Introduction
Esophageal carcinoma is the eighth most common and
most severe malignant tumor worldwide (1). Despite the different therapeutic
regimens employed with patients, including surgical resections,
radiotherapy and chemotherapy, the survival rate has continued to
be low for many decades. There are two strategies associated with
cancer immunotherapy: one strategy induces or enhances
immune-mediated tumor cell destruction, and the other counteracts
tumor cell evasion from the host's immune system or boosts
immune-mediated destruction (2). In
our previous study, dendritic cells were used as the basis of a
tumor vaccine for stimulating the immune system to target the
antigens associated with the tumor (3). It has been well established that
angiogenesis forms the basis of tumor development and metastasis;
thus, suppressing tumor-associated angiogenesis has become a
favorable strategy for the treatment of cancer (4). Active immunization with a whole
endothelial cell vaccine can control tumor size and metastasis by
restraining tumor angiogenesis in mice (5). Human umbilical vein endothelial cell
(HUVEC) vaccines have been effective in anti-angiogenesis immunity
against melanoma, glioblastoma, colorectal cancer and
hepatocellular carcinoma (5–8).
However, the anti-angiogenic effect of the HUVEC vaccine on ESCC is
still unknown. Xenograft mouse models have often been used to
evaluate the anticancer effect in ESCC. Transplantable tumors in
immunocompetent mice can be used to design therapeutic strategies
and analyze the underlying immunological mechanisms; however, the
value of these animal models has often been questioned when in
comparison with patients (9). As a
substitute, reconstituted mouse models, which consist of
immunodeficient mice that are repopulated with human immune cells,
are promising alternatives (10).
In the past few decades, modified transplantation models, which
support the transplantation and progression of a human
hematolymphoid system, have become a significant experimental
technique within this field of research (11). NOD/SCID mice are thought to be ideal
models for immunodeficient and immune reconstruction studies
(12). A previous study also
demonstrated that NOD/SCID mice transplanted with hominine
hematopoietic tissue or peripheral blood mononuclear cells (PBMC)
could reconstitute a functional immune system (13). Considering the role of the HUVEC
vaccine in antitumor and anti-angiogenic activities in many types
of cancers, the present study engrafted human PBMCs into NOD/SCID
mice to reconstruct a humanized immune system in order to evaluate
the antitumor and anti-angiogenic impact of the HUVEC vaccine on
ESCC, and further investigate the underlying mechanism.
Materials and methods
Mice and cells
A total of 30 female NOD/SCID mice, aged 4–5 weeks,
weighing 18–22 g, were acquired from Vital River Company (Beijing,
China). All mice used in the subsequent experiments were treated
and observed according to the guidelines of Zhengzhou University
Animal Ethics Committee (Henan, China). The animals were housed
under specific pathogen-free conditions (light on from 6:00 am to
6:00 pm, temperature 20–26°C, relative humidity: 40–60%) and
allowed access to a commercial mice chow and sterilized water ad
libitum. The mice were sacrificed by carbon dioxide euthanasia
at the 16th week after the initiation of the experiment, and the
tumor volume of the mice should not exceed 1, 000 mm3.
Total murine serum immunoglobulin (Ig) was detected by ELISA in all
mice and only non-leaky mice were used for analyses. The esophageal
tumor cell line EC9706 was cultured in Dulbecco's modified Eagle's
medium containing 10% fetal bovine serum (FBS) (both from
Biological Industries, Kibbutz Beit Haemek, Israel) with 5%
CO2 at 37°C. The primary HUVECs were separated from the
umbilical cord, and cultured with endothelial cell media, as
previously described (14).
Quantitation of IgG levels in
mice
The NOD/SCID mice blood samples with heparin were
centrifuged at 800 × g for 10 min and the isolated serum samples
were stored until subsequent use. The contents of mouse IgG was
detected using an IgG ELISA kit (Arigo, Taiwan, China). The
specimens were determined in triplicate by spectrophotometry at an
absorbance of 450 nm. According to the standards of Jackson
Laboratory (Bar Harbor, ME, USA), an IgG content <1 µg/ml
indicated no immune leakage; these mice were termed the non-leaky
mice.
Immune reconstruction of mice
The present study collected human PBMCs from healthy
volunteers as described in our previous study (15). Written informed consent was obtained
from healthy volunteers. The PBMCs were separated by density
gradient centrifugation at 1,500 × g for 20 min with lymphocyte
medium (TBD Sciences, Tianjin, China). Non-leaky NOD/SCID mice were
randomly divided into 2 groups: i) the vehicle group (n=8); and ii)
the PBMC group (n=16). The mice in the PBMC group were engrafted
with PBMCs (4×107 cells/mouse) intraperitoneally. After
4 weeks, the expression of cluster of differentiation
(CD)-45+ T-lymphocytes in the peripheral blood (obtained
from the retroorbital venous plexus) of the mice was detected via
flow cytometry. When the level of CD45+ T-lymphocytesin
the peripheral blood was >0.1%, the immune reconstruction of the
mice was considered successful (16).
Flow cytometry
The mouse blood samples were obtained from the
retroorbital venous plexus. Single-cell suspensions were generated
from engrafted mice spleens and filtered by 70-µm filter paper.
Blood (100 µl) or single-cell splenic suspension (1×106)
was treated with 10 µl anti-human fluorescein isothiocyanate
(FITC)-conjugated CD45 antibody (cat. no. ab134199) or
FITC-conjugated mouse IgG1 isotype control (cat. no. ab91356; both
from Abcam, Cambridge, UK) in the dark for 30 min at 4°C. Before
washing stained cells with PBS, all of the erythrocytes were
treated with FACS lysing solution (BD Biosciences, San Jose, CA,
USA) and then resuspended with 1 ml PBS containing 2% FBS. Finally,
T-lymphocytes were analyzed by flow cytometry.
Vaccination protocols in mice
Subconfluent HUVECs were trypsinized, centrifuged,
fixed with 0.025% glutaraldehyde and then suspended in PBS prior to
storage at −80°C. The successfully immune reconstituted mice
(humanized mice) were randomly allocated into 2 groups (n=8): i)
the PBMC group; and ii) the PBMC + HUVEC group. The mice in the
PBMC + HUVEC group were immunized 5 times with the HUVEC vaccine
(5×106 cells/mouse) in the axillary lymphonodus area,
once a week. The PBMC group was processed in the same manner using
PBS instead. Then the mice were subcutaneously injected with EC9706
cells (6×106/mouse) into their left flank. After the
tumor became palpable, the size was measured every other day using
vernier caliper. The long and short diameters were measured, and
the volume of the tumor (V) was calculated using the following
formula: V (mm3) = 0.5 × length × width2.
Immunohistochemistry
Tumor tissues and spleens resected from mice were
fixed in 4% paraformaldehyde, then embedded in paraffin for
subsequent sectioning. Each sample was cut into 4-µm sections and
incubated with the primary antibody anti-CD31 overnight at 4°C
(1:50 dilution; cat. no. ab28364; Abcam) in order to estimate the
microvascular content. The number of stained vessels were
quantified using microvessel density (MVD). Another section was
incubated with mouse anti-human CD45 antibody (1:100 dilution; cat.
no. ab187271; Abcam) overnight at 4°C in order to analyze the
infiltration of the spleen, which was then further incubated with
rabbit anti-mouse antibody at room temperature for 2 h (1:1,000
dilution; cat. no. sc-358914; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA). Following this, the sections were stained with
diaminobenzidine (ZSGB-BIO; OriGene Technologies, Inc., Beijing,
China) and hematoxylin. The sections were then observed under an
optical microscope (magnification, ×200).
Hemoglobin assay
A hemoglobin assay was conducted as previously
described (3). Briefly, the amount
of hemoglobin in tumors was verified using Drabkin's reagent
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) according to the
manufacturer's instructions. An equal quantity of tumor tissues was
homogenized in 1 ml oxidizing agent, and the samples were
centrifuged at 12,000 × g for 20 min. The samples were then
detected at an absorbance of 540 nm in a spectrophotometer (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) after incubation at 37°C
for 30 min. The relative contents of hemoglobin in tumor specimens
were then compared with the vehicle group.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed to determine the changes in
vascular endothelial growth factor receptor 2 (VEGFR2) and CD144
mRNA levels. Total RNA from tumor samples was extracted with TRIzol
(Invitrogen; Thermo Fisher Scientific Inc.), and cDNA was
synthesized with a RT-qPCR kit (Takara Bio, Inc., Otsu, Japan).
Target mRNA was quantified using the following primer pairs: GAPDH
forward, 5′-GAAGGCTGGGGCTCATTT-3′ and reverse,
5′-GAGGAGGCATTGCTGATGAT-3′; VEGFR2 forward,
5′-GAGTGAGGAAGGAGGACGAAGG-3′ and reverse,
5′-CCCGTAGGATGATGACAAGAAGTAGC-3′; CD144 forward,
5′-AAACACCTCACTTCCCCATC-3′ and reverse, 5′-ACCTTGCCCACATATTCTCC-3′.
The qPCR thermocycling conditions were as follows: Initial
denaturation at 95°C for 10 min, followed by 40 cycles of
denaturation at 95°C for 10 sec and annealing at 60°C for 30 sec.
Experiments were repeated 3 times for each sample. The 7500 Fast
Real-Time PCR System v2.0.5 (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was used to process the results.
Western blot analysis
The tumor tissues were homogenized with Tissue
Protein Extraction Reagent (Boster Biological Technology, Co.,
Ltd., Wuhan, China) to detect protein expression. The lysates were
processed and protein content was measured by bicinchoninic acid
assay (BCA; Beyotime Institute of Biotechnology, Shanghai, China).
Then, 50 µg protein of each group was isolated by 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes (PVDF; EMD
Millipore, Billerica, MA, USA) and subsequently incubated in
blocking buffer (5% skimmed milk, 1% Tween-20 in 20 mmol/l
Tris-buffered saline) at room temperature for 2 h. Then, the PVDF
membranes were incubated with anti-VEGFR2 (1:200 dilution; cat. no.
ab42228) or anti-CD144 (1:500 dilution; cat. no. ab166715) (both
from Abcam) primary antibodies overnight at 4°C, followed by
incubation with rabbit anti-mouse horseradish peroxidase
(HRP)-conjugated secondary antibody (1:1,000 dilution; cat. no.
sc-358914; Santa Cruz Biotechnology, Inc.) at room temperature for
2 h. In addition, the blots detected by enhanced chemiluminescence
kit (Invitrogen; Thermo Fisher Scientific Inc.) according to the
manufacturer's instructions. Equal loading of protein was confirmed
by probing with monoclonal mouse anti-human β-actin antibody
(1:1,000 dilution; cat. no. sc-130065; Santa Cruz Biotechnology
Inc.).
HUVEC membrane protein was prepared using the
Membrane Protein Extraction Kit (Beyotime Institute of
Biotechnology, Shanghai, China) to detect antibody levels. An equal
quantity of proteins was separated and then transferred to the
membranes. The membranes were incubated overnight with mouse serum
obtained from the mice, and then incubated with a rabbit anti-human
secondary antibody labeled with horseradish peroxidase (cat. no.
ab6759; Abcam). The results were finally quantified with ImageJ
software (National Institutes of Health, Bethesda, MD, USA).
Detection of the anti-HUVEC
antibody
ELISAs were performed in order to determine the
levels of anti-HUVEC antibody in the serum of mice, as described in
our previous study (3). Briefly,
HUVEC membrane proteins were loaded onto ELISA plates (JetBioFil,
Guangzhou, China). Then the plates were incubated with 100 µl serum
(1:50), which was obtained from mice the following day. The ELISA
was conducted with 3,3′,5,5′-tetramethyl benzidine (Beijing
Solarbio Science & Technology Co., Ltd., Beijing, China) and
the reaction was terminated using H2SO4 after
incubation with rabbit anti-mouse HRP-conjugated IgG (1:1,000
dilution; cat. no. sc-358914; Santa Cruz Biotechnology, Inc.) at
37°C for 2 h, and the optical density at an absorbance of 450 nm
was measured. Each sample was evaluated in 3 separate
experiments.
Statistical analysis
Quantitative data was expressed as the mean ±
standard deviation. Student's t-test (two-sided) was conducted for
analysis between two subgroups. Statistical significance between
>2 subgroups were evaluated by one-way analysis of variance
followed with Bonferroni and Tamhane post hoc tests. GraphPad Prism
(GraphPad Software, Inc., La Jolla, CA, USA) was used for all
statistical tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
Establishment of the humanized mouse
model
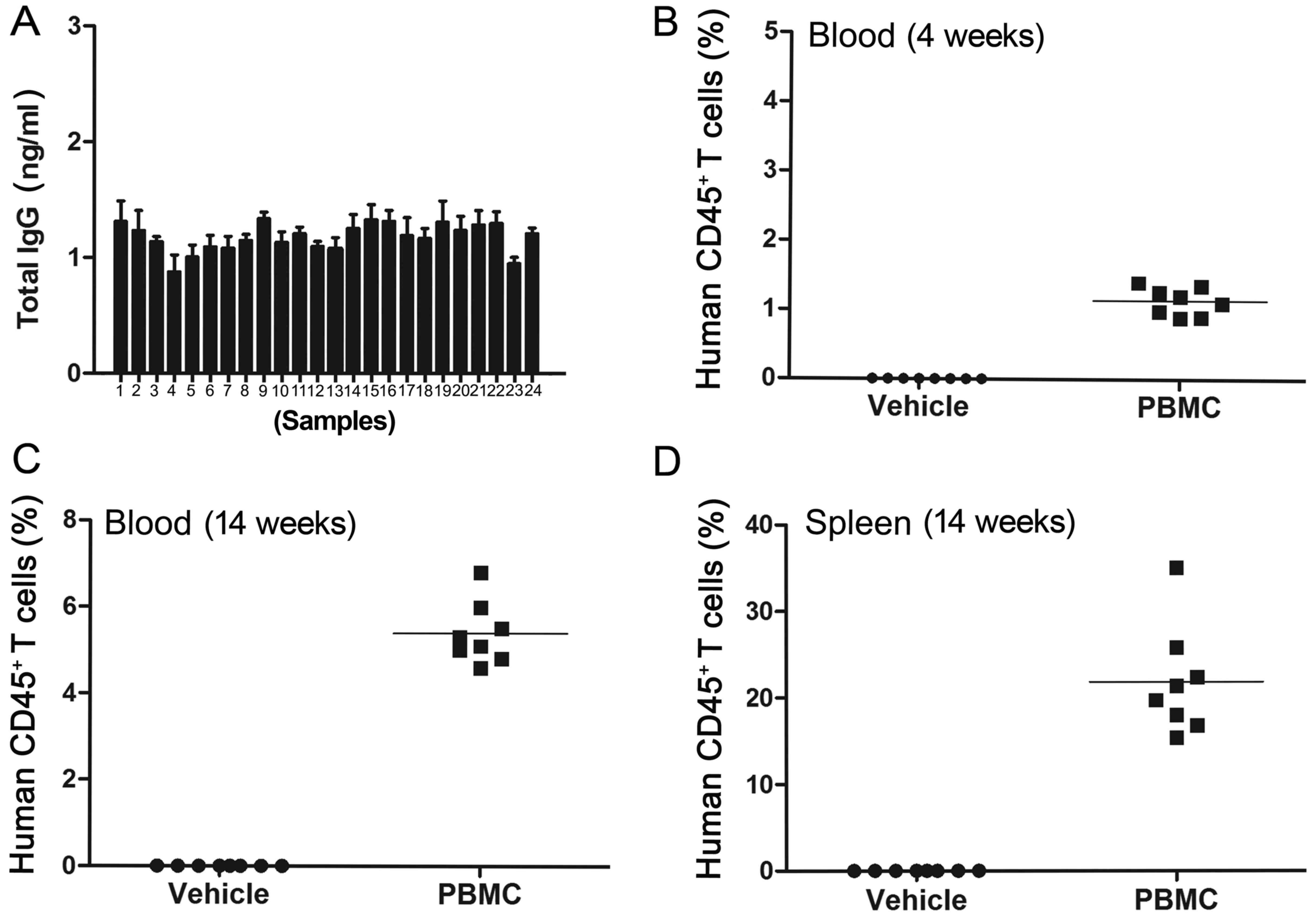
Total murine serum IgG was determined by ELISA. The
results indicated that the total murine serum IgG was <1 µg/ml
in all 24 tested mice, which indicated that all of the mice had no
immune leakage (Fig. 1A). The
non-leaky mice were then used to establish the humanized mouse
model.
The levels of human CD45+ T-lymphocytes
in mouse peripheral blood and spleens were assessed by flow
cytometry at different time-points after PBMC administration. The
peripheral blood levels of human CD45+ T-lymphocytes in
the PBMC group was >0.1% at 4 weeks after PBMC administration
(P<0.01; Fig. 1B), which further
increased to >5% after 14 weeks post-PBMC application
(P<0.001; Fig. 1C). Furthermore,
the level of human CD45+ T-lymphocytes in the mouse
spleens of the PBMC group was significantly higher when compared
with the vehicle group at 14 weeks after the PBMC injections
(P<0.01; Fig. 1D).
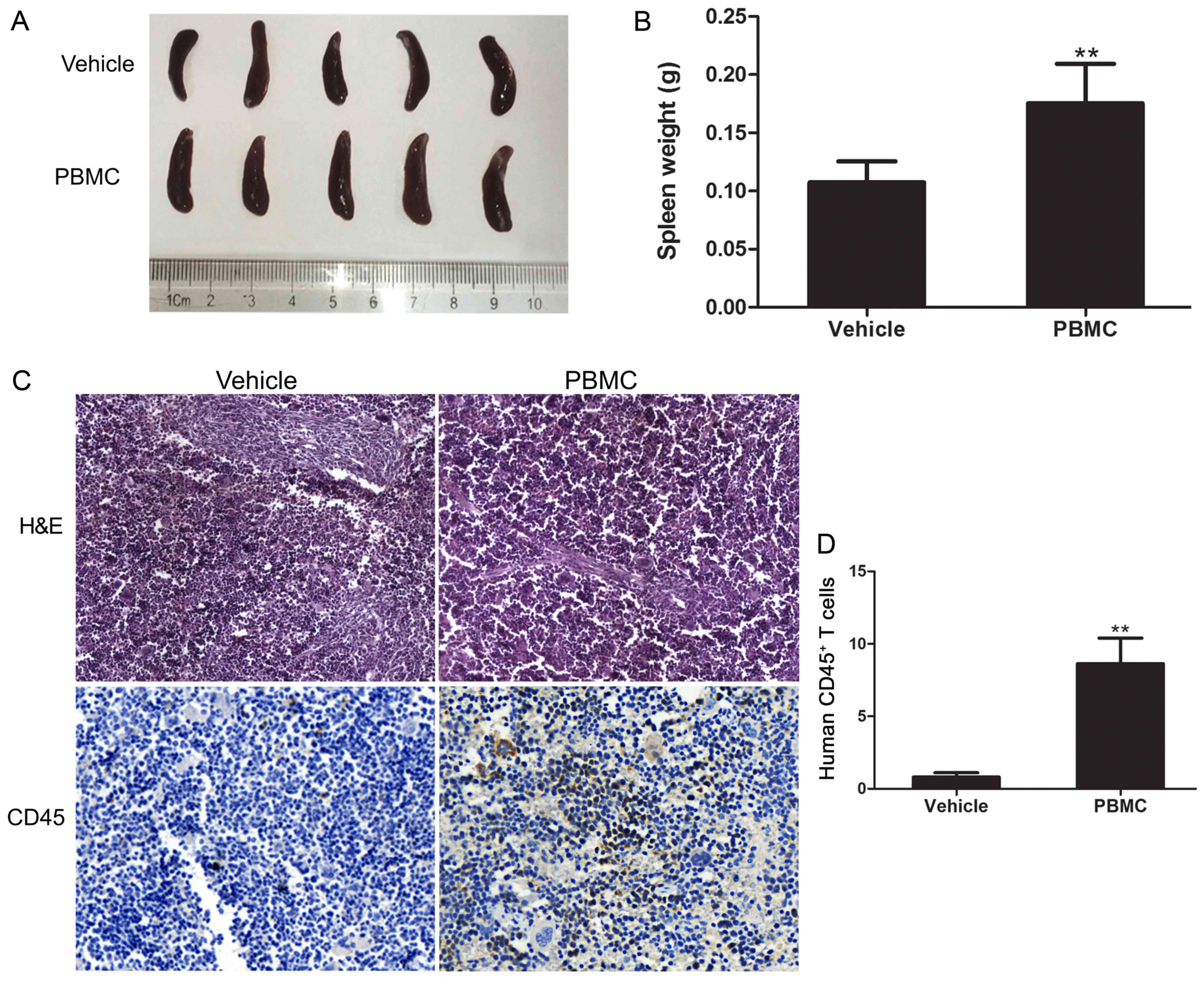
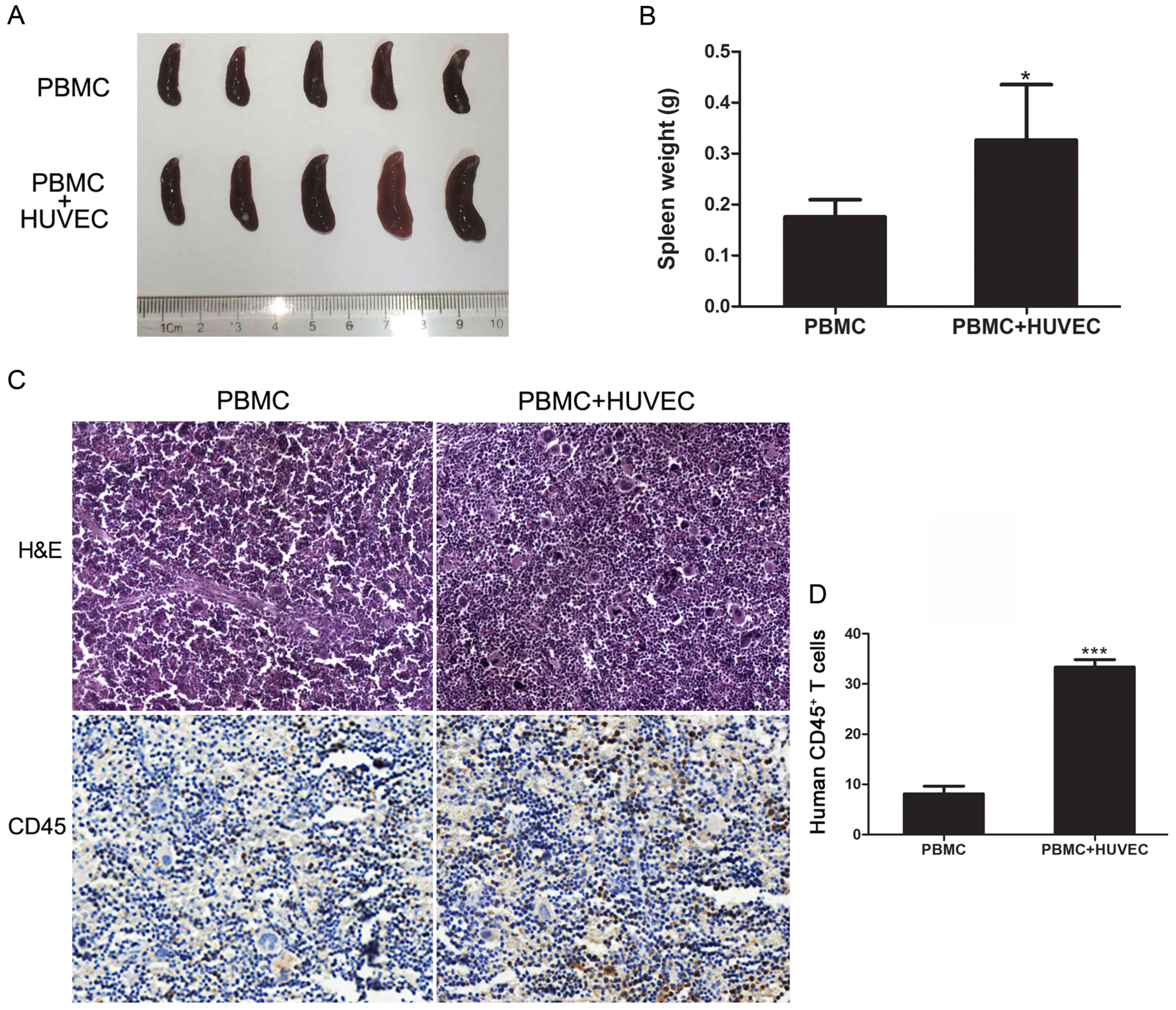
To further assess the establishment of the humanized
engraftment mouse model, the present study examined the recruitment
of T-lymphocytes in the spleen 14 weeks after the PBMC injections.
The mouse spleens were excised and then weighed; the results
revealed that the size and weight of the spleens in the PBMC group
were significantly greater when compared with the control group
(P<0.01; Fig. 2A and B). Then,
the expression of the human CD45 leukocyte common antigen in the
spleens was analyzed by immunohistochemical staining. The results
demonstrated that the spleens of the PBMC group of mice exhibited
extensive human CD45+ T-lymphocyte levels, while the
vehicle group did not present CD45 positive signals (P<0.01;
Fig. 2C and D). In conclusion, the
results indicated that the humanized mouse model was established
successfully.
HUVEC vaccine inhibits the
angiogenesis of ESCCs in the humanized mouse model
A previous study demonstrated that immunization with
the HUVEC vaccine prevented tumor development and metastasis by
utilizing the immune system to attack tumor vasculature in myeloma
tumor models and Lewis lung carcinoma (17). However, there are increasing
concerns regarding the value of this mouse model when translating
the research to clinical application (18). Thus, an increasing number of studies
have employed humanized mice to study human malignancies and
biological processes (19–21). In the present study, the humanized
mice were vaccinated weekly with either PBS or the HUVEC vaccine
for 5 weeks. Then, esophageal carcinoma EC9706 cells were
implanted, measured over time and then the resulting tumors were
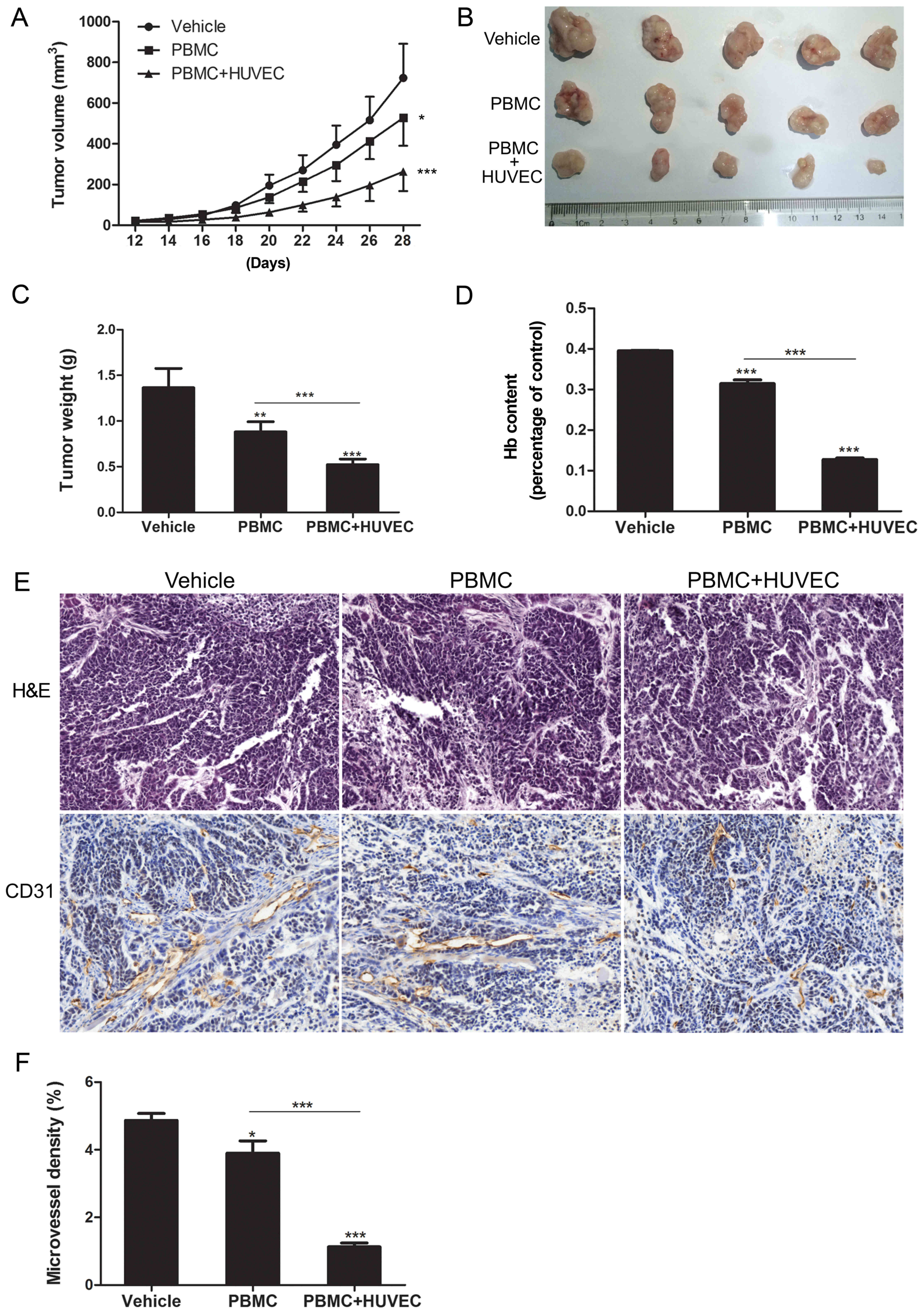
removed for analyses. When compared with the vehicle group, tumor
size was reduced in the PBMC group (P<0.05) and the PBMC + HUVEC
group (P<0.001; Fig. 3A).
Notably, when compared with the PBMC group, tumor size was
significantly suppressed in the PBMC + HUVEC group (P<0.001;
Fig. 3B and C).
The present study then assessed the response of the
HUVEC vaccine by analyzing angiogenesis in the tumor specimens. The
hemoglobin assay revealed that the hemoglobin content in the PBMC +
HUVEC group was significantly less than in the two other groups
(P<0.001; Fig. 3D). In addition,
immunohistochemistry was conducted to investigate CD31 staining (an
angiogenesis marker), and to analyze the MVD. The MVD in the PBMC +
HUVEC group was significantly less than that observed in the
vehicle and PBMC groups (P<0.001; Fig. 3E and F). These results indicated
that mice immunized with HUVECs may have inhibited ESCC growth via
the suppression of tumor angiogenesis in the humanized mouse
model.
Detection of antigens and antibodies
in solid tumors and serum
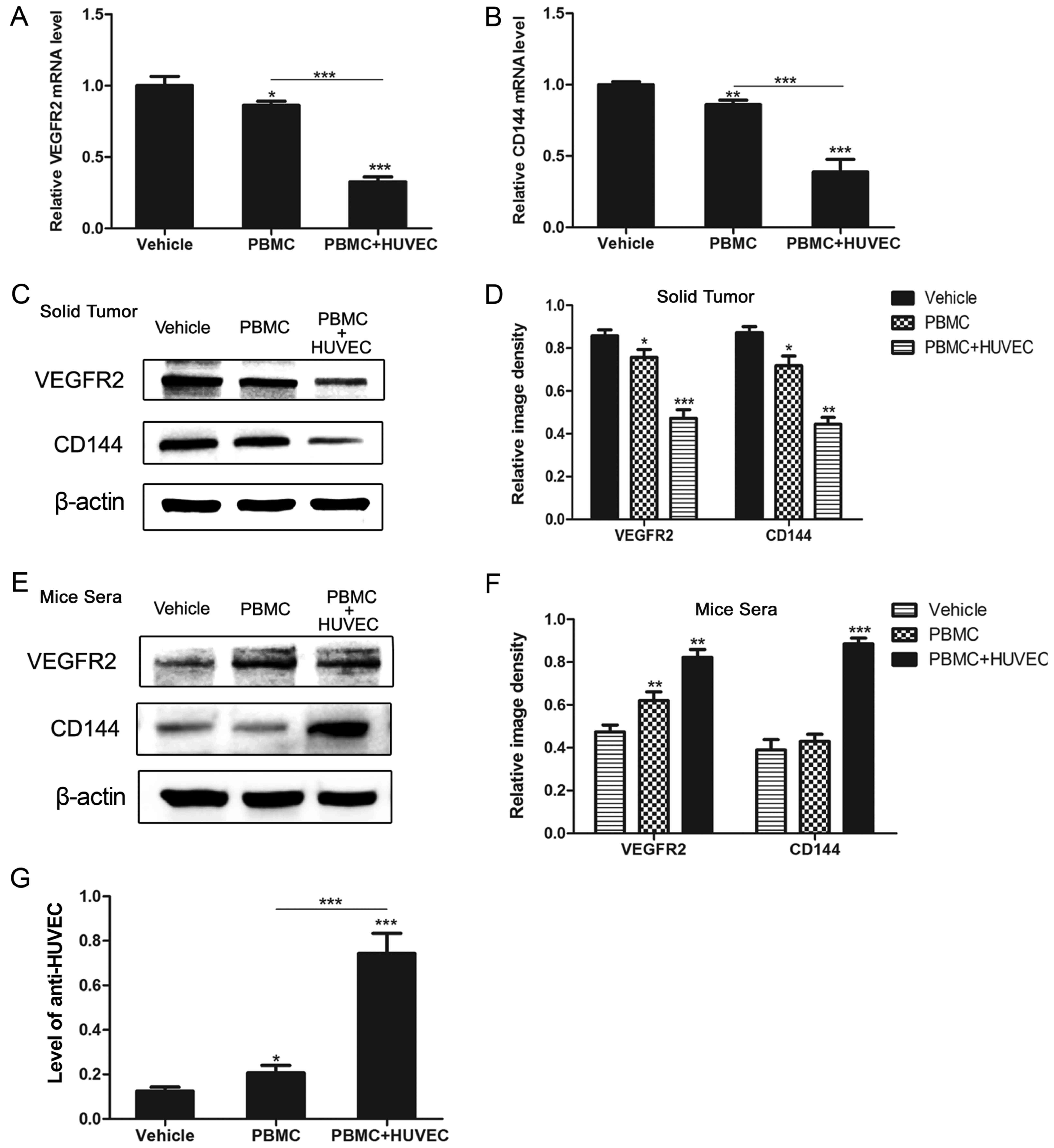
To evaluate angiogenesis-associated antigens and
antibodies in mice, the expression of VEGFR2 and CD144 in solid
tumors and serum was detected. After 4 weeks of tumor development,
all of the mice were sacrificed, and the tumors and serum samples
obtained were analyzed for mRNA and protein expression. The results
of RT-qPCR indicated that the mRNA levels of VEGFR2 and CD144 in
the solid tumors of the PBMC + HUVEC group were significantly
reduced when compared with the two other groups (P<0.01;
Fig. 4A and B). The levels of
VEGFR2 and CD144 proteins in the solid tumors of the PBMC + HUVEC
group were also significantly reduced (P<0.01; Fig. 4C and D). Mouse sera was also
investigated using western blotting; and the results revealed that
the serum antibody levels were significantly increased in the PBMC
+ HUVEC group (P<0.01; Fig. 4E and
F). In order to investigate the levels of the anti-HUVEC
antibody in the serum of mice, ELISA analysis was performed, and
the levels in mice immunized with the HUVEC vaccine were more
marked (P<0.001; Fig. 4G). The
results demonstrated that the HUVEC vaccine increased the levels of
angiogenesis-associated antigens in humanized mice.
Immunization with the HUVEC vaccine
increases T-lymphocyte recruitment in the spleen
In order to evaluate the immune responses elicited
by the HUVEC vaccine, the present study detected T-lymphocyte
recruitment in mice after the last immunization. The weight and
volume of the spleen were increased in the PBMC + HUVEC group
(P<0.05; Fig. 5A and B). The
results of splenic immunohistochemical staining demonstrated that
the levels of human CD45+ T-lymphocytes in the PBMC +
HUVEC group were significantly higher than in the PBMC group
(P<0.001; Fig. 5C and D).
Overall, the results indicated that mice immunized with the HUVEC
vaccine had increased T-lymphocyte recruitment in the spleens of
humanized mice.
Discussion
A number of animal models have been developed and
used in basic tumor research. Due to the technical and ethical
considerations, investigations into the immunobiology of
allotransplantation can only be performed using animals, which are
limited when translating the results to humans (22). Progress has been made with the
development of different chimeric animal models, which can serve
important roles in studies associated with cancer, transplantation
biology, infectious diseases, and autoimmunity (23). In the present study, human PBMC were
engrafted into immunodeficient hosts, established mice with human
lymphocytes, to characterize the effects of the HUVEC vaccine on
esophageal carcinoma (16). Based
on previous studies, mice were transplanted with PBMCs at a dose of
4×107/mouse, which has been shown to have an increased
success rate of immune reconstruction and a reduced risk of graft
vs. host disease (16,20,24).
Among the various transplantation methods, the present study
engrafted human PBMCs into NOD/SCID mice via an intraperitoneal
injection because the method is more readily manipulated (22). After 4 weeks of immunization with
human PBMCs, human CD45+ T-lymphocytes were detected in
mice. In addition, the weight of the spleens in the PBMC group was
greater than in the vehicle group. The present study demonstrated
the existence of T-lymphocytes to further assess the engraftment
model. As a result, a large number of lymphocytes were identified
in the spleen.
Angiogenesisis a vital part of sustained tumor
development and progression, and involves comprehensive crosstalk
among stromal cells, pericytes, tumor cells and vascular
endothelial cells (4,25). Therefore, inhibition of angiogenesis
is currently being studied as a new anticancer therapy. Since the
first vaccine targeting tumor angiogenesis was reported,
proliferative HUVECs as a vaccine to induce antitumor immunity has
been studied in several in vivo tumor models (5–8,17,26).
However, further investigation is required before the HUVEC vaccine
can be used clinically, and the antitumor effect has not been
studied previously in esophageal carcinoma. In the present study,
the humanized mouse model immunized with the HUVEC vaccine
significantly suppressed the size of the tumor. To further explore
the underlying mechanism, the hemoglobin content and MVD of tumor
specimens were evaluated. The results revealed that mice immunized
with the HUVEC vaccine had reduced ESCC development due to
suppression of tumor angiogenesis.
VE-Cadherin (CD144), an endothelial-specific
adhesion protein that serves as an endothelial marker, mediates
vascular stability and angiogenesis (27–29).
VEGFR2, an overexpressed angiogenesis-associated antigen is
appropriate for targeting the tumor vasculature via anti-angiogenic
therapy, and is perceived to be a specific endothelial molecule
(30). The present study detected
the mRNA and protein levels of angiogenesis-associated antigens in
the tumor specimen. It was demonstrated that the HUVEC vaccine
decreased the levels of angiogenesis-associated antigens. The
expression of antibodies and the ability of anti-HUVEC cells to
proliferate were then evaluated in vitro in the serum from
immunized mice. The results revealed that there were higher levels
of antibodies in the sera of mice immunized with the HUVEC
vaccine.
A critical role of the immune system is to suppress
the progression of tumors and inhibit tumor growth. To further
demonstrate that the HUVEC vaccine can induce these specific
cellular immune responses, the present study monitored the levels
of T-lymphocyte infiltration in the spleens. The weight of the
spleens in the PBMC + HUVEC group was significantly higher than
that observed in the PBMC group. Moreover, immunohistochemical
staining indicated that a high level of human T-lymphocytes
infiltrated the spleens of the PBMC + HUVEC group, which provided
evidence that the HUVEC vaccine successfully induced immune
responses in the humanized mouse model.
In conclusion, the present study revealed that the
HUVEC vaccine inhibited ESCC growth by reducing tumor angiogenesis
in a humanized mouse model. Thus, the results of the present study
provide a potential model for evaluating immunotherapeutic
approaches and highlights the potential use of HUVECs.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81572972), the
Science and Technology Innovation Talents Support Plan of
University in Henan Province (grant no. 15HASTIT038) and the
Colleges and Universities in Henan Province Key Scientific Research
Plan (grant no. 15A310030).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Author's contributions
JL and HL conceived and designed the study and the
experiments. HL, JZ, YY, GJ, XZ, DW, CX, KL, XC, XL and FT
performed the experiments and analyzed the data. HL, YY and JL
drafted, wrote or revised the manuscript. All authors read and
approved the manuscript and agreed to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
All mice used in the study were treated and observed
according to the guidelines of the Zhengzhou University Animal
Ethics Committee (Henan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang Y: Epidemiology of esophageal
cancer. World J Gastroenterol. 19:5598–5606. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pujol JL, De Pas T, Rittmeyer A, Vallieres
E, Kubisa B, Levchenko E, Wiesemann S, Masters GA, Shen R,
Tjulandin SA, et al: Safety and immunogenicity of the PRAME cancer
immunotherapeutic in patients with resected non-small cell lung
cancer: A phase I dose escalation study. J Thorac Oncol.
11:2208–2217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang Y, Lu J, Liu H, Jin G, Bai R, Li X,
Wang D, Zhao J, Huang Y, Liu K, et al: Dendritic cells loading
autologous tumor lysate promote tumor angiogenesis. Tumour Biol.
37:15687–15695. 2016. View Article : Google Scholar
|
|
4
|
Weis SM and Cheresh DA: Tumor
angiogenesis: Molecular pathways and therapeutic targets. Nat Med.
17:1359–1370. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xu M, Xing Y, Zhou L, Yang X, Yao W, Xiao
W, Ge C, Ma Y, Yang J, Wu J, et al: Improved efficacy of
therapeutic vaccination with viable human umbilical vein
endothelial cells against murine melanoma by introduction of OK432
as adjuvant. Tumour Biol. 34:1399–1408. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tanaka M, Tsuno NH, Fujii T, Todo T, Saito
N and Takahashi K: Human umbilical vein endothelial cell vaccine
therapy in patients with recurrent glioblastoma. Cancer Sci.
104:200–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mu X, Sang Y, Fang C, Shao B, Yang L, Yao
K, Zhao X, Gou J, Wei Y, Yi T, et al: Immunotherapy of tumors with
human telomerase reverse transcriptase immortalized human umbilical
vein endothelial cells. Int J Oncol. 47:1901–1911. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu M, Zhou L, Zhang P, Lu Y, Ge C, Yao W,
Xing Y, Xiao W, Dong Y, Wu J, et al: Enhanced antitumor efficacy by
combination treatment with a human umbilical vein endothelial cell
vaccine and a tumor cell lysate-based vaccine. Tumour Biol.
34:3173–3182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ellis LM and Fidler IJ: Finding the tumor
copycat. Therapy fails, patients don't. Nat Med. 16:974–975. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sanmamed MF, Rodriguez I, Schalper KA,
Onate C, Azpilikueta A, Rodriguez-Ruiz ME, Morales-Kastresana A,
Labiano S, Perez-Gracia JL, Martín-Algarra S, et al: Nivolumab and
urelumab enhance antitumor activity of human T lymphocytes
engrafted in Rag2−/−IL2Rγnull immunodeficient
mice. Cancer Res. 75:3466–3478. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Theocharides AP, Rongvaux A, Fritsch K,
Flavell RA and Manz MG: Humanized hemato-lymphoid system mice.
Haematologica. 101:5–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Li H, Liu J, Guan Y, Huang L, Tang
H and He J: Immune reconstitution from peripheral blood mononuclear
cells inhibits lung carcinoma growth in NOD/SCID mice. Oncol Lett.
8:1638–1644. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Banuelos SJ, Shultz LD, Greiner DL,
Burzenski LM, Gott B, Lyons BL, Rossini AA and Appel MC: Rejection
of human islets and human HLA-A2.1 transgenic mouse islets by
alloreactive human lymphocytes in immunodeficient NOD-scid
and NOD-Rag1nullPrf1null mice. Clin
Immunol. 112:273–283. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu J, Zhao J, Liu K, Zhao J, Yang H, Huang
Y, Qin Z, Bai R, Li P, Ma J, et al: MAPK/ERK1/2 signaling mediates
endothelial-like differentiation of immature DCs in the
microenvironment of esophageal squamous cell carcinoma. Cell Mol
Life Sci. 67:2091–2106. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jin G, Zhao J, Yang YI, Liu K, Jiang Y,
Zhang X, Zhang Y, Huang Y, Lu J and Dong Z: JAK/STAT3 signaling
pathway mediates endothelial-like differentiation of immature
dendritic cells. Oncol Lett. 10:3471–3477. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
King M, Pearson T, Shultz LD, Leif J,
Bottino R, Trucco M, Atkinson MA, Wasserfall C, Herold KC, Woodland
RT, et al: A new Hu-PBL model for the study of human islet
alloreactivity based on NOD-scid mice bearing a targeted mutation
in the IL-2 receptor gamma chain gene. Clin Immunol. 126:303–314.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen XY, Zhang W, Zhang W, Wu S, Bi F, Su
YJ, Tan XY, Liu JN and Zhang J: Vaccination with viable human
umbilical vein endothelial cells prevents metastatic tumors by
attack on tumor vasculature with both cellular and humoral
immunity. Clin Cancer Res. 12:5834–5840. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Holzapfel BM, Wagner F, Thibaudeau L,
Levesque JP and Hutmacher DW: Concise review: Humanized models of
tumor immunology in the 21st century: Convergence of cancer
research and tissue engineering. Stem Cells. 33:1696–1704. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Brehm MA, Shultz LD and Greiner DL:
Humanized mouse models to study human diseases. Curr Opin
Endocrinol Diabetes Obes. 17:120–125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shultz LD, Ishikawa F and Greiner DL:
Humanized mice in translational biomedical research. Nat Rev
Immunol. 7:118–130. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shultz LD, Brehm MA, Garcia-Martinez JV
and Greiner DL: Humanized mice for immune system investigation:
Progress, promise and challenges. Nat Rev Immunol. 12:786–798.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gong Z, Xu H, Su Y, Wu W, Hao L and Han C:
Establishment of a novel bladder cancer xenograft model in
humanized immunodeficient mice. Cell Physiol Biochem. 37:1355–1368.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yacoub-Youssef H, Marcheix B, Calise D,
Thiers JC, Therville N, Benoist H, Blaes N, Segui B, Dambrin C and
Thomsen M: Engraftment of human T, B and NK cells in CB.17
SCID/beige mice by transfer of human spleen cells. Transpl Immunol.
15:157–164. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ji M, Jin X, Phillips P and Yi S: A
humanized mouse model to study human immune response in
xenotransplantation. Hepatobiliary Pancreat Dis Int. 11:494–498.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Folkman J: Angiogenesis: An organizing
principle for drug discovery? Nat Rev Drug Discov. 6:273–286. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei YQ, Wang QR, Zhao X, Yang L, Tian L,
Lu Y, Kang B, Lu CJ, Huang MJ, Lou YY, et al: Immunotherapy of
tumors with xenogeneic endothelial cells as a vaccine. Nat Med.
6:1160–1166. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Peiffer DS, Zimmerman NP, Wang LS, Ransom
BW, Carmella SG, Kuo CT, Siddiqui J, Chen JH, Oshima K, Huang YW,
et al: Chemoprevention of esophageal cancer with black raspberries,
their component anthocyanins, and a major anthocyanin metabolite,
protocatechuic acid. Cancer Prev Res. 7:574–584. 2014. View Article : Google Scholar
|
|
28
|
Dejana E and Giampietro C: Vascular
endothelial-cadherin and vascular stability. Curr Opin Hematol.
19:218–223. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vestweber D: VE-cadherin: The major
endothelial adhesion molecule controlling cellular junctions and
blood vessel formation. Arterioscler Thromb Vasc Biol. 28:223–232.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Okaji Y, Tsuno NH, Saito S, Yoneyama S,
Tanaka M, Nagawa H and Takahashi K: Vaccines targeting tumour
angiogenesis-a novel strategy for cancer immunotherapy. Eur J Surg
Oncol. 32:363–370. 2006. View Article : Google Scholar : PubMed/NCBI
|