Introduction
Gastric cancer (GC) is the second most common
malignant tumor in terms of morbidity and mortality. There are
900,000 new cases of GC and 700,000-related deaths worldwide each
year (1). However, no biomarkers
for GC are routinely used to predict its occurrence and
prognosis.
Homeobox A5 (HOXA5) is located on chromosome
7, has a total length of 2,319 bp and is a member of the HOX
family. Its encoded protein, HOXA5, contains 403 amino acids. HOXA5
is a transcription factor that is involved in regulating human
embryonic development and adult stem cell differentiation (2,3). The
role of HOXA5 in cancer progression has been gradually clarified.
It has been demonstrated that HOXA5 inhibits the activity of the
Wingless (Wnt) pathway through the regulation of β-catenin
inhibitory proteins and forms a negative feedback loop with Wnt
signals to inhibit the stemness characteristics of colorectal
cancer cells (4). Retinoic acid
(RA) is a promising anticancer drug, and it has been found that the
enforced overexpression of HOXA5 induces the apoptosis and
differentiation of breast cancer cells. Further mechanistic
research has revealed that the RA response element upstream of the
HOXA5 promoter combines with RA receptor β (RARβ), thus
mediating the anticancer effects of RA (5). F-actin reorganization and filopodia
formation are known to promote cell movement, and HOXA5 in lung
cancer cells has been shown to decrease the level of myosin-X
proteins expressed on filopodia tips and to reduce the number of
pseudopods, thereby preventing cancer cell metastases. Survival
analysis has indicated that HOXA5 expression is associated with a
better overall and disease-free survival of patients with non-small
cell lung cancer expressing wild-type EGFR (6). These studies have suggested that
HOXA5 may be a broad-spectrum tumor suppressor gene with the
potential for wide-ranging significance and applications.
A recent study found that HOXA5 was
significantly differentially methylated between GC tissues and
adjacent non-cancer tissues (7).
However, to the best of our knowledge, no information is available
to date regarding the expression of HOXA5 and its function in human
GC. Thus, in this study, in order to explore the vital role of
HOXA5 in the tumorigenesis and progression of GC, the expression of
HOXA5 in 81 patients with GC was detected by immunohistochemistry,
and the association between HOXA5 expression and the
clinicopathological characteristics of the patients with GC was
analyzed. Additionally, the potential value of HOXA5 as a
prognostic indicator was assessed in patients with GC. Finally, the
effects of HOXA5 on cell proliferation were investigated in
vitro and in vivo.
Materials and methods
Patients and specimens
A total of 81 pairs of GC tissues and matched
paracancerous tissues (≥6 cm away from the tumor) for
immunohistochemistry and reverse transcription-quantitative PCR
(RT-qPCR) were collected from patients with GC after surgical
resection at the First Affiliated Hospital of Chongqing Medical
University from January to October, 2009. Due to the long storage
time, only 72 in 81 pairs of tissue samples were qualified for
RT-qPCR. All patients had complete pathological parameters and
follow-up information, including 53 males and 28 females with an
average age of 64.1 years (range, 42–83 years). TNM staging was as
follows: 5 cases of stage I, 23 cases of stage II, 48 cases of
stage III and 5 cases of stage IV. In total, 30 pairs of GC and
matched paracancerous specimens for western blot analysis were
collected from patients with GC admitted to the First Affiliated
Hospital of Chongqing Medical University from July to August, 2016.
The patient cohort comprised 18 male and 12 female patients with an
average age of 58.3 years (range, 31–83 years). All patients
underwent total or subtotal gastrectomy and did not receive
radiotherapy and chemotherapy prior to surgery. The use of human
tissue samples and experimental protocols were approved by the
Medical Ethics Review Committee of the First Affiliated Hospital of
Chongqing Medical University and written informed consent was
obtained from all patients.
Cells and lentiviral transduction
The human GC cell line, SGC7901, was obtained from
the Chinese Academy of Sciences Shanghai Cell Bank (Shanghai,
China) and cultured in RPM-1640 (HyClone, Shanghai, China)
supplemented with 10% fetal bovine serum (HyClone) in an incubator
with 5% CO2 at 37°C. The overexpression vector
pLV-HOXA5-GFP-puro, the control vector pLV-GFP-puro and polybrene
were obtained from Hanbio Technology (Shanghai, China). The SGC7901
cells were seeded at 1×105/well in 6-well plates (1
ml/well). After 10 h, lentiviruses were added into the plates at a
MOI of 40. Polybrene was added at a final concentration of 5 µg/ml
to each well. The culture media were replaced after 15 h. After 72
h, the cells were examined to determine the transduction efficiency
under a fluorescence microscope (Olympus, Tokyo, Japan) and
puromycin (Beyotime, Shanghai, China) was then added at a final
concentration of 2 µg/ml to each well. After selection for 2 weeks,
puromycin was removed and the transduced cells were used in further
experiments.
Antibodies and reagents
Rabbit anti-human HOXA5 (Cat. #ab82645), p21 (Cat.
#ab227443) and c-Myc (Cat. #ab32072) antibodies were purchased from
Abcam (Cambridge, UK). Rabbit anti-human Ki67 (Cat. #AP19895b)
antibody was purchased from Abgent (San Diego, CA, USA). Rabbit
anti-human GAPDH (Cat. #10494-1-AP), cyclin D1 (Cat. #12363-1-AP)
and cyclin E (Cat. #11554-1-AP) antibodies and horseradish
peroxidase (HRP)-conjugated goat anti-rabbit secondary antibody
(Cat. #SA00001-15) were purchased from Proteintech (Wuhan, China).
Immunohistochemistry-related reagents, western blot
analysis-related reagents, CCK-8 reagents and DAPI were purchased
from Beyotime (Shanghai, China). All reagents used in RT-qPCR were
purchased from Takara (Dalian, China). The
5-ethynyl-2′-deoxyuridine (EdU) proliferation assay kit was
obtained from Ribobio (Guangzhou, China). All primers used for
RT-qPCR were obtained from Sangon Biotech (Shanghai, China).
RT-qPCR
RT-qPCR was used to assess the differences in
HOXA5 mRNA expression levels between the GC tissues and
adjacent paracancerous tissues. Total RNA was extracted using
TRIzol reagent (Takara, Dalian, China) and reverse transcribed into
cDNA using the reverse transcription kit from Takara. The reverse
transcription conditions were as follows: 37°C, 15 min; 85°C, 5
sec. Two-step RT-qPCR was performed using a SYBR-Green assay on a
CFX96 PCR machine (Bio-Rad, USA) according to the instruction. The
PCR conditions were as follows: Pre-denaturation at 95°C for 30
sec; 45 cycles of denaturation at 95°C for 5 sec and annealing at
60°C for 30 sec, extension at 65°C for 1 min. The data were
analyzed using the ΔΔCq method (8).
The primers used are shown in Table
I.
 | Table I.The sequences of the PCR primers used
in this study. |
Table I.
The sequences of the PCR primers used
in this study.
| Gene | Primer |
|---|
| GAPDH | F:
5′-CTTTGGTATCGTGGAAGGACTC-3′ |
|
| R:
5′-GTAGAGGCAGGGATGATGTTCT-3′ |
| HOXA5 | F:
5′-AGCCACAAATCAAGGACACA-3′ |
|
| R:
5′-GCTCGCTCACGGAACTATG-3′ |
|
E-cadherin | F:
5′-TGGCTTCCCTCTTTCATCTCC-3′ |
|
| R:
5′-TCATAGTTCCGCTCTGTCTTTGG-3′ |
|
N-cadherin | F:
5′-CGTGAAGGTTTGCCAGTGTGA-3′ |
|
| R:
5′-CCTGGCGTTCTTTATCCCG-3′ |
|
Vimentin | F:
5′-TCAATGTTAAGATGGCCCTTG-3′ |
|
| R:
5′-TGAGTGGGTATCAACCAGAGG-3′ |
| CD44 | F:
5′-TTACTCTGCTGCGTTGTCATTG-3′ |
|
| R:
5′-ACAACACCACGCCCAGAGGA-3′ |
| EpCAM | F:
5′-CGCAGCTCAGGAAGAATGTGT-3′ |
|
| R:
5′-AGCCATTCATTTCTGCCTTCAT-3′ |
| Lgr5 | F:
5′-GAGCTGCCTTCCAACCTCAG-3′ |
|
| R:
5′-CCCGCAAGACGTAACTCCTC-3′ |
| CCND1 | F:
5′-ATGTTCGTGGCCTCTAAGATGA-3′ |
|
| R:
5′-CAGGTTCCACTTGAGCTTGTTC-3′ |
| CCNB1 | F:
5′-AATAAGGCGAAGATCAACATGGC-3′ |
|
| R:
5′-TTTGTTACCAATGTCCCCAAGAG-3′ |
| Ki67 | F:
5′-ATCGTCCCAGGTGGAAGAGTT-3′ |
|
| R:
5′-ATAGTAACCAGGCGTCTCGTGG-3′ |
| MMP2 | F:
5′-GACATACATCTTTGCTGGAGAC-3′ |
|
| R:
5′-TTCAGGTAATAGGCACCCTT-3′ |
| MMP7 | F:
5′-CGGATGGTAAGCAGTCTAGGG-3′ |
|
| R:
5′-AGGTTGGATACATCACTGCATTAG-3′ |
| VEGFA | F:
5′-CACACAGGATGGCTTGAAGA-3′ |
|
| R:
5′-AGGGCAGAATCATCACGAAG-3′ |
| VEGFR2 | F:
5′-CCAGATGACAACCAGACGGA-3′ |
|
| R:
5′-AGCCTTCAGATGCCACAGACTC-3′ |
| EGFR | F:
5′-AGGCACGAGTAACAAGCTCAC-3′ |
|
| R:
5′-ATGAGGACATAACCAGCCACC-3′ |
| ABCC1 | F:
5′-GTGATGGCCATGAAGACCAAGA-3′ |
|
| R:
5′-GCCAGCTCCCAGGCATAAAG-3′ |
| ABCG2 | F:
5′-TCTCTTCTTCCTGACGACCAA-3′ |
|
| R:
5′-AAACCACACTCTGACCTGCTG-3′ |
|
Caspase-3 | F:
5′-TACCAGTGGAGGCCGACTTC-3′ |
|
| R:
5′-GCACAAAGCGACTGGATGAAC-3′ |
| BCL2 | F:
5′-CTGCACCTGACGCCCTTCACC-3′ |
|
| R:
5′-CACATGACCCCACCGAACTCAAAGA-3′ |
| BECN1 | F:
5′-GGCTGAGAGACTGGATCAGG-3′ |
|
| R:
5′-CTGCGTCTGGGCATAACG-3′ |
| p21 | F:
5′-AGCGGAACAAGGAGTCAG-3′ |
|
| R:
5′-CGTTAGTGCCAGGAAAGAC-3′ |
| p53 | F:
5′-GCGTAAACGCTTCGAGATGTT-3′ |
|
| R:
5′-TTTTTATGGCGGGAAGTAGACTG-3′ |
| C-Myc | F:
5′-ACACCCGAGCAAGGACGCGA-3′ |
|
| R:
5′-CGCGGGAGGCTGCTGGTTTTC-3′ |
|
β-catenin | F:
5′-GAGTGCTGAAGGTGCTATCTGTC-3′ |
|
| R:
5′-CTGAACAAGAGTCCCAAGGAGA-3′ |
Western blot analysis
The cancer tissues and matched paracancerous tissues
were lysed using RIPA lysis buffer, and the protein concentration
was measured using the BCA assay after the lysates were harvested
by centrifugation (12,000 rpm) at 4°C. Subsequently, 20 µg of
protein samples were separated by electrophoresis on a 10% sodium
dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and transferred onto
polyvinylidene fluoride membranes. The membranes were incubated
overnight at 4°C with HOXA5 antibody (1:1,000), p21 antibody
(1:1,000), p53 antibody (1:1,000), cyclin D1 antibody (1:2,000),
cyclin E antibody (1:500), c-Myc antibody (1:1,000) and Ki67
antibody (1:1,000) following blocking of the non-specific binding
sites for 2 h in 5% non-fat milk. After washing with TBS-T,
membranes were incubated with secondary antibody (1:2,000) at 37°C
for 2 h. GAPDH was used as a loading control. An enhanced
chemiluminescence kit (ECL) was used for visualizing immunobands
and the signal intensity of each band was measured by Fusion
software (Vilber Lourmat, Paris, France) to calculate protein
levels.
Immunohistochemistry
Tissues were fixed by formalin and embedded in
paraffin blocks, and a series of sections (4-mm-thick) were
prepared. The tissue antigens were repaired by sodium citrate
buffer (pH 6.0). The sections were incubated with rabbit anti-human
HOXA5 antibody at 4°C overnight after non-specific binding was
blocked with goat serum. The scoring criteria were as follows: 5
fields were randomly selected from each section under a light
microscope (Olympus), and the percentage of positively stained
cells to total cells was calculated. The sections were scored as
follows: 0, <5%; 1, 5–25%; 2, 26–50%; 3, 51–75%; and 4, >75%.
Additionally, the staining intensity was scored as follows: 0, no
staining; 1, light yellow; 2, brownish yellow; and 3, brown. The
total score was the sum of staining range and intensity, and was
classified as low expression (≤3 points) or high expression (≥4
points).
CCK-8 and colony-forming assays
For the CCK-8 assay, the cells were seeded at
2×103/well in 96-well plates (200 µl/well). After cell
adherence, 20 µl CCK-8 reagent were added to each well on days 0,
1, 2 and 3. Following incubation at 37°C for 1.5 h, the OD value at
450 nm was measured using a microplate reader (Thermo Fisher
Scientific, Waltham, MA, USA), and the growth curve was then
plotted. For the colony formation assay, the cells were seeded at
500/well in 6-well plates (1 ml/well) and cultured in complete
medium. The culture medium was replaced every 4 days. After 15
days, the cells were fixed with 4% paraformaldehyde and stained
with crystal violet (Beyotime, Shanghai, China) at room temperature
for 10 min. Images were obtained to calculate the number of
clones.
Flow cytometry
At least 5×105 cells were suspended and
fixed with 70% ethanol for 2 h. Propidium iodide (IP) was used to
stain the cells and the cell cycle distribution was detected using
a flow cytometer (BD Biosciences, San Jose, CA, USA).
5-ethynyl-2′-deoxyuridine (EdU)
assay
The cells were seeded at 2×103/well in
96-well plates (200 µl/well) and were treated with 50 µM EdU for 2
h at 37°C. After washing with PBS and fixing with 4%
paraformaldehyde for 30 min, the cells were permeabilized with 0.5%
Triton X-100 for 5 min. The cells were then incubated with 100 µl
1× Apollo reaction cocktail for 30 min. DAPI was then used to stain
the nuclei for 15 min. Images were obtained under a fluorescence
microscope (Olympus) to calculate the proliferation rate.
Nude mouse models
A total of 12 nude mice aged 4 weeks old (female,
weighing 13 to 16 g) were obtained from the Animal Experimental
Center of Chongqing Medical University and were maintained at
24±2°C under a 40–70% humidity with free access to food and water.
The experiment was divided into SGC7901-NC group and SGC7901-HOXA5
group, with 6 mice in each group. When mice were 5 weeks old,
2×106 cells suspended in 100 µl PBS were injected
subcutaneously into the left flank. The tumor volume was monitored
by measuring diameters and calculated as LxS2/2 (L
indicates the long side and S indicates the short side) every 3
days. After 3 weeks, the nude mice were sacrificed by ether
anesthesia combined with carbon dioxide asphyxiation. The flow rate
of the gas was 20% of the chamber volume per minute and the death
of the mice was judged according to breathing, heartbeat, nerve
reflex and muscle relaxation after 5 min. The tumor tissues were
stained with hematoxylin and eosin (H&E) for pathological
confirmation. The animal experimental protocols were approved by
the Medical Ethics Review Committee of the First Affiliated
Hospital of Chongqing Medical University.
GEPIA analysis
Gene Expression Profiling Interactive Analysis
(GEPIA, http://gepia.cancer-pku.cn), an
online cancer microarray database, was used to analyze the effect
of HOXA5 on the overall survival of GC patients (9). Patients were divided into high
expression group and low expression group according to the median
expression level of HOXA5 mRNA (Normalized by GAPDH) and the
overall survival was assessed by a Kaplan-Meier survival plot.
Statistical analysis
All experiments were repeated 3 times and the data
were analyzed using SPSS 19.0 software. Measurement data are
expressed as the means ± standard deviation (SD). Comparisons were
made between two groups using the Student's t-test. Wilcoxon
signed-rank tests were used to assess the differences in
HOXA5 mRNA expression levels between the GC tissues and
adjacent paracancerous tissues of 72 patients. The correlation
between HOXA5 protein expression and each clinicopathological
parameter was examined using χ2 tests. Survival curves
for different expression levels of HOXA5 were plotted according to
the Kaplan-Meier method, and prognostic significance of all factors
was calculated by the log-rank test. Significant parameters
(P<0.05) in the univariate model were further evaluated using
multivariate Cox regression, after which the independence and
hazard ratio (HR) of each prognostic factor could be calculated.
Differences were considered statistically significant at a P-value
<0.05. The correlation between the expression of HOXA5
mRNA and 23 genes related to malignant biological behavior in 30 GC
tissues was determined using Spearman's correlation analysis.
Results
Decreased expression of HOXA5 in human
GC tissues
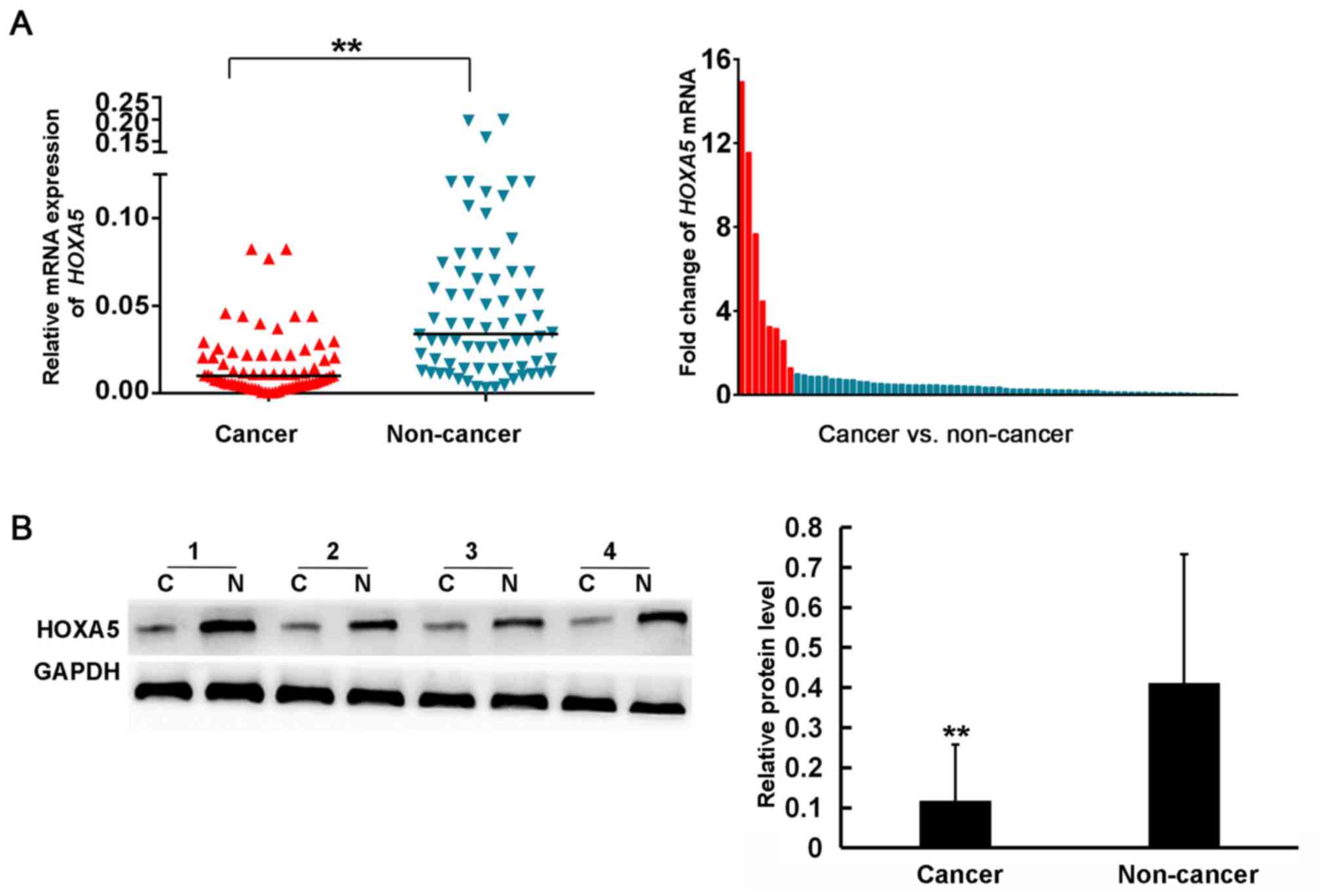
To examine the role of HOXA5 in GC, RT-qPCR was used
to detect the mRNA expression of HOXA5 in 72 pairs of GC
issues and adjacent paracancerous tissues. The results demonstrated
that 63 patients (87.5%) exhibited a lower mRNA expression of
HOXA5 in GC tissues compared to the matched non-cancer
tissues (Fig. 1A). The average mRNA
expression level of HOXA5 in the cancer tissues was
3.23-fold lower than that in the adjacent non-cancer tissues.
Subsequently, western blot analysis was performed to determine
whether HOXA5 was also downregulated at the protein level in 30 GC
patients. As shown in Fig. 1B, the
average protein expression level of HOXA5 in the GC tissues was
3.73-fold lower than that in the non-cancer tissues (0.11±0.14 vs.
0.41±0.32, respectively). These results suggested that HOXA5
expression was decreased in GC.
HOXA5 acts as a tumor suppressor gene
in GC
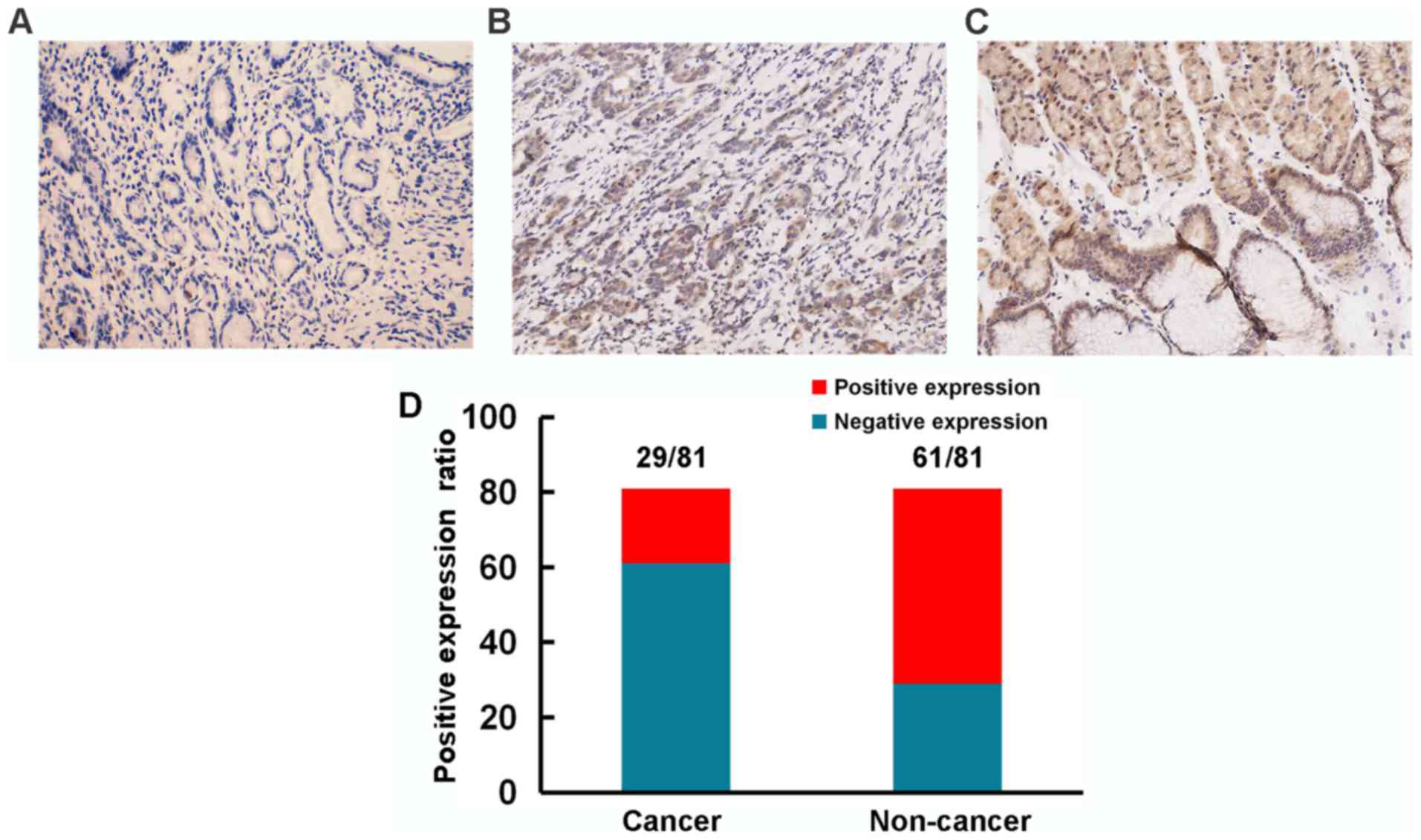
To further illuminate the role of HOXA5 in the
progression of GC, we used immunohistochemistry to detect the
expression of HOXA5 in samples from 81 patients with GC and
analyzed the association between HOXA5 and clinical parameters. We
found that HOXA5 staining was mainly located in the nucleus and
cytoplasm (Fig. 2). Compared with a
75.3% positive expression of HOXA5 (61/81) in the adjacent
non-cancer tissues, the positive expression in the GC tissues was
only 35.8% (29/81).
The GC specimens were then divided into a high
expression group and a low expression group, according to the
immunohistochemistry score for HOXA5 expression. As shown in
Table II, a decreased HOXA5
expression was significantly associated with tumor size and
histological grade. These results thus suggest that HOXA5
acted as a tumor suppressor gene in the progression of GC.
| Table II.Association between the expression of
HOXA5 and clinical parameters of patients with gastric cancer
(GC). |
Table II.
Association between the expression of
HOXA5 and clinical parameters of patients with gastric cancer
(GC).
|
|
| HOXA5 |
|
|
| Index | Case | + | − | χ2 | P-value |
|---|
| Sex |
|
|
| 0.974 | 0.324 |
|
Male | 53 | 21 | 32 |
|
|
|
Female | 28 | 8 | 20 |
|
|
| Age (years) |
|
|
| 0.822 | 0.365 |
|
<60 | 31 | 13 | 18 |
|
|
|
≥60 | 50 | 16 | 34 |
|
|
| Histological
grade |
|
|
| 7.814 | 0.005b |
|
Well-differentiated | 39 | 20 | 19 |
|
|
|
Poorly-differentiated | 42 | 9 | 33 |
|
|
| Tumor size
(cm) |
|
|
| 10.65 | 0.001b |
|
<5 | 39 | 21 | 18 |
|
|
| ≥5 | 42 | 8 | 34 |
|
|
| TNM stage |
|
|
| 2.486 | 0.115a |
|
1+2 | 38 | 17 | 21 |
|
|
|
3+4 | 43 | 12 | 31 |
|
|
| Lymph node
involvement |
|
|
| 0.510 | 0.475 |
| No | 24 | 10 | 14 |
|
|
|
Yes | 57 | 19 | 38 |
|
|
| Vascular
invasion |
|
|
| 2.523 | 0.112 |
|
Absent | 16 | 3 | 13 |
|
|
|
Present | 65 | 26 | 39 |
|
|
| Distant
metastasis |
|
|
| 0.008 | 0.928 |
| No | 78 | 28 | 50 |
|
|
|
Yes | 3 | 1 | 2 |
|
|
Decreased expression of HOXA5 protein
as a prognostic marker for GC
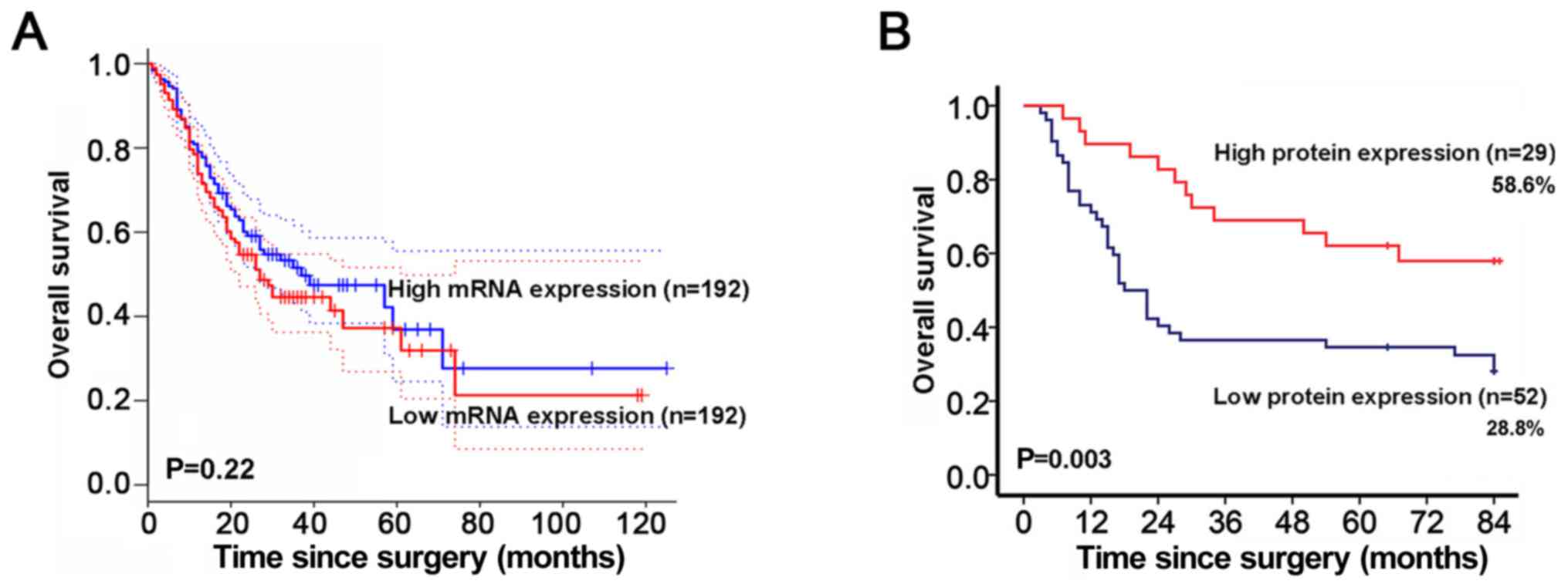
To investigate the prognostic value of HOXA5 for GC,
survival analysis from GEPIA database was obtained. HOXA5
mRNA was not associated with the overall survival of patients with
GC (Fig. 3A). Given that HOXA5
functions as a transcription factor, we further assessed the
association between HOXA5 protein expression and survival according
to the immunohistochemistry score. It is worth noting that the 75%
survival duration of the patients in the low protein expression
group was 10 months, which was significantly shorter than that of
30 months for the patients in the high protein expression group.
Patients with a low protein expression level of HOXA5 had an
overall survival rate of 28.8% in comparison to 58.6% for patients
with a high protein expression level (Fig. 3B).
According to a univariate analysis of the prognostic
factors of GC, the overall survival was found to be significantly
associated with each of the following: Histological grade
(P=0.031), tumor size (P<0.001), lymph node involvement
(P<0.001), TNM stage (P<0.001), distant metastasis
(P<0.001), and decreased HOXA5 expression (P=0.003). To
investigate whether a decreased HOXA5 is an independent prognostic
marker in patients with GC, a multivariate Cox regression analysis
was performed. We found that tumor size, lymph node involvement,
TNM stage, distant metastasis and a decreased HOXA5 expression were
significantly associated with overall survival after controlling
for other prognostic factors in the multivariate Cox regression
analysis (Table III). These data
suggested that a decreased HOXA5 expression was an independent risk
factor for the prognosis of GC and could be a prognostic marker for
patients with GC.
| Table III.Kaplan-Meier univariate survival
analysis and multivariate Cox regression analysis of prognostic
factors in gastric cancer (GC) for overall survival. |
Table III.
Kaplan-Meier univariate survival
analysis and multivariate Cox regression analysis of prognostic
factors in gastric cancer (GC) for overall survival.
|
| Univariate | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variables | P-value | HR | 95% CI | P-value |
|---|
| Sex | 0.307 |
|
|
|
| Age (years) | 0.943 |
|
|
|
| Histological
grade | 0.031a | 0.683 | 0.379–1.234 | 0.206 |
| Tumor size |
<0.001b | 2.697 | 1.309–5.558 | 0.007b |
| TNM stage |
<0.001b | 3.083 | 1.474–6.447 | 0.003b |
| Lymph node |
<0.001b | 3.787 | 1.308–10.962 | 0.014a |
| Vascular
invasion | 0.284 | 0.714 | 0.334–1.526 | 0.385 |
| Distant
metastasis |
<0.001b | 9.263 | 2.068–41.502 | 0.004b |
| HOXA5
expression | 0.003b | 0.429 | 0.202–0.912 | 0.028a |
Identification of potential target
genes of HOXA5
The progression of GC involves a series of cellular
biological events, including the loss of control of cell
proliferation, an enhanced angiogenesis, the acquisition of
epithelial-mesenchymal transition (EMT) and stemness, and sustained
invasion and drug resistance. Our data suggested that HOXA5
inhibited the progression of GC. In order to further explore the
underlying mechanisms, we used RT-qPCR to detect the expression of
the following genes in 30 GC tissues, including
tumorigenesis-related genes (p21, p53 and β-catenin),
proliferation-related genes (cyclin B1, cyclin D1, c-Myc and
Ki67), angiogenesis-related genes [vascular endothelial
growth factor (VEGF)A, VEGFR2 and epidermal growth
factor (EGFR)], metastasis-related genes [matrix
metalloproteinase (MMP2) and (MMP7)],
apoptosis-related genes (BCL2 and caspase-3), EMT
markers (vimentin, N-cadherin and E-cadherin), stemness markers
(Lgr5, CD44 and EpCAM) and multi-drug resistance
genes (ABCC1, ABCG2 and Beclin1) (10–15). A
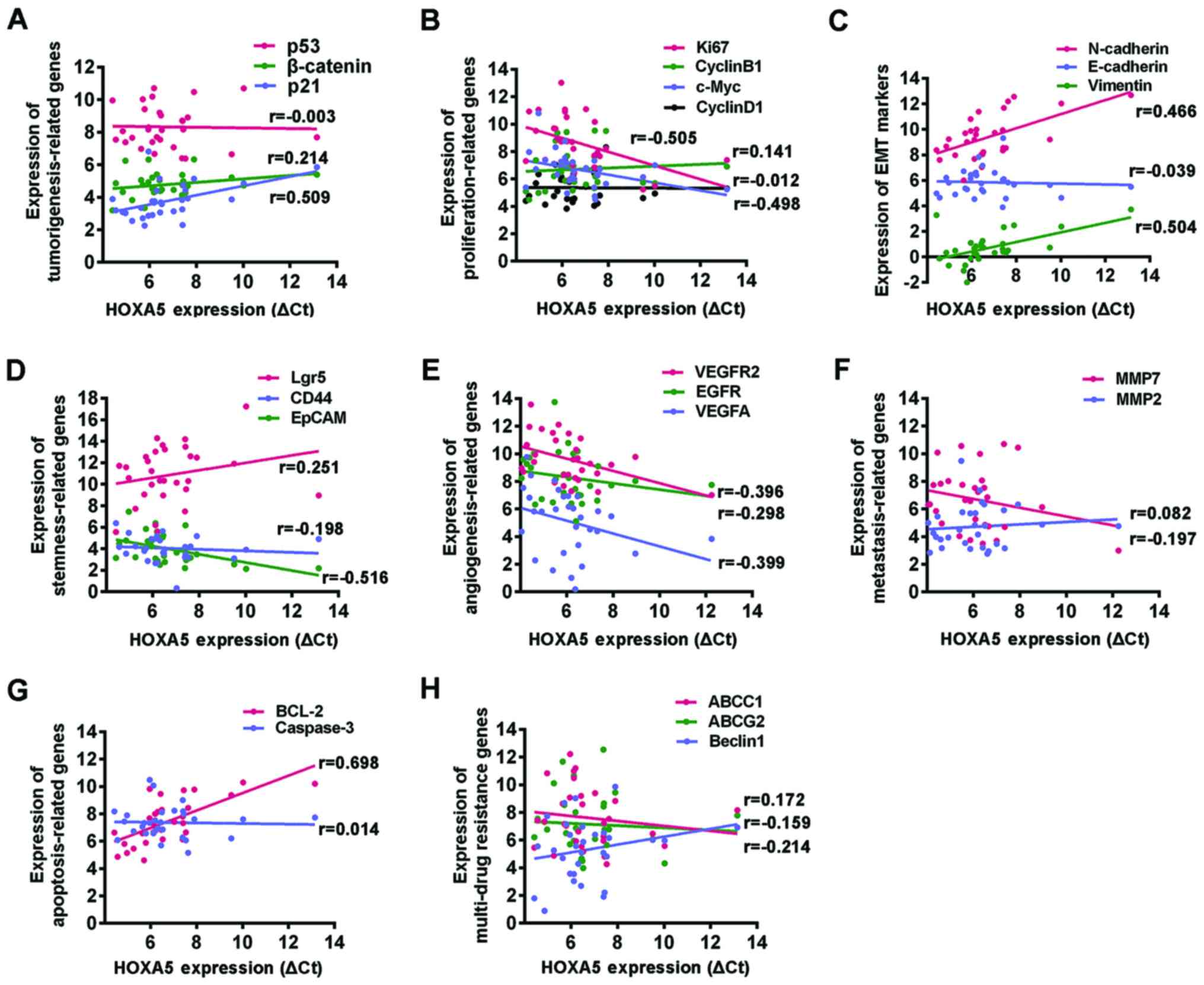
Spearman's correlation model was used to examine the correlation
between each of these genes and HOXA5 mRNA expression. We
found that HOXA5 mRNA expression positively correlated with
the tumor suppressor gene, p21, and negatively correlated
with the proliferation-related genes, c-Myc and Ki67,
the angiogenesis genes, VEGFA and VEGAFR2, and the
stemness marker, EpCAM. No correlation was observed with the other
genes examined. Surprisingly, although HOXA5 exhibited a role in
inhibiting the progression of GC, its mRNA expression positively
correlated with the mesenchymal factors, vimentin and N-cadherin,
which indicated the acquisition of EMT; it also positively
correlated with the anti-apoptotic molecule, BCL2, which is known
to play an important role in apoptosis resistance (Fig. 4 and Table IV).
| Table IV.Spearman's correlation analysis of
the correlation between the expression of HOXA5 and
malignant behavior- related genes. |
Table IV.
Spearman's correlation analysis of
the correlation between the expression of HOXA5 and
malignant behavior- related genes.
|
| Spearman's
correlation analysis |
|---|
|
|
|
|---|
| Variables | r value | P-value |
|---|
|
Carcinogenesis-related genes |
|
p21 | 0.509 |
<0.01b |
|
p53 | −0.003 | 0.987 |
|
β-catenin | 0.214 | 0.256 |
|
Proliferation-related genes |
|
CCNB1 | 0.141 | 0.456 |
|
CCND1 | −0.012 | 0.951 |
|
Ki67 | −0.505 |
<0.01a |
|
C-Myc | −0.498 |
<0.01a |
| EMT markers |
|
E-cadherin | −0.039 | 0.836 |
|
N-cadherin | 0.466 | 0.01a |
|
Vimentin | 0.504 |
<0.01a |
| Stemness
markers |
|
Lgr5 | 0.251 | 0.181 |
|
CD44 | −0.198 | 0.295 |
|
EpCAM | −0.516 |
<0.01a |
|
Angiogenesis-related genes |
|
VEGFA | −0.399 | 0.029b |
|
VEGFR2 | −0.396 | 0.03b |
|
EGFR | −0.298 | 0.109 |
| Metastasis-related
genes |
|
MMP2 | 0.082 | 0.668 |
|
MMP7 | −0.197 | 0.296 |
| Apoptosis-related
genes |
|
BCL2 | 0.698 |
<0.01a |
|
Caspase-3 | −0.014 | 0.941 |
| Multi-drug
resistance genes |
|
ABCC1 | −0.214 | 0.256 |
|
ABCG2 | −0.159 | 0.401 |
|
Beclin1 | 0.172 | 0.364 |
Enforced overexpression of HOXA5
suppresses the aberrant proliferation of GC cells
The immunohistochemistry results revealed that HOXA5
expression negatively correlated with the size of the tumor.
Moreover, correlation analysis revealed that Ki67, c-Myc and p21
(these 3 genes are involved in the regulation of tumor
proliferation) may be potential target genes of HOXA5. This
indicated that HOXA5 may exert its anticancer effects through the
negative regulation of cancer cell proliferation. To examine the
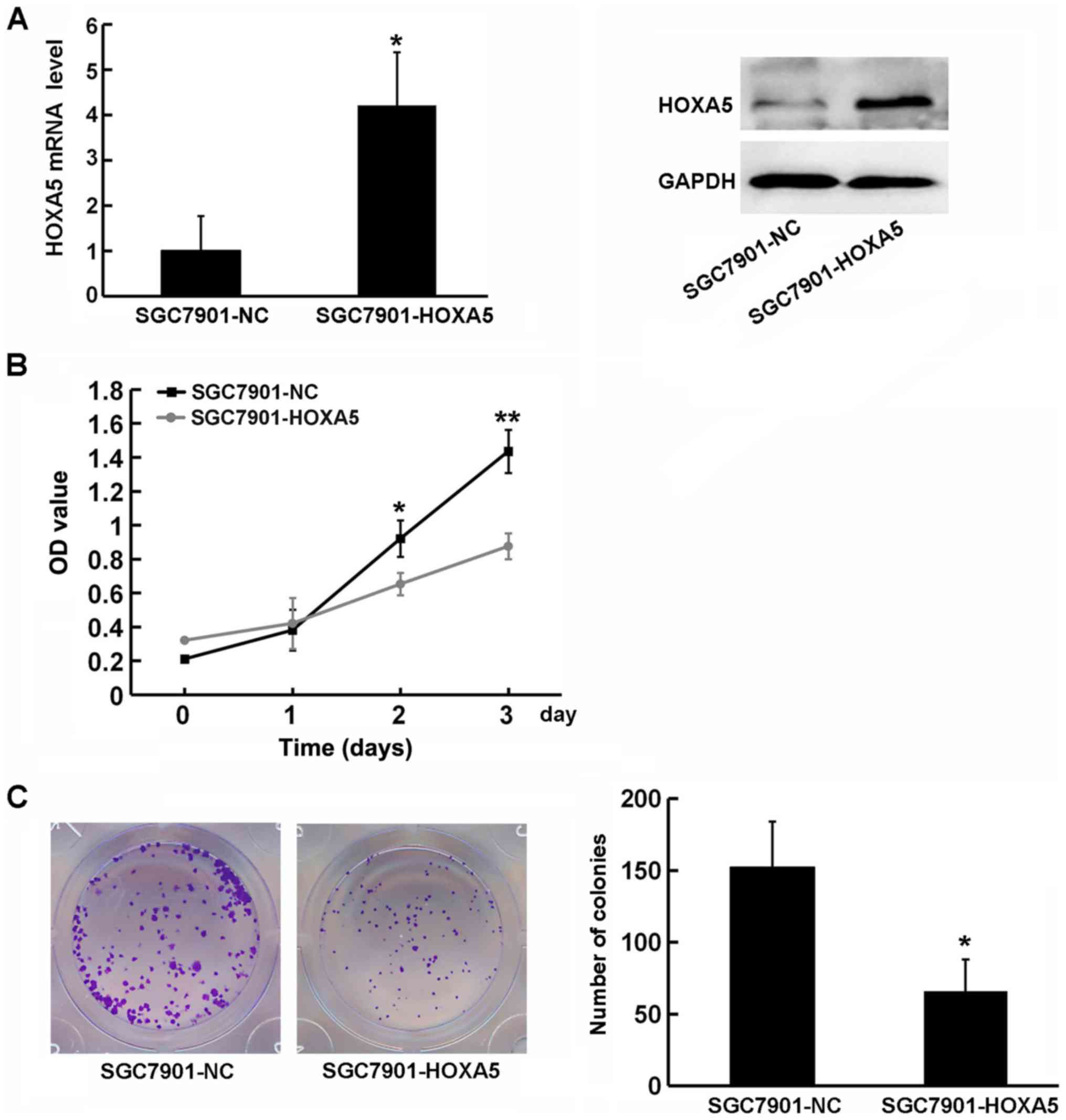
function of HOXA5 in the progression of GC, we used HOXA5
overexpression vectors to transfect GC SGC7901 cells, which express
a low level of HOXA5, and detected its effect on cell proliferation
using a CCK-8 assay (Fig. 5A). The
growth curve indicated that HOXA5-overexpressing cells
(SGC7901-HOXA5) exhibited a significantly decreased proliferation
compared with the negative control cells (SGC7901-NC) (Fig. 5B). Similarly, a colony formation
assay revealed that HOXA5 overexpression resulted in an impairment
of SGC7901 colony formation (Fig.
5C). These results indicated that the upregulation of HOXA5 in
GC cells was beneficial in inhibiting cell growth, and that HOXA5
suppressed the proliferation of GC cells.
Enforced overexpression of HOXA5
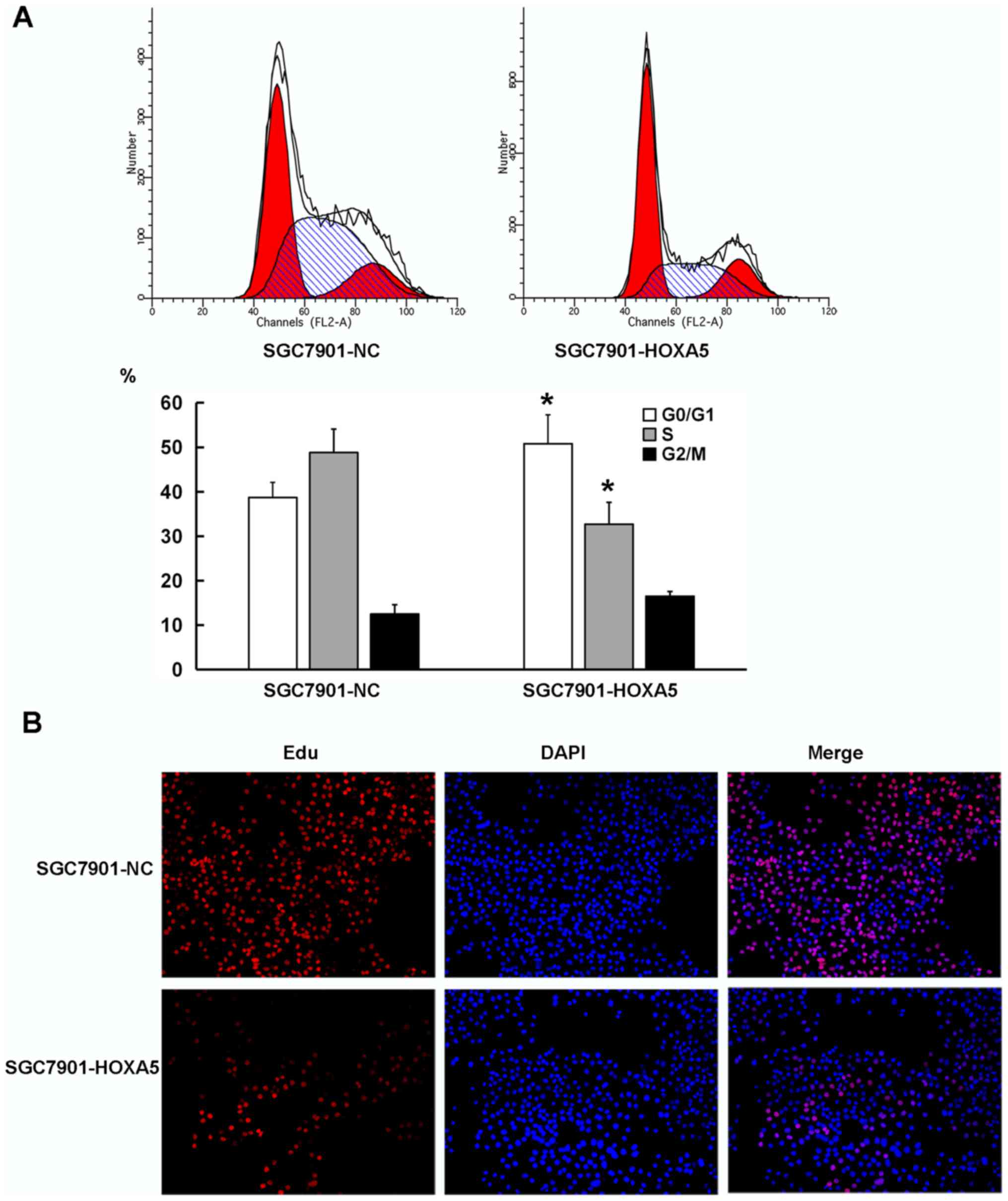
decelerates the G1-S phase transition of GC cells
Cell proliferation depends on the proper progression
of the cell cycle, particularly in the G1-S phase
transition. As shown in Fig. 6A,
the results from flow cytometry revealed that compared with the
SGC7901-NC cells, the SGC7901-HOXA5 cells exhibited a significantly
increased percentage of cells in the G0/G1
phase (38.7% vs. 50.8%), but an decreased percentage of cells in
the S phase (48.6% vs. 32.7%). We then employed an EdU staining
assay to detect the cell proliferation rate of the cells in the
SGC7901-NC and SGC7901-HOXA5 groups. Fluorescence microscopy
revealed a lower proliferative activity in the SGC7901-HOXA5 group
(19.2%) than that noted in the SGC7901-NC group (55.7%), which was
consistent with the flow cytometry data (Fig. 6B). These results indicated that the
enforced overexpression of HOXA5 decelerated the G1-S
transition of the GC cells.
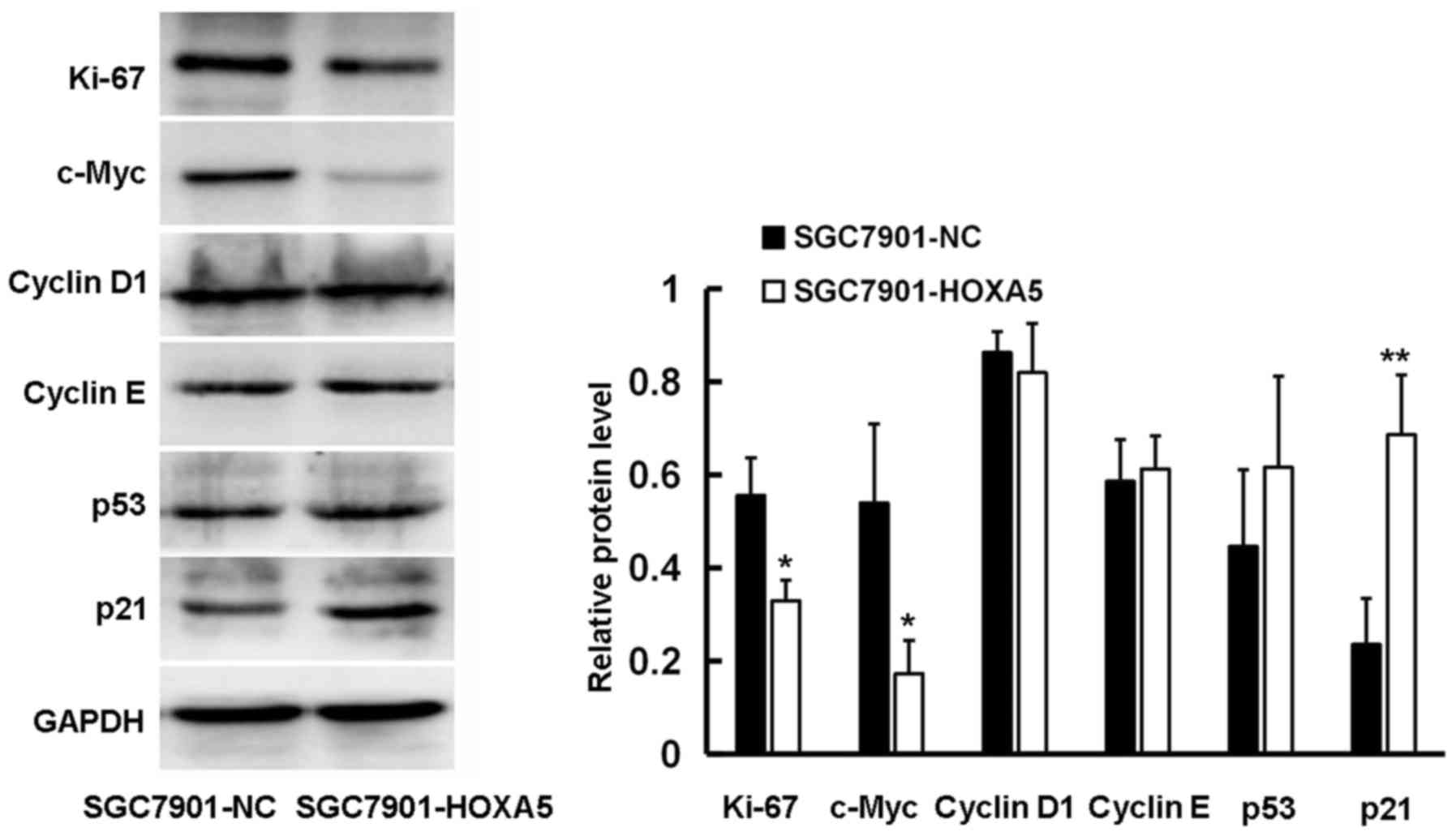
HOXA5 regulates the expression of cell
cycle-related proteins
Our data suggested that HOXA5 decelerated the cell
cycle, thus inhibiting the proliferation of GC cells. In order to
further clarify the underlying mechanisms, we used western blot
analysis to detect the effect of HOXA5 on a series of cell
cycle-related proteins. We found that HOXA5 promoted the expression
of p21 and inhibited the expression of c-Myc and Ki67 at the
protein level, although it had no marked effect on the expression
of p53, cyclin D1 and cyclin E (Fig.
7). This may be the underlying mechanism through which HOXA5
exerts its inhibitory effect on GC cell proliferation.
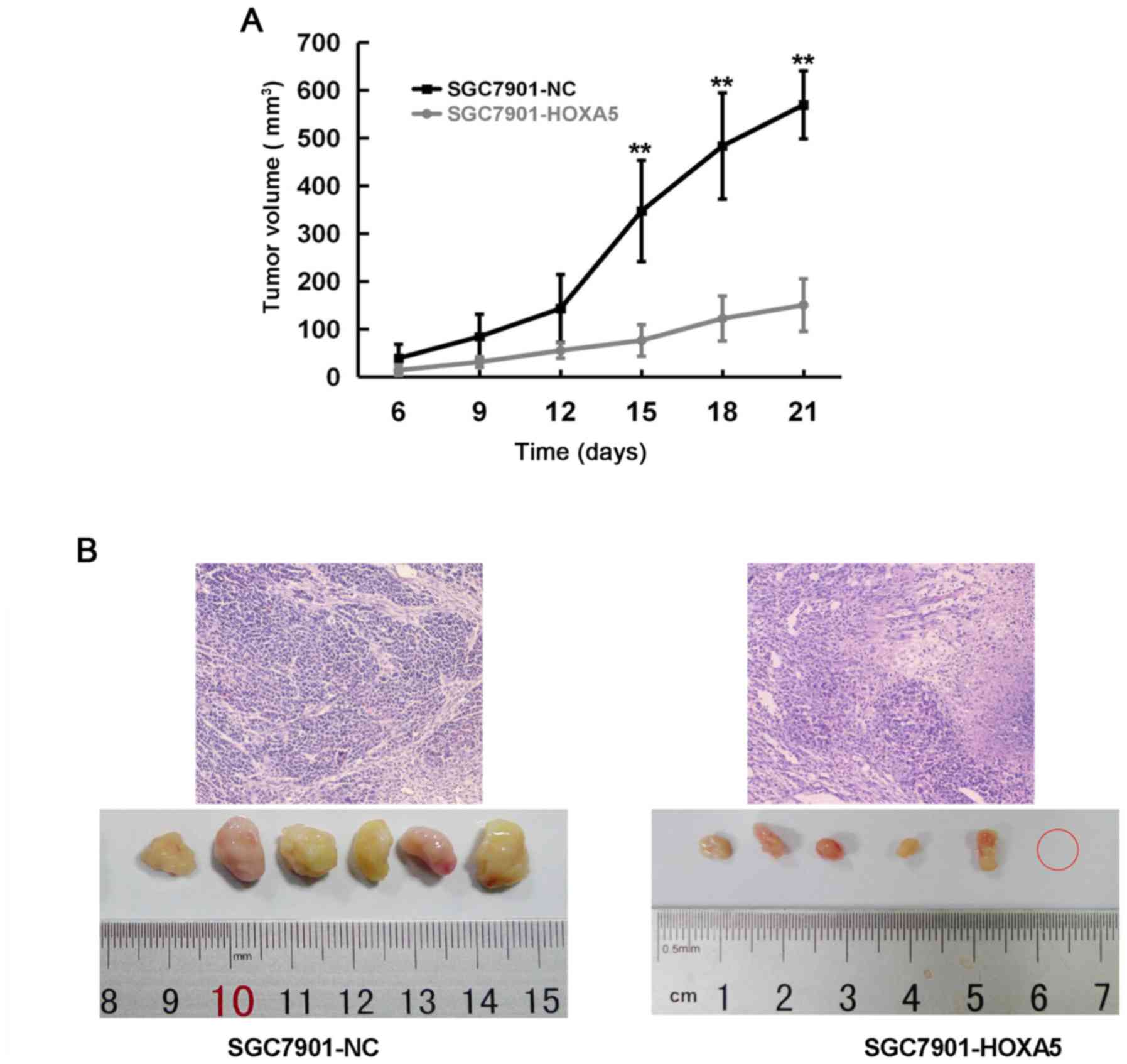
HOXA5 suppresses the proliferation and
promotes the differentiation of GC cells in vivo
Xenograft experiments in vivo are the gold
standard to determine cell proliferation. Thus, we injected
SGC7901-NC or SGC7901-HOXA5 cells subcutaneously into nude mice to
observe their different effects on proliferation. As shown in the
tumor growth curve, the SGC7901-HOXA5 group began to display a
smaller tumor volume than that of the SGC7901-NC group from day 15
(Fig. 8A). In addition, H&E
staining revealed that compared with the NC group, the tumors in
the SGC7901-HOXA5 group were loosely arranged, with more
mesenchymal and inflammatory cell infiltration and fewer abnormal
cell mitoses. The overexpression of HOXA5 promoted the tumor cells
to enter a more differentiated state (Fig. 8B). These results suggested that
HOXA5 suppressed the proliferation and promoted the differentiation
of GC cells in vivo.
Discussion
In the present study, we demonstrated that HOXA5 was
downregulated in GC and that the underexpression of HOXA5 was
associated with the progression of GC. In addition, we also
demonstrated that the underexpression of HOXA5 was an independent
prognostic factor for patients with GC. Furthermore, we revealed
that HOXA5 may exert its anticancer effects through the inhibition
of the abnormal proliferation of GC cells.
GC is the second most common malignant tumor
worldwide and is associated with a poor prognosis. Despite the
existence of comprehensive treatments based on surgery and
chemotherapy, the 5-year survival rate of patients with GC has not
improved significantly. It is thus of great clinical significance
to find an effective molecular target for the early diagnosis and
treatment of GC.
HOXA5 is a member of the homeobox gene family, and
it encodes the 29 kDa HOXA5 protein, which functions as a critical
master regulatory factor in controlling embryonic development and
adult stem cell differentiation. Recently, accumulating evidence
has revealed that the human HOXA5 protein limits the aggressiveness
of breast cancer and colon cancer (4,5). To
clarify the associatoin between HOXA5 and GC, we detected the
expression of HOXA5 using RT-qPCR and western blot analysis in 30
patients with GC and found that HOXA5 expression was significantly
lower in GC tissues than that in adjacent non-cancer tissues. This
result was consistent with a previous finding, in that HOXA5 was
hypermethylated in GC tissues (7).
A recent study reported that HOXA5 was a marker of a good prognosis
in patients with colon cancer (4),
although another study demonstrated that the overexpression of
HOXA5 was associated with a poor prognosis in non-small cell lung
cancer (16). In this study, we
found that a decreased HOXA5 expression in GC tissues was
associated with larger tumors that were poorly differentiated and a
higher TNM stage.
Although that the mRNA level of HOXA5 was not
directly associated with the prognosis of patients with GC,
patients with a low HOXA5 protein expression had a significantly
shorter overall survival than patients with a high expression. One
possible explanation for this is that HOXA5 is a transcription
factor which functions as a protein and there may be
post-transcriptional modifications during the process of
translation of HOXA5 mRNA. Further multivariate analysis
confirmed that a decreased HOXA5 expression, together with tumor
size, lymph node involvement, distant metastasis and TNM stage,
were independent prognostic factors in patients with GC. Therefore,
our results support the notion that a decreased HOXA5 expression
promotes the progression of GC and is an indicator of a poor
prognosis of patients with GC.
As is widely known, cancer progression involves a
series of cellular biological events, including the loss of control
of cell proliferation, angiogenesis, EMT, stemness, invasion and
drug resistance (17–20). In this study, we found that the mRNA
expression of HOXA5 positively correlated with the broad
spectrum tumor suppressor gene, p21. Although it has been
shown that HOXA5 promotes apoptosis by the transcriptional
regulation of p53 in breast cancer cells expressing wild-type
p53 (21), our study did not
show an association of HOXA5 with p53. In addition,
HOXA5 was found to negatively correlate with the
proliferation-related genes, c-Myc and Ki67, the
angiogenesis-related genes, VEGFA and VEGAFR2, and
the stemness marker EpCAM, suggesting that HOXA5 may
suppress GC progression by inhibiting abnormal proliferation,
angiogenesis and the acquisition of stemness. These results are
consistent with previous findings on breast cancer and colon cancer
(4,5). It is worth noting that HOXA5
exhibited a significant positive correlation with the mesenchymal
factors, N-cadherin and vimentin, and the anti-apoptotic molecule,
BCL2. It is well known that the upregulation of mesenchymal
molecules can induce cytoskeletal remodeling, polarity loss and an
increased migratory capacity, which are early markers of metastasis
in epithelium-derived tumors (22).
However, taking into account that HOXA5 is expressed in the
mesenchyme and regulates organ patterning through signal pathways
such as the hedgehog (Hh) and transforming growth factor-β (TGF-β)
pathways in the processes of respiratory and digestive tract
development, it is not difficult to envision that HOXA5 also
increases N-cadherin and vimentin expression in GC (23). Although HOXA5 may increase
BCL2 expression according to our data, the association
between HOXA5 and apoptosis remains to be further verified given
the presence of multiple complex signaling pathways in the
regulation of apoptosis. Our results suggested that HOXA5 may
inhibit the proliferation, angiogenesis and the stemness of GC
through mediators or signaling pathways. Despite possibly
facilitating EMT, HOXA5 exerted an inhibitory effect on the
progression of GC.
Previous studies have reported that HOXA5 inhibits
the proliferation of breast and lung cancer cells (24,25).
Moreover, in this study, the results from immunohistochemistry also
indicated that HOXA5 may exert its anticancer effect through the
negative regulation of cancer cell proliferation. To further
confirm the role of HOXA5 in GC cell proliferation, we
overexpressed HOXA5 by the transduction of lentiviral vectors in
SGC7901 cells. As was suspected, the enforced overexpression of
HOXA5 significantly inhibited the proliferation of GC cells, as
shown by CCK-8 and colony formation assays. The results from flow
cytometry and EdU assay revealed that HOXA5 overexpression induced
arrested cell cycle at the G1 phase. The G1-S
transition is regulated by a series of factors (26,27).
In this study, we demonstrated that HOXA5
upregulated p21 and downregulated c-Myc and Ki67, although the
expression of two important G1 phase proteins, cyclin D1
and cyclin E, was not altered. The cyclin-dependent kinase
inhibitor, p21, inhibits the activity of the cyclin D1-CDK4 and
cyclin E-CDK2 complexes and prevents the cells from entering the S
phase from the G1 phase, which is the most important
pathway to block the G1-S transition (28,29).
By contrast, the proliferation-related proteins, c-Myc and Ki67,
contribute to the progression of the cell cycle. It is worth noting
that the correlation model in tissues revealed that the correlation
between these three genes and HOXA5 was unimpressive. Our
function assays in cells demonstrated that the overexpression of
HOXA5 can cause significant p21 upregulation and Ki67, c-Myc
downregulation. One possible explanation is that HOXA5 is a type of
transcription factor and can function through downstream signaling
pathways (27), or direct
transcriptional activation or inhibition of target genes (30) in the form of a protein. Once HOXA5
protein in the GC cells was upregulated, the downstream target
genes revealed obvious changes. Therefore, we hypothesized that
HOXA5 suppressed the aberrant proliferation of GC cells through the
regulation of p21, c-Myc, and Ki67. This may be the mechanism
through which HOXA5 exerts its anti-tumor effects on GC. Finally,
nude mouse models also verified that HOXA5 suppressed the
proliferation of GC cells in vivo.
In conclusion, in this study, we demonstrated that
HOXA5 was a tumor suppressor gene and was decreased in GC.
Its underexpression may be used as a direct prognostic indicator of
a negative outcome. Our research may provide an opportunity for
developing a novel therapeutic target as well as a prognostic
marker in GC.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Key
Clinical Specialties Construction Program of China [grant no.
(2012)649].
Availability of data and materials
All data generated or analyzed during this study are
included in this published article and are freely available to any
researchers.
Authors' contributions
ZW conceived and designed the experiments; XP, LZ
and AC performed the experiments and analyzed the data; and XP
wrote the manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The use of human tissue samples and experimental
protocols were approved by the Medical Ethics Review Committee of
the First Affiliated Hospital of Chongqing Medical University and
written informed consent was obtained from all patients. The animal
experimental protocols were approved by the Medical Ethics Review
Committee of the First Affiliated Hospital of Chongqing Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jiang H, Shah S and Hilt DC: Organization,
sequence, and expression of the murine S100 beta gene.
Transcriptional regulation by cell type-specific cis-acting
regulatory elements. J Biol Chem. 268:20502–20511. 1993.PubMed/NCBI
|
|
3
|
Boucherat O, Montaron S, Bérubé-Simard FA,
Aubin J, Philippidou P, Wellik DM, Dasen JS and Jeannotte L:
Partial functional redundancy between Hoxa5 and Hoxb5 paralog genes
during lung morphogenesis. Am J Physiol Lung Cell Mol Physiol.
304:L817–L830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ordóñez-Morán P, Dafflon C, Imajo M,
Nishida E and Huelsken J: HOXA5 counteracts stem cell traits by
inhibiting wnt signaling in colorectal cancer. Cancer Cell.
28:815–829. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen H, Zhang H, Lee J, Liang X, Wu X, Zhu
T, Lo PK, Zhang X and Sukumar S: HOXA5 acts directly downstream of
retinoic acid receptor beta and contributes to retinoic
acid-induced apoptosis and growth inhibition. Cancer Res.
67:8007–8013. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang CC, Su KY, Chen HY, Chang SY, Shen
CF, Hsieh CH, Hong QS, Chiang CC, Chang GC, Yu SL and Chen JJ:
HOXA5 inhibits metastasis via regulating cytoskeletal remodelling
and associates with prolonged survival in non-small-cell lung
carcinoma. PLoS One. 10:e1241912015.
|
|
7
|
Loh M, Liem N, Vaithilingam A, Lim PL,
Sapari NS, Elahi E, Mok ZY, Cheng CL, Yan B, Pang B, et al: DNA
methylation subgroups and the CpG island methylator phenotype in
gastric cancer: A comprehensive profiling approach. BMC
Gastroenterol. 14:552014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Feng QY, Hu ZX, Song XL and Pan HW:
Aberrant expression of genes and proteins in pterygium and their
implications in the pathogenesis. Int J Ophthalmol. 10:973–981.
2017.PubMed/NCBI
|
|
11
|
Marhaba R, Klingbeil P, Nuebel T,
Nazarenko I, Buechler MW and Zoeller M: CD44 and EpCAM:
Cancer-initiating cell markers. Curr Mol Med. 8:784–804. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Simon T, Gagliano T and Giamas G: Direct
effects of anti-angiogenic therapies on tumor cells: VEGF
signaling. Trends Mol Med. 23:282–292. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y, Pan K, Wang P, Cao Z, Wang W, Wang
S, Hu N, Xue J, Li H, Jiang W, et al: HBP1-mediated regulation of
p21 protein through the Mdm2/p53 and TCF4/EZH2 pathways and its
impact on cell senescence and tumorigenesis. J Biol Chem.
291:12688–12705. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gupta A, Kaur CD, Jangdey M and Saraf S:
Matrix metalloproteinase enzymes and their naturally derived
inhibitors: Novel targets in photocarcinoma therapy. Ageing Res
Rev. 13:65–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Spitzwieser M, Pirker C, Koblmüller B,
Pfeiler G, Hacker S, Berger W, Heffeter P and Cichna-Markl M:
Promoter methylation patterns of ABCB1, ABCC1 and ABCG2 in human
cancer cell lines, multidrug-resistant cell models and tumor,
tumor-adjacent and tumor-distant tissues from breast cancer
patients. Oncotarget. 7:73347–73369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang ML, Nie FQ, Sun M, Xia R, Xie M, Lu
KH and Li W: HOXA5 indicates poor prognosis and suppresses cell
proliferation by regulating p21 expression in non small cell lung
cancer. Tumour Biol. 36:3521–3531. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ilson DH: Angiogenesis in gastric cancer:
Hitting the target? Lancet. 383:4–6. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mikhail S, Albanese C and Pishvaian MJ:
Cyclin-dependent kinase inhibitors and the treatment of
gastrointestinal cancers. Am J Pathol. 185:1185–1197. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Okimoto RA, Breitenbuecher F, Olivas VR,
Wu W, Gini B, Hofree M, Asthana S, Hrustanovic G, Flanagan J,
Tulpule A, et al: Inactivation of Capicua drives cancer metastasis.
Nat Genet. 49:87–96. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang B, Chen Q, Cao Y, Ma X, Yin C, Jia Y,
Zang A and Fan W: LGR5 is a gastric cancer stem cell marker
associated with stemness and the EMT signature genes NANOG,
NANOGP8, PRRX1, TWIST1, and BMI1. PLoS One. 11:e1689042016.
View Article : Google Scholar
|
|
21
|
Raman V, Martensen SA, Reisman D, Evron E,
Odenwald WF, Jaffee E, Marks J and Sukumar S: Compromised HOXA5
function can limit p53 expression in human breast tumours. Nature.
405:974–978. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nieto MA: Context-specific roles of EMT
programmes in cancer cell dissemination. Nat Cell Biol. 19:416–418.
2017. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Aubin J, Déry U, Lemieux M, Chailler P and
Jeannotte L: Stomach regional specification requires Hoxa5-driven
mesenchymal-epithelial signaling. Development. 129:4075–4087.
2002.PubMed/NCBI
|
|
24
|
Teo WW, Merino VF, Cho S, Korangath P,
Liang X, Wu RC, Neumann NM, Ewald AJ and Sukumar S: HOXA5
determines cell fate transition and impedes tumor initiation and
progression in breast cancer through regulation of E-cadherin and
CD24. Oncogene. 35:5539–5551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Xu L and Jiang L: MiR-1271
promotes non-small-cell lung cancer cell proliferation and invasion
via targeting HOXA5. Biochem Biophys Res Commun. 458:714–719. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang HW, Chung M, Kudo T and Meyer T:
Competing memories of mitogen and p53 signalling control cell-cycle
entry. Nature. 549:404–408. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ratti S, Ramazzotti G, Faenza I, Fiume R,
Mongiorgi S, Billi AM, McCubrey JA, Suh PG, Manzoli L, Cocco L and
Follo MY: Nuclear inositide signaling and cell cycle. Adv Biol
Regul. 67:1–6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Waldman T: The inaugural use of gene
editing for the study of tumor suppressor pathways in human
cells-p21WAF1/CIP1. Cancer Res. 76:4598–4601. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
LaBaer J, Garrett MD, Stevenson LF,
Slingerland JM, Sandhu C, Chou HS, Fattaey A and Harlow E: New
functional activities for the p21 family of CDK inhibitors. Genes
Dev. 11:847–862. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feng F, Ren Q, Wu S, Saeed M and Sun C:
Hoxa5 increases mitochondrial apoptosis by inhibiting
Akt/mTORC1/S6K1 pathway in mice white adipocytes. Oncotarget.
8:95332–95345. 2017. View Article : Google Scholar : PubMed/NCBI
|