Introduction
Pancreatic cancer is the third most common cause of
cancer-associated mortality in the United States, and is the
leading cause of mortality and morbidity worldwide (1). Advances in medical technology and
chemotherapy over the past few decades have increased overall
survival; however, since long-term survival rates are still low,
novel therapies are required (2,3). At
present, surgical resection is considered a major treatment for
patients with pancreatic cancer. Since pancreatic cancer has no
early symptoms, in the majority of patients tumor metastasis is
observed before resection of malignant tumors (4,5). Even
if surgical resection is performed early, almost all patients
experience recurrence following surgery, or eventually reach a
pathological state due to metastatic growth (5). Therefore, therapeutic strategies that
specifically inhibit or prevent invasion or metastasis may
significantly improve the prognosis of pancreatic cancer.
Interferon (IFN)-γ promotes the transcriptional
activation of IFN-γ-inducible genes, predominantly through the
Janus kinase (JAK)/signal transducer and activator of transcription
(STAT) intracellular signaling pathway (6). When IFN-γ binds to cell surface
receptors, the receptor-associated tyrosine kinases JAK1 and JAK2
are activated. JAKs phosphorylate STAT-1; phosphorylated (p)-STAT1
is dimerized and translocated to the nucleus, where it binds to the
IFN-γ-activated site element of IFN-γ responsive genes, thus
resulting in gene activation (7).
In general, STAT1 is considered a tumor suppressor; however,
paradoxical evidence supporting the tumor-promoting function of
STAT1 has emerged. Previous studies have demonstrated that extended
IFN signaling or constitutive STAT1 signaling promotes tumor
growth, as well as resistance to chemotherapy and radiation
(8–13). STAT1 enhances cancer cell growth,
invasion and exhibits various activities, including inhibition of
cell death and induction of therapeutic resistance (14).
Rhus verniciflua Stokes (RVS) is a member of
the Anacardiaceae family, which is also known as the lacquer tree,
and has been used for the treatment of gastric diseases, including
tumors, for centuries in Traditional Korean Medicine (TKM)
(15,16). RVS contains phenolic compounds, such
as fustin, fisetin, sulfuretin, butein, gallic acid and kaempferol.
It has been reported that RVS exhibits antifibrogenic (17), antiproliferative (16,18),
antioxidant (19,20) and antitumorigenic (21–23)
activities. In clinical use, urushiol should be removed from RVS,
as it may induce an allergic skin rash in sensitive individuals.
Allergen-removed RVS (aRVS) has also been revealed to exert
anticancer effects in preclinical studies, and has potential as an
anticancer therapeutic agent for the treatment of patients with
advanced cancer, including pancreatic cancer (24–26).
Since cancer cell invasion and metastasis involve
STAT1 activation and aRVS has been reported to possess
antitumorigenic activity, the present study investigated whether
aRVS serves a pivotal role in this process. The present results
demonstrated that activation and upregulation of STAT1 by IFN-γ
initiated the induction of pancreatic cancer cell invasion and
metastasis, whereas aRVS suppressed these carcinogenic processes,
potentially through inhibition of IFN-γ-induced STAT1
activation.
Materials and methods
Materials
aRVS was obtained from Bflux Pharma Corp (Seoul,
South Korea). The voucher specimen was registered and deposited at
East-West Medical Research Institute, Kyung Hee University (Seoul,
South Korea). The preparative process and quality control of aRVS
was conducted according to the standard operational procedure of
Bflux Pharma Corp (Korean patent no. 10-2016-0101802) (27). Briefly, RVS stalk, which was 10
years old, was purchased from Kyung Hee Herb Pharm (Wonju, Korea).
The dried RVS stalk was chopped into pieces, and extracted twice
with a 4-fold volume of purified water at 105-110°C for 4 h. The
extract was filtered using a 1-mm filtration device, and was vacuum
concentrated. The concentrate was lyophilized to powder form,
resulting in a 3% yield of aRVS extract. Quality control was
performed using high-performance liquid chromatography (fisetin,
>10.0%; fustin, >8.0%; gallic acid, >8.0%; sulfuretin,
>1.5%; urushiol, not detected), a pesticide detection test and a
residual microorganism test. aRVS was dissolved in 50% methanol as
a 200 mg/ml stock solution and stored at −20°C. Further dilution
was conducted in cell culture medium. Dulbecco's modified Eagle's
medium (DMEM), RPMI-1640 medium, fetal bovine serum (FBS), 0.25%
trypsin-EDTA and antibiotic-antimycotic (100 U/ml penicillin and
streptomycin) were purchased from Gibco; Thermo Fisher Scientific,
Inc. (Waltham, MA, USA). MTT, dimethyl sulfoxide (DMSO) and SDS
were purchased from Sigma-Aldrich (Merck KgaA, Darmstadt, Germany).
Matrigel was obtained from BD Biosciences (Franklin Lakes, NJ,
USA), type I collagen was purchased from Costar (Corning
Incorporated, Corning, NY, USA), and IFN-γ was purchased from
BioLegend, Inc. (San Diego, CA, USA). Antibodies against p-Src
(cat. no. 2101), Src (cat. no. 2108), X-linked inhibitor of
apoptosis protein (XIAP) (cat. no. 2042), cleaved-caspase-3 (cat.
no. 9661), p-JAK1(cat. no. 3331), p-JAK2 (cat. no. 8082), p-STAT1
(Y701, S727) (cat. no. 7649), STAT1 (cat. no. 9172), p-STAT3 (Y705)
(cat. no. 9145), p-focal adhesion kinase (FAK) (cat. no. 3284) and
FAK (cat. no. 13009), as well as secondary antibodies (anti-rabbit;
cat. no. 7074, anti-mouse; cat. no. 7076), were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Matrix
metalloproteinase 9 (MMP9) (cat. no. sc-21733), B-cell lymphoma 2
(Bcl2) (cat. no. sc-492), cyclin D1 (cat. no. sc-718), JAK1 (cat.
no. sc-277), JAK2 (cat. no. sc-278), STAT3 (cat. no. sc-482) and
β-actin (cat. no. sc-47778) antibodies were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). MUC4 (cat. no. ab60720)
antibody was purchased from Abcam (Abcam, Cambridge, MA).
Cell culture
A total of three pancreatic cancer cell lines
(PANC-1, HPAC and Bxpc3) were obtained from the American Type
Culture Collection (Manassas, VA, USA). The PANC-1 and HPAC
immortalized human pancreatic cancer cell lines were cultured in
DMEM supplemented with 10% FBS and 100 U/ml
penicillin-streptomycin. Bxpc3 pancreatic cancer cells were
cultured in RPMI-1640 supplemented with 10% FBS and 100 U/ml
penicillin-streptomycin. All cells were maintained at 37°C in a
humidified atmosphere containing 5% CO2. After attaining
80% confluence, the cells were subcultured by trypsinization with
trypsin-EDTA solution.
Cell viability assay
Cell viability and cell number were determined using
an MTT (purity >95%) assay. HPAC (1×104 cells/well),
PANC (1×104 cells/well) and Bxpc3 (1×104
cells/well) were respectively seeded into 96-well plates with 100
µl culture medium and were treated with the indicated
concentrations (50, 100 and 200 µM) of aRVS for 24 h, and 20 ng/ml
IFN-γ for 24 h at 37°C. Subsequently, 20 µl MTT (5 mg/ml) solution
was added to each well, and the cells were incubated for 4 h at
37°C. Once the medium was carefully removed, 150 µl DMSO was added
and agitated to dissolve the formazan crystals. The absorbance at
490 nm was measured using Tecan Sunrise Eliza-Reader (Tecan Group
Ltd., Mannedorf, Switzerland). For relative quantification, the
value of absorbance in each group was normalized to that of the
control group.
Western blotting
Cells were treated with IFN-γ (20 ng/ml) and aRVS
(200 µg/ml), or with IFN-γ (20 ng/ml) alone, for 24 or 48 h at
37°C, and were then washed and collected. Total protein was
extracted using radioimmunoprecipitation assay buffer (Cell
Signaling Technology Inc.) containing a complete protease inhibitor
cocktail tablet (Roche Diagnostics, Mannheim, Germany), and
proteins were then incubated on ice. After 10 min, cells were
centrifuged at 16,000 × g for 10 min at 4°C, and the protein
concentration was measured using the bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). Subsequently,
30 µg protein samples were separated by 10% SDS-PAGE, after which,
proteins were transferred to a polyvinylidene difluoride membrane.
The membrane was blocked with 5% skim milk in Tris-buffered saline
with 0.1% Tween-20 (TBST) at room temperature for 1 h. The membrane
was washed three times with TBST and incubated with the primary
antibodies overnight at 4°C. The membrane was then washed three
times in TBST, and incubated with horseradish peroxidase
(HRP)-conjugated goat anti-rabbit (dilution 1:2,000; cat. no. 7074)
and HRP-conjugated goat anti-mouse secondary antibodies (dilution
1:2,000; cat. no. 7076) (both Cell Signaling Technology, Inc.) for
1 h at room temperature. The immunoreactive proteins were
visualized using SuperSignal™ West Pico PLUS Chemiluminescent
Substrate (cat. no. 34580) or SuperSignal™ West Dura Extended
Duration Substrate (cat. no. 34075) (Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocols. Images were
captured using an Imagequant™ LAS 4000 (GE Healthcare Japan, Tokyo,
Japan).
Dilutions for primary antibodies used in the present
study are as follows; MUC4 (1:1,000), p-Src (1:1,000), Src
(1:1,000), XIAP (1:1,000), cleaved-caspase-3 (1:1,000), p-JAK1
(1:1,000), p-JAK2 (1:1,000), p-STAT1 (Y701, S727) (1:1,000), STAT1
(1:1,000), p-STAT3 (Y705) (1:1,000), p-FAK (1:1,000), FAK
(1:1,000), JAK1 (1:1,000), JAK2 (1:1,000), STAT3 (1:1,000) and
β-actin (1:2,000).
Matrigel-invasion assay
The in vitro invasion assay was performed
using a 24-well Transwell unit (pore size, 8 µm) with polycarbonate
membranes (Costar; Corning Incorporated). The upper and lower sides
of the membrane were coated with Matrigel (1 mg/ml) and type I
collagen (0.5 mg/ml). The lower chamber was filled with 10%
FBS-containing medium or serum-free medium containing IFN-γ (20
ng/ml). Cells (1×105/ml) were placed in the upper
chamber of the Transwell unit and were cultured for 18 h at 37°C
with or without aRVS (200 µg/ml). The noninvading cells on the
upper surface of membrane were removed from the chamber, and the
invading cells on the lower surface of the membrane were stained
with Quick-Diff stain kit (BD Biosciences). Briefly, the cells were
fixed in REASTAIN Quick-Diff Fix for 5 min, and were then stained
with REASTAIN Quick-Diff Red followed by REASTAIN Quick-Diff Blue
for 10 min each. The number of invasive cells was counted in five
randomly selected fields under a Nikon Ti-U microscope (Nikon
Corporation, Tokyo, Japan) at ×200 magnification.
Wound healing assay
Once cells reached 80% confluence, they were
pretreated with mitomycin C (25 µg/ml) in serum-free medium for 30
min to suppress cell proliferation before a wound was made to the
cell monolayer using a 200-µl pipette tip (28,29).
In all subsequent experimental steps, medium contained mitomycin C.
After washing with serum-free medium, cells were incubated with 10%
FBS-containing medium or serum-free medium containing IFN-γ (20
ng/ml) in the presence or absence of aRVS (200 µg/ml) for 24 h at
37°C. The migration of the cells at the edge of the scratch was
analyzed at 0, 18 and 24 h, when microscopic images of the cells
were captured. Images were captured using a digital camera system
(Nikon Corporation) connected to a light microscope (Olympus
America, Inc., Melville, NY, USA) at different time points.
Statistical analysis
The results obtained from each experiment are
expressed as the means ± standard deviation from at least three
independent experiments. Statistical analysis was performed using
one way analysis of variance followed by Tukey's post hoc test for
multiple comparisons. GraphPad Prism 5.0 software (GraphPad
Software, Inc., San Diego, CA, USA) was used to analyze data.
P<0.05 was considered to indicate a statistically significant
difference.
Results
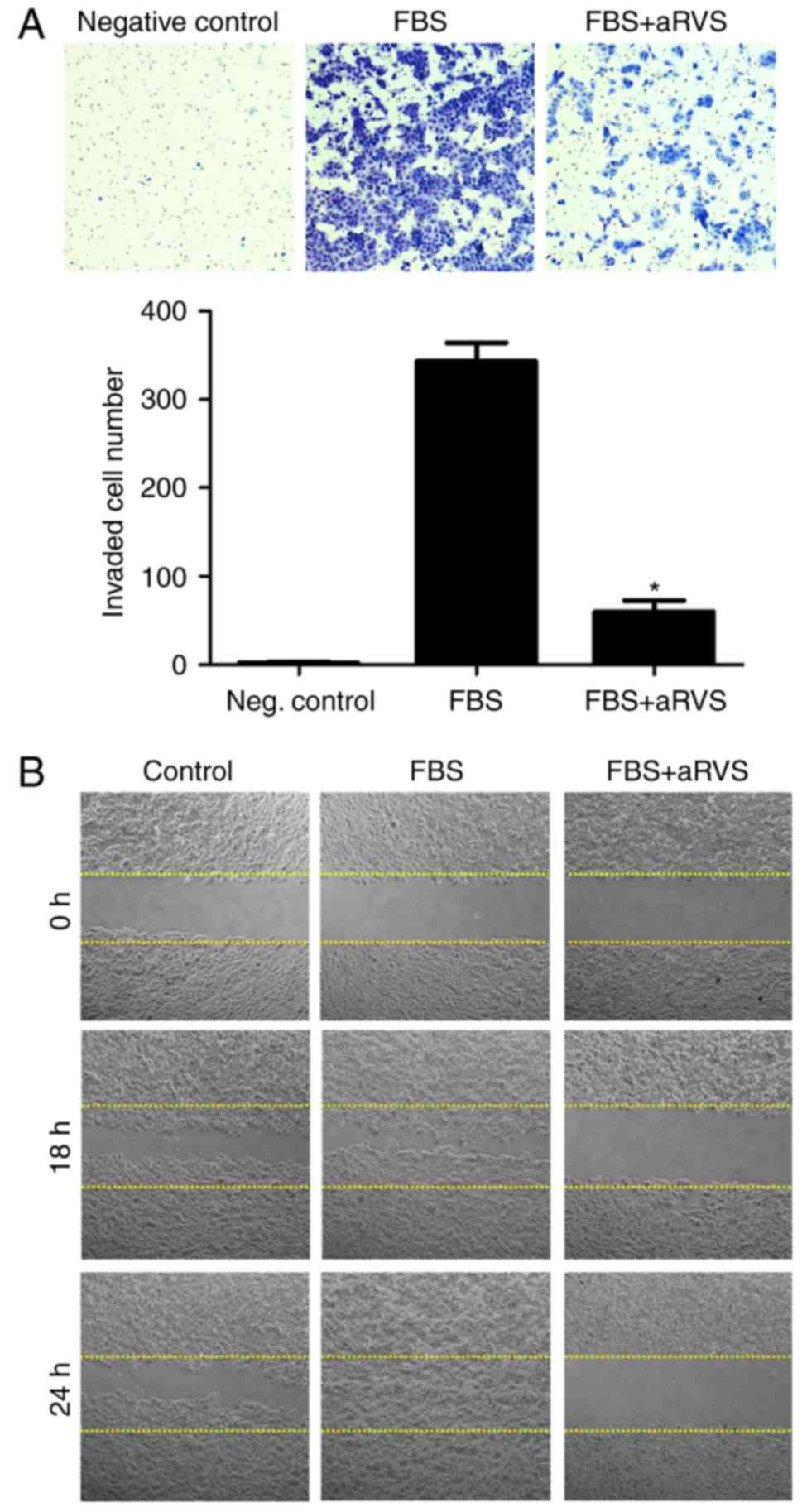
aRVS inhibits cell invasion and
migration of pancreatic cancer cells
To evaluate the therapeutic potential of aRVS in
pancreatic cancer, its effects on invasion and migration were
analyzed using Matrigel invasion and wound healing assays. The
preliminary results demonstrated that HPAC cells had the highest
metastatic capacity among several pancreatic cancer cell lines
(data not shown); therefore, HPAC cells were used for the following
experiments. As shown in Fig. 1A,
cells treated with aRVS experienced a significant decrease in the
number of invading cells compared with those treated with 10% FBS.
The wound healing assay also indicated that 10% FBS-treated cells
migrated across the wound much faster than aRVS-treated cells
(Fig. 1B). These results indicated
that aRVS significantly inhibited the invasion and migration of
HPAC pancreatic cancer cells.
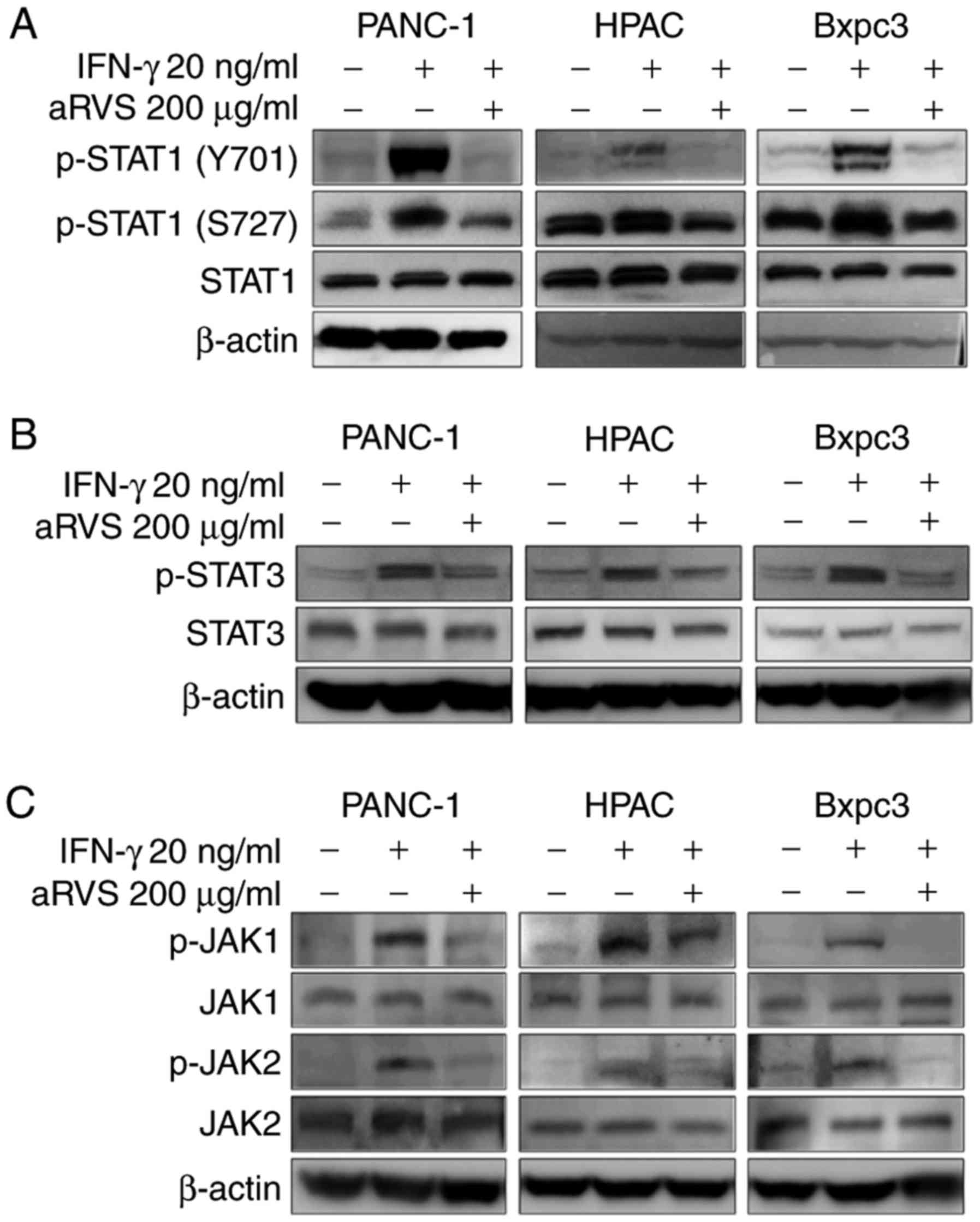
aRVS downregulates the IFN-γ-induced
STAT1 signaling pathway
STAT1 and STAT3 transcription factors serve an
important role in tumorigenesis. Despite the controversial nature
of the effects of STAT1 on tumorigenesis, STAT3 strongly induces
cancer invasion or migration (14,30,31).
Therefore, the present study examined whether aRVS regulated the
activity of STAT1 or STAT3, thus leading to tumor invasion and
migration. The results demonstrated that aRVS inhibited the
IFN-γ-induced activation of STAT1 and STAT3; however, STAT1 was
more strongly inhibited by aRVS in IFN-γ-stimulated PANC-1, HPAC
and Bxpc3 cells (Fig. 2A and B). In
the process of STAT1 activation, the phosphorylation of Ser727 is
required for the dimerization of STAT1, and the phosphorylation of
Tyr701 is essential for translocation of STAT1 into the nucleus
(32,33). Upon examination, the present study
demonstrated that aRVS inhibited the phosphorylation of both serine
and tyrosine residues, thus suggesting that aRVS may block the
phosphorylation of both residues. However, the suppression of
Tyr701 phosphorylation in STAT1 by aRVS was more marked than that
of Ser727. Based on these results, the present study aimed to
determine whether aRVS inhibited the upstream kinases of the STAT
signaling pathway. As shown in Fig.
2C, aRVS also inhibited the activity of JAK1 and JAK2.
aRVS inhibits IFN-γ-induced invasion,
migration and proliferation
The present study also investigated whether the
activation of STAT1 by IFN-γ induced invasion, migration and
proliferation of pancreatic cancer cells, and examined whether this
was inhibited by aRVS. The results demonstrated that IFN-γ induced
the invasion, migration and proliferation of pancreatic cancer
cells, and that aRVS treatment significantly inhibited these
effects (Fig. 3). The wound healing
assay indicated that IFN-γ-treated cells migrated much faster than
cells treated with aRVS (Fig. 3B).
In the invasion assay, treatment with aRVS resulted in a
significantly reduced number of invasive cells compared with in the
IFN-γ-treated group (Fig. 3A). In
addition, aRVS significantly inhibited IFN-γ-induced proliferation
of pancreatic cancer cells in a concentration-dependent manner
(Fig. 3C). The present study also
confirmed that the inhibitory effects of aRVS were not due to
toxicity (data not shown).
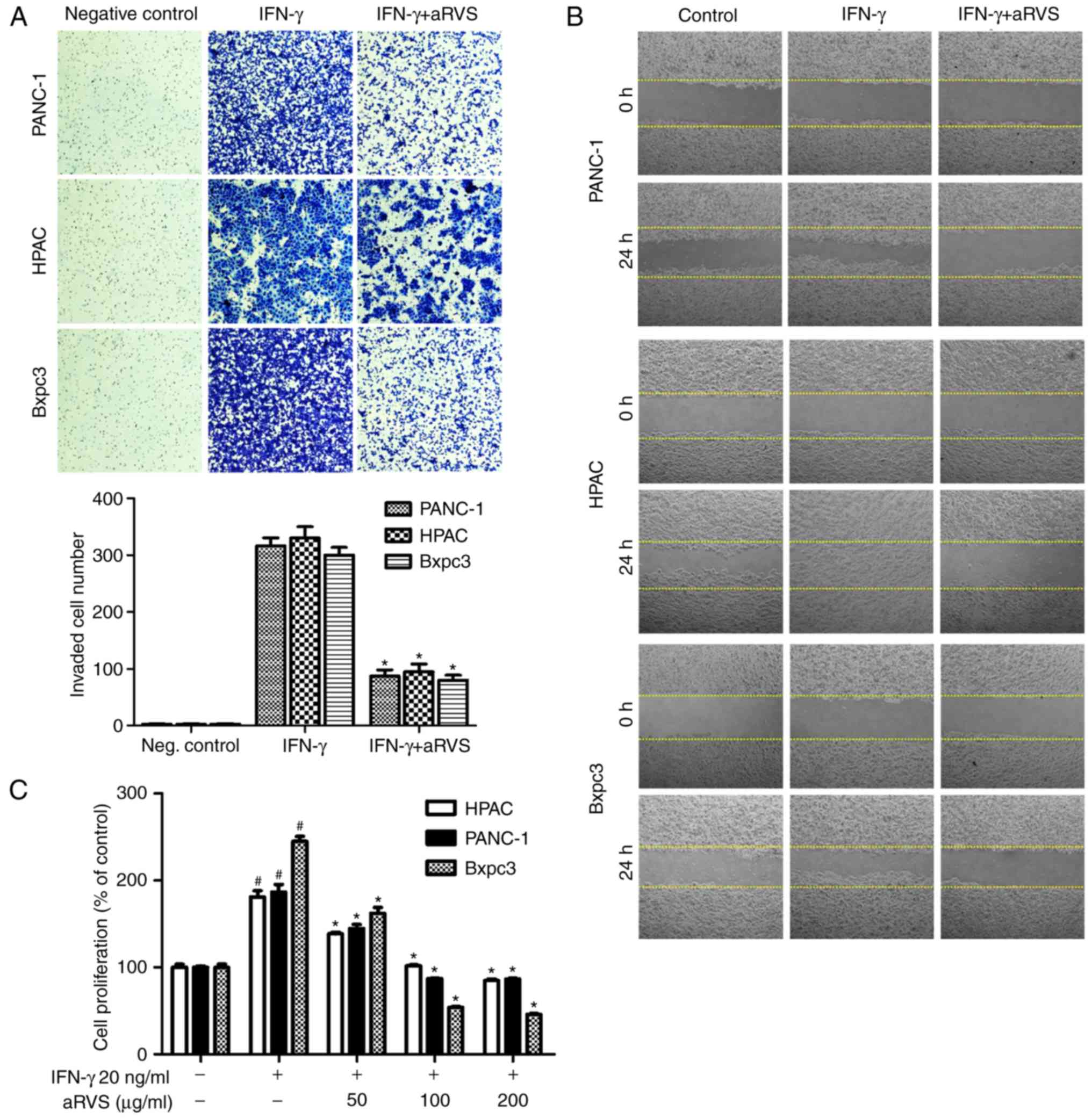 | Figure 3.aRVS suppresses IFN-γ-induced
invasion and migration of various pancreatic cancer cells. (A)
Matrigel-invasion assay determined the inhibitory effects of aRVS
on PANC-1, HPAC and Bxpc3 pancreatic cancer cell invasion. Cells
were treated with IFN-γ and aRVS, or with IFN-γ alone, and equal
numbers of the cells were seeded into the upper chamber of a
Matrigel-coated Transwell system. After 18 h at 37°C, non-invading
cells on the upper part of the membrane were removed with a cotton
swab, and the invasive cells were fixed and stained. The number of
cells was quantified in five random fields. Quantitative results
were obtained from ×200 magnification images. *P<0.01 vs. the
IFN-γ- treated group. (B) Wound healing assay determined the
effects of aRVS on the migratory ability of PANC-1, HPAC and Bxpc3
cells. Confluent cells were scratched with a 200-µl sterile pipette
tip at the center of the well, and cells were treated with IFN-γ
and aRVS, or with IFN-γ alone for 24 h at 37°C. Images of the
wounded monolayer were captured at the indicated time points under
a light microscope with ×100 magnification. (C) Serum-starved cells
(1×104 cells) were stimulated with IFN-γ in the presence
or absence of aRVS, and the number of viable cells was measured
using the MTT assay. #P<0.05 vs. the Control group;
*P<0.05 vs. the IFN-γ-treated group. Data are presented as the
means ± standard error of the mean from three independent
experiments. aRVS, allergen-removed Rhus verniciflua Stokes;
IFN-γ, interferon-γ. |
Our preliminary experiments revealed that HPAC cell
mobility was the most abundant and aRVS exhibited potent inhibitory
effects on all three pancreatic cell lines. Subsequently, the
expression levels of MMP9, which targets various extracellular
proteins during invasion and metastasis, were analyzed, in order to
confirm the ability of aRVS to inhibit the invasion and metastasis
of pancreatic cancer cells. The results indicated that the
expression levels of MMP9 were lower in cells treated with aRVS
compared with in those treated with IFN-γ (Fig. 4). Based on these results, it was
suggested that aRVS may inhibit the invasion and metastasis of
pancreatic cancer cells via the inhibition of STAT1.
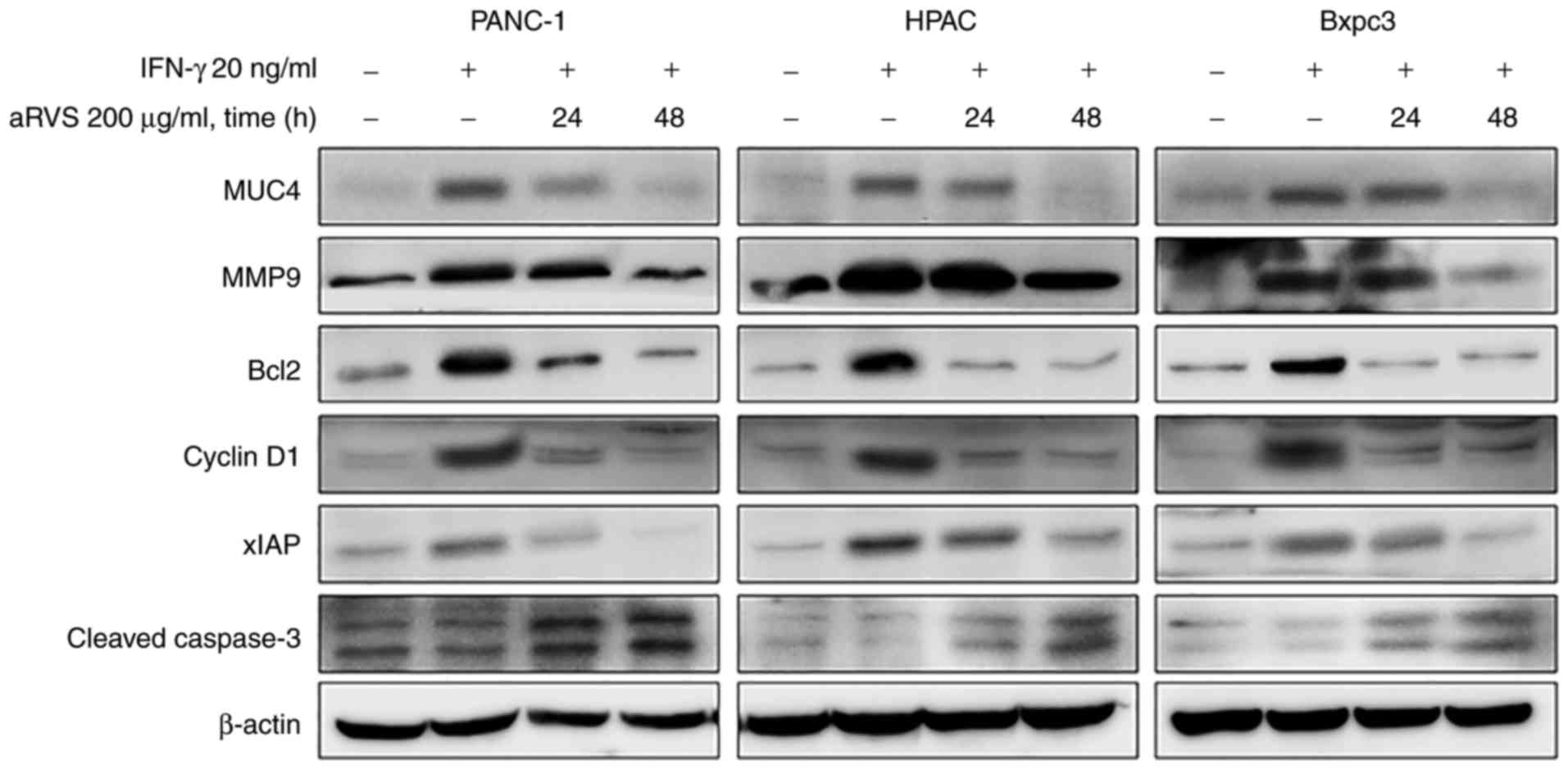 | Figure 4.aRVS regulates the expression of
MUC4, and anti-apoptotic and cell cycle regulatory proteins. Cells
were treated with IFN-γ and aRVS, or with IFN-γ alone for the
indicated time points, and whole cell extracts were collected.
Protein expression was determined by western blotting with
antibodies against MUC4, MMP9, Bcl2, cyclin D1, xIAP and cleaved
caspase-3 proteins. β-actin was used as a loading control.
Representative data of three independent experiments are shown.
aRVS, allergen-removed Rhus verniciflua Stokes; Bcl2, B-cell
lymphoma 2; IFN-γ, interferon-γ; MMP9, matrix metalloproteinase 9;
MUC4, mucin 4; xIAP, X-linked inhibitor of apoptosis protein. |
aRVS downregulates mucin 4 (MUC4)
expression in pancreatic cancer cells
It has previously been indicated the involvement of
MUC4 in pancreatic cancer cell motility and invasion; furthermore,
it promotes resistance to apoptosis when cells are treated with
various chemotherapeutic agents (34). In addition, the activated
transcription factors STAT1 and STAT3 have been reported to serve
as potential regulators of MUC4 expression in pancreatic cancer
cells (35). Based on these
reports, the present study determined whether aRVS could affect
MUC4 expression in pancreatic cancer cells. PANC-1, HPAC and Bxpc3
cells were untreated for 48 h, or were treated with IFN-γ with or
without aRVS for 24 and 48 h. The cells were then harvested and
MUC4 expression was evaluated using western blotting. aRVS reduced
MUC4 expression in the three pancreatic cancer cell lines; MUC4 was
almost eliminated following treatment with aRVS for 48 h (Fig. 4).
aRVS downregulates cell cycle
regulatory and anti-apoptotic proteins
The present study also examined the effects of aRVS
on the expression levels of various proteins associated with cell
cycle regulation and apoptosis. Treatment with aRVS reduced the
expression of the cell cycle regulatory protein cyclin D1. In
addition, the expression levels of the anti-apoptotic molecules
Bcl2 and XIAP were decreased by aRVS, whereas the expression of the
cleaved form of the proapoptotic molecule caspase-3 was increased
(Fig. 4). These results suggested
that aRVS may modulate the cell cycle and the intrinsic
mitochondrial apoptotic pathway in pancreatic cancer cells.
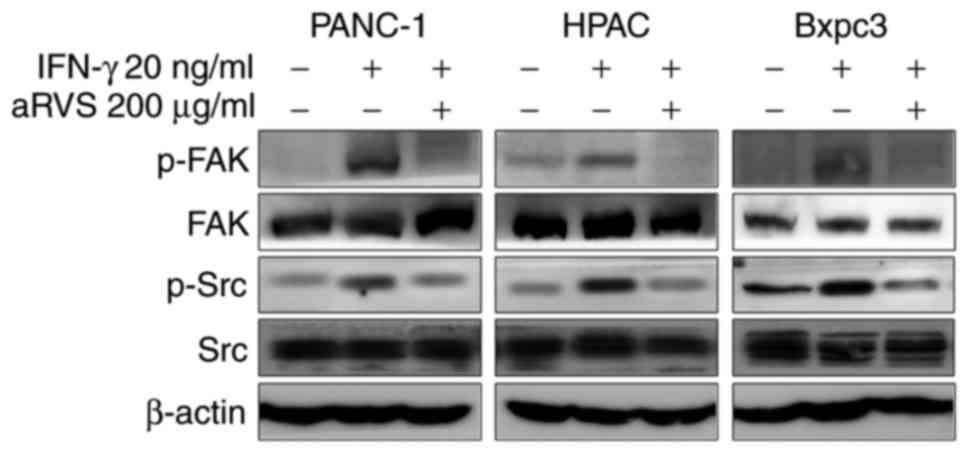
aRVS downregulates FAK and Src
signaling
The FAK protein has an important role in cellular
motility and invasion; recently, studies have been conducted
regarding the association between FAK and STAT1 (36,37).
In addition, the phosphorylation and activation of FAK by Src is
required for actin stress fiber formation, and for focal adhesion
assembly during cell adhesion and cell spreading. Therefore, the
present study aimed to determine whether aRVS was involved in the
activities of FAK and Src. The results demonstrated that FAK and
Src were activated by IFN-γ, whereas aRVS reduced the
phosphorylation of FAK and Src in PANC-1, HPAC and Bxpc3 cells
(Fig. 5).
Discussion
RVS and aRVS have been reported to induce apoptosis,
inhibit angiogenesis, and possess antioxidant and antiproliferative
activities (20,24,26,38,39).
aRVS and RVS consist of multiple constituents, including phenolic
acids (gallic acid, protocatechuic acid, etc.), flavonoids
(fisetin, sulfuretin, fustin, butein, quercetin, etc.), and other
constituents (chlorogenic acid, kaempferol-3-O-glucoside,
p-coumaric acid, etc.). Kim et al previously described the
chemical structures of the aforementioned constituents (26). In TKM, aRVS has a long history of
use due to its various efficacies and low toxicity; however, there
is still uncertainty about the specific mechanism of aRVS treatment
for the treatment of patients with cancer. Therefore, the present
study evaluated the anticancer effect of aRVS using various
pancreatic cancer cell lines. The present study indicated that aRVS
treatment affected the regulation of MUC4 and FAK expression via
the inhibition of JAK/STAT1 signaling, thus potentially reducing
invasion and metastasis. To the best of our knowledge, the present
study is the first to demonstrate that aRVS may modulate MUC4 and
identified MUC4 as a potential therapeutic target for pancreatic
cancer. Therefore, mucin expression may have an important role in
carcinogenesis progression of pancreatic cancer. This finding may
be valuable for a large number of patients with pancreatic cancer,
in whom significant overexpression of MUC4 has been detected in
pancreatic cancer compared with normal pancreatic expression
(34).
One of the notable discoveries of the present study
is the modulation of MUC4. MUC4 is a member of the membrane-binding
mucins, which is known to be overexpressed in pancreatic cancer
cells. Conversely, it is not expressed in normal pancreatic tissue,
whereas its expression is steadily increased with the stage of
disease progression and is associated with a poor prognosis
(40,41). Other studies have also reported the
oncogenic role of MUC4 and revealed that MUC4 induces the survival,
invasion and metastasis of pancreatic cancer (42,43).
MUC4 also induces epithelial-to-mesenchymal transition through the
stabilization of N-cadherin expression in pancreatic cancer cells
(44). Not only MUC4, but also MUC1
and MUC16, are overexpressed in pancreatic cancer cells and
contribute to its pathogenesis (45,46).
Considering the importance of MUC4 in the pathogenesis of
pancreatic cancer, the present study identified a downregulation
mechanism of MUC4 induced by aRVS.
Several cytokines, including interleukin (IL)-4,
IL-6, tumor necrosis factor-α and IFN-γ, have been reported to be
involved in overexpression of MUC4 via the JAK/STAT pathway,
particularly STAT1 and STAT3 (47).
Previous studies regarding the MUC4 promoter have identified the
binding sites for several transcription factors, including STAT1
and STAT3 (34,35,48).
These findings suggested that the STAT transcription factors may
have an important role in the transcriptional regulation of MUC4.
In the present study, STAT1 and STAT3 phosphorylation, and MUC4
expression were reduced in pancreatic cells treated with aRVS;
furthermore, the phosphorylation of the upstream kinases JAK1 and
JAK2 were also reduced. These results suggested that the inhibition
of MUC4 by aRVS may have an effect on STAT expression.
The induction of apoptosis and the inhibition of
cell proliferation are important mechanisms underlying the
anticancer action of numerous drugs from natural sources (49). The present study revealed that aRVS
treatment inhibited cell proliferation and induced apoptosis, via
the activation of caspase-3 and the suppression of Bcl-2, XIAP and
cyclin D1. These results indicated that aRVS may be involved in the
intrinsic apoptosis pathway. The downregulation of MUC4 has also
been revealed to induce the intrinsic pathway of apoptosis
(50), which may be a possible
mechanism by which aRVS induces apoptosis in pancreatic cancer
cells.
Cell motility is an important process in tumor
invasion and is an attractive therapeutic target for advanced
pancreatic cancer. Previous studies have reported that MUC4
modulates the mobility, morphology and actin-cytoskeleton of cancer
cells (42,51). In the present study, aRVS inhibited
the mobility and invasion of pancreatic cancer cells, and the
results suggested that the inhibitory effects of aRVS may be caused
by downregulation of MUC4. FAK is also overexpressed in invasive
tumors, and induces the invasion and metastasis of cancer cells, in
addition to re-organization of the cytoskeleton and MMPs (52). The present study revealed that aRVS
inhibited the phosphorylation of FAK and Src, without altering
total FAK and Src levels. Therefore, it may be suggested that aRVS
reduces cell mobility and invasion by regulating FAK and Src
signaling through STAT1 and MUC4.
In conclusion, the present study is the first, to
the best of our knowledge, to report that aRVS downregulated MUC4.
In addition, this study provides further evidence regarding the
molecular mechanism underlying the effects of aRVS on pancreatic
cancer. Considering the overexpression of MUC4, and its association
with chemotherapeutic resistance, in pancreatic cancer (50,53),
MUC4-targeted chemotherapy using aRVS may be a potential
therapeutic strategy. Overall, aRVS may serve an important role in
the downregulation of MUC4 and offers a potential for the
development of novel therapies for pancreatic cancer. Further
studies are required to assess the therapeutic value of aRVS in
preclinical models.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (grant no.
NRF-2016R1A6A1A03011325), and was supported by the Traditional
Korean Medicine R&D program funded by the Ministry of Health
& Welfare through the Korea Health Industry Development
Institute (KHIDI) (grant no. HB16C0067).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
SWY and BP designed the research. YK performed the
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Conroy T, Desseigne F, Ychou M, Bouché O,
Guimbaud R, Bécouarn Y, Adenis A, Raoul JL, Gourgou-Bourgade S, de
la Fouchardière C, et al: FOLFIRINOX versus gemcitabine for
metastatic pancreatic cancer. N Engl J Med. 364:1817–1825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Von Hoff DD, Ervin T, Arena FP, Chiorean
EG, Infante J, Moore M, Seay T, Tjulandin SA, Ma WW, Saleh MN, et
al: Increased survival in pancreatic cancer with nab-paclitaxel
plus gemcitabine. N Engl J Med. 369:1691–1703. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Warshaw AL and Fernández-del Castillo C:
Pancreatic carcinoma. N Engl J Med. 326:455–465. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li D, Xie K, Wolff R and Abbruzzese JL:
Pancreatic cancer. Lancet. 363:1049–1057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stark GR: How cells respond to interferons
revisited: From early history to current complexity. Cytokine
Growth Factor Rev. 18:419–423. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Darnell JE Jr, Kerr IM and Stark GR:
Jak-STAT pathways and transcriptional activation in response to
IFNs and other extracellular signaling proteins. Science.
264:1415–1421. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khodarev NN, Beckett M, Labay E, Darga T,
Roizman B and Weichselbaum RR: STAT1 is overexpressed in tumors
selected for radioresistance and confers protection from radiation
in transduced sensitive cells. Proc Natl Acad Sci USA.
101:1714–1719. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Khodarev NN, Minn AJ, Efimova EV, Darga
TE, Labay E, Beckett M, Mauceri HJ, Roizman B and Weichselbaum RR:
Signal transducer and activator of transcription 1 regulates both
cytotoxic and prosurvival functions in tumor cells. Cancer Res.
67:9214–9220. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khodarev NN, Roach P, Pitroda SP, Golden
DW, Bhayani M, Shao MY, Darga TE, Beveridge MG, Sood RF, Sutton HG,
et al: STAT1 pathway mediates amplification of metastatic potential
and resistance to therapy. PLoS One. 4:e58212009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weichselbaum RR, Ishwaran H, Yoon T,
Nuyten DS, Baker SW, Khodarev N, Su AW, Shaikh AY, Roach P, Kreike
B, et al: An interferon-related gene signature for DNA damage
resistance is a predictive marker for chemotherapy and radiation
for breast cancer. Proc Natl Acad Sci USA. 105:18490–18495. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pitroda SP, Wakim BT, Sood RF, Beveridge
MG, Beckett MA, MacDermed DM, Weichselbaum RR and Khodarev NN:
STAT1-dependent expression of energy metabolic pathways links
tumour growth and radioresistance to the Warburg effect. BMC Med.
7:682009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kharma B, Baba T, Matsumura N, Kang HS,
Hamanishi J, Murakami R, McConechy MM, Leung S, Yamaguchi K, Hosoe
Y, et al: STAT1 drives tumor progression in serous papillary
endometrial cancer. Cancer Res. 74:6519–6530. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meissl K, Macho-Maschler S, Müller M and
Strobl B: The good and the bad faces of STAT1 in solid tumours.
Cytokine. 89:12–20. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Son YO, Lee KY, Lee JC, Jang HS, Kim JG,
Jeon YM and Jang YS: Selective antiproliferative and apoptotic
effects of flavonoids purified from Rhus verniciflua Stokes
on normal versus transformed hepatic cell lines. Toxicol Lett.
155:115–125. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jang HS, Kook SH, Son YO, Kim JG, Jeon YM,
Jang YS, Choi KC, Kim J, Han SK, Lee KY, et al: Flavonoids purified
from Rhus verniciflua Stokes actively inhibit cell growth
and induce apoptosis in human osteosarcoma cells. Biochim Biophys
Acta. 1726:309–316. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee SH, Nan JX, Zhao YZ, Woo SW, Park EJ,
Kang TH, Seo GS, Kim YC and Sohn DH: The chalcone butein from Rhus
verniciflua shows antifibrogenic activity. Planta Med. 69:990–994.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Samoszuk M, Tan J and Chorn G: The
chalcone butein from Rhus verniciflua Stokes inhibits
clonogenic growth of human breast cancer cells co-cultured with
fibroblasts. BMC Complement Altern Med. 5:52005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee JC, Lim KT and Jang YS: Identification
of Rhus verniciflua Stokes compounds that exhibit free radical
scavenging and anti-apoptotic properties. Biochim Biophys Acta.
1570:181–191. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lim KT, Hu C and Kitts DD: Antioxidant
activity of a Rhus verniciflua Stokes ethanol extract. Food Chem
Toxicol. 39:229–237. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kitts DD and Lim KT: Antitumorigenic and
cytotoxic properties of an ethanol extract derived from Rhus
verniciflua Stokes (RVS). J Toxicol Environ Health A. 64:357–371.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee JC, Kim J and Jang YS: Ethanol-eluted
extract of Rhus verniciflua stokes inhibits cell growth and induces
apoptosis in human lymphoma cells. J Biochem Mol Biol. 36:337–343.
2003.PubMed/NCBI
|
|
23
|
Lim KT, Lee SJ, Heo KS and Lim K: Effects
of glycoprotein isolated from Rhus verniciflua stokes on
TPA-induced apoptosis and production of cytokines in cultured mouse
primary splenocytes. Toxicol Lett. 145:261–271. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Choi W, Jung H, Kim K and Lee S, Yoon S,
Park J, Kim S, Cheon S, Eo W and Lee S: Rhus verniciflua stokes
against advanced cancer: A perspective from the Korean Integrative
Cancer Center. J Biomed Biotechnol. 2012:8742762012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee S, Kim K, Jung H, Lee S, Cheon S, Kim
S, Eo W and Choi W: Efficacy and safety of standardized
allergen-removed Rhus verniciflua Stokes extract in patients with
advanced or metastatic pancreatic cancer: A Korean single-center
experience. Oncology. 81:312–318. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim JH, Shin YC and Ko SG: Integrating
traditional medicine into modern inflammatory diseases care:
Multitargeting by Rhus verniciflua Stokes. Mediators Inflamm.
2014:1545612014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ryu JC: Toxicity free lacquer solution and
device for producing the same. Korean Patent 10-2016-0101802. Filed
August 10, 2016; issued August 10. 2016.
|
|
28
|
Nyegaard S, Christensen B and Rasmussen
JT: An optimized method for accurate quantification of cell
migration using human small intestine cells. Metab Eng Commun.
3:76–83. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang Z, Wang Y, Farhangfar F, Zimmer M and
Zhang Y: Enhanced keratinocyte proliferation and migration in
co-culture with fibroblasts. PLoS One. 7:e409512012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Teng Y, Ross JL and Cowell JK: The
involvement of JAK-STAT3 in cell motility, invasion, and
metastasis. JAKSTAT. 3:e280862014.PubMed/NCBI
|
|
31
|
Bu LL, Deng WW, Huang CF, Liu B, Zhang WF
and Sun ZJ: Inhibition of STAT3 reduces proliferation and invasion
in salivary gland adenoid cystic carcinoma. Am J Cancer Res.
5:1751–1761. 2015.PubMed/NCBI
|
|
32
|
Barnholt KE, Kota RS, Aung HH and Rutledge
JC: Adenosine blocks IFN-gamma-induced phosphorylation of STAT1 on
serine 727 to reduce macrophage activation. J Immunol.
183:6767–6777. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kovarik P, Stoiber D, Eyers PA, Menghini
R, Neininger A, Gaestel M, Cohen P and Decker T: Stress-induced
phosphorylation of STAT1 at Ser727 requires p38 mitogen-activated
protein kinase whereas IFN-gamma uses a different signaling
pathway. Proc Natl Acad Sci USA. 96:13956–13961. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Andrianifahanana M, Singh AP, Nemos C,
Ponnusamy MP, Moniaux N, Mehta PP, Varshney GC and Batra SK:
IFN-gamma-induced expression of MUC4 in pancreatic cancer cells is
mediated by STAT-1 upregulation: A novel mechanism for IFN-gamma
response. Oncogene. 26:7251–7261. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Perrais M, Pigny P, Ducourouble MP,
Petitprez D, Porchet N, Aubert JP and Van Seuningen I:
Characterization of human mucin gene MUC4 promoter: Importance of
growth factors and proinflammatory cytokines for its regulation in
pancreatic cancer cells. J Biol Chem. 276:30923–30933. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie B, Zhao J, Kitagawa M, Durbin J, Madri
JA, Guan JL and Fu XY: Focal adhesion kinase activates Stat1 in
integrin-mediated cell migration and adhesion. J Biol Chem.
276:19512–19523. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang L and Zou W: Inhibition of integrin
β1 decreases the malignancy of ovarian cancer cells and potentiates
anticancer therapy via the FAK/STAT1 signaling pathway. Mol Med
Rep. 12:7869–7876. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Choi HS, Kim MK, Choi YK, Shin YC, Cho SG
and Ko SG: Rhus verniciflua Stokes (RVS) and butein induce
apoptosis of paclitaxel-resistant SKOV-3/PAX ovarian cancer cells
through inhibition of AKT phosphorylation. BMC Complement Altern
Med. 16:1222016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Choi HS, Seo HS, Kim SR, Choi YK, Jang BH,
Shin YC and Ko SG: Anti-inflammatory and anti-proliferative effects
of Rhus verniciflua Stokes in RAW264.7 cells. Mol Med Rep.
9:311–315. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Andrianifahanana M, Moniaux N, Schmied BM,
Ringel J, Friess H, Hollingsworth MA, Büchler MW, Aubert JP and
Batra SK: Mucin (MUC) gene expression in human pancreatic
adenocarcinoma and chronic pancreatitis: A potential role of MUC4
as a tumor marker of diagnostic significance. Clin Cancer Res.
7:4033–4040. 2001.PubMed/NCBI
|
|
41
|
Chauhan SC, Singh AP, Ruiz F, Johansson
SL, Jain M, Smith LM, Moniaux N and Batra SK: Aberrant expression
of MUC4 in ovarian carcinoma: Diagnostic significance alone and in
combination with MUC1 and MUC16 (CA125). Mod Pathol. 19:1386–1394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chaturvedi P, Singh AP, Moniaux N,
Senapati S, Chakraborty S, Meza JL and Batra SK: MUC4 mucin
potentiates pancreatic tumor cell proliferation, survival, and
invasive properties and interferes with its interaction to
extracellular matrix proteins. Mol Cancer Res. 5:309–320. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bafna S, Singh AP, Moniaux N, Eudy JD,
Meza JL and Batra SK: MUC4, a multifunctional transmembrane
glycoprotein, induces oncogenic transformation of NIH3T3 mouse
fibroblast cells. Cancer Res. 68:9231–9238. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rachagani S, Macha MA, Ponnusamy MP,
Haridas D, Kaur S, Jain M and Batra SK: MUC4 potentiates invasion
and metastasis of pancreatic cancer cells through stabilization of
fibroblast growth factor receptor 1. Carcinogenesis. 33:1953–1964.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hollingsworth MA and Swanson BJ: Mucins in
cancer: Protection and control of the cell surface. Nat Rev Cancer.
4:45–60. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Haridas D, Chakraborty S, Ponnusamy MP,
Lakshmanan I, Rachagani S, Cruz E, Kumar S, Das S, Lele SM,
Anderson JM, et al: Pathobiological implications of MUC16
expression in pancreatic cancer. PLoS One. 6:e268392011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Heinrich PC, Behrmann I, Haan S, Hermanns
HM, Müller-Newen G and Schaper F: Principles of interleukin
(IL)-6-type cytokine signalling and its regulation. Biochem J.
374:1–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kossow C, Jose D, Jaster R, Wolkenhauer O
and Rateitschak K: Mathematical modelling unravels regulatory
mechanisms of interferon-γ-induced STAT1 serine-phosphorylation and
MUC4 expression in pancreatic cancer cells. IET Syst Biol. 6:73–85.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shishodia S and Aggarwal BB: Guggulsterone
inhibits NF-kappaB and IkappaBalpha kinase activation, suppresses
expression of anti-apoptotic gene products, and enhances apoptosis.
J Biol Chem. 279:47148–47158. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bafna S, Kaur S, Momi N and Batra SK:
Pancreatic cancer cells resistance to gemcitabine: The role of MUC4
mucin. Br J Cancer. 101:1155–1161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ponnusamy MP, Singh AP, Jain M,
Chakraborty S, Moniaux N and Batra SK: MUC4 activates HER2
signalling and enhances the motility of human ovarian cancer cells.
Br J Cancer. 99:520–526. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chang YM, Kung HJ and Evans CP:
Nonreceptor tyrosine kinases in prostate cancer. Neoplasia.
9:90–100. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Skrypek N, Duchêne B, Hebbar M, Leteurtre
E, van Seuningen I and Jonckheere N: The MUC4 mucin mediates
gemcitabine resistance of human pancreatic cancer cells via the
concentrative nucleoside transporter family. Oncogene.
32:1714–1723. 2013. View Article : Google Scholar : PubMed/NCBI
|