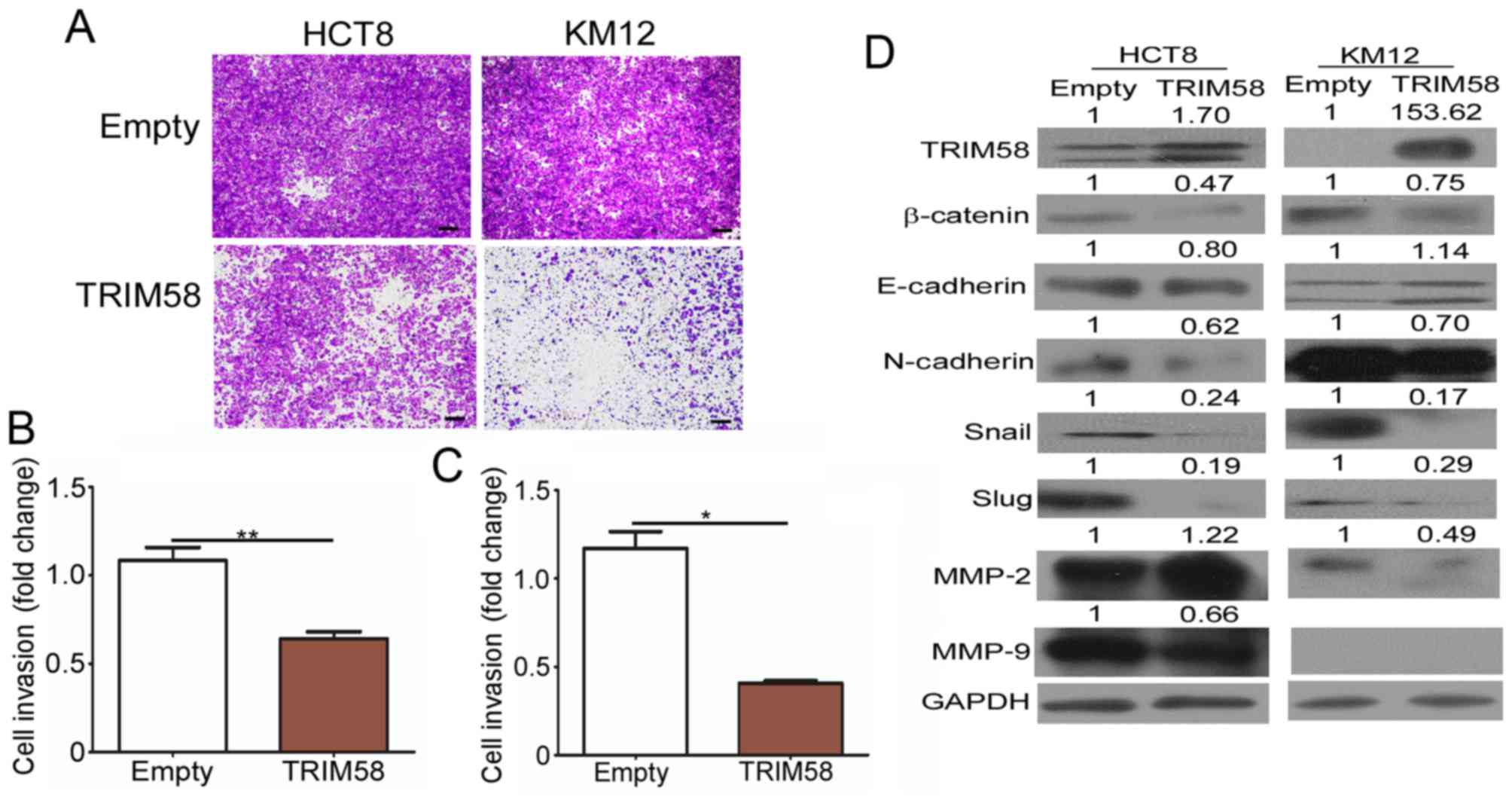
Evidence has shown that local and distant invasion
is highly associated with cancer malignancy (11). We thus conducted invasion assays to
determine whether high TRIM58 expression negatively impacts CRC
development by interfering with the invasion of CRC cells. As shown
in Fig. 4A-C, a significant
reduction in the number of invading HCT8 and KM12 CRC cells was
observed following transient overexpression of TRIM58 in these
cells in the invasion assays. Since the EMT program is a prominent
regulator of invasion, and MMPs have been shown to play an active
role in tumor invasion and metastasis (12), we detected the TRIM58-mediated
changes in EMT proteins and MMP expression (Fig. 4D). Western blot results showed that
TRIM58 overexpression downregulated the levels of most mesenchymal
proteins, including N-cadherin, Snail and Slug, in both HCT8 and
KM12 cells, while it upregulated the epithelial protein E-cadherin
in KM12 cells. Furthermore, β-catenin, a key regulator of EMT, was
downregulated when TRIM58 was overexpressed in both tested cell
lines. Moreover, we also determined the MMP2 and MMP9 levels and
found that the MMP2 level was decreased in KM12 cells and MMP9 was
reduced in the HCT8 cells under TRIM58 overexpression. The above
results suggested that TRIM58 plays an important role in inhibiting
the invasive ability of CRC cells during cancer progression and
might be regulated by Wnt-β-catenin-mediated regulation of EMT
signaling and MMP expression.
Silencing of TRIM58 overexpression
reverses CRC cell invasion and related protein levels
To further determine the effect of TRIM58 on cell
invasion, we neutralized TRIM58 overexpression by introducing
siRNAs into stable TRIM58-overexpressing cells. First, we tested
the efficiency of the TRIM58 siRNAs by RT-PCR and western blotting.
The results (Fig. 5A and C) showed
that siT58#1 and siT58#2 effectively reduced the mRNA and protein
levels of TRIM58 in SW620 cells. We then mixed siT58#1 and siT58#2
and transfected the mixture into the TRIM58-overexpressing HCT8
cells (termed siTRIM58), while HCT8 cells were transfected with
empty vector and siCtrl as the control (termed Ctrl), and stable
TRIM58-expressing HCT8 cells were transfected with siCtrl (termed
TRIM58) (Fig. 5B and D). The
results showed that the invasive ability recovered and was slightly
stronger than that in the Ctrl-HCT8 cells after knocking down
TRIM58 in TRIM58-overexpressing HCT8 cells (siTRIM58) compared to
TRIM58-overexpressing HCT8 cells transfected with the control siRNA
(TRIM58) (Fig. 5E and F).
Additionally, expression of mesenchymal proteins (N-cadherin,
Snail, Slug), MMP2 and MMP9 were prominently increased and
E-cadherin was reduced after silencing of TRIM58 in HCT8
overexpression cells, which was similar to the results in the Ctrl
HCT8 cells (Fig. 5G). The above
results further demonstrated that TRIM58 induced CRC cell
progression by enhancing the invasion modulated by EMT signaling
and MMPs.
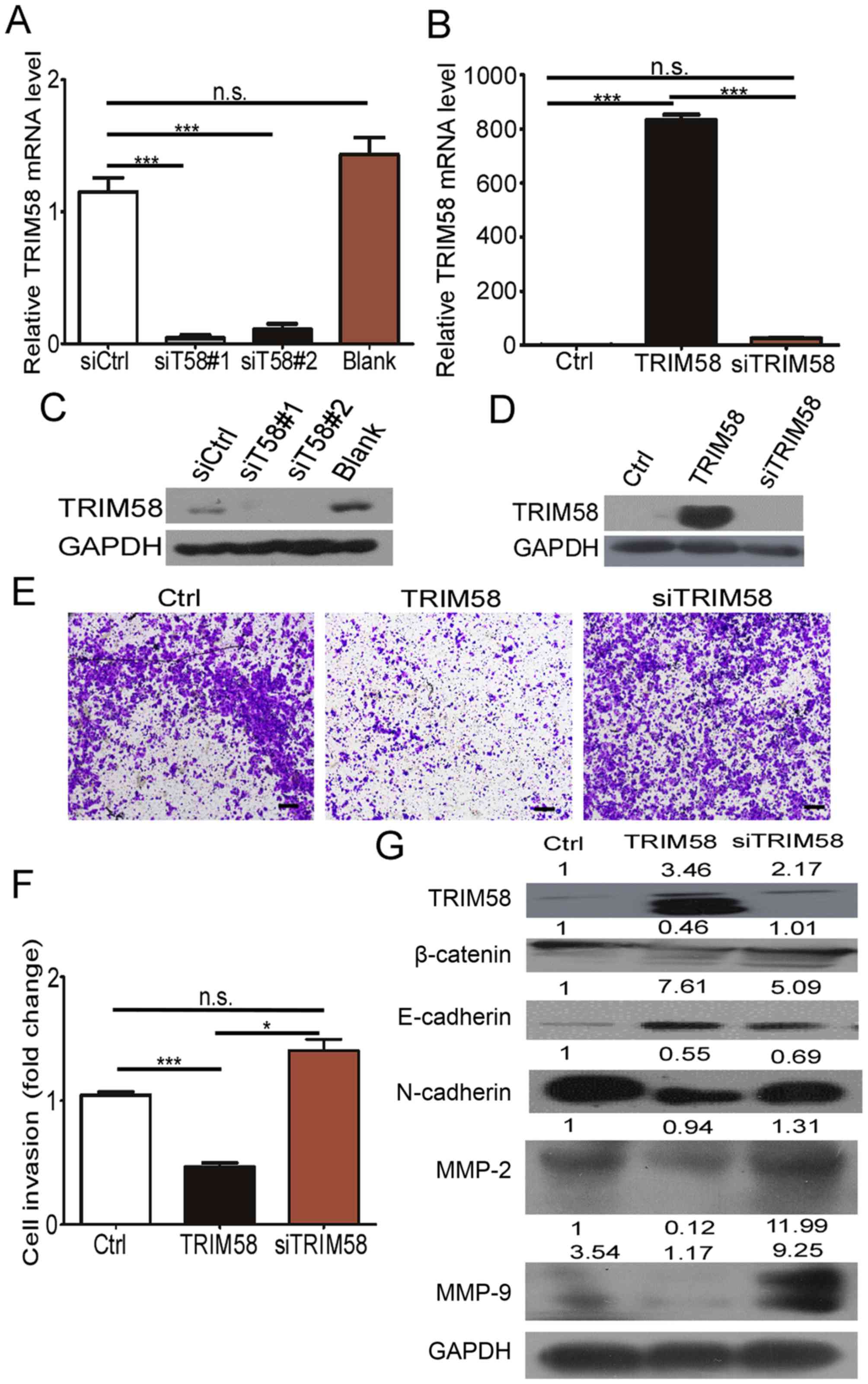 | Figure 5.Silencing of TRIM58 overexpression
reversed invasive ability in the CRC cells and related protein
level. (A and C) siRNA silencing of TRIM58 mRNA and protein in CRC
cell lines was confirmed by RT-PCR and western blot analysis.
Blank, cells treated with ddH2O as a blank control. n.s.
indicates not significant, n=3, P>0.05; ***: n=3, P<0.0001,
one-way ANOVA with Dunnett's post hoc test (A). (B and D) Knockdown
of TRIM58 mRNA and protein in stable TRIM58-overexpressing CRC cell
lines was confirmed by RT-PCR and western blot analysis. n.s.
indicates not significant, n=3, P>0.05; ***: n=3, P<0.0001,
one-way ANOVA with Bonferroni post hoc test (B). (E) Invasion
assays indicated that the cell invasion inhibition of TRIM58
overexpression was neutralized after introducing TRIM58 siRNA into
the stable TRIM58-overexpressing HCT8 cells, and the cell invasion
was similar to that in Ctrl cells with empty vector that were
transfected with siCtrl. All the results were collected after 24 h.
Scale bar, 100 µm. (F) Statistical analysis of the number of
invasive HCT8 cells. The number of invasive HCT8 cells treated with
empty or TIRIM58 expression vectors was quantified. **: n=3,
P<0.01, and *: n=3, P<0.05, one-way ANOVA with Bonferroni
post hoc test. (G) Western blot analysis and semi-quantative
statistical results for EMT-related proteins and MMPs. n.s.
indicates not significant, n=3, P>0.05. TRIM, tripartite motif
protein; CRC, colorectal cancer; MMPs, matrix
metalloproteinases. |
Discussion
Discovery of potential biomarkers for early tumor
detection is now a major concern of researchers. Various aberrant
genes have been reported and translated into clinical diagnostic
applications.
The TRIM family is a large protein family comprising
more than 70 members. The majority of TRIM proteins are believed to
be E3 ubiquitin ligases because of their RING domain, which
frequently displays E3 ligase activity. Indeed, several TRIM
proteins have been shown to modify target proteins with
ubiquitination and to be associated with many biological processes,
and abnormal changes in their abundance or activity are commonly
involved in pathological conditions, including cancer (13,14).
Accumulated reports have shown that TRIM proteins participate in
tumorigenesis and progression of breast cancer, liver cancer,
gastric cancer, osteosarcoma and skin cancer (15–18).
For example, TRIM31 indirectly overactivates the mTORC pathway by
ubiquitinating the TSC1-TSC2 complex to promote hepatocellular
carcinoma progression (15).
However, the pro-tumorigenesis TRIM family has
primarily been studied for their oncogenic functions, and less
attention has been paid to tumor-suppressor gene functions. TRIM58
was recently shown to act as a tumor-suppressor gene and to play a
crucial role in the tumorigenesis of different cancers (7,8).
Accordingly, our data also indicated that TRIM58 downregulation in
CRC was significantly correlated with poor clinicopathological
characteristics, especially T status. We therefore hypothesized
that TRIM58 might be a tumor suppressor whose aberrant expression
plays a crucial role in CRC tumorigenesis and progression. To
confirm this hypothesis, we performed a series of in vitro
assays, including proliferation, colony formation, migration and
invasion assays, to investigate the role of TRIM58 in CRC cell
growth and motility, which are the crucial factors leading to tumor
malignancy. Surprisingly, our results showed that TRIM58
overexpression only inhibited cell invasion, which is likely
associated with EMT and MMP regulation, but not other malignant
behaviors, such as cell proliferation, colony formation and
migration.
These results not only support our view that TRIM58
may function as a tumor suppressor in CRC, expanding upon its
previously reported role in other cancers, but also, interestingly,
demonstrated a distinct function of TRIM58. Aberrant TRIM58
expression only influenced the CRC cell invasion capacity, in
contrast with the results of other studies showing that TRIM58
suppression enhances proliferation, colony formation, migration and
invasion in lung and liver tumors. The functional differences among
these tumor types is likely due to phenotypic distinctions and
genomic pattern variations in tumors derived from diverse origins
(19,20). Furthermore, our results revealed
that although TRIM58 overexpression inhibited CRC cell invasion, it
had little effect on CRC cell motility in the wound healing assays.
This is likely because cell motility or directional movement is one
of several invasion processes that TRIM58 has little or no effect
on in the CRC cells used in this study (21). We also tested the expression of EMT
genes that have been confirmed as important cell invasion
regulators. Consistent with the TRIM58-induced changes in cell
invasion, Snail and Slug, the key transcriptional genes that
control cell-cell adhesion, cell shape, degradation of E-cadherin
protein and basement membrane integrity during EMT, were
downregulated when TRIM58 was overexpressed. Furthermore, the
expression level of the mesenchymal protein N-cadherin was reduced
when TRIM58 was overexpressed and rescued after silencing of TRIM58
by siRNA, which is opposite to the trend observed for E-cadherin,
an epithelial cell gene involved in EMT. These findings indicate
that TRIM58 regulates EMT and thus influences CRC invasion.
Further investigation indicated that the expression
of β-catenin, a crucial downstream effector of canonical
Wnt-β-catenin signaling during EMT (16), is negatively regulated by TRIM58 and
that TRIM58 might affect EMT by modulating Wnt-β-catenin signaling.
However, further studies are needed.
Collectively, we suggest that TRIM58 is a tumor
suppressor that inhibits tumorigenesis and progression by
regulating EMT via Wnt-β-catenin signaling, and TRIM58 suppression
may be a marker for early CRC detection.
Acknowledgements
We thank for the support by the National Key
Clinical Discipline.
Funding
This work was financially supported by the National
Natural Science Foundation (grant no. 81573078 to LW), the Natural
Science Foundation of Guangdong Province (grant no. 2017A030313805
to DC and 2016A030311021 to LW), the Science and Technology Program
of Guangzhou City (grant no. 201604020174 to DC), and the Science
and Technology Planning Project of Tianhe District of Guangzhou
(201504KW044 to HW).
Availability of data and materials
The datasets and materials used during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
DC conceived the project, designed the experiments
and edited the manuscript. LW edited the manuscript and provided
overall support. XZ performed the experiments, analyzed the
results, conducted statistical analysis of data and wrote the
manuscript. HW assessed the IHC microarray scores and performed
RT-qPCR and western blotting of the tissue samples. ML collected
the clinical samples, counted the HCT8 and KM12 cell colonies,
assessed the IHC microarray scores and edited the manuscript. JC
provided the tissue microarray. YL, QL and ZY helped culture cells
and in vitro assay. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The study was approved by the Human Medical Ethics
Committee of Sun Yat-Sen University (SYSU). Informed consent was
obtained from all patients enrolled in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflicts of
interest to disclose.
Glossary
Abbreviations
Abbreviations:
|
TRIM
|
tripartite motif protein
|
|
CRC
|
colorectal cancer
|
|
EMT
|
epithelial-mesenchymal transition
|
|
OS
|
overall survival
|
|
MMP
|
matrix metalloproteinase
|
|
TCGA
|
The Cancer Genome Atlas
|
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fessler E and Medema JP: Colorectal cancer
subtypes: Developmental origin and microenvironmental regulation.
Trends Cancer. 2:505–518. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shia J, Schultz N, Kuk D, Vakiani E,
Middha S, Segal NH, Hechtman JF, Berger MF, Stadler ZK, Weiser MR,
et al: Morphological characterization of colorectal cancers in The
Cancer Genome Atlas reveals distinct morphology-molecular
associations: Clinical and biological implications. Mod Pathol.
30:599–609. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gibson TB, Ranganathan A and Grothey A:
Randomized phase III trial results of panitumumab, a fully human
anti-epidermal growth factor receptor monoclonal antibody, in
metastatic colorectal cancer. Clin Colorectal Cancer. 6:29–31.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Los M, Roodhart JML and Voest EE: Target
practice: Lessons from phase III trials with bevacizumab and
vatalanib in the treatment of advanced colorectal cancer.
Oncologist. 12:443–450. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thom CS, Traxler EA, Khandros E, Nickas
JM, Zhou OY, Lazarus JE, Silva AP, Prabhu D, Yao Y, Aribeana C, et
al: Trim58 degrades Dynein and regulates terminal erythropoiesis.
Dev Cell. 30:688–700. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qiu X, Huang Y, Zhou Y and Zheng F:
Aberrant methylation of TRIM58 in hepatocellular carcinoma and its
potential clinical implication. Oncol Rep. 36:811–818. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kajiura K, Masuda K, Naruto T, Kohmoto T,
Watabnabe M, Tsuboi M, Takizawa H, Kondo K, Tangoku A and Imoto I:
Frequent silencing of the candidate tumor suppressor TRIM58 by
promoter methylation in early-stage lung adenocarcinoma.
Oncotarget. 8:2890–2905. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rhodes DR, Kalyana-Sundaram S, Mahavisno
V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ,
Kincead-Beal C, Kulkarni P, et al: Oncomine 3.0: Genes, pathways,
and networks in a collection of 18,000 cancer gene expression
profiles. Neoplasia. 9:166–180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dufour A and Overall CM: Missing the
target: Matrix metalloproteinase antitargets in inflammation and
cancer. Trends Pharmacol Sci. 34:233–242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meroni G and Diez-Roux G: TRIM/RBCC, a
novel class of ‘single protein RING finger’ E3 ubiquitin ligases.
BioEssays. 27:1147–1157. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Elabd S, Meroni G and Blattner C: TRIMming
p53's anticancer activity. Oncogene. 35:5577–5584. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo P, Ma X, Zhao W, Huai W, Li T, Qiu Y,
Zhang Y and Han L: TRIM31 is upregulated in hepatocellular
carcinoma and promotes disease progression by inducing
ubiquitination of TSC1-TSC2 complex. Oncogene. 37:478–488. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawabata H, Azuma K, Ikeda K, Sugitani I,
Kinowaki K, Fujii T, Osaki A, Saeki T, Horie-Inoue K and Inoue S:
TRIM44 is a poor prognostic factor for breast cancer patients as a
modulator of NF-κB signaling. Int J Mol Sci. 18:E19312017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu G, Guo Y, Xu D, Wang Y, Shen Y, Wang F,
Lv Y, Song F, Jiang D, Zhang Y, et al: TRIM14 regulates cell
proliferation and invasion in osteosarcoma via promotion of the AKT
signaling pathway. Sci Rep. 7:424112017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Horn EJ, Albor A, Liu Y, El-Hizawi S,
Vanderbeek GE, Babcock M, Bowden GT, Hennings H, Lozano G, Weinberg
WC, et al: RING protein Trim32 associated with skin carcinogenesis
has anti-apoptotic and E3-ubiquitin ligase properties.
Carcinogenesis. 25:157–167. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zehir A, Benayed R, Shah RH, Syed A,
Middha S, Kim HR, Srinivasan P, Gao J, Chakravarty D, Devlin SM, et
al: Mutational landscape of metastatic cancer revealed from
prospective clinical sequencing of 10,000 patients. Nat Med.
23:703–713. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hause RJ, Pritchard CC, Shendure J and
Salipante SJ: Classification and characterization of microsatellite
instability across 18 cancer types. Nat Med. 22:1342–1350. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Friedl P and Alexander S: Cancer invasion
and the microenvironment: Plasticity and reciprocity. Cell.
147:992–1009. 2011. View Article : Google Scholar : PubMed/NCBI
|