Introduction
Sex-determining region Y (SRY)-box 9 (SOX9) is a
member of a transcription factor family characterized by
similarities in the high-mobility group DNA-binding domain of SRY
(1). It plays crucial roles in the
development and differentiation of diverse tissues, such as
cartilage (1,2) and testes (3). During chondrogenesis, SOX9
transactivates the genes encoding extracellular matrix (ECM)
proteins, including type II collagen and aggrecan, which function
as a foothold for chondrocytes (4).
During male gonadogenesis, SOX9 specifies the Sertoli cell lineage
and upregulates the transcription of a gene encoding anti-Müllerian
hormone, which inhibits the development of the female gonads
(3). SOX9 is also crucial for the
development and maintenance of ectodermal tissues. In adult skin,
SOX9 is expressed in the sebaceous glands, sweat glands, outer root
sheath of the hair follicles (5),
and basal layer of the human epidermis, where it acts as an
important regulator of epidermis and cutaneous appendages through
pro-proliferative and anti-apoptotic activities (6).
Recently, SOX9 was shown to be involved in the
initiation and/or progression of various types of cancers,
including prostate (7), breast
(8) and colorectal (9) cancers, basal cell carcinoma of the
skin (10) and esophageal squamous
cell carcinoma (SCC) (11). These
studies have described higher SOX9 expression levels in cancer
cells than in their normotypic counterparts (7–11). In
addition, three of the studies have reported that high SOX9
expression levels in cancer cells are positively correlated with
poor clinical outcomes (8,9,11).
However, in oral squamous cell carcinoma (OSCC), both the
expression profile of SOX9 and its possible clinicopathological
correlates remain unknown.
We previously described the importance of ECM
proteins in the pathogenesis of OSCC (12,13)
and salivary gland tumors (14,15).
We demonstrated that the keratinocyte-derived, basement
membrane-type heparan sulfate proteoglycan perlecan is deposited in
the intraepithelial spaces during oral epithelial carcinogenesis
(12). Indeed, we revealed that
perlecan was abundantly expressed around the invasive nests of OSCC
and that this increased expression in the stromal space was a
useful indicator of stromal invasion (13). Furthermore, perlecan acts as a
reservoir for growth factors, such as keratinocyte growth factor
(14) and vascular endothelial
growth factor (16). Perlecan
expression is regulated by the NF-κB pathway, which regulates the
transcription of the perlecan-encoding HSPG2 gene in the
microenvironment of prostate cancer (17). Transcriptional profiling of
oncogene-expressing cells with a SOX9 deletion has
demonstrated that SOX9 is a potent positive regulator of ECM
(including perlecan) gene transcription (18).
Given the above reports, we aimed to characterize
the immunohistochemical expression profiles of SOX9 in surgical
samples of oral mucosa that contained foci of epithelial dysplasia,
carcinoma in situ (CIS), and SCC as well as in cultured
cells of OSCC origin. In addition, we investigated the use of SOX9
expression as a novel prognostic marker for OSCC.
Materials and methods
Tissue samples
Forty-nine consecutive patients (27 males and 22
females) who underwent primary surgical treatment for OSCC from
January 2008 to December 2011 were included in the present study.
Patients with recurrent tumors were excluded. The follow-up
clinical information was obtained from the patient medical records.
The follow-up period was measured from the date of surgery until
death or until the final follow-up record. Clinical stage was
determined according to the Union for International Cancer Control
TNM classification system, 7th edition (19). The median age of the patients at
surgery was 69 years (range, 22–88 years).
Surgical specimens were fixed in 10% formalin and
embedded in paraffin. After examining hematoxylin and eosin
(H&E)-stained sections from each specimen, the paraffin block
from each patient that simultaneously contained (maximum possible)
the SCC area, including the invasive front, and areas of adjacent
epithelia appearing normal, dysplastic, or as CIS was selected.
Serial 2.5-µm-thick sections were prepared for H&E staining and
immunohistochemistry. In addition, samples of metastatic lymph
nodes were collected from 17 of the 19 patients with late
metastasis in the cervical lymph nodes. The Ethics Board of the
Niigata University Graduate School of Medicine and Dental Sciences
(Oral Life Science) reviewed and approved the experimental protocol
for analyzing the surgical materials (approval no. 12-10-13) and
written informed consent was obtained from the patients.
Antibodies
The following antibodies were used for
immunostaining: Anti-SOX9 (1:1,500; rabbit polyclonal; cat. no.
HPA001758; Atlas Antibodies, Stockholm, Sweden); anti-perlecan
(1:6,000; rabbit polyclonal) was established by Saku and Furthmayr
as previously described (20);
anti-keratin 17 (K17) (1:100; mouse monoclonal; cat. no. M7046;
clone E3; Dako, Glostrup, Denmark); and anti-Ki-67 (1:100; mouse
monoclonal; cat. no. M7240; clone MIB-1; Dako). For western
blotting, a different anti-SOX9 antibody (1:1,000; rabbit
monoclonal; cat. no. 82630; clone D8G8H; Cell Signaling Technology,
Inc., Danvers, MA, USA) was used.
Immunohistochemistry
Immunohistochemical analysis of paraffin sections
was performed using the ChemMate EnVision system (Dako) according
to the manufacturer's instructions. Deparaffinized and rehydrated
sections were immersed in 0.3% hydrogen peroxide in methanol for 30
min at room temperature to block endogenous peroxidase activities.
For SOX9, K17 and Ki-67 immunostaining, antigen retrieval was
performed by first autoclaving sections in citric acid buffer (pH
6.0) at 121°C for 10 min. To analyze the stromal expression of
perlecan, the sections were incubated with hyaluronidase (Type IS,
330 UI/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 30
min prior to immunostaining. After antigen retrieval, the sections
were incubated with 5% milk protein in phosphate-buffered saline
(PBS) for 30 min and then with the primary antibodies overnight at
4°C. After washing with PBS, the sections were reacted with
EnVision (Dako) for 60 min at room temperature. Peroxidase reaction
products were developed with 3,3′-diaminobenzidine, and the
sections were counterstained with hematoxylin. In the control
experiments, the primary antibodies were replaced with the
appropriate negative control immunoglobulin (cat. no. X0931 for
mouse IgG1 and cat. no. X0903 for rabbit Ig fraction; Dako).
SOX9 expression
To investigate the localization of SOX9 in squamous
cell epithelial lesions, the combined H&E and
immunohistochemical findings were used to first define the foci on
a given section on a case-by-case basis, which demonstrated normal
epithelia, epithelial dysplasia, and CIS, if present. Nuclear and
cytoplasmic positivity was evaluated individually for foci of each
type of epithelia and for SCC; foci were positive if they occupied
≥10% of the area of stained cells. For the SCC components, the
proportion of the entire area of the SCC demonstrating
SOX9-positive cancer cells was manually semi-quantified on the
representative sections of each specimen.
Cell lines
Human cell lines derived from OSCC HSC-3 and HSC-4
cells were obtained from Riken BRC Cell Bank (Tsukuba, Japan). To
analyze the association between SOX9 expression and metastasis, two
cell lines, namely HSC-3, a metastatic cell line that was
established from the metastatic lymph node of a 63-year-old man
with poorly differentiated SCC and HSC-4, a non-metastatic cell
line that was established from a metastatic lymph node in a
63-year-old man with well-differentiated SCC, were selected
(21). The cells were maintained in
Minimum Essential Medium (Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal calf serum (FCS) under a
humidified atmosphere with 5% CO2 and 95% air at
37°C.
Immunofluorescence
The HSC-3 and HSC-4 cells were seeded on a coverslip
in 35-mm dishes at a cell concentration of 0.25×105
cells/dish. After 3 days, the cells were washed with PBS and fixed
with 4% paraformaldehyde for 30 min at room temperature. Then, the
cells were permeabilized with 0.2% Triton X-100 in PBS, and then
incubated with 5% milk protein in PBS at 4°C overnight to block
non-specific binding. The cells were incubated with the anti-SOX9
primary antibody for 60 min. After washing with PBS, the cells were
reacted with Alexa Fluor™ 568 goat anti-rabbit IgG secondary
antibody (1:200; cat. no. A11011; Thermo Fisher Scientific, Inc.).
The coverslips were mounted with ImmunoSelect Antifading Mounting
Medium DAPI (Dianova, Hamburg, Germany), and the cells were
observed using a Nikon Eclipse E600W immunofluorescence microscope
(Nikon Corp., Tokyo, Japan). The fluorescence intensities for SOX9
and DAPI staining were measured using the NIS-Elements AR software
(Nikon Corp.).
Scratch wound-healing assay
Artificial wounds were inflicted on the confluent
monolayers of HSC-3 and HSC-4 cells in 60-mm culture dishes using a
200-µl pipette tip. Every 6 h for 36 h, the process of migration
into the wound areas was monitored under a phase-contrast
microscope equipped with a digital camera. Wound surface area was
measured on the captured images of four fields using ImageJ
(version 1.50i; National Institutes of Health, Bethesda, MD, USA).
Percent (%) closure of the wound surface area was calculated as
open surface area/total surface area (surface area at 0 h) ×
100.
Growth curve analysis
The cells were seeded at a concentration of
0.25×105 cells in 60-mm2 dishes and incubated
at 37°C for 6 days. Thereafter, the cells were counted on days 2, 4
and 6. The average number of proliferating cells was calculated
from triplicate experiments.
Real-time reverse
transcription-polymerase chain reaction (RT-PCR)
HSC-3 and HSC-4 cells were lysed with ISOGEN (Nippon
Gene Co., Ltd., Tokyo, Japan) for the extraction of total RNA. cDNA
was synthesized from each 5-µg RNA sample using the SuperScript III
First-Strand Synthesis system (Thermo Fisher Scientific, Inc.),
followed by PCR. RT-PCR was performed with specific primers for
SOX9 using a MiniOpticon Real-Time PCR Detection system
CFB-3120 (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Gene
expression levels were quantified by the ΔΔCq method (22) using CFX Manager Software version 2.1
(Bio-Rad Laboratories). The expression of the peptidylprolyl
isomerase A (PPIA) gene was used to normalize for variance,
and the expression levels of specific genes were represented as
ratios relative to the expression level of PPIA from the
same master reaction. PCR primer pairs used for each gene were:
5′-AGCTCTGGAGACTTCTGAACGAGAG-3′ and 5′-CGTTCTTCACCGACTTCCTCC-3′ for
SOX9; and 5′-GCAGTAATGGGTTACTTCTGAAAC-3′ and
5′-TGCCTCAGGTAATACATTACAGAC-3′ for PPIA.
Western blotting
For the extraction of total protein, the HSC-3 and
HSC-4 cells were lysed at day 5 after seeding with M-PER™ Mammalian
Protein Extraction reagent (Thermo Fisher Scientific, Inc.).
Thereafter, the cell lysates were used for western blotting as
described elsewhere (23). Primary
antibody for SOX9 was used at a dilution of 1:1,000. Labeled
protein bands were visualized using the ECL Prime Western Blotting
Detection system (GE Healthcare UK, Ltd., Little Chalfont, UK).
Statistical analysis
The association between SOX9 expression profiles and
the patient characteristics were studied using Chi-squared test
(n=49). Survival curves (disease-specific survival and relapse-free
survival) were plotted using the Kaplan-Meier method (n=49), and
statistical differences were compared using the Wilcoxon test
(n=49). A P-value of <0.05 was considered statistically
significant. Differences in cell growth and migration between the
HSC-3 and HSC-4 cells were assessed by the Student's t-test
(growth, n=6; migration, n=4). All statistical analyses were
performed using XLSTAT-Premium (Addinsoft Corp., New York, NY,
USA).
Results
SOX9 expression profiles in oral
epithelial lesions
Epithelial dysplasia, CIS and SCC were
differentiated according to the expression of perlecan (24), Ki-67 (25) and K17 (26). The tissue specimens from the 49
patients contained 29 foci of normal epithelia, 48 foci of
epithelial dysplasia and 41 foci of CIS (Table I). In normal epithelia (Fig. 1A-E), very weak staining for SOX9 in
the nuclei of parabasal cells was detected in 16 of the 29 foci
(Fig. 1B), and no signals were
detectable in the rest. In epithelial dysplasia with a two-phase
appearance (moderate degree) (Fig.
1F-J), SOX9 positivity was detected in the nuclei of the
proliferating cells in the lower half (Fig. 1G), which were positive for Ki-67
(Fig. 1I) in 38 of the 48 foci. In
differentiated CIS (Fig. 1K-O),
which exhibited a significant degree of keratinization toward the
surface, Ki-67 positivity was mainly observed in the basal zone,
including the first layer of epithelium (Fig. 1N), whereas the upper half layer
(nearer the surface) was positive for K17 (Fig. 1O). Nuclear positivity for SOX9 was
detected throughout the epithelial layer (Fig. 1L) in 38 of the 41 foci. Perlecan was
deposited in the intraepithelial space (Fig. 1M). The number of SOX9-positive cells
was significantly higher than the number of Ki-67-positive cells
(Fig. 1L and N).
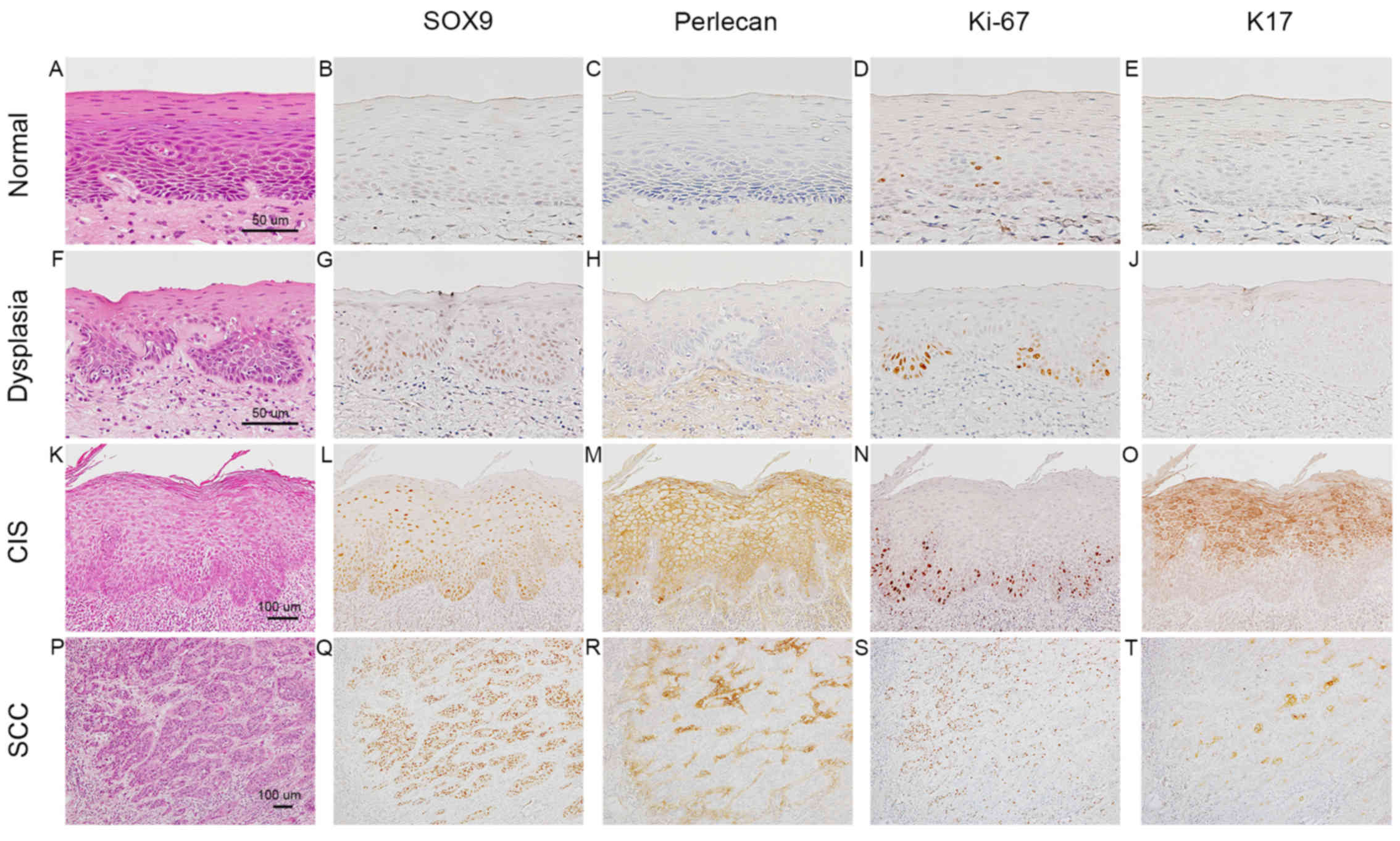 | Figure 1.Immunohistochemical expression
profiles of SOX9 in normal oral epithelia, epithelial dysplasia,
carcinoma in situ (CIS) and squamous cell carcinoma (SCC).
Normal epithelia (A-E), epithelial dysplasia (F-J), differentiated
CIS (K-O), SCC (P-T). Hematoxylin and eosin (H&E) staining (A,
F, K and P); immunoperoxidase staining for SOX9 (B, G, L and Q),
perlecan (C, H, M and R), Ki-67 (D, I, N and S) and keratin (K)17
(E, J, O and T). In normal epithelia (A), SOX9 (B) and perlecan (C)
are not evident. In epithelial dysplasia (F), weak SOX9 positivity
is observed in the nuclei of the parabasal cells (G), parts of
which are also positive for Ki-67 (I). In differentiated CIS (K),
the lower half of the epithelial layer is occupied by SOX9-positive
cells (L), around which perlecan is intraepithelially deposited
(M). In SCC (P), SOX9 expression is enhanced further (Q) and
perlecan is diffusely deposited in the cancer stroma (R). SOX9,
SRY-box 9. |
 | Table I.SOX9 expression profiles in oral
epithelial lesions. |
Table I.
SOX9 expression profiles in oral
epithelial lesions.
|
|
| SOX9-positive foci,
n (%) |
|---|
|
|
|
|
|---|
| Lesion | No. of foci | Nuclear | Cytoplasmic |
|---|
| Normal
epithelia | 29 | 16 (55) | 0 (0) |
| Epithelial
dysplasia | 48 | 38 (79) | 0 (0) |
| CIS | 41 | 38 (93) | 0 (0) |
| SCC | 49 | 43 (88) | 43 (88) |
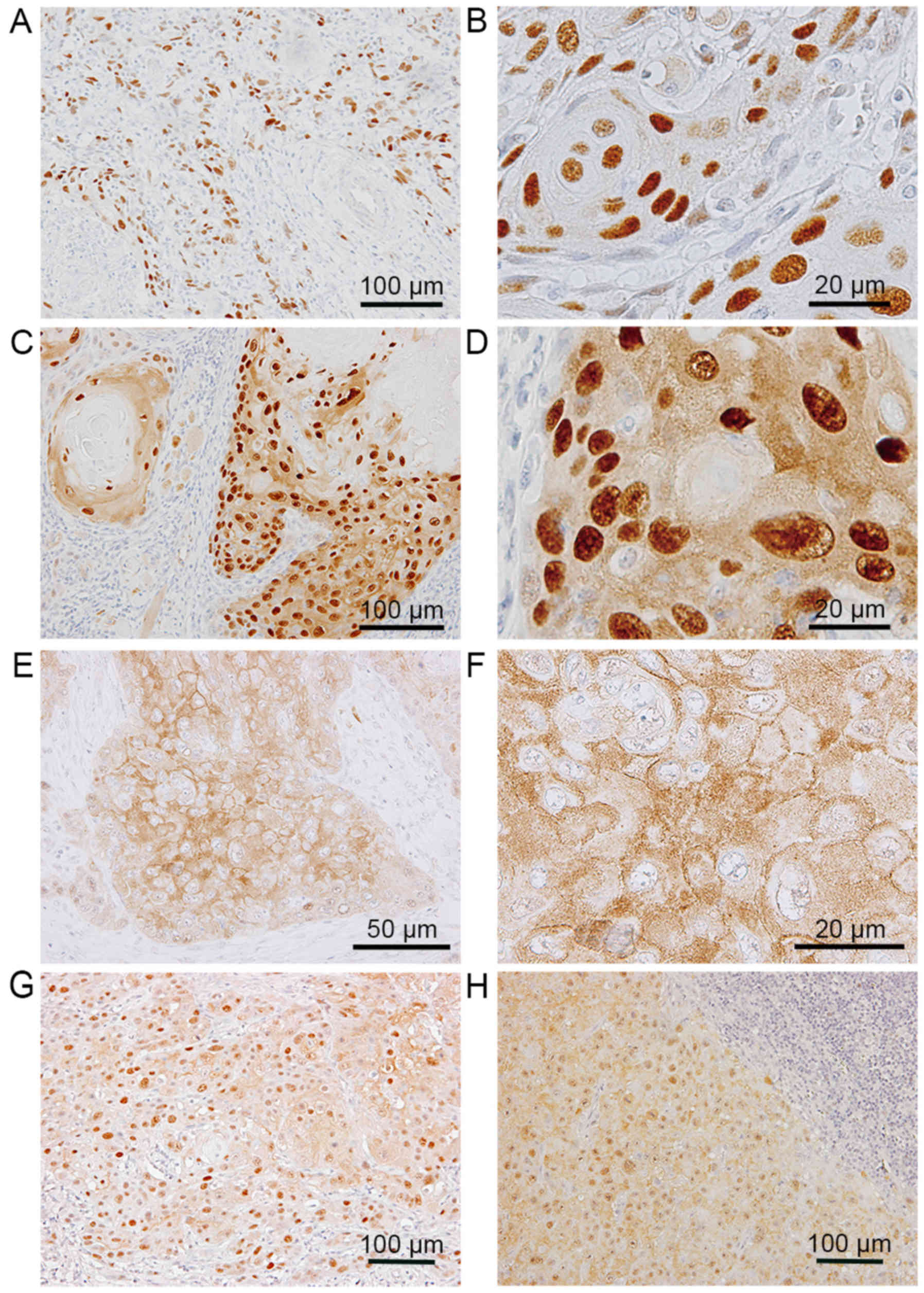
In the invasive nests of SCC (Fig. 1P-T), SOX9 was strongly expressed in
SCC cells in 45 of the 49 cases (Fig.
1Q), and perlecan was abundantly expressed in the stromal space
around the nests (Fig. 1R). K17
positivity was mainly observed in the centers of SCC nests
(Fig. 1T), and was not always
coincident with the area demonstrating SOX9-positive cells. The
immunolocalization of SOX9 was observed in the nuclei of the SCC
cells in 43 of the 45 cases (Fig. 2A
and B). Intriguingly, in 43 of the 45 cases, positive signals
for SOX9 were observed in both the cytoplasm and nuclei of SCC
cells (Fig. 2C and D); however,
cytoplasmic staining was not detectable in epithelial dysplasia and
CIS (Table I). In addition, in some
subdivisions of the SCC cell areas, SOX9 positivity was found only
in the cytoplasm (Fig. 2E and
F).
Correlation between SOX9 localization
and clinicopathological factors
After determining the proportions of the nuclear and
cytoplasmic SOX9-positive areas to the entire area of the SCC, 49
tumors were categorized into two groups using a cut-off point of
25% SOX9 positivity for both nuclear and cytoplasmic proportions.
Consequently, 39 tumors (80%) were categorized as having high
nuclear expression and 35 tumors (71%) were categorized as
exhibiting high cytoplasmic expression. The correlation of high
nuclear and cytoplasmic expression of SOX9 with the
clinicopathological factors is summarized in Table II. High nuclear SOX9 expression did
not correlate with any clinicopathological factors. However, high
cytoplasmic SOX9 expression significantly correlated with regional
recurrence (P=0.026) and female gender (P=0.037), although both
nuclear and cytoplasmic SOX9 expression did not correlate with the
pathological T factor, clinical stage or histologic grade.
| Table II.Relationship between
clinicopathological factors of the OSCC cases and SOX9
expression. |
Table II.
Relationship between
clinicopathological factors of the OSCC cases and SOX9
expression.
|
|
| Nuclear SOX9 | Cytoplasmic
SOX9 |
|---|
|
|
|
|
|
|---|
| Clinicopathological
factors | n | High | Low | P-value | High | Low | P-value |
|---|
| Age (years) |
|
<Median | 24 | 20 | 4 | 0.524 | 16 | 8 | 0.470 |
|
≥Median | 25 | 19 | 6 |
| 19 | 6 |
|
| Sex |
|
Male | 27 | 21 | 6 | 0.727 | 16 | 11 | 0.037 |
|
Female | 22 | 18 | 4 |
| 19 | 3 |
|
| Tumor site |
|
Tongue | 28 | 21 | 7 | 0.796 | 22 | 6 | 0.246 |
| Buccal
mucosa | 5 | 5 | 0 |
| 3 | 2 |
|
| Oral
floor | 5 | 4 | 1 |
| 2 | 3 |
|
|
Gingiva | 7 | 6 | 1 |
| 6 | 1 |
|
|
Palate | 1 | 1 | 0 |
| 1 | 0 |
|
|
Other | 3 | 2 | 1 |
| 1 | 2 |
|
| Pathological T
factor |
| T1 | 17 | 15 | 2 | 0.567 | 12 | 5 | 0.827 |
| T2 | 26 | 19 | 7 |
| 18 | 8 |
|
| T3 | 2 | 2 | 0 |
| 2 | 0 |
|
| T4 | 4 | 3 | 1 |
| 3 | 1 |
|
| Lymph node
metastasis |
|
Absent | 38 | 31 | 7 | 0.521 | 27 | 11 | 0.914 |
|
Present | 11 | 8 | 3 |
| 8 | 3 |
|
| Clinical stage |
| I | 17 | 15 | 2 | 0.388 | 12 | 5 | 0.561 |
| II | 23 | 16 | 7 |
| 15 | 8 |
|
|
III | 3 | 3 | 0 |
| 3 | 0 |
|
| IV | 6 | 5 | 1 |
| 5 | 1 |
|
| Histological
grade |
| G1 | 42 | 32 | 10 | 0.148 | 30 | 12 | 0.694 |
| G2 +
G3 | 7 | 7 | 0 |
| 5 | 2 |
|
| Local
recurrence |
|
Absent | 40 | 33 | 7 | 0.287 | 29 | 11 | 0.726 |
|
Present | 9 | 6 | 3 |
| 6 | 3 |
|
| Regional
recurrence |
|
Absent | 30 | 24 | 6 | 0.929 | 18 | 12 | 0.026 |
|
Present | 19 | 15 | 4 |
| 17 | 2 |
|
| Late distant
metastasis |
|
Absent | 36 | 30 | 6 | 0.280 | 23 | 13 | 0.052 |
|
Present | 13 | 9 | 4 |
| 12 | 1 |
|
Additionally, we examined the metastatic cervical
lymph nodes in 12 of the 14 cases with high cytoplasmic SOX9
expression in the primary lesion. Similar to the primary lesions
(Fig. 2G), the metastatic SCC cells
demonstrated cytoplasmic SOX9 expression (Fig. 2H).
Cytoplasmic SOX9 positivity is
significantly correlated with poor clinical outcomes
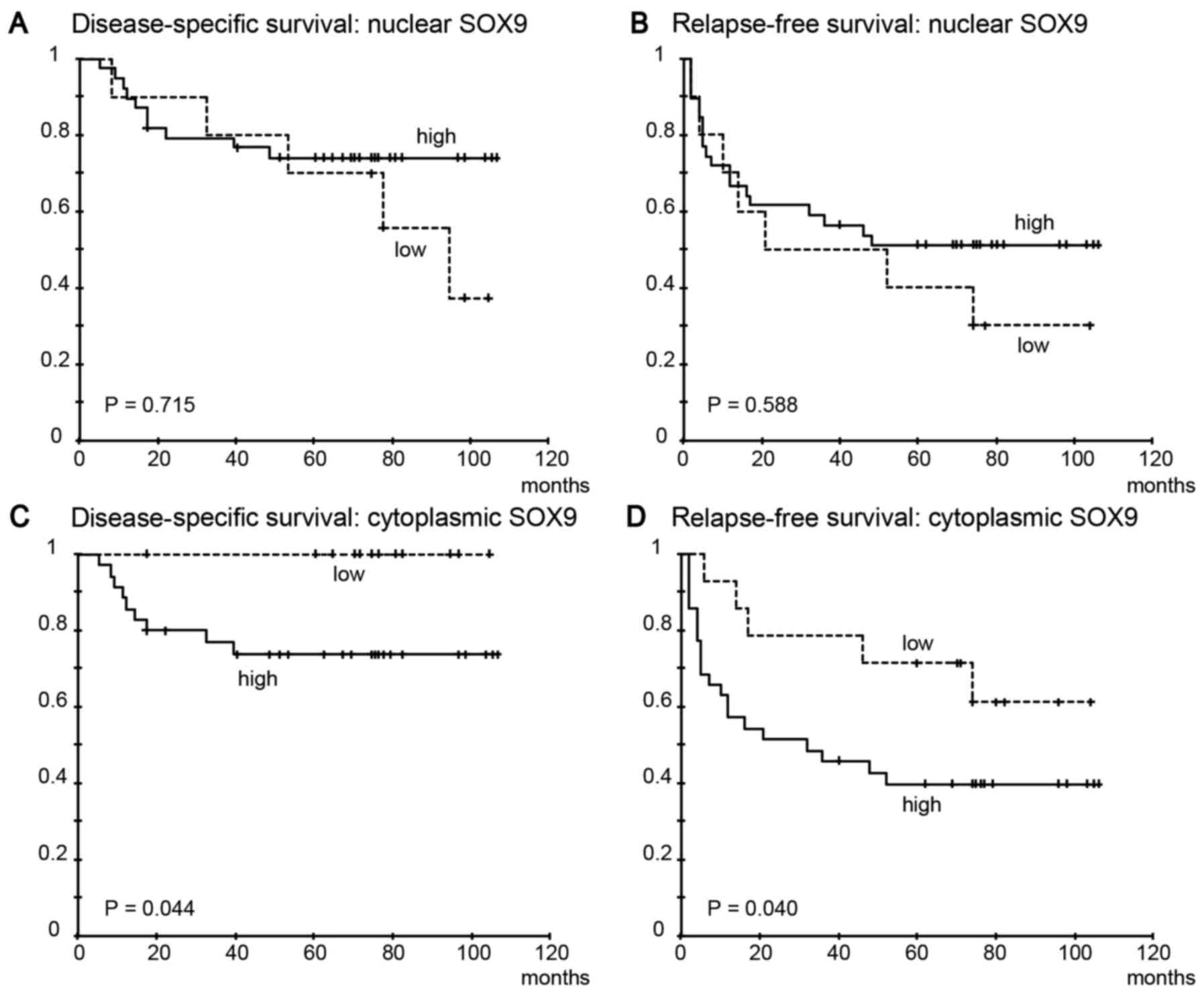
High nuclear SOX9 expression was not significantly
correlated with disease-specific survival (Fig. 3A, P=0.715) or relapse-free survival
(Fig. 3B, P=0.588). However, high
cytoplasmic SOX9 expression was significantly correlated with both
shorter disease-specific survival (Fig.
3C, P=0.044) and relapse-free survival (Fig. 3D, P=0.040).
Cytoplasmic expression of SOX9 is
correlated with higher cell migration ability
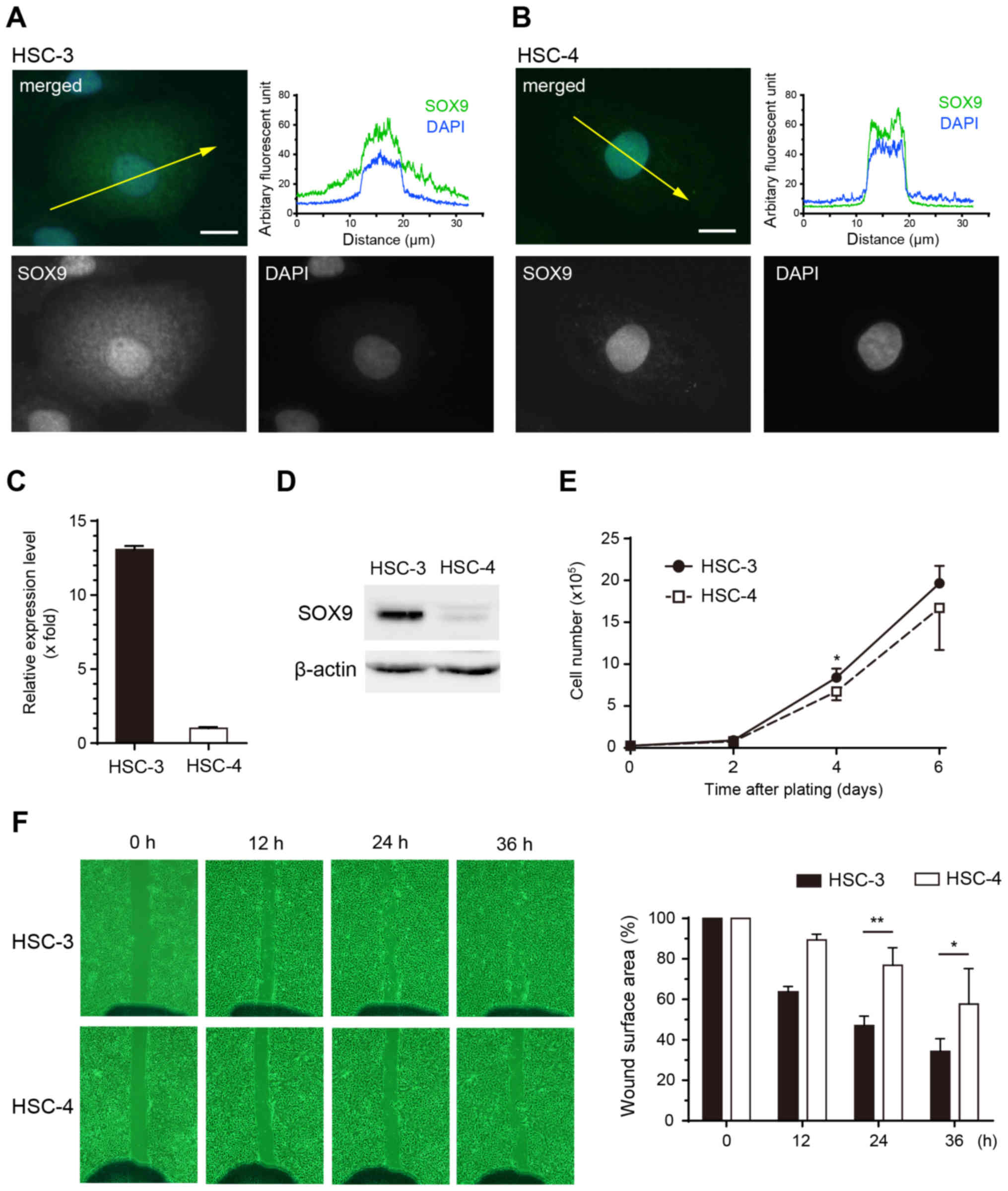
Immunofluoresence staining revealed that SOX9 was
localized in both the nuclei and cytoplasm of HSC-3 cells (Fig. 4A). In contrast, the localization of
SOX9 was limited to the nuclei in HSC-4 cells (Fig. 4B). Both mRNA and protein expression
levels of SOX9 were much higher in HSC-3 cells than these levels in
HSC-4 cells (Fig. 4C and D).
Although there was no significant difference in cell growth
(Fig. 4E), migration ability was
higher in HSC-3 cells than that in HSC-4 cells (Fig. 4F). Taken together, cytoplasmic SOX9
expression in HSC-3 cells might be related to the higher migration
ability of HSC-3 cells.
Discussion
In the present study, we demonstrated for the first
time the elevated expression of SOX9 in oral epithelial lesions,
including epithelial dysplasia, CIS and SCC through
immunohistochemical analysis and posited that the cytoplasmic
expression of SOX9 in SCC cells is a potential prognostic factor
for OSCC.
We successfully demonstrated that SOX9 expression
was elevated in a stepwise manner in epithelial dysplasia, CIS and
SCC. However, only weak signals were detected in the parabasal
cells of the normal epithelia (Fig.
1). Similar to our results, several studies have reported that
compared with non-neoplastic cells in various organs, cancer cells
express SOX9 at higher levels (6,7,11). In
epithelial dysplasia and CIS, SOX9-positive cells were distributed
in the layer farther from the surface, whereas the number of
Ki-67-positive proliferating cells was higher; these data suggest
that SOX9 expression is positively correlated with cell
proliferation during carcinogenesis. The reduction of endogenous
SOX9 expression by RNA interference together with the increased
expression of the cell cycle inhibitor p27 in prostatic cancer cell
lines has been shown to result in growth arrest (7). Meanwhile, the knockdown of SOX9 along
with the upregulation of p21 in lung adenocarcinoma cell lines has
been reported to inhibit cell growth (27). Conversely, in esophageal SCC cells,
SOX9 overexpression has been shown to induce cell proliferation and
tumorigenicity through the activation of PI3K/Akt signaling
(11). Besides, SOX9 expression is
regulated by the Wnt signaling pathway (7). Consistent with our previous report,
the nuclear transition of β-catenin was observed in the lower layer
of epithelial dysplasia (28),
suggesting that the upregulation of SOX9 expression in oral mucosal
epithelial cells may be controlled by the Wnt pathway.
The present study revealed that SOX9 expression was
localized in the cytoplasm as well as nuclei of SCC cells but not
of the cells in epithelial dysplasia and CIS (Fig. 1 and Table I). Chakravarty et al reported
that in the invasive ductal carcinoma of the breast, SOX9
accumulation in the cytoplasm was correlated with enhanced cell
proliferation (8). We speculate
that the elevated SOX9 expression in this study resulted in its
accumulation in the cytoplasm given the fact that cytoplasmic SOX9
was not detected in HSC-4 cells, which only weakly expressed SOX9,
but was detected in HSC-3 cells, which showed strongly expressed
SOX9 (Fig. 4). Malki et al
demonstrated the retention of synthesized SOX9 in the cytoplasm via
interaction with the microtubule network in male pre-Sertoli cells
(29). Cytoplasmic SOX9 expression
in SCC cells may be induced through a similar mechanism. However,
more detailed investigations should be conducted since cytoplasmic
protein localization also results from post-translational
modifications, such as acetylation (30,31).
Furthermore, we demonstrated that cytoplasmic SOX9
expression in a higher proportion of SCC cells was significantly
correlated with the late occurrence of lymph node metastasis and
shorter relapse-free and disease-specific survival (Table II and Fig. 3); these data indicate that SOX9
could be a useful prognostic marker for OSCC, similar to its use in
various types of human cancers (8,9,11).
SOX9 orchestrates the cellular dynamics that involve migration,
invasion and metastasis in addition to cell proliferation (18). Larsimont et al demonstrated
that SOX9 was found to positively regulate the expression of
several ECM components (collagens and laminins), cell adhesion
(integrins), and key regulators of actin cytoskeleton dynamics
(gelsolin and palladin) in mouse models of cutaneous basal cell
carcinoma (18).
Moreover, SOX9 is known to promote
epithelial-mesenchymal transition (EMT) together with SLUG
(32,33), one of the key transcriptional
factors in EMT (34). When SOX9 and
SLUG were overexpressed in breast cancer cells, they cooperatively
facilitated EMT, resulting in elevated tumorigenesis and metastasis
(32). In addition, SOX9 and SLUG
synergistically activated the expression of tenascin C and
periostin, which are important ECM components in cancer invasion
and metastasis (33).
In the present study, we illustrated the elevated
deposition of perlecan around SOX9-positive CIS cells and in the
stromal spaces in SCC (Fig. 1).
HSPG2, which encodes perlecan, is one of the key target
genes of SOX9 (18). In contrast,
perlecan is required for the induction of SOX9 expression in
chondrogenic differentiation (35).
Therefore, a positive feedback loop between SOX9 and perlecan
appears to act in cancer progression. Regarding K17, which is
highly expressed in oral epithelial malignancies (26) and promotes cell proliferation and
migration (36), we observed no
evidence of colocalization with SOX9. Further investigation is
warranted to determine the potential role of SOX9 in regulating K17
expression.
In conclusion, SOX9 may play important roles in the
carcinogenesis and progression of OSCC. SOX9 expression represents
a potential marker for predicting poor prognosis in patients with
OSCC. However, future studies are warranted to elucidate the
precise mechanisms underlying the cytoplasmic localization of SOX9
expression.
Acknowledgements
The authors would like to thank Ms. Shizuko Horiuchi
and Ms. Yuki Takahashi for assistance in the tissue sample
preparation.
Funding
The present study was supported in part by a
Grant-in Aid for Scientific Research from the Japan Society for the
Promotion of Science (JSPS KAKENHI grant no. 15K11006 to MY and
JSPS KAKENHI (grant no. 15K11069 to SM).
Availability of data and materials
This research's data is on ‘figshere’. (https://figshare.com/projects/Cytoplasmic_expression_of_SOX9_as_a_poor_prognostic_factor_for_oral_squamous_cell_carcinoma_/30118).
Author's contributions
YS, MY, SM, RT and JIT conceived and designed the
study. YS, MY, SM, JC and TA performed the experiments. YS and MY
wrote the paper. SM, JC, RT and JIT reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was certified by the Ethics
Committee of Niigata University. The Ethics Board of the Niigata
University Graduate School of Medicine and Dental Sciences (Oral
Life Science) reviewed and approved the experimental protocol for
analyzing the surgical materials (approval no. 12-10-13).
Patient consent for publication
Written informed consent was obtained from all the
patients.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Jo A, Denduluri S, Zhang B, Wang Z, Yin L,
Yan Z, Kang R, Shi LL, Mok J, Lee MJ and Haydon RC: The versatile
functions of Sox9 in development, stem cells, and human diseases.
Genes Dis. 1:149–161. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pritchett J, Athwal V, Roberts N, Hanley
NA and Hanley KP: Understanding the role of SOX9 in acquired
diseases: Lessons from development. Trends Mol Med. 17:166–174.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Harley VR, Clarkson MJ and Argentaro A:
The molecular action and regulation of the testis-determining
factors, SRY (sex-determining region on the Y chromosome) and SOX9
[SRY-related high-mobility group (HMG) box 9]. Endocr Rev.
24:466–487. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Akiyama H: Control of chondrogenesis by
the transcription factor Sox9. Mod Rheumatol. 18:213–219. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vidal VP, Chaboissier MC, Lutzkendorf S,
Cotsarelis G, Mill P, Hui CC, Ortonne N, Ortonne JP and Schedl A:
Sox9 is essential for outer root sheath differentiation and the
formation of the hair stem cell compartment. Curr Biol.
15:1340–1351. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi G, Sohn KC, Li Z, Choi DK, Park YM,
Kim JH, Fan YM, Nam YH, Kim S, Im M, et al: Expression and
functional role of Sox9 in human epidermal keratinocytes. PLoS One.
8:e543552013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang H, McKnight NC, Zhang T, Lu ML, Balk
SP and Yuan X: SOX9 is expressed in normal prostate basal cells and
regulates androgen receptor expression in prostate cancer cells.
Cancer Res. 67:528–536. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chakravarty G, Moroz K, Makridakis NM,
Lloyd SA, Galvez SE, Canavello PR, Lacey MR, Agrawal K and Mondal
D: Prognostic significance of cytoplasmic SOX9 in invasive ductal
carcinoma and metastatic breast cancer. Exp Biol Med. 236:145–155.
2011. View Article : Google Scholar
|
|
9
|
Lü B, Fang Y, Xu J, Wang L, Xu F, Xu E,
Huang Q and Lai M: Analysis of SOX9 expression in colorectal
cancer. Am J Clin Pathol. 130:897–904. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vidal VP, Ortonne N and Schedl A: SOX9
expression is a general marker of basal cell carcinoma and
adnexal-related neoplasms. J Cutan Pathol. 35:373–379. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hong Y, Chen W, Du X, Ning H, Chen H, Shi
R, Lin S, Xu R, Zhu J, Wu S, et al: Upregulation of sex-determining
region Y-box 9 (SOX9) promotes cell proliferation and
tumorigenicity in esophageal squamous cell carcinoma. Oncotarget.
6:31241–31254. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ikarashi T, Ida-Yonemochi H, Ohshiro K,
Cheng J and Saku T: Intraepithelial expression of perlecan, a
basement membrane-type heparan sulfate proteoglycan reflects
dysplastic changes of the oral mucosal epithelium. J Oral Pathol
Med. 33:87–95. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maruyama S, Shimazu Y, Kudo T, Sato K,
Yamazaki M, Abé T, Babkair H, Cheng J, Aoba T and Saku T:
Three-dimensional visualization of perlecan-rich neoplastic stroma
induced concurrently with the invasion of oral squamous cell
carcinoma. J Oral Pathol Med. 43:627–636. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Maruyama S, Cheng J, Yamazaki M, Liu A and
Saku T: Keratinocyte growth factor colocalized with perlecan at the
site of capsular invasion and vascular involvement in salivary
pleomorphic adenomas. J Oral Pathol Med. 38:377–385. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Toyoshima K, Kimura S, Cheng J, Oda Y,
Mori KJ and Saku T: High-molecular-weight fibronectin synthesized
by adenoid cystic carcinoma cells of salivary gland origin. Jpn J
Cancer Res. 90:308–319. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hasegawa M, Cheng J, Maruyama S, Yamazaki
M, Abé T, Babkair H, Saito C and Saku T: Differential
immunohistochemical expression profiles of perlecan-binding growth
factors in epithelial dysplasia, carcinoma in situ, and squamous
cell carcinoma of the oral mucosa. Pathol Res Pract. 212:426–436.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Warren CR, Grindel BJ, Francis L, Carson
DD and Farach-Carson MC: Transcriptional activation by NFkappaB
increases perlecan/HSPG2 expression in the desmoplastic prostate
tumor microenvironment. J Cell Biochem. 115:1322–1333. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Larsimont JC, Youssef KK, Sánchez-Danés A,
Sukumaran V, Defrance M, Delatte B, Liagre M, Baatsen P, Marine JC,
Lippens S, et al: Sox9 controls self-renewal of oncogene targeted
cells and links tumor initiation and invasion. Cell Stem Cell.
17:60–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
UICC International Union Against Cancer:
TNM Classification of Malignant Tumours. Sobin LH, Gospodarowicz MK
and Wittekind C: 7th edition. Wiley-Blackwell; Hoboken, NJ: pp.
25–28. 2009
|
|
20
|
Saku T and Furthmayr H: Characterization
of the major heparan sulfate proteoglycan secreted by bovine aortic
endothelial cells in culture. Homology to the large molecular
weight molecule of basement membranes. J Biol Chem. 264:3514–3523.
1989.PubMed/NCBI
|
|
21
|
Momose F, Araida T, Negishi A, Ichijo H,
Shioda S and Sasaki S: Variant sublines with different metastatic
potentials selected in nude mice from human oral squamous cell
carcinomas. J Oral Pathol Med. 18:391–395. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamazaki M, Maruyama S, Abé T, Essa A,
Babkair H, Cheng J and Saku T: MFG-E8 expression for progression of
oral squamous cell carcinoma and for self-clearance of apoptotic
cells. Lab Invest. 94:1260–1272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ahsan MS, Yamazaki M, Maruyama S,
Kobayashi T, Ida-Yonemochi H, Hasegawa M, Ademola Henry A, Cheng J
and Saku T: Differential expression of perlecan receptors,
α-dystroglycan and integrin β1, before and after invasion of oral
squamous cell carcinoma. J Oral Pathol Med. 40:552–559. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kobayashi T, Maruyama S, Cheng J,
Ida-Yonemochi H, Yagi M, Takagi R and Saku T: Histopathological
varieties of oral carcinoma in situ: Diagnosis aided by
immunohistochemistry dealing with the second basal cell layer as
the proliferating center of oral mucosal epithelia. Pathol Int.
60:156–166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mikami T, Cheng J, Maruyama S, Kobayashi
T, Funayama A, Yamazaki M, Adeola HA, Wu L, Shingaki S, Saito C and
Saku T: Emergence of keratin 17 vs. loss of keratin 13: Their
reciprocal immunohistochemical profiles in oral carcinoma in situ.
Oral Oncol. 47:497–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang SS, Fang WT, Hou YH, Huang SF, Yen
BL, Chang JL, Li SM, Liu HP, Liu YL, Huang CT, et al: Upregulation
of SOX9 in lung adenocarcinoma and its involvement in the
regulation of cell growth and tumorigenicity. Clin Cancer Res.
16:4363–4373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Alvarado CG, Maruyama S, Cheng J,
Ida-Yonemochi H, Kobayashi T, Yamazaki M, Takagi R and Saku T:
Nuclear translocation of β-catenin synchronized with loss of
E-cadherin in oral epithelial dysplasia with a characteristic
two-phase appearance. Histopathology. 59:283–291. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Malki S, Berta P, Poulat F and
Boizet-Bonhoure B: Cytoplasmic retention of the sex-determining
factor SOX9 via the microtubule network. Exp Cell Res. 309:468–475.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oz Bar M, Kumar A, Elayyan J, Reich E,
Binyamin M, Kandel L, Liebergall M, Steinmeyer J, Lefebvre V and
Dvir-Ginzberg M: Acetylation reduces SOX9 nuclear entry and
ACAN gene transactivation in human chondrocytes. Aging Cell.
15:499–508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sim H, Argentaro A and Harley VR: Boys,
girls and shuttling of SRY and SOX9. Trends Endocrinol Metab.
19:213–222. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo W, Keckesova Z, Donaher JL, Shibue T,
Tischler V, Reinhardt F, Itzkovitz S, Noske A, Zürrer-Härdi U, Bell
G, et al: Slug and Sox9 cooperatively determine the mammary stem
cell state. Cell. 148:1015–1028. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fazilaty H, Gardaneh M, Akbari P, Zekri A
and Behnam B: SLUG and SOX9 cooperatively regulate tumor initiating
niche factors in breast cancer. Cancer Microenviron. 9:71–74. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Savagner P, Yamada KM and Thiery JP: The
zinc-finger protein slug causes desmosome dissociation, an initial
and necessary step for growth factor-induced epithelial-mesenchymal
transition. J Cell Biol. 137:1403–1419. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sadatsuki R, Kaneko H, Kinoshita M, Futami
I, Nonaka R, Culley KL, Otero M, Hada S, Goldring MB, Yamada Y, et
al: Perlecan is required for the chondrogenic differentiation of
synovial mesenchymal cells through regulation of Sox9 gene
expression. J Orthop Res. 35:837–846. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Khanom R, Nguyen CT, Kayamori K, Zhao X,
Morita K, Miki Y, Katsube K, Yamaguchi A and Sakamoto K: Keratin 17
is induced in oral cancer and facilitates tumor growth. PLoS One.
11:e01611632016. View Article : Google Scholar : PubMed/NCBI
|