Introduction
Tumor metastasis significantly affects the prognosis
of patients with gastric cancer, and is the primary cause of
treatment failure (1). Mechanisms
of tumor metastasis are complex and the tumor microenvironment,
enriched in cytokines, growth factors and tumor cell-derived
vesicles, is key in its pathophysiology. Receptor activator of
nuclear factor-κB ligand (RANKL), an important cytokine belonging
to the tumor necrosis factor (TNF) family, promotes osteoclast
maturation and migration. In addition to being secreted by
osteoclast cells, previous studies have revealed that RANKL is
secreted by infiltrating T cells; whereas RANK is expressed on the
surface of various cancer cells, including breast, renal and lung
cancer cells (2–6). According to our previous study, RANK
is also expressed in gastric cancer cells (7), and infiltrating T cells have been
found to be abundant in gastric cancer tissues (8,9).
Collectively, these studies indicate that RANKL may also promote
gastric cancer cell migration, although there is no supporting data
at present.
Lipid rafts, comprised of assemblies of cholesterol,
sphingolipids and certain types of proteins, form sorting platforms
for targeted proteins (10) and are
essential in a variety of signaling processes, including cell
migration, through the regulation of proteins located in the cell
membrane (11,12). Lipid rafts are reported to be able
to control human melanoma cell migration by regulating focal
adhesion disassembly (13), and
promote breast cancer cell migration by restricting interactions
between CD44 and ezrin (14). A
previous study showed lipid rafts to be critical for RANK functions
in osteoclasts (15). Based on
this, it was hypothesized that lipid rafts may be involved in
RANKL-induced cancer cell migration.
Caveolin-1 (Cav-1), a pivotal component of lipid
rafts, is a membrane-bound scaffolding protein that regulates
signal transduction (16). The role
of Cav-1 in cancer remains controversial; it can regulate a number
of metastatic cancer cells, either negatively or positively. Cav-1
reportedly inhibits cell migration and invasion via the suppression
of epithelial-mesenchymal transition in pancreatic cancer cells
(17), and has been shown to reduce
the metastatic capacity of colon cancer cells (18). By contrast, the expression of Cav-1
appears to be increased in prostate tumors, lung cancer, melanoma
cells and renal cell carcinoma (18–21),
thereby favoring tumor progression and migration (22). RANKL induces the expression of
Cav-1, which is immediately conveyed to lipid rafts to promote
osteoclastogenesis (23).
As there has been no previous study reporting the
effect of Cav-1 on RANKL-induced cell migration, the present study
aimed to identify the potential roles and mechanisms of RANKL/RANK
in gastric cancer cell migration and metastasis. The results
indicated that the proto-oncogene tyrosine-protein kinase Src
(c-Src)/Cav-1 pathway and lipid raft aggregation may be the primary
mechanisms involved in RANKL-induced gastric cancer cell
migration.
Materials and methods
Cell culture
The MGC803, BGC823 and SGC7901 (gastric cancer),
H460 (lung cancer), ACHN (renal cancer) and MDA-MB-231 (breast
cancer) cells were purchased from the Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). MGC803, BGC823 and
SGC7901, H460 and ACHN cells were cultured in Roswell Park Memorial
Institute (RPMI)-1640 medium (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). MDA-MB-231 cells were cultured in L15 medium
(Gibco; Thermo Fisher Scientific, Inc.) RPMI-1640 and L15 media
were supplemented with 10% fetal bovine serum (FBS), penicillin
(100 U/ml) and streptomycin (100 mg/ml) in an atmosphere of 95% air
and 5% CO2 at 37°C.
Cell treatment
We added sRANKL (PeproTech, Inc., Rocky Hill, NJ,
USA) to cancer cells to final concentration of 10 µg/ml for 0, 5,
10, 30 or 60 min. We added 10 µM PP2 (Sigma-Aldrich St. Louis, MO,
USA) or Nystatin (50 µg/ml; cat. no. N3503; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany and/or its affiliates) 1 h prior to
sRANKL. To detect the lipid raft aggregation, we used CTXB (1
mg/ml; cat. no. SAE0069-500UG; Sigma-Aldrich; Merck KGaA).
Western blot analysis
Western blot analysis was performed as previously
described (24). The following
antibodies were used: Anti-phospho-Scr (1:250; rabbit monoclonal;
cat. no. 6943S; Cell Signaling Technology, Danvers, MA, USA),
anti-Scr (1:1,000; mouse monoclonal; cat. no. 2110s; Cell Signaling
Technology), anti-phospho-Cav-1 (1:250; rabbit polyclonal; cat. no.
3251s; Cell Signaling Technology), anti-Cav-1 (1:1,000; rabbit
polyclonal; cat. no. sc-894; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA), anti-phospho-Akt (1:500; rabbit polyclonal; cat.
no. 9271L; Cell Signaling Technology), anti-Akt (1:1,000; rabbit
polyclonal; cat. no. 9272S; Cell Signaling Technology),
anti-phospho-ERK1/2 (1:500; rabbit polyclonal; cat. no. sc-16982;
Santa Cruz Biotechnology), anti-ERK1/2 (1:1,000; rabbit polyclonal;
cat. no. 9102S; Santa Cruz Biotechnology), anti-RANK (1:500; rabbit
polyclonal; cat. no. A303-897A; Bethyl Laboratories, Inc.,
Montgomery, TX, USA), anti-β-actin (1:1,000; rabbit polyclonal;
cat. no. sc-1616-R; Santa Cruz Biotechnology), followed by
incubation with appropriate secondary antibodies. Secondary goat
anti-rabbit (1:1,000) and goat anti-mouse antibodies were purchased
from Santa Cruz Biotechnology, Inc.
Transwell assay
The cells were pretreated with appropriate solvent
control (dimethyl sulfoxide) or various concentrations of
inhibitors (PP2: 10 µM; Nystatin: 50 µg/ml) for 60 min in
serum-free media. The treated cells were plated in the upper insert
of a 24-well chemotaxis chamber (2×104 cells/well; 8-µm
pore size; Corning Inc., Corning, NY, USA) in serum-free medium.
Medium containing 2.5% serum (0.5 ml) and recombinant RANKL (1 µ1),
with DMSO or inhibitors, was added to the bottom well and incubated
for 24 h. The porous inserts were carefully removed, and the cells
was stained and counted at ×200 magnification (Olympus Corp.,
Tokyo, Japan) in at least five different fields of each filter.
Fluorescence microscopy
The MGC803 cells were first treated with PP2 or
nystatin for 1 h, and then RANKL was added at a final concentration
of 1 µg/ml for 10 min. The cells were fixed in 4.4%
paraformaldehyde for 20 min, permeabilized with 0.2% Triton X-100
for 15 min, and then blocked with 5% bovine serum albumin (BSA;
Sigma-Aldrich, Merck KGaA) for 1 h. The slides were incubated with
CTXB antibody or anti-RANK antibody for 1 h and then with
FITC-conjugated goat anti-mouse or anti-rabbit IgG were added for 1
h. Images were captured with a fluorescence microscope (Olympus
Corp.).
Surface RANK expression analysis
Surface RANK expression was determined by flow
cytometry as previously described (24). The following antibodies were used:
Mouse anti-RANK (1:500; mouse monoclonal; cat. no. MAB683; R&D
Systems, Minneapolis, MN, USA) or isotype control (R&D
Systems), FITC-conjugated anti-mouse secondary antibody (1:200;
mouse monoclonal; cat. no. sc-2356; Santa Cruz Biotechnology).
Transfection with small interfering
(si)RNA
The cells were cultivated at a density of
2×105/well in 6-well plates. After 24 h, the cells were
transfected with siRNA using Lipofectamine™ 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The CAV-1 siRNAs were designed to target
the sequence 5′-AACCAGAAGGGACACACAGTT-3′. The cells were treated
with or without RANKL at 48 h post-transfection. The gene silencing
effect was evaluated by western blot analysis.
Patients and tissue samples
Specimens of gastric adenocarcinoma tissue were
collected from 228 patients who underwent surgical resection at the
First Hospital of China Medical University (Shenyang, China) from
March 2006 to October 2011. None of the patients had received
operative radiotherapy, chemotherapy or immunotherapy previously.
Age, sex, pathological tumor-node-metastasis (pTNM) stage and
Lauren grade were evaluated following medical charts and
pathological records. The pTNM stage was examined according to the
seventh edition of the AJCC Cancer Staging Manual (25). The Lauren grade was assigned
according to the classification of the World Health Organization.
The First Hospital of China Medical University Ethical Committee
approved the study, and no consent was required due to the
retrospective nature of the study.
Immunohistochemistry
Formalin-fixed paraffin-embedded tumor specimens
were collected from the Department of Pathology at the First
Hospital of China Medical University. The immunohistochemical
staining observed with Olympus microscope (Olympus Corp.) was
performed using the biotinstreptavidin method (UltraSensitive S-P
kit; MaixinBio, Shanghai, China) as previously described (26). Two observers, who had no prior
information of the clinical or pathological parameters, performed
the evaluation of results independently. The immunoreactivity was
scored based on the intensity of staining (negative, 0; weak, 1;
moderate, 2; strong, 3).
Statistical analysis
The experimental data are summarized and presented
as the mean ± standard deviation. The significance of differences
was analyzed statistically using Student's two-tailed t-test,
P<0.05 was considered to indicate a statistically significant
difference. Each experiment was repeated at least three times.
Statistical analyses were performed using the SPSS statistical
package software (SPSS for Windows, version 20.0; IBM Corp.,
Armonk, NY, USA).
Results
RANKL induces the migration of gastric
cancer cells via phosphoinositide 3-kinase (PI3K)/Akt and ERK
pathways
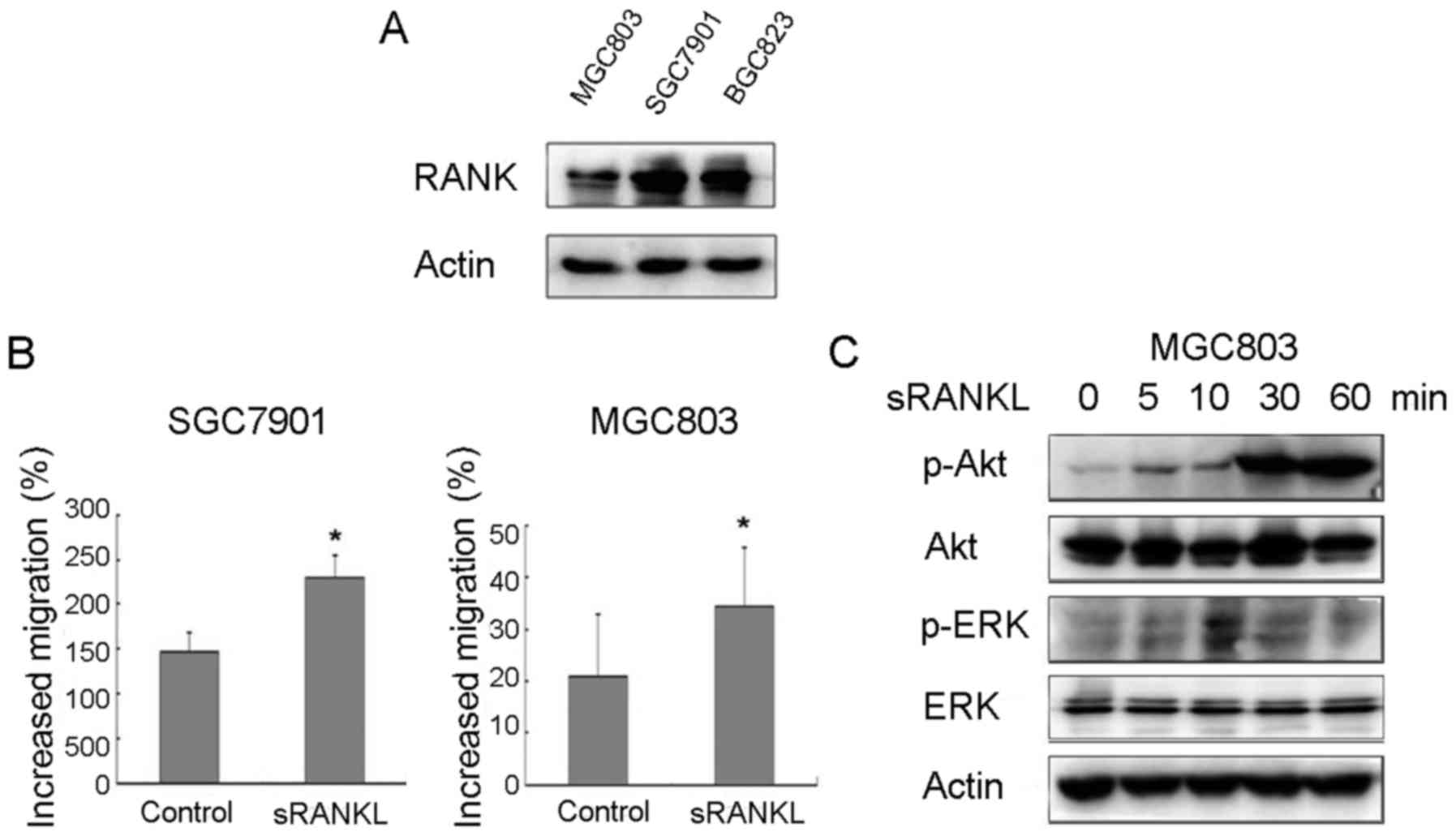
The western blot analysis revealed the expression of
RANK in MGC803, BGC823 and SGC7901 cell lines. Stimulation of the
MGC803 and SGC7901 cells with 1.0 µg/ml RANKL significantly
increased cell migration by 63.8 and 56.3%, respectively (Fig. 1B). As RANKL had no effect on the
proliferation of MGC803 or SGC7901 cells (data not shown), the
increased number of MGC803 and SGC7901 cells traversing the filter
may have resulted from increased migratory abilities. The
downstream signaling of RANKL/RANK was also examined in BGC803
cells; Akt and ERK were markedly increased in response to RANKL
treatment (Fig. 1C). Therefore, the
RANKL/RANK pathway appeared to be significantly involved in the
migration of gastric cancer cells.
Lipid rafts are involved in
RANKL-induced migration
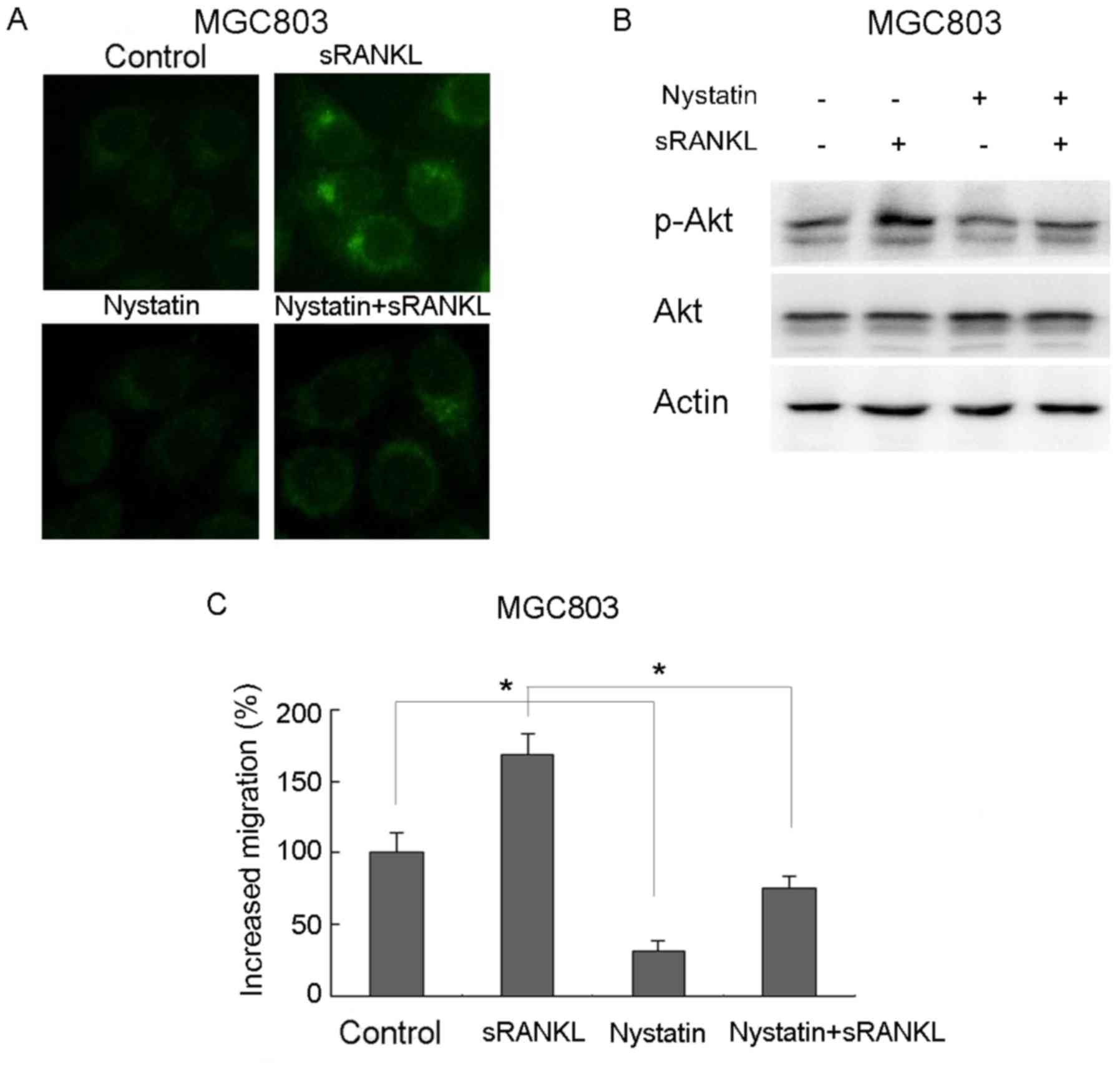
Lipid rafts represent a major platform for signaling
regulation in cancer. To examine the involvement of lipid rafts in
RANKL-induced gastric cancer cell migration, the MGC803 cells were
pretreated with nystatin, a lipid raft inhibitor, for 1 h, followed
by RANKL treatment for 10 min. The immunofluorescence indicated
that RANKL significantly induced lipid raft aggregation, which was
reversed by nystatin (Fig. 2A).
Downstream signals, including the activation of Akt, were also
markedly promoted by RANKL, but were decreased by pretreatment with
nystatin (Fig. 2B). Nystatin also
decreased RANKL-induced gastric cancer cell migration from 168.8 to
75.6% (Fig. 2C). These results
suggested that the aggregation of lipid rafts was associated with
RANKL-induced gastric cancer cell migration.
Cav-1 promotes the migration of
RANKL-induced gastric cancer cells via interactions with RANK
To investigate the effect of Cav-1 on gastric cancer
cell migration, the activation of Cav-1 was examined. The results
showed that RANKL not only activated Cav-1 in a time-dependent
manner (Fig. 3A), but also
triggered an interaction between RANK and Cav-1 (Fig. 3B). The knockdown of Cav-1 by siRNA
suppressed RANKL-induced lipid raft aggregation, accompanied by a
decrease in the activation of Akt and ERK in MGC803 cells (Fig. 3C and D). Cav-1 knockdown also
significantly reduced RANKL-induced gastric cancer cell migration
from 176.2 to 18.5% (Fig. 3E).
These results suggested that Cav-1 promoted RANKL-induced gastric
cancer cell migration via interactions with RANK.
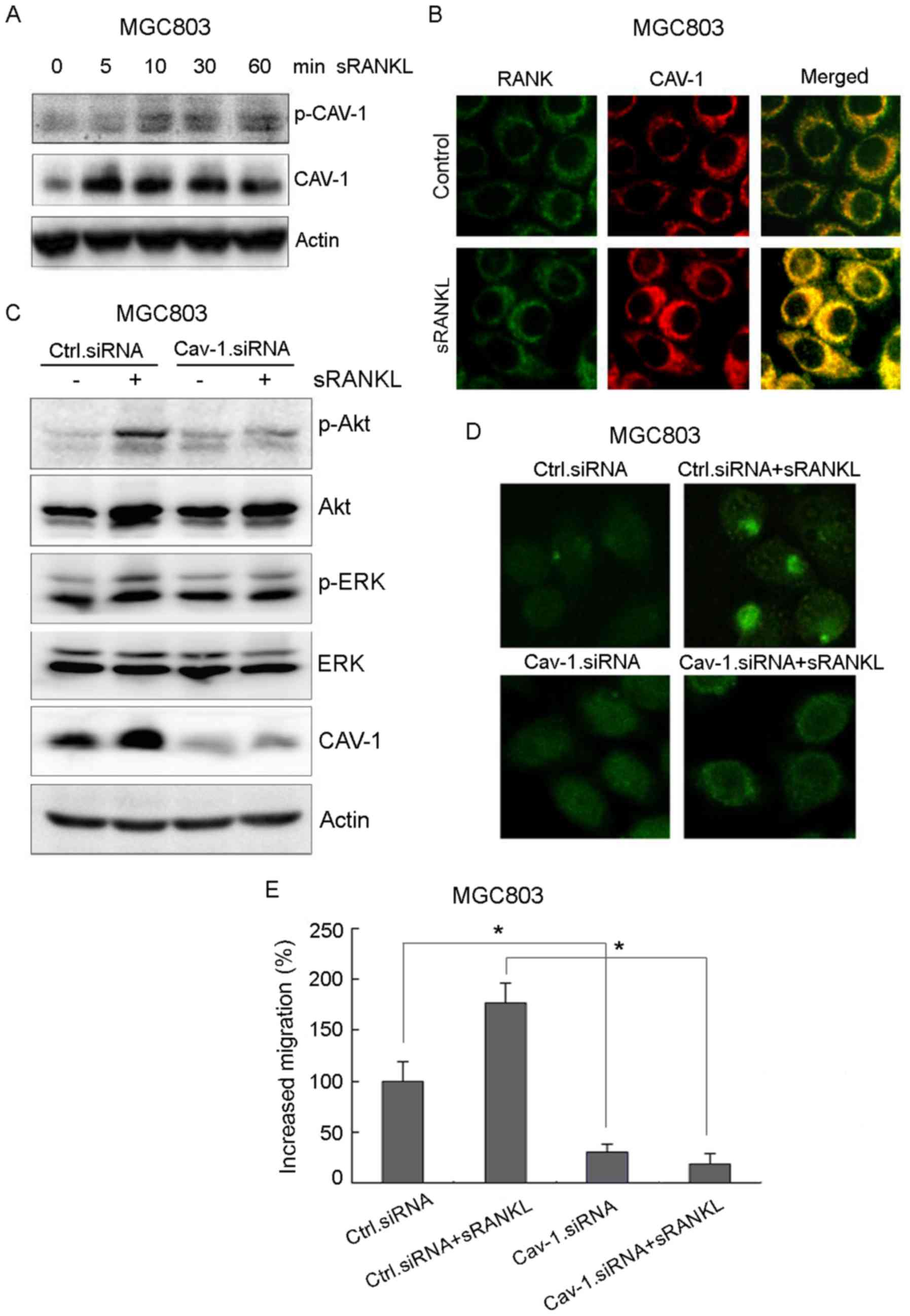 | Figure 3.Cav-1 promotes RANKL-induced gastric
cancer cell migration via interaction with RANK. (A) MGC803 cells
were treated with 1 µg/ml recombinant RANKL at indicated times, and
the activation of Cav-1, Akt and ERK was examined by western blot
analysis. (B) MGC803 cells were treated with 1 µg/ml recombinant
RANKL for 10 min, and the interaction between Cav-1 and RANK was
analyzed by immunofluorescence at high magnification (×40). RANK
and Cav-1 were indicated as green and red respectively. (C) Cav-1
siRNA or control siRNA were transfected into MGC803 cells. Lipid
raft status was analyzed by immunofluorescence following incubation
with CTXB (magnification, ×40). (D) Cav-1 siRNA or control siRNA
transfected cells were treated with 1 µg/ml recombinant RANKL for
10 min, and the activation of Cav-1, Akt and ERK was examined by
western blot analysis. (E) Migration activity of MGC803 cells was
measured with a Transwell assay following treatment with 1 µg/ml
recombinant RANKL for 24 h. Error bars represent the standard
deviation of three independent experiments. *P<0.05, vs.
corresponding control cells (Student's t-test). RANK, receptor
activator of nuclear factor-κB; sRANKL, soluble RANK ligand; Cav-1,
caveolin-1; ERK, extracellular signal-regulated kinase; siRNA,
small interfering RNA; p-, phosphorylated; Ctrl, control. |
RANKL induces the activity of
caveolin-1 via c-Src
To characterize the downstream mechanisms occurring
due to the activation of Cav-1, the cells were incubated with RANKL
over different periods of time and examined for the activation of
c-Src. As shown in Fig. 4A, c-Src
was rapidly activated and reached a peak at 10 min. The c-Src
inhibitor PP2 inhibited the activation of Cav-1 and Akt/ERK
(Fig. 4A). The immunofluorescence
and Transwell experiments revealed that PP2 significantly
suppressed lipid raft aggregation and RANKL-induced migration
(Fig. 4B and C). Collectively,
these results suggested that the c-Src-mediated activation of Cav-1
promoted RANKL-induced gastric cancer cell migration.
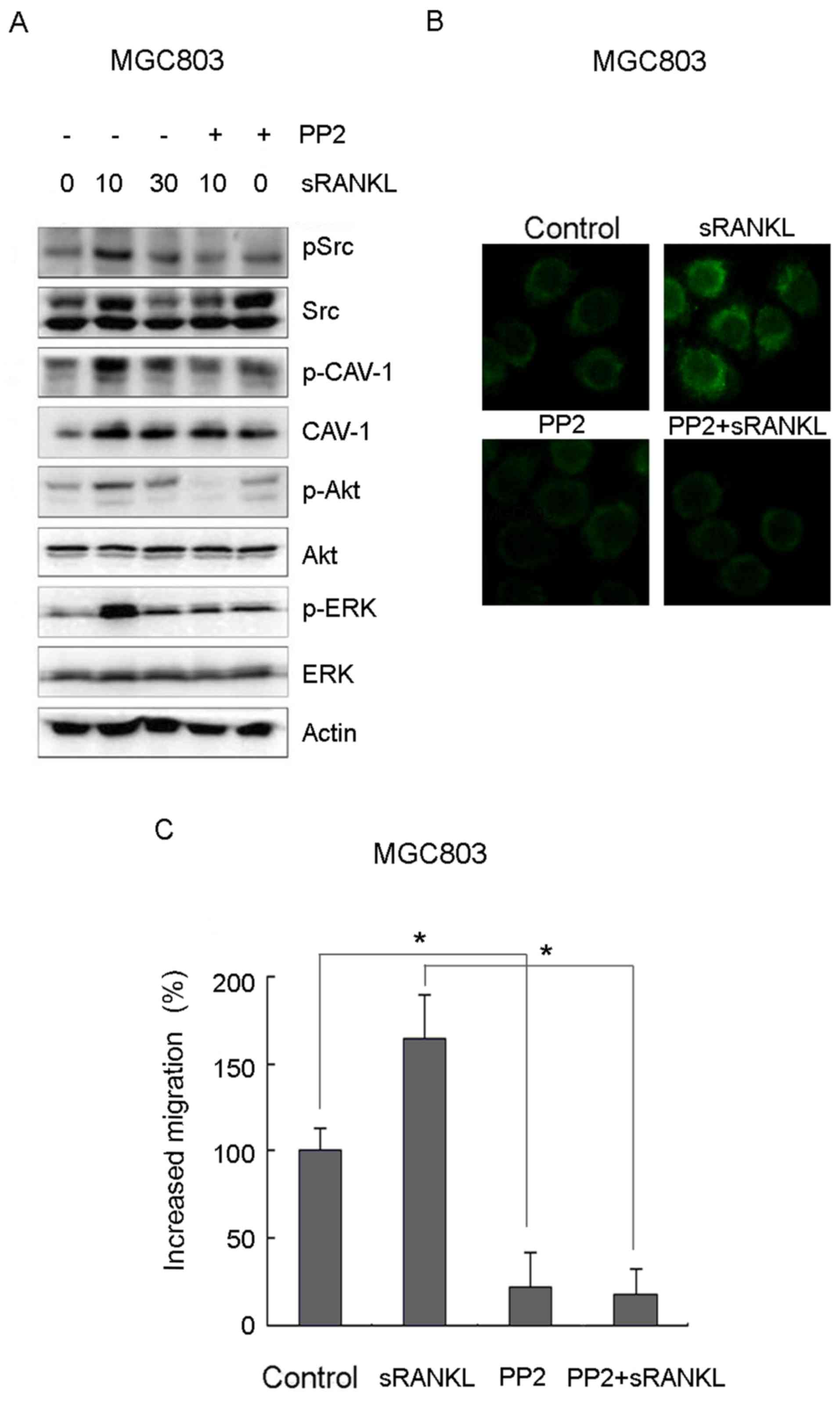 | Figure 4.Src-mediated activation of Cav-1
promotes RANKL-induced gastric cancer cell migration. (A) MGC803
cells were pretreated with 10 µM PP2 or control for 1 h, following
incubation with 1 µg/ml recombinant RANKL for the indicated times.
The expression levels of pSrc/Src, pCav-1/Cav-1, pAkt/Akt, pERK/ERK
were examined by western blot analysis. (B) MGC803 cells were
pretreated with or without 10 µM PP2 for 1 h, and then treated with
or without 1 µg/ml recombinant RANKL for 10 min. Lipid raft status
was observed by immunofluorescence at high magnification (×40). (C)
Cell migration was examined by Transwell assays. Error bars
represent the standard deviation. Data are representative of three
independent experiments. *P<0.05, vs. corresponding control
cells (Student's t-test). RANK, receptor activator of nuclear
factor-κB; sRANKL, soluble RANK ligand; Cav-1, caveolin-1; ERK,
extracellular signal-regulated kinase; p-, phosphorylated. |
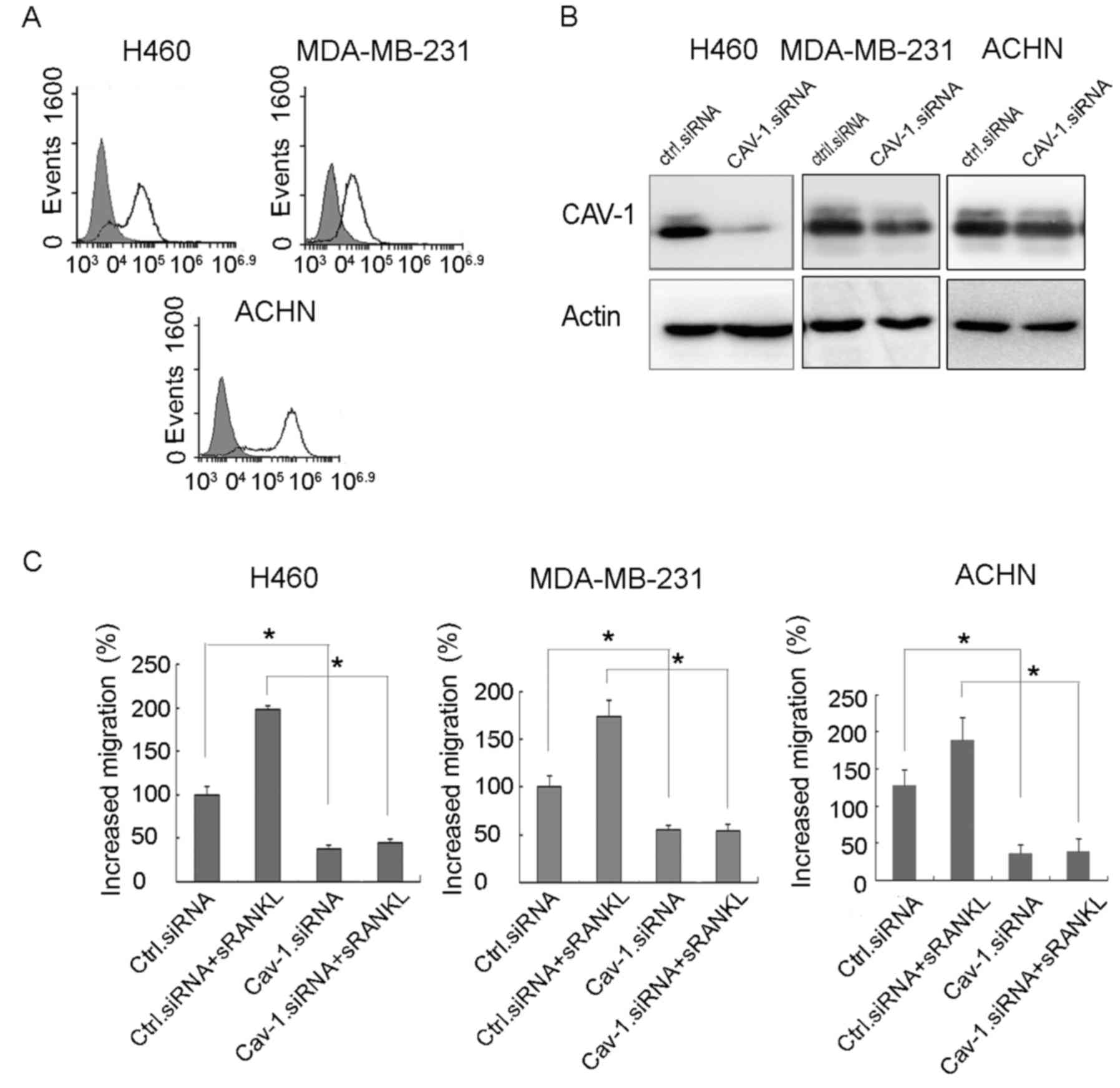
RANKL-induced migration is suppressed
by Cav-1 knockdown
The expression of RANK was examined in a variety of
cancer cells by flow cytometry. The results showed that H460 (lung
cancer), ACHN (renal cancer) and MDA-MB-231 (breast cancer) cells
expressed RANK on their surface (Fig.
5A). The knockdown of Cav-1 by siRNA significantly suppressed
RANKL-induced migration of the cancer cells (Fig. 5B and C).
Cav-1 is independently a poor
predictive factor for the overall survival rate of patients with
gastric cancer
To examine the association between RANK and Cav-1,
228 histologically confirmed gastric cancer samples were selected
for investigation. The follow-up time ranged between 3 and 83
months, with a mean follow-up time of 38 months. The immunostaining
confirmed that Cav-1 was expressed in 56.5% of patients (Table I), whereas 47.4% were positive for
RANK. The correlation between the expression of RANK or Cav-1 and
patient characteristics is shown in Table I. The expression of RANK, observed
in 58.3% of the diffuse patients, was correlated with Lauren
classification. The prognostic value of Cav-1 in patients with
RANK-positive cells was also analyzed. Within this population, a
higher expression of Cav-1 was correlated with poor survival rate
(P=0.025), as the mean overall survival rate of patients was 45
months in the Cav-1-positive arm, compared with 64 months in the
Cav-1-negative arm (Fig. 6). In
patients with RANK-positive cells, univariate analysis revealed
that the positive expression of Cav-1, T stage, N stage and pTNM
stage indicated poor prognosis. The multivariate analysis indicated
that Cav-1, T stage and N stage were independent predictors for
patients with RANK-positive cells (Table II). These results demonstrated that
the expression of Cav-1 was predictive of poor prognosis in
patients with RANK-positive gastric cancer cells.
 | Table I.Correlation of the expression of RANK
and Cav-1 with clinic-pathological parameters in 228 patients with
gastric cancer. |
Table I.
Correlation of the expression of RANK
and Cav-1 with clinic-pathological parameters in 228 patients with
gastric cancer.
|
|
| RANK | Cav-1 |
|---|
|
|
|
|
|
|---|
| Factor | n | Negative (%) | Positive (%) | P-value | Negative (%) | Positive (%) | P-value |
|---|
| Number | 228 | 120 (52.6) | 108 (47.4) |
| 74 (43.5) | 154 (56.5) |
|
| Age (years) |
|
|
| 0.111 |
|
| 0.089 |
| ≤60 | 107 | 50 (46.7) | 57 (53.3) |
| 41 (38.3) | 66 (61.7) |
|
| >60 | 121 | 70 (57.9) | 51 (42.1) |
| 33 (27.3) | 88 (72.7) |
|
| Sex |
|
|
| 0.307 |
|
| 0.439 |
| Male | 162 | 89 (54.9) | 73 (45.1) |
| 50 (30.9) | 112 (69.1) |
|
| Female | 66 | 31 (47.0) | 35 (53.0) |
| 24 (36.4) | 42 (63.6) |
|
| T stage |
|
|
| 0.500 |
|
| 0.714 |
| T1 | 2 | 0 (0) | 2 (100) |
| 1 (50.0) | 1 (50.0) |
|
| T2 | 18 | 10 (55.6) | 8 (44.4) |
| 4 (22.2) | 14 (77.8) |
|
| T3 | 36 | 17 (47.2) | 19 (52.8) |
| 12 (33.3) | 24 (66.7) |
|
| T4 | 172 | 93 (54.1) | 79 (45.9) |
| 57 (33.1) | 115 (66.9) |
|
| N stage |
|
|
| 0.869 |
|
| 0.149 |
| N1 | 51 | 27 (52.9) | 24 (47.1) |
| 19 (37.3) | 32 (62.7) |
|
| N2 | 36 | 21 (58.3) | 15 (41.7) |
| 14 (38.9) | 22 (61.1) |
|
| N3 | 47 | 25 (53.2) | 22 (46.8) |
| 9 (19.1) | 38 (80.9) |
|
| N4 | 94 | 47 (50.0) | 47 (50.0) |
| 32 (34.0) | 62 (66.0) |
|
| pTNM stage |
|
|
| 0.540 |
|
| 0.323 |
| I+II | 55 | 31 (56.4) | 24 (43.6) |
| 21 (38.2) | 34 (61.8) |
|
| III+IV | 173 | 89 (51.4) | 84 (48.6) |
| 53 (30.6) | 120 (69.4) |
|
| Lauren grade |
|
|
|
<0.001 |
|
| 0.059 |
| Intestinal | 89 | 62 (69.7) | 27 (30.3) |
| 21 (23.6) | 68 (76.4) |
|
| Diffuse | 98 | 35 (35.7) | 63 (64.3) |
| 39 (39.8) | 59 (60.2) |
|
| Mixed | 41 | 23 (56.1) | 18 (43.9) |
| 14 (34.1) | 27 (65.9) |
|
| Location |
|
|
| 0.672 |
| Cardia | 28 | 15 (53.6) | 13 (46.4) |
| 10 (35.7) | 18 (64.3) |
|
| Body | 20 | 13 (65.0) | 7 (35.0) |
| 6 (30.0) | 14 (70.0) |
|
| Antrum | 147 | 74 (50.3) | 73 (49.7) |
| 46 (31.3) | 101 (68.7) |
|
| Other | 33 | 18 (54.5) | 15 (45.5) |
| 12 (36.4) | 21 (63.6) |
|
| Histological
classification |
|
|
|
<0.001 |
|
| 0.023 |
| Well | 12 | 8 (66.7) | 4 (33.3) |
| 3 (25.0) | 9 (75.0) |
|
| Moderate | 75 | 53 (70.7) | 22 (29.3) |
| 16 (21.3) | 59 (78.7) |
|
| Poor | 141 | 59 (41.8) | 82 (58.2) |
| 55 (39.0) | 86 (61.0) |
|
| Table II.Cox univariate and multivariate
analyses of overall survival in patients with receptor activator of
nuclear factor-κB-positive gastric cancer (n=228). |
Table II.
Cox univariate and multivariate
analyses of overall survival in patients with receptor activator of
nuclear factor-κB-positive gastric cancer (n=228).
|
| Univariate | Multivariate |
|---|
|
|
|
|
|---|
| Biomarker | Hazard | 95% CI | P-value | Hazard | 95% CI | P-value |
|---|
| Age | 1.489 | 0.876–2.530 | 0.142 |
|
|
|
| T stage | 2.812 | 1.410–5.609 | 0.003 | 2.559 | 1.292–5.065 | 0.007 |
| N stage | 1.518 | 1.176–1.960 | 0.001 | 1.496 | 1.156–1.936 | 0.002 |
| pTNM stage | 3.688 | 1.468–9.263 | 0.005 |
|
|
|
| Lauren | 1.102 | 0.738–1.645 | 0.635 |
|
|
|
| Caveolin-1 | 2.392 | 1.082–5.289 | 0.031 | 2.603 | 1.174–5.773 | 0.019 |
Discussion
The RANKL/RANK pathway is a classical pathway for
osteoclast maturation and activation, whereby RANKL interacts with
RANK to recruit TNF-receptor associated factor, resulting in the
activation of nuclear factor-FB, c-Jun N-terminal kinase, p38, ERK
and Akt (27–29). In breast, lung and prostate cancer
cells, the inhibition of PI3K and mitogen-activated protein kinase
kinase 1/2 can reduce RANKL-induced migration (30–32).
According to the results of the present study, RANK was expressed
in gastric cancer cells. Furthermore, RANKL significantly increased
the migration ability of gastric cancer cells, accompanied by the
activation of Akt and ERK. As gastric cancer tissues are enriched
in infiltrating T cells capable of secreting RANKL, RANKL-induced
migration may represent a pivotal mechanism for gastric cancer
metastasis. Drugs, including denosumab, which target the RANKL/RANK
pathway, likely inhibit this process and can be potentially used as
novel therapeutic intervention for treating metastatic gastric
cancer.
Previous studies have provided evidence in support
of the involvement of lipid rafts in cancer cell invasion and
metastasis (33–35). Yamaguchi et al reported the
requirement of lipid rafts for invadopodia formation and
extracellular matrix degradation in human breast cancer cells
(36). Chinni et al showed
that C-X-C motif chemokine ligand 12/C-X-C chemokine receptor type
4 transactivates human epidermal growth factor receptor 2 in lipid
rafts to promote prostate cancer cell migration (37). In the present study, the finding
that RANKL triggered lipid raft aggregation, which was reversed by
nystatin, and reduced RANKL-induced migration in gastric cancer
cells indicated the importance of lipid rafts in gastric cancer
cell migration. Lipid rafts are known to be regulated by other
important factors, including Cav-1. Cav-1 can also result in
further clustering of lipid rafts mediated by the activation of
several downstream signaling pathways (36,38).
In the present study, Cav-1 was shown to be involved in
RANKL-induced lipid raft aggregation and cell migration. It was
confirmed that certain RANK-expressing gastric cancer cells also
express Cav-1, which was significantly correlated with the poor
prognosis in individuals with RANK-positive cells. Univariate and
multivariate analyses demonstrated that the expression of Cav-1 was
an independent predictor of poor overall survival rate in these
patients. Furthermore, the involvement of Cav-1 in RANKL-induced
cell migration was confirmed in several cancer cell lines. These
findings indicated that Cav-1 is essential not only for appropriate
RANK-localization within the lipid raft, but also for RANKL-induced
lipid raft aggregation and cancer cell migration.
Although the data obtained in the present study
revealed that Cav-1 was rapidly activated by RANKL, the question
regarding the key mediator remains unanswered. The tyrosine protein
kinase c-Src is known to be involved in the regulation of cellular
metabolism, survival and proliferation. In cancer cells, the
activation of c-Src results in increased tumor progression,
invasion and metastasis (39–42).
Furthermore, RANKL has shown potential in activating c-Src in
breast cancer cells (30). Previous
reports have suggested that the interaction between Cav-1 and
Rho-GTPases promotes metastasis by controlling the activation of
c-Src, Ras and Erk (43). In the
present study, the activation of Cav-1 accompanied that of c-Src.
In addition, the activation of Cav-1, lipid raft aggregation and
cell migration were almost completely reversed by the PP2-mediated
inhibition of c-Src function, which is an important regulator in
several signaling pathways (44).
These results suggested that the c-Src-mediated activation of Cav-1
promoted RANKL-induced gastric cancer cell migration.
In conclusion, RANKL-induced gastric cancer cell
migration is at least partially dependent on lipid rafts and its
main component, Cav-1, and is promoted by the activation of c-Src
and Cav-1. These findings demonstrate a detailed mechanism
underlying the effect of RANK on gastric cancer cell migration.
This may shed light on the potential drug targets for novel
treatment of metastatic gastric cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Science and Technology Major Project of the Ministry of Science and
Technology of China (grant no. 2017ZX09304025), the National
Natural Science Foundation of China (grant nos. 81572374 and
81302128), the Liaoning BaiQianWan Talents Program (grant no.
2014921032), the General Project of Liaoning Province Department of
Education (grant no. LZ2015073), the Foundation for Selected
Overseas Chinese Scholar 2015 Science and Technology Plan Project
of Liaoning Province (grant nos. 2016007010 and 2015020457) and The
Key Research and Development Program of Shenyang (grant no.
17-230-9-01).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YW, YL and XQ conceived and designed the study. YW,
QW, XZ, LZ, JQ, ZL, LX, YZ, KH, YF and XC performed the
experiments. YS provided the samples and collected the patient
information. XC and YW contributed in the statistical analysis. YW
wrote the manuscript. XQ, YL and XC reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The First Hospital of China Medical University
Ethical Committee approved the study. No consent was required due
to the retrospective nature of the study.
Patient consent for publication
No consent was required due to the retrospective
nature of the study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mikami S, Katsube K, Oya M, Ishida M,
Kosaka T, Mizuno R, Mochizuki S, Ikeda T, Mukai M and Okada Y:
Increased RANKL expression is related to tumour migration and
metastasis of renal cell carcinomas. J Pathol. 218:530–539. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jones DH, Nakashima T, Sanchez OH,
Kozieradzki I, Komarova SV, Sarosi I, Morony S, Rubin E, Sarao R,
Hojilla CV, et al: Regulation of cancer cell migration and bone
metastasis by RANKL. Nature. 440:692–696. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen LM, Kuo CH, Lai TY, Lin YM, Su CC,
Hsu HH, Tsai FJ, Tsai CH, Huang CY and Tang CH: RANKL increases
migration of human lung cancer cells through intercellular adhesion
molecule-1 up-regulation. J Cell Biochem. 112:933–941. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rao S, Cronin SJF, Sigl V and Penninger
JM: RANKL and RANK: From mammalian physiology to cancer treatment.
Trends Cell Biol. 28:213–223. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tan W, Zhang W, Strasner A, Grivennikov S,
Cheng JQ, Hoffman RM and Karin M: Tumour-infiltrating regulatory T
cells stimulate mammary cancer metastasis through RANKL-RANK
signalling. Nature. 470:548–553. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang X, Song Y, Song N, Zhang Y, Zhang L,
Wang Y, Wang Z, Qu X and Liu Y: RANKL/RANK pathway abrogates
cetuximab sensitivity in gastric cancer cells via activation of
EGFR and c-Src. Onco Targets Ther. 10:73–83. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Choi HS, Ha SY, Kim HM, Ahn SM, Kang MS,
Kim KM, Choi MG, Lee JH, Sohn TS, Bae JM, et al: The prognostic
effects of tumor infiltrating regulatory T cells and myeloid
derived suppressor cells assessed by multicolor flow cytometry in
gastric cancer patients. Oncotarget. 7:7940–7951. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mao F, Kong H, Zhao YL, Peng LS, Chen W,
Zhang JY, Cheng P, Wang TT, Lv YP, Teng YS, et al: Increased
tumor-infiltrating CD45RA-CCR7-regulatory T-cell subset with
immunosuppressive properties foster gastric cancer progress. Cell
Death Dis. 8:e30022017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lingwood D and Simons K: Lipid rafts as a
membrane-organizing principle. Science. 327:46–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Babina IS, McSherry EA, Donatello S, Hill
AD and Hopkins AM: A novel mechanism of regulating breast cancer
cell migration via palmitoylation-dependent alterations in the
lipid raft affiliation of CD44. Breast Cancer Res. 16:R192014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin BJ, Tsao SH, Chen A, Hu SK, Chao L and
Chao PG: Lipid rafts sense and direct electric field-induced
migration. Proc Natl Acad Sci USA. 114:8568–8573. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang R, Bi J, Ampah KK, Ba X, Liu W and
Zeng X: Lipid rafts control human melanoma cell migration by
regulating focal adhesion disassembly. Biochim Biophys Acta.
1833:3195–3205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Donatello S, Babina IS, Hazelwood LD, Hill
AD, Nabi IR and Hopkins AM: Lipid raft association restricts
CD44-ezrin interaction and promotion of breast cancer cell
migration. Am J Pathol. 181:2172–2187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ha H, Kwak HB, Lee SK, Na DS, Rudd CE, Lee
ZH and Kim HH: Membrane rafts play a crucial role in receptor
activator of nuclear factor kappaB signaling and osteoclast
function. J Biol Chem. 278:18573–18580. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Parton RG and Simons K: The multiple faces
of caveolae. Nat Rev Mol Cell Biol. 8:185–194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Salem AF, Bonuccelli G, Bevilacqua G,
Arafat H, Pestell RG, Sotgia F and Lisanti MP: Caveolin-1 promotes
pancreatic cancer cell differentiation and restores membranous
E-cadherin via suppression of the epithelial-mesenchymal
transition. Cell Cycle. 10:3692–3700. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nimri L, Barak H, Graeve L and Schwartz B:
Restoration of caveolin-1 expression suppresses growth,
membrane-type-4 metalloproteinase expression and
metastasis-associated activities in colon cancer cells. Mol
Carcinog. 52:859–870. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L, Ren C, Yang G, Goltsov AA, Tabata K
and Thompson TC: Caveolin-1 promotes autoregulatory, Akt-mediated
induction of cancer-promoting growth factors in prostate cancer
cells. Mol Cancer Res. 7:1781–1791. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fecchi K, Travaglione S, Spadaro F,
Quattrini A, Parolini I, Piccaro G, Raggi C, Fabbri A, Felicetti F,
Caré A, et al: Human melanoma cells express FGFR/Src/Rho signaling
that entails an adhesion-independent caveolin-1 membrane
association. Int J Cancer. 130:1273–1283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Joo HJ, Oh DK, Kim YS, Lee KB and Kim SJ:
Increased expression of caveolin-1 and microvessel density
correlates with metastasis and poor prognosis in clear cell renal
cell carcinoma. BJU Int. 93:291–296. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu W, Yin NC, Liu H and Nan KJ: Cav-1
promote lung cancer cell proliferation and invasion through lncRNA
HOTAIR. Gene. 641:335–340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hada N, Okayasu M, Ito J, Nakayachi M,
Hayashida C, Kaneda T, Uchida N, Muramatsu T, Koike C, Masuhara M,
et al: Receptor activator of NF-κB ligand-dependent expression of
caveolin-1 in osteoclast precursors, and high dependency of
osteoclastogenesis on exogenous lipoprotein. Bone. 50:226–236.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song N, Liu S, Zhang J, Liu J, Xu L, Liu Y
and Qu X: Cetuximab-induced MET activation acts as a novel
resistance mechanism in colon cancer cells. Int J Mol Sci.
15:5838–5851. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Washington K: 7th edition of the AJCC
cancer staging manual: Stomach. Ann Surg Oncol. 17:3077–3079. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang L, Teng Y, Fan Y, Wang Y, Li W, Shi
J, Ma Y, Li C, Shi X, Qu X and Liu Y: The E3 ubiquitin ligase Cbl-b
improves the prognosis of RANK positive breast cancer patients by
inhibiting RANKL-induced cell migration and metastasis. Oncotarget.
6:22918–22933. 2015.PubMed/NCBI
|
|
27
|
Darnay BG, Ni J, Moore PA and Aggarwal BB:
Activation of NF-kappaB by RANK requires tumor necrosis factor
receptor-associated factor (TRAF) 6 and NF-kappaB-inducing kinase.
Identification of a novel TRAF6 interaction motif. J Biol Chem.
274:7724–7731. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee SY, Reichlin A, Santana A, Sokol KA,
Nussenzweig MC and Choi Y: TRAF2 is essential for JNK but not
NF-kappaB activation and regulates lymphocyte proliferation and
survival. Immunity. 7:703–713. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen T and Feng X: Cell-based assay
strategy for identification of motif-specific RANK signaling
pathway inhibitors. Assay Drug Dev Technol. 4:473–482. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang L, Teng Y, Zhang Y, Liu J, Xu L, Qu
J, Hou K, Yang X, Liu Y and Qu Y: C-Src-mediated RANKL-induced
breast cancer cell migration by activation of the ERK and Akt
pathway. Oncol Lett. 3:395–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nakamura ES, Koizumi K, Kobayashi M,
Saitoh Y, Arita Y, Nakayama T, Sakurai H, Yoshie O and Saiki I:
RANKL-induced CCL22/macrophage-derived chemokine produced from
osteoclasts potentially promotes the bone metastasis of lung cancer
expressing its receptor CCR4. Clin Exp Metastasis. 23:9–18. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mori K, Le Goff B, Charrier C, Battaglia
S, Heymann D and Redini F: DU145 human prostate cancer cells
express functional receptor activator of NFkappaB: New insights in
the prostate cancer bone metastasis process. Bone. 40:981–990.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Baillat G, Siret C, Delamarre E and Luis
J: Early adhesion induces interaction of FAK and Fyn in lipid
domains and activates raft-dependent Akt signaling in SW480 colon
cancer cells. Biochim Biophys Acta. 1783:2323–2331. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Babina I, McSherry EA, Donatello S, Hill
AD and Hopkins AM: A novel mechanism of regulating breast cancer
cell migration via palmitoylation-dependent alterations in the
lipid raft affiliation of CD44. Breast Cancer Res. 16:R192014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chinni SR, Sivalogan S, Dong Z, Filho JC,
Deng X, Bonfil RD and Cher ML: CXCL12/CXCR4 signaling activates
Akt-1 and MMP-9 expression in prostate cancer cells: The role of
bone microenvironment-associated CXCL12. Prostate. 66:32–48. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yamaguchi H, Takeo Y, Yoshida S, Kouchi Z,
Nakamura Y and Fukami K: Lipid rafts and caveolin-1 are required
for invadopodia formation and extracellular matrix degradation by
human breast cancer cells. Cancer Res. 69:8594–8602. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chinni SR, Yamamoto H, Dong Z, Sabbota A,
Bonfil RD and Cher ML: CXCL12/CXCR4 transactivates HER2 in lipid
rafts of prostate cancer cells and promotes growth of metastatic
deposits in bone. Mol Cancer Res. 6:446–457. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Patra SK: Dissecting lipid raft
facilitated cell signaling pathways in cancer. Biochim Biophys
Acta. 1785:182–206. 2008.PubMed/NCBI
|
|
39
|
Zhang J, Wang S, Jiang B, Huang L, Ji Z,
Li X, Zhou H, Han A, Chen A, Wu Y, et al: c-Src phosphorylation and
activation of hexokinase promotes tumorigenesis and metastasis. Nat
Commun. 8:137322017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Summy JM and Gallick GE: Src family
kinases in tumor progression and metastasis. Cancer Metastasis Rev.
22:337–358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Irby RB and Yeatman TJ: Role of Src
expression and activation in human cancer. Oncogene. 19:5636–5642.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mitra SK and Schlaepfer DD:
Integrin-regulated FAK-Src signaling in normal and cancer cells.
Curr Opin Cell Biol. 18:516–523. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Buczynski G and Potter RL: Nucleoside
diphosphate kinase from Xenopus oocytes; partial purification and
characterization. Biochim Biophys Acta. 1041:296–304. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Thomas SM and Brugge JS: Cellular
functions regulated by Src family kinases. Annu Rev Cell Dev Biol.
13:513–609. 1997. View Article : Google Scholar : PubMed/NCBI
|