Introduction
It has been widely acknowledged that glioblastoma
multiforme (GBM), characterized by its poor 2-year survival rate
and high mortality rate, is the most aggressive and malignant
subtype of glioma (1). Canonical
therapy for GBM includes surgical resection of tumors followed by
radiotherapy (2). However, these
therapies remain largely palliative as the majority of patients
relapse within a year of their initial operation (3). Previous studies have indicated that
the activation of cell survival pathways following ionizing
radiation (IR) contributes to the induction of recurrence (4). In addition, aberrations in tumor
suppressor genes also serve critical roles in tumor relapse
(5). Notably, the induction of
recurrence following radioresistance is commonly associated with
the activation of DNA double strand break (DSB) repair (6,7). There
exist two major pathways for DNA DSB repair, which are
nonhomologous end-joining (NHEJ) and homologous recombination (HR).
It has been demonstrated that IR-induced DSB repair is
predominantly through the NHEJ pathway, which is an intrinsically
error-prone pathway and occurs in all phases of the cell cycle
(8). A number of proteins influence
NHEJ, including the DNA-dependent protein kinase catalytic subunit,
the Ku 70/80 heterodimer, serine-protein kinase ATM(ATM), γ-histone
H2AX (γ-H2AX), and TP53-binding protein 1 (53BP1) (9–12).
Studies have demonstrated that cells deficient in the
NHEJ-associated proteins mentioned above are hypersensitive to IR
(13). Therefore, a novel
therapeutic drug targeting the NHEJ pathway may be a promising
radiosensitization approach for the treatment of glioblastoma
following IR.
Mangiferin,
1,3,6,7-tetrahydroxyxanthone-C2-β-D-glucoside, is isolated
from the leaves, stem barks, fruit peels and roots of
Mangiferina indica (14).
Known as an antioxidant, anti-diabetic and anti-inflammatory
compound (15–18), mangiferin also exhibits
anti-neoplastic effects in lung cancer (19–21),
colon cancer (20,22), leukemia (19,23–26)
and ovarian cancer (27). A
previous study demonstrated the decreased proliferation and
increased apoptosis induced by mangiferin in glioma cells via the
induction of microRNA-15b and the inhibition of matrix
metalloproteinase-9 expression (28). Further studies on mangiferin
demonstrated that it suppresses the invasiveness of glioma cells by
inhibiting the activation of the phosphatidylinositol
3-kinase/RAC-α serine/threonine-protein kinaseand mitogen-activated
protein kinase signaling pathways (29). Additionally, mangiferin enhances
recognition memory by increasing neurotrophin and cytokine levels
(30). However, to the best of our
knowledge, there are no data on whether mangiferin is able to
improve radiosensitivity in GBM cells. In the present study, it was
demonstrated that treatment with mangiferin was able to sensitize
U-87 MG and U-118MG cells to IR. As it remained unclear which
repair pathways were the most relevant targets of mangiferin in GBM
following IR, the two pathways were examined, the
radiosensitization effect of mangiferin was observed to be mediated
by inhibition of the NHEJ pathway. Taken together, these findings
demonstrated a novel function of mangiferin by inhibiting NHEJ of
DSBs generated by IR. Therefore, the present study on mangiferin
offered a potential novel strategy by which to increase the
sensitivity of glioblastoma to radiotherapy.
Materials and methods
Reagents
Mangiferin was purchased from Shanghai PureOne
Technology (Shanghai, China). The purity of mangiferin was >95%,
as demonstrated by high-performance liquid chromatography.
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS)
and 0.25% trypsin were purchased from Gibco (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). MTT (cat. no. M5655), dimethyl
sulfoxide (DMSO; cat. no. D2650), 0.25% trypsin solution (cat. no.
T4049), bovine serum albumin (BSA; cat. no. V900933), DAPI (cat.
no. D9542), paraformaldehyde (PFA; cat. no. 16005), HEPES (cat. no.
H3375), Triton X-100 (cat. no. H9284), 2 mM sodium orthovanadate
(cat. no. S6508), sodium fluoride (cat. no. S7920), 1 mM edetic
acid (cat. no. E9884), phenylmethylsulfonyl fluoride (PMSF; cat.
no. 78830), aprotinin (cat. no. A11530), leupeptin (cat. no.
L2884), penicillin and streptomycin (cat. no. V900929), and
L-glutamine (cat. no. G3126) were purchased from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). The Annexin V-fluorescein
isothiocyanate (FITC)/propidium iodide (PI) apoptosis detection kit
(cat. no. C1063) was purchased from Beyotime Institute of
Biotechnology (Haimen, China).
Cell culture, cell survival and DNA
damage assay
The human glioblastoma of unknown origin cell
lineU-87 MG (cat. no. HTB-14) and human glioblastoma cell line
U-118 MG (cat. no. HTB-15) were obtained from the American Type
Culture Collection (ATCC; Manassas, VA, USA). Short tandem repeat
(STR) DNA profiling of U-87 MG cells and U-118 MG cells was
performed using the Cell ID System (cat. no. G9500; Promega
Corporation, Madison, WI, USA), and the products were analyzed
using an ABI 3130 Genetic Analyzer (Applied Biosystems; Thermo
Fisher Scientific, Inc.). Although the U-87 MG cell line from the
ATCC is not the original cell line from the University of Uppsala
(31), and the U-118 MG and U138
lines exhibit cross contamination (32), these cell lines remain widely used
in the study of glioblastoma (33,34).
Furthermore, U-87 MG cells may be engineered with various
expression vectors. Thus, it was decided to use the U87 and U-118
MG cells from the ATCC in the present study for the in vitro
and in vivo experiments. A rat immortalized neuronal Schwann
cell line (cat. no. CRL-2765) was also purchased from the ATCC.
Cells were cultured in DMEM containing 10% FBS, 100 µg/ml
streptomycin, 100 U/ml penicillin and 0.03% L-glutamine and
maintained at 37°C with 5% CO2 in a humidified
atmosphere.
Cells in the logarithmic growth phase were seeded in
a 96-well plate (3×104 cells/well) and incubated at 37°C
for 24 h. Mangiferin (25 µg/ml) and control solvent (DMSO) were
added and incubated for 48 h (28).
The cells were irradiated with a calibrated Shepherd &
Associates Mark I self-shielded 137Cs γirradiator (JL
Shepherd & Associates Inc., San Fernando, CA, USA), at a dose
of 1.84 Gy/min for 2 min and 43 sec. As a control, mock irradiation
(0 Gy) was performed by placing the plates containing the cells in
the irradiator for the designated time period without turning on
the machine (35). For
dose-dependent and time course study, different dosage of
mangiferin (12.5, 25, 50 and 100 µg/ml) were added and incubated
for 12, 24, 36 and 48 h, respectively. A total of 0.05 mg (10 µl of
5 mg/ml) MTT was added to each well and incubated at 37°C for 4 h;
the medium was removed and termination buffer (SDS-HCl) was added
to each well. The absorbance at 570 nm was measured with a
spectrophotometer (Model 3550 Microplate Reader; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Cell viability was
calculated as follows: Cell viability (%) = [optical density (OD)
570 nm (drug)/OD 570 nm (control)] × 100. Cellular apoptosis was
measured with the Annexin V-FITC/PI apoptosis detection kit. In
brief, cells were harvested with 0.25% trypsin and washed twice
with cold PBS. A total of 1×105 cells were resuspended
in 1X binding buffer, and incubated with 5 µl FITC-conjugated
Annexin V and 5 µl PI for 15 min at room temperature in the dark.
Samples were analyzed using a FACSAria II machine (BD Biosciences,
San Jose, CA, USA). Data were analyzed with Cell Quest Pro software
(version 5.2.1; BD Biosciences).
8-Hydroxy-2′-deoxyguanosine (8-OHdG) is a modified
nucleotide base and by-product of DNA damage that is excreted upon
DNA repair (36). By measuring the
levels of 8-OHdG, DNA damage percentages were determined using a
commercial ELISA kit (cat. no. ADI-EKS-350; Enzo Life Sciences,
Inc., Farmingdale, NY, USA).
Immunofluorescent staining
U-87 MG, U-118 MG and Schwann cells were seeded into
a 6-well culture plate at a density of 4×105 cells/well
and cultured for 24 h. Following treatment with mangiferin or
control solvent, cells were exposed to 5-Gy radiation using the
Mark I 137Cs irradiator. Following irradiation,
immunofluorescent staining of γ-H2AX was performed as previously
described (35). Cells were fixed
with 4% PFA for 30 min at room temperature, and permeabilized with
1% Triton X-100. Following blocking with 5% BSA for 1 h at room
temperature, cells were incubated with rabbit polyclonal γ-H2AX
(phospho-S139) antibody (1:100 dilution; cat. no. ab11174, Abcam,
Cambridge, MA, USA) overnight at 4°C. The cells were incubated with
fluorescein isothiocyanate-conjugated goat anti-rabbit
immunoglobulin (Ig)G polyclonal antibody (1:500 dilution; cat. no.
111-095-003, Jackson ImmunoResearch Laboratories, Inc., West Grove,
PA, USA) for 2 h at room temperature. Following staining with DAPI,
cells were washed and analyzed immediately under a fluorescence
microscope (×200 magnification; Olympus Corporation, Tokyo, Japan).
At least 500 cells were counted per slide, and cells containing
>10 foci were scored as positive.
NHEJ/HR I-SceI reporter assay
In the NHEJ/HR I-SceI reporter assay, green
fluorescent protein (GFP) expression was quantified (by flow
cytometry) in U87-DRGFP cells transfected with an I-SceI plasmid as
previously described (37,38). In brief, a single DSB was generated
in the plasmid substrate pEGFP-N1 (Addgene, Inc., Cambridge, MA,
USA) containing an NHEJ or HR reporter cassette by cleavage between
the promoter and GFP reporter gene with I-SceI (cat. no.
R0694; New England BioLabs, Inc., Ipswich, MA, USA) or
HindIII (cat. no. R0104; New England BioLabs, Inc.,)
restriction enzymes. The linear products were purified using a gel
purification kit (cat. no. DP209; Tiangen Biotech Co., Ltd.,
Beijing, China). Serum-starved (overnight) U-87 MG cells were
transfected with 1 µg NHEJ reporter constructor 2 µg HR reporter
construct, and 0.1 µg pDsRed-N1 as the internal control. Cells were
harvested 48 h subsequently. The percentages of GFP-positive cells
(NHEJ or HR-repaired cells) were quantitated using a FACSAria II
machine (BD Biosciences). For each assay, 1×105 cells
were processed and data were analyzed with Cell Quest Pro software
(Version 5.2.1; BD Biosciences).
Western blot analysis
U-87 MG cells and U-118 MG cells were treated with
50 µM mangiferin or DMSO for 48 h, and the cells were exposed to
5-Gy radiation using the Mark I 137Cs irradiator.
Adherent and floating cells were collected. The cell pellets were
resuspended with lysis buffer and lysed at 4°C for 15 min. The
lysis buffer consisted of 50 mM HEPES (pH 7.4), 1% Triton X-100, 2
mM sodium orthovanadate, 100 mM sodium fluoride, 1 mM edetic acid,
1 mM PMSF, 10 mg/l aprotinin and 10 mg/l leupeptin. Following
12,000 × g centrifugation for 15 min at 4°C, the protein content of
the supernatant was determined by Bradford protein assay (cat. no.
P0006; Beyotime Institute of Biotechnology). Equal amounts of the
total protein (10 µg) were separated on 4–12% NuPAGE Bis-Tris gels
(cat. no. NP0327BOX; Life Technologies; Thermo Fisher Scientific,
Inc.) and transferred to PVDF membranes (cat. no. ISEQ00010; EMD
Millipore, Billerica, MA, USA). The membranes were soaked in
blocking buffer (5% BSA; cat. no. V900933; Sigma-Aldrich; Merck
KGaA) at room temperature for 1 h, and incubated with primary
antibodies at 4°C overnight. The following primary antibodies were
used in this study. Rabbit polyclonal 53BP1 antibody (1:1,000
dilution; cat. no. ab36823), rabbit polyclonal phospho-53BP1 (S25)
antibody (1:1,000 dilution; cat. no. ab70323), mouse monoclonal
phospho-ATM (S1981) antibody (1:1,000 dilution; cat. no. ab19304),
rabbit polyclonal γ-H2AX (phosphor-S139) antibody (1:1,000
dilution; cat. no. ab11174) were purchased from Abcam. Mouse
monoclonal β-actin antibody (1:5,000 dilution; cat. no. sc-47778)
was purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). Following incubation with specific primary antibodies, the
membranes were washed and incubated with the corresponding
secondary antibodies at room temperature for 2 h. For the secondary
antibodies, horseradish peroxidase (HRP)-conjugated goat anti-mouse
IgG polyclonal antibody (1:5,000 dilution; cat. no. 115-035-003)
and HRP-conjugated goat anti-rabbit polyclonal IgG (1:5,000
dilution; cat. no. 111-035-003) were purchased from Jackson
ImmunoResearch Laboratories, Inc. The membranes were washed and
visualized via enhanced chemiluminescence (cat. no. 345818; EMD
Millipore).
Tumor xenograft study
To establish tumor xenografts, 5×106
U-118 MG cells were injected into BALB/c nude male mice (5–6 weeks
old; 16–18 g body weight; Affiliated Laboratory Animal Center of
Sichuan Academy of Medical Science and Sichuan Provincial People's
Hospital, Chengdu, China). Following implantation, the tumors were
allowed to grow to a size of 100–550 mm3. No mouse
bearing multiple tumors was identified in the present study.
Furthermore, the largest diameter exhibited by a single
subcutaneous tumor was 1.1 cm. The mice were randomly divided into
4 groups (n=80 in total and n=20 per group) as follows: i) The
control group, in which mice with U-118 MG-derived tumors were left
untreated; ii) the IR group, in which mice received 25-Gy IR
following tumor formation by U-118 MG cells; iii) the mangiferin
group, in which mice were intraperitoneally administered with 5
mg/kg mangiferin following tumor formation by U-118 MG cells; and
iv) the IR+mangiferin group, in which mice were intraperitoneally
administered with 5 mg/kg mangiferin and subjected to 25-Gy IR
following tumor formation by U-118 MG cells. Following treatment,
mouse body weights were measured every day. On day 20, 10 mice in
each group were sacrificed, and 10 mice in each group were used for
survival analysis. The subcutaneous tumors were removed from the
sacrificed mice and weighed. Meanwhile, the volume of the tumors
was determined in three dimensions with Vernier calipers, according
to the following formula: Tvol = length × width × depth × 0.5.
Animal handling was approved by the Ethics Committee of Sichuan
Academy of Medical Science and Sichuan Provincial People's
Hospital, and all animals were kept in a 12 h light/dark cycle with
free access to water and food (26°C and 40–60% humidity), which is
in accordance with individual ventilated cage requirements at the
Sichuan Academy of Medical Science and Sichuan Provincial People's
Hospital.
Statistical analysis of the data
The experiments were repeated three times, and all
data are expressed as the mean ± standard error of the mean from at
least three independent experiments. Data analysis was performed
using GraphPad Prism 5.0 software (GraphPad Software, Inc., La
Jolla, CA, USA). Statistical significance between multiple groups
was determined by one- or two-way analysis of variance with a
Bonferroni post hoc test, and between two groups using a Student's
t-test. Survival analyses were carried out using Kaplan-Meier
curves. P<0.05 was considered to indicate a statistically
significant difference.
Results
Mangiferin inhibits cell viability
following IR
Based on a previous study on mangiferin (21,27),
it was hypothesized that mangiferin may increase the sensitivity of
glioblastoma cells to radiation. Therefore, to substantiate this
hypothesis and to examine the inhibitory role of mangiferin in GBM
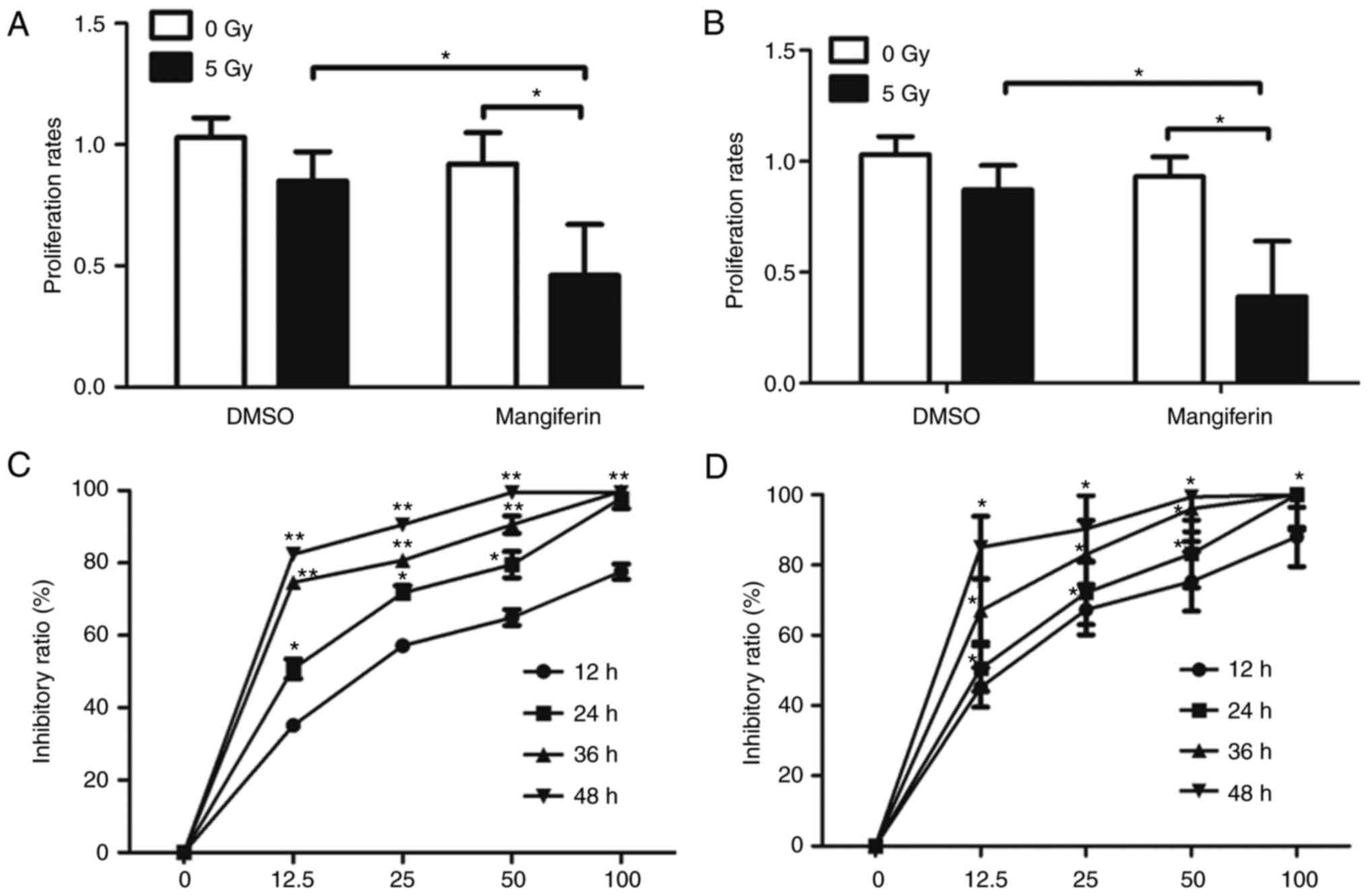
cells following IR, MTT assays were performed on U-87 MG (Fig. 1A) and U-118 MG cells (Fig. 1B). As predicted, the proliferation
rates of U-87 MG cells treated with 25 µg/ml mangiferin
following5-Gy IR were decreased significantly compared with those
of the mock-treated cells (Fig.
1A), indicating that mangiferin may be a potential
radiosensitive agent in the treatment of GBM following IR. Although
the U87 cell line from ATCC is not the original cell line from the
University of Uppsala (31), the
cells remain widely used in the study of glioblastoma (33,34).
To further ascertain the potential inhibitory role of mangiferin
following IR, the radiosensitizing potential of mangiferin in U-118
MG cells was also determined. As presented in Fig. 1B, the proliferation rates of U-118
MG cells were greatly inhibited following combined treatment with
mangiferin and IR. To further assess whether mangiferin was able to
radiosensitize GBM cells by inhibiting DNA repair, a radiation
survival assay was performed to assess this in a dose and
time-dependent manner. Consistent with the proliferation data
(Fig. 1A and B), with 5 Gy
radiation, U-87 MG cells (Fig. 1C)
and U-118 MG cells (Fig. 1D)
exhibited marked sensitivity to radiation following treatment with
mangiferin. Notably, this radiosensitization was dose-dependent,
meaning that a lesser degree of sensitization was observed in cells
treated with lower concentrations of mangiferin. Furthermore, this
radiosensitization was also time-dependent. On the basis of these
experiments, a concentration of 25 µg/ml mangiferin was used for
the subsequent experiments. Taken together, these results suggested
that mangiferin may enhance the radiosensitivity of glioblastoma
cells to radiation, and therefore inhibit the viability of cells
following radiation.
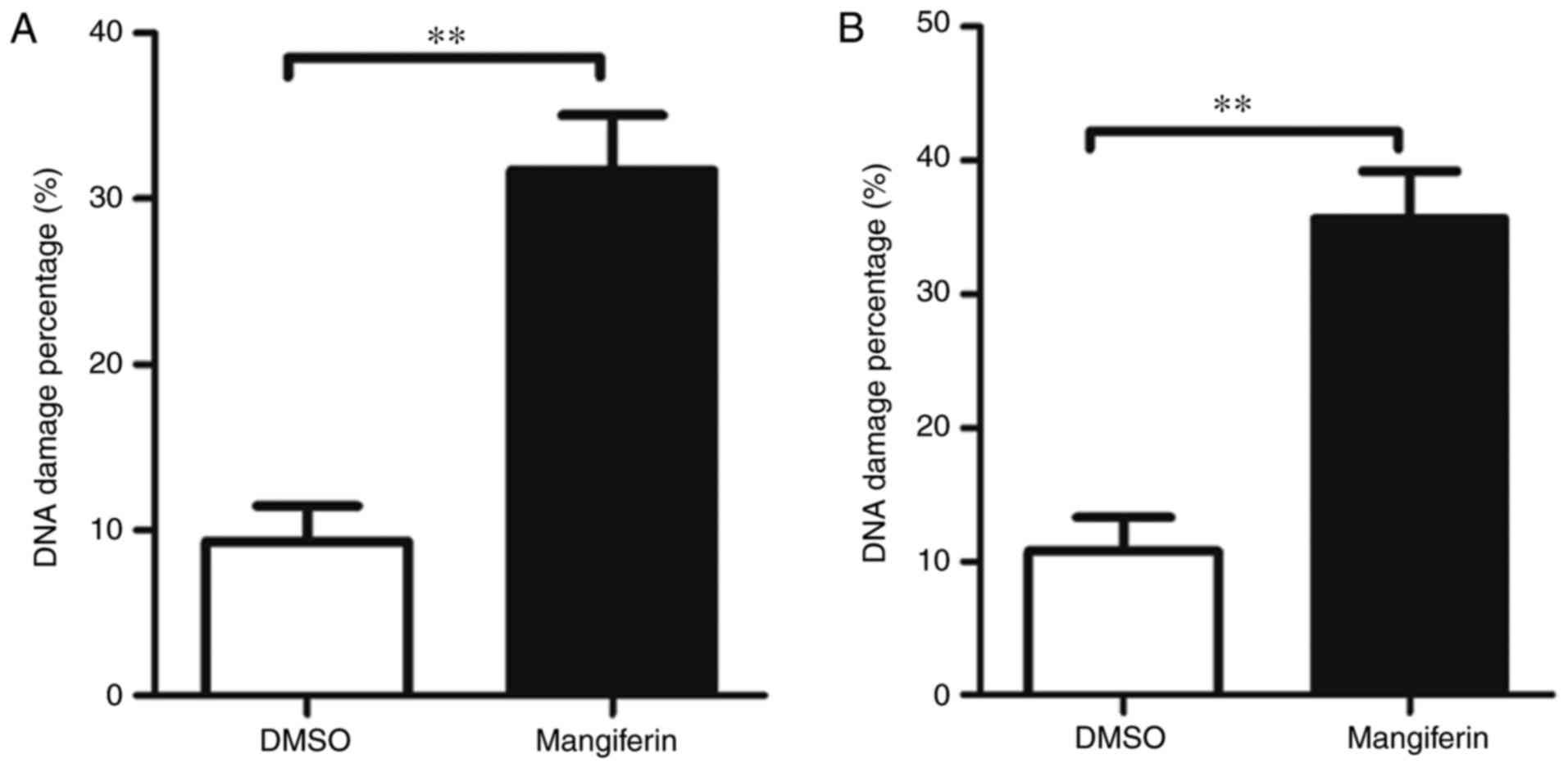
Mangiferin inhibits DNA damage repair
via the NHEJ pathway
To examine the mechanism of mangiferin-induced
radiosensitivity in glioblastoma cells, the present study examined
whether mangiferin was able to increase the DNA damage percentages
in GBM cells following IR. As indicated by Fig. 2A (U-87 MG cells) and Fig. 2B (U-118 MG cells),
mangiferin-treated GBM cells exhibited increased numbers of DNA
damage percentages, which correlated with the high degree of
radiosensitization observed in Fig.
1. These observations indicated that mangiferin possibly
mediated radiosensitization through the inhibition of DNA damage
repair.
Since in eukaryotic cells there exist two major DSB
repair pathways following radiation, one of which is the fast and
efficient, although error-prone, NHEJ DNA repair pathway. The other
is error-free HR, which is considered to be a more accurate
mechanism for DSB repair as it uses homologous sequences for repair
synthesis. However, as there is competition and crosstalk between
these two repair pathways (39),
the present study subsequently investigated by which pathway
mangiferin inhibited DSB repair and thus induced the limited
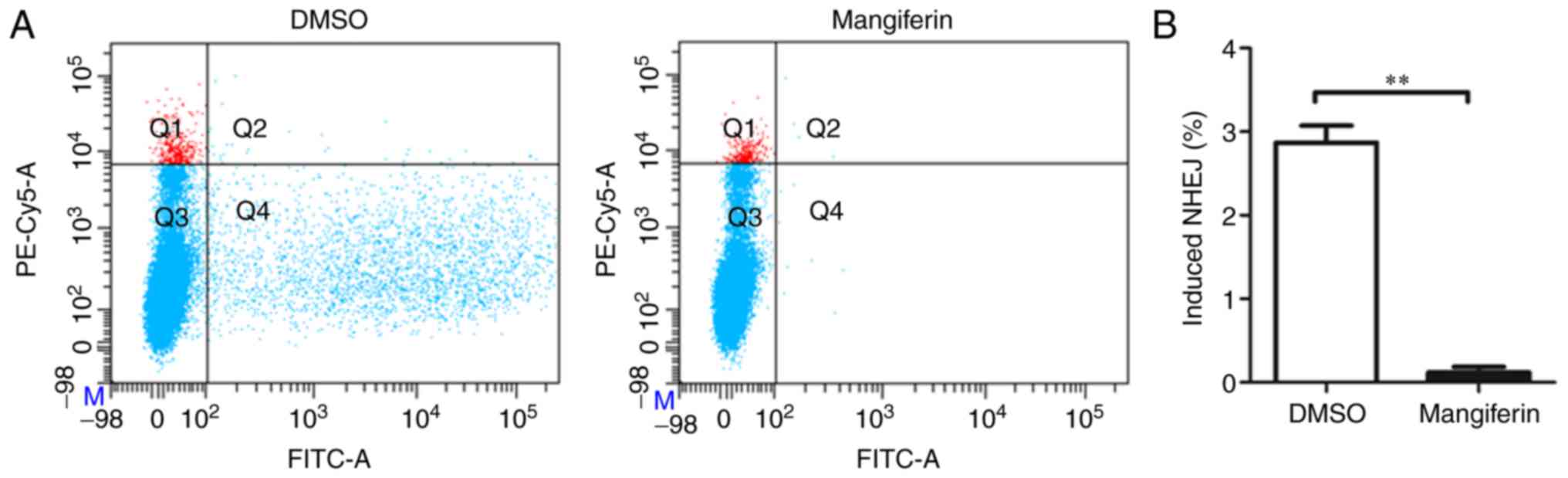
proliferation following radiation. Notably, treatment with
mangiferin resulted in NHEJ repair defects in cells that were more
severe than those in the control solvent cells (Fig. 3A). Statistical analysis of NHEJ
percentages further confirmed this (Fig. 3B). Therefore, NHEJ repair was
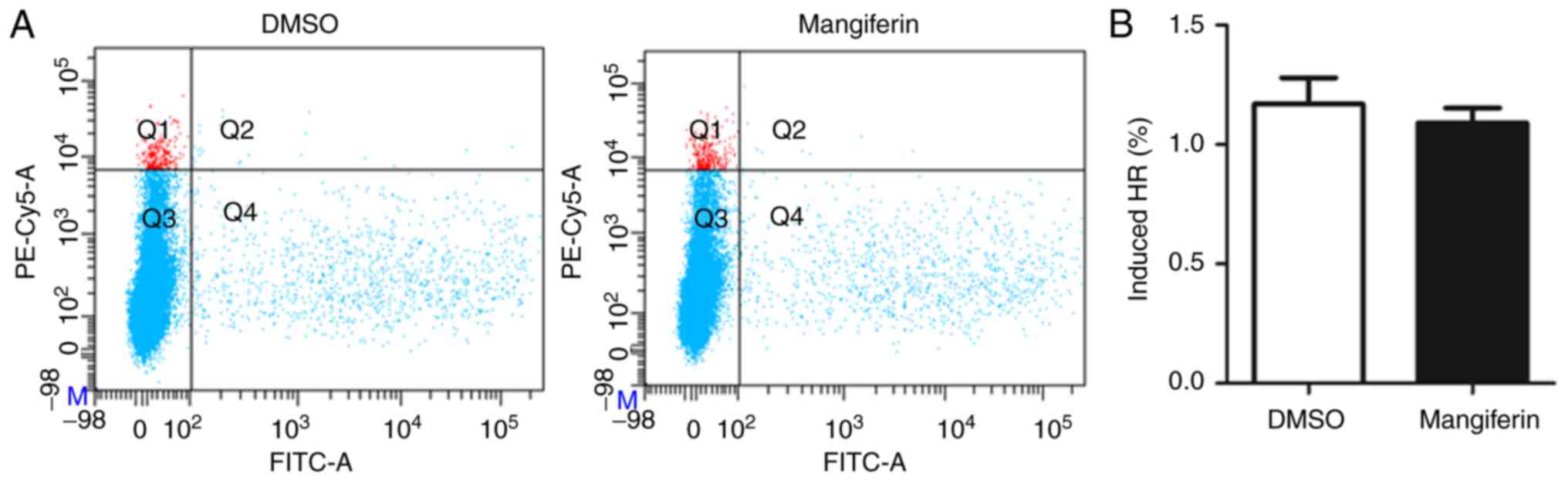
significantly inhibited by mangiferin. In contrast to the
inhibitory effects of mangiferin on NHEJ repair, pretreatment of
cells with mangiferin led to decreased HR repair percentages
(Fig. 4A), although this decrease
was not statistically significant (P>0.05; Fig. 4B).
To further determine the mechanisms of
mangiferin-mediated inhibition of NHEJ repair in GBM cells, the
present study investigated the phosphorylation levels of ATM, 53BP1
and γ-H2AX. It was identified that mangiferin attenuated the
activation of ATM, 53BP1 andγ-H2AX in U-87 MG cells. Similarly, the
phosphorylation of ATM, 53BP1 andγ-H2AX was also markedly impaired
by pretreatment with mangiferin in U-118 MG cells (Fig. 5). Collectively, these data suggested
that the radiosensitization effect of mangiferin was due to its
inhibition of the NHEJ repair pathway.
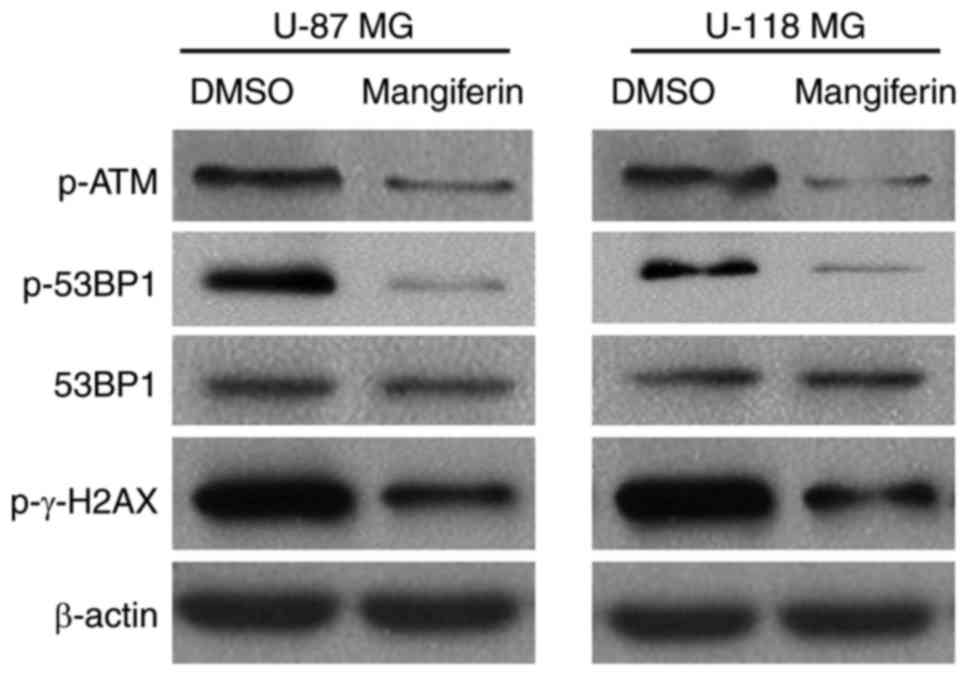 | Figure 5.Mangiferin inhibits key proteins in
non-homologous end-joining repair. Western blotting of phospho-ATM,
phospho-53BP1, 53BP1, phospho-γ-H2AX and β-actin. Left, U-87 MG
cells; right, U-118 MG cells. Images are representative of three
independent experiments. ATM, serine-protein kinase ATM; 53BP1,
TP53-binding protein 1; γ-H2AX, γ-histone H2AX; p, phosphorylated;
DMSO, dimethyl sulfoxide. |
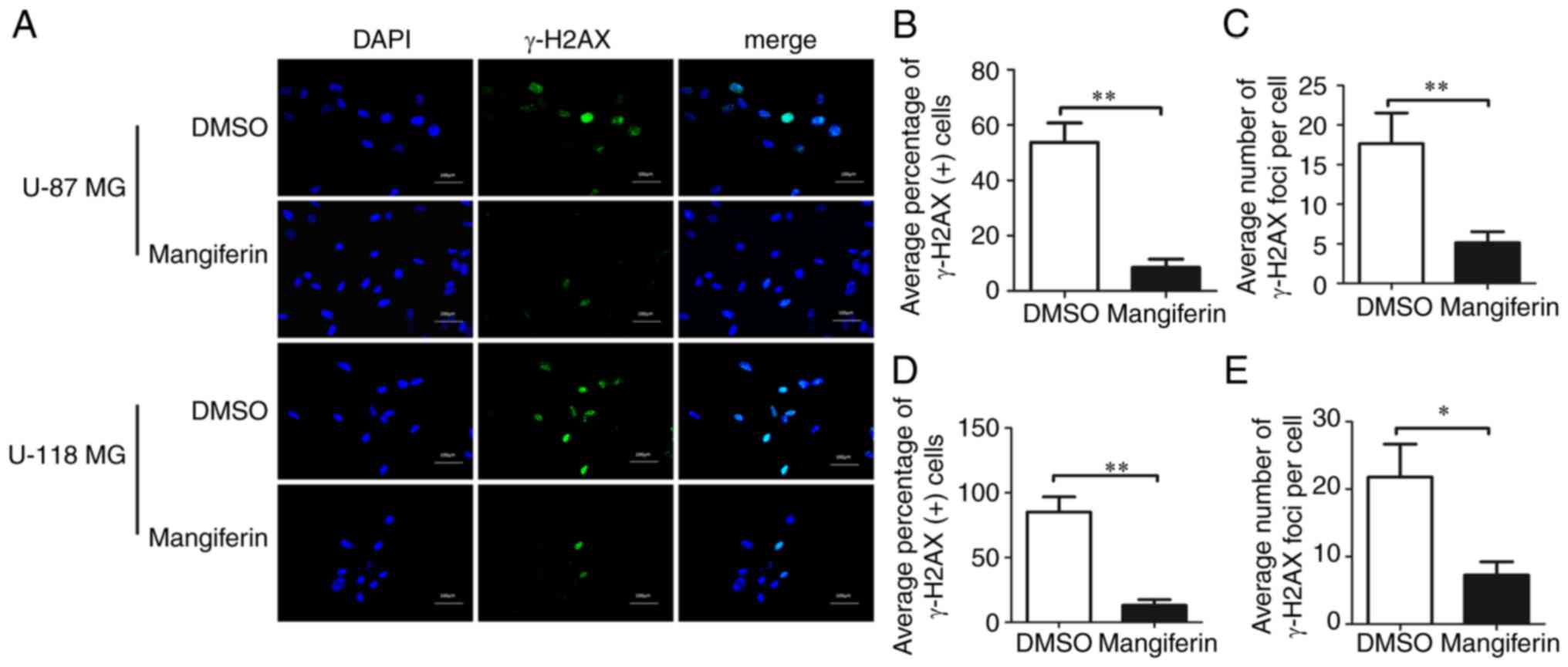
Mangiferin decreases the number of
γ-H2AX foci
To validate the findings on mangiferin-mediated
inhibition of NHEJ repair, immunocytochemistry staining of U-87 MG
and U-118 MG cells and assessed the repair percentages by
investigating the number of γ-H2AX foci (Fig. 6). Immunocytochemistry analysis
revealed that pretreatment with mangiferin decreased the
percentages ofγ-H2AX foci (Fig.
6A). Notably, statistical analysis of the average percentages
of γ-H2AX-positive U-87 MG cells (Fig.
6B) and U-118 MG cells (Fig.
6D) clearly indicated that pretreatment with mangiferin
decreased the number of γ-H2AX foci in cells. Similar to the data
onγ-H2AX-positive cell percentages, the average numbers of γ-H2AX
foci per cell were significantly decreased in U-87 MG cells
(Fig. 6C) and U-118 MG cells
(Fig. 6E). These results clearly
indicated that mangiferin was able to potently inhibit NHEJ,
resulting in DSB repair defects by decreasing the number of γ-H2AX
foci in GBM cells.
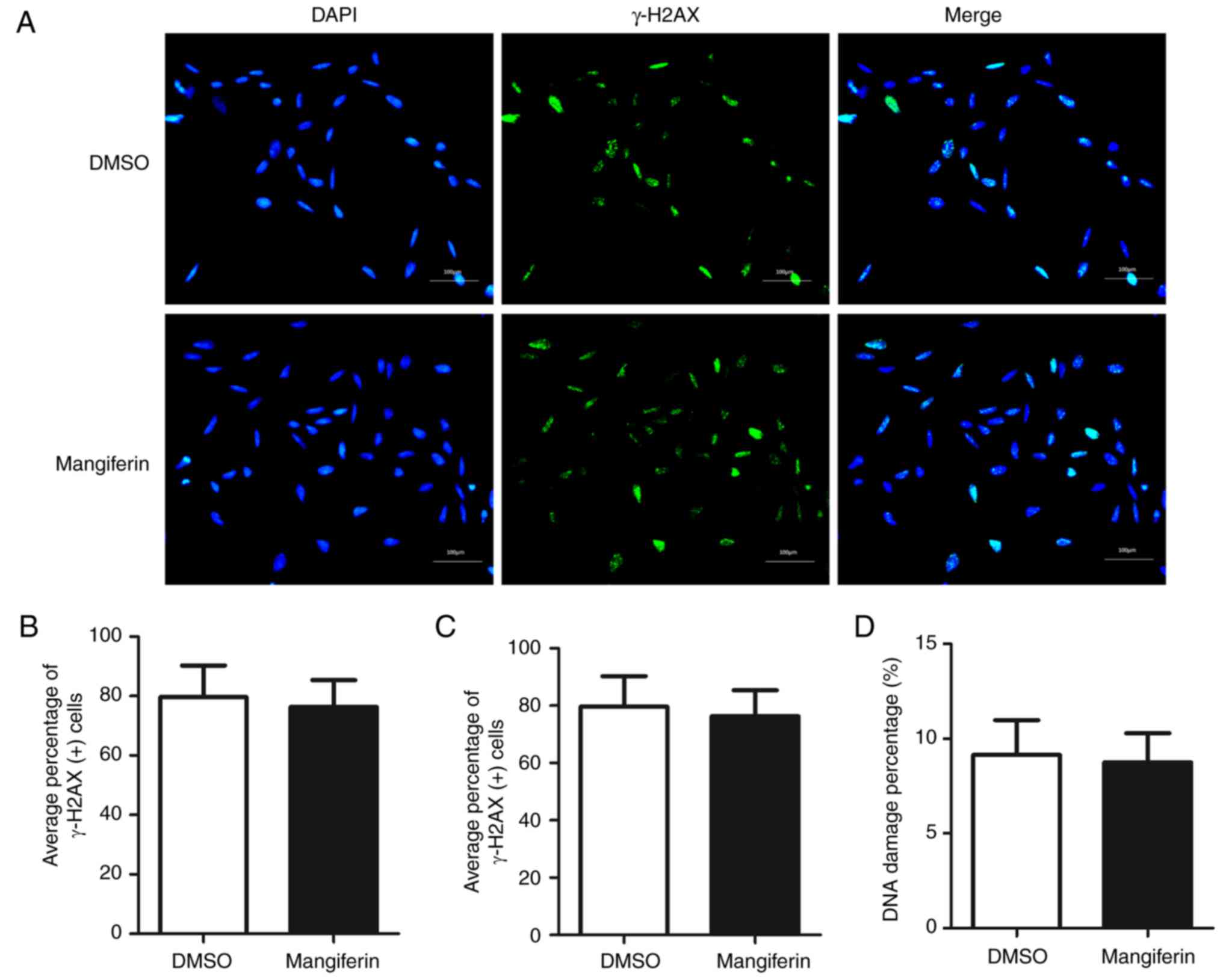
Mangiferin does not inhibit DSB repair
in Schwann cells
To investigate whether mangiferin was able to induce
the inhibition of DSB repair in neuronal cells, DSB repair was
examined in rat immortalized neuronal Schwann cells. In Fig. 7A, cells pretreated with mangiferin
and control solvent exhibited comparatively similar numbers of
γ-H2AX foci. Statistical analysis of the average percentages of
γ-H2AX-positive cells (Fig. 7B) and
average numbers of γ-H2AX foci per cell (Fig. 7C) clearly demonstrated that
pretreatment with mangiferin was not able to attenuate DSB repair
in Schwann cells. Further analysis of the DNA damage percentages
also indicated that pretreatment with mangiferin was not able to
inhibit DSB repair in rat neuronal Schwann cells. These data
suggested that mangiferin selectively inhibited DSB repair in GBM
cells, and was unable to trigger inhibition in normal neuronal
cells.
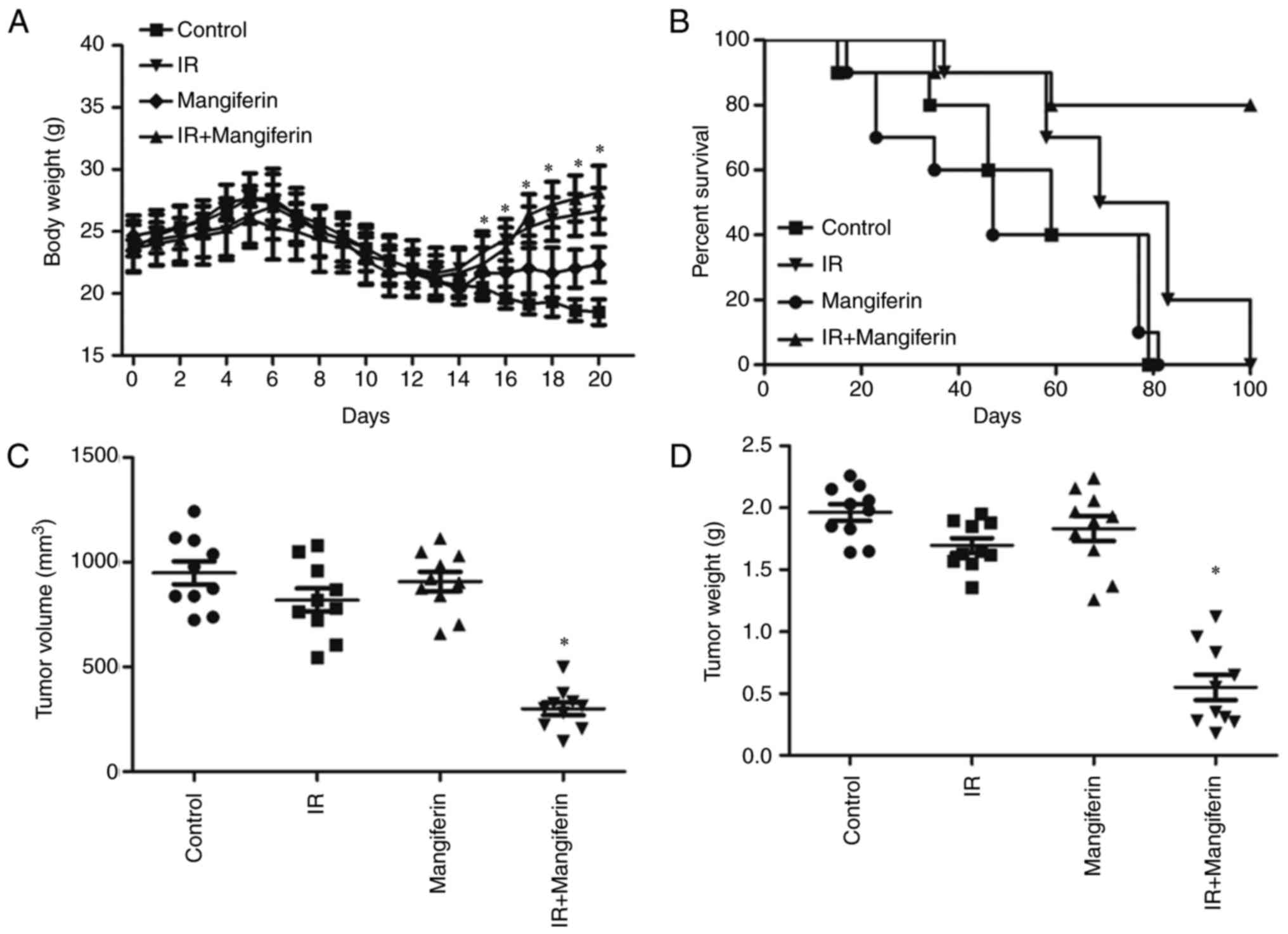
Mangiferin increases radiosensitivity
in vivo
Finally, to determine the effect of mangiferin on
radiosensitivity in xenograft tumors, subcutaneous tumors were
generated in nude mice using U-118 MG cells (Fig. 8). As presented in Fig. 8A, following treatment with IR and
mangiferin, tumor-bearing mice exhibited gradually increased body
weights. Subsequently, tumors from mangiferin and IR co-treated
mice exhibited marked reductions in the tumor volume (Fig. 8C) and the tumor weight (Fig. 8D), demonstrating that treatment with
mangiferin and IR was able to inhibit tumor growth compared with IR
alone, mangiferin alone and the normal saline control. Furthermore,
the survival curve of xenograft mice clearly indicated that the
irradiated mice pretreated with mangiferin had a comparatively
prolonged life span (Fig. 8B).
Taken together, these results demonstrated that pretreatment with
mangiferin increased radiosensitivity in glioblastoma, in
vitro and in vivo.
Discussion
With a poor 2-year survival rate, GBM is the most
aggressive primary brain tumor in adults and the leading cause of
mortality (40). Standard therapy
for GBM includes surgical resection followed by radiotherapy and
chemotherapy. However, GBM relapse rates remain high, likely due to
the activation of DNA repair systems in cancer cells. Therefore,
previous studies have focused on the improvement of
radiosensitivity to IR by blocking DSB repair pathways (41–43).
In the current study, markedly inhibited proliferation was observed
with combined therapy of radiation and pretreatment with
mangiferin, suggesting that mangiferin may be a potential
radiosensitive agent. Furthermore, the improved sensitivity was
dose- and time-dependent. Based on these observations, it was
hypothesized that the improved radio sensitivity of GBM cells by
pretreatment with mangiferin was mediate via the inhibition of DSB
repair. Through analysis of DNA damage percentages, NHEJ and HR
percentages, and γ-H2AX foci numbers, the hypothesis that
mangiferin inhibited DNA repair by inhibiting the NHEJ pathway was
validated. More importantly, experiments on rat immortalized
neuronal Schwann cells revealed that the DSB repair-blocking effect
of mangiferin was only triggered in tumor cells. Furthermore,
tumor-bearing mouse data indicated that mangiferin was able to
increase the sensitivity of the xenograft tumors to IR and maybe a
potential therapeutic drug for the treatment of GBM.
Previous studies on mangiferin revealed that
mangiferin may exert anticancer effects by inhibiting the Notch
signaling pathway (27) and
transcriptional coactivator YAP1 signaling pathway (He et
al, unpublished data) in ovarian cancer. A study on non-small
cell lung cancer A549 cells also revealed the anti-neoplastic
effects of mangiferin, mediated by inducing apoptosis and cell
cycle arrest at the G2/M phase (21). Other studies have revealed that
mangiferin mediates apoptosis via the activation of nuclear
factor-κB by inducing tumor necrosis factor expression (44). Furthermore, in lung cancer-bearing
animals, the activity of glutathione transferase, quinone reductase
and uridine 5′-diphosphate-glucuronosyl transferase is enhanced by
mangiferin (45). In addition, in
K562 cells, mangiferin inhibits telomerase activity and induces
apoptosis (23,24). Therefore, mangiferin has been
reported to possess anti-neoplastic functions in lung (19,20)
and colon cancer (20,22), leukemia (19,23–26),
lung (19,26) and prostate cancer (19). However, there exists controversy as
to whether mangiferin is able to inhibit the proliferation of GBM
cells. According to a study by Pardo Andreu et al (30), mangiferin stimulates the
proliferation of human U-138 MG glioblastoma cells. However, a
study by Xiao et al (28)
demonstrated the increased apoptosis of U-87 MG cells mediated by
mangiferin. Furthermore, doubts have been raised about U-87 MG
cells originating from the ATCC due to potential contamination
(31). Therefore, in the present
study, U-87 MG s and U-118 MG cells were used. With mock radiation,
slightly inhibited proliferation of mangiferin-treated cells was
observed, although this was not statistically significant. This was
primarily due to the low concentration used in the present study.
In accordance with the study of Xiao et al (28), with the increased concentration of
mangiferin, increased apoptosis was also observed in U-87 MG and
U-118 MG cells (data not shown). As the primary purpose of the
present study was to examine whether mangiferin was able to
increase the radiosensitivity of GBM cells, a dosage of 25 µg/ml
mangiferin was selected for the present study. With the time course
and dose-dependence experiment, the radiosensitive role of
mangiferin was observed to be time-and dose-dependent. Further
analysis of DNA damage percentages demonstrated that
mangiferin-treated GBM cells had more damaged DNA. As there exist
two principal pathways of DNA damage repair, the mechanisms
underlying mangiferin-inhibited DNA repair required further
study.
It has been widely postulated that two principal
pathways, NHEJ and HR, are involved in the DSB repair in eukaryotic
cells. To facilitate efficient repair, these two principal repair
pathways cooperate and compete with each other at DSB sites
(46). Although NHEJ appears to
compete with HR for DSB, key proteins, including ATM and 53BP1,
have been reported to influence HR through a complex regulatory
network (39). 53BP1 is a highly
conserved DNA damage checkpoint protein, and an important regulator
of genome stability by mediating DSB repair (47). A previous study demonstrated that
53BP1-null mice are hypersensitive to radiation due to defects in
NHEJ (48). Therefore, to examine
the radiosensitive role of mangiferin, the phospho-53BP1 expression
levels were examined, and decreased activation of 53BP1 mediated by
pretreatment with mangiferin was observed. Furthermore,
accumulating evidence suggests that ATM serves a critical role in
regulating the cellular response to IR (49). A previous study reported that
ATM-dependent phosphorylation was a prerequisite for the activation
of 53BP1 at DSB sites following IR (12). Therefore, in the present study, it
was observed that there was decreased activation of ATM and 53BP1
following treatment with mangiferin and radiation, indicating that
mangiferin inhibited DSB repair. In the current study, in order to
ensure equal loading, 53BP1 and β-actin were used as loading
controls, and their protein expression was quantified. Furthermore,
as previously reported, the activation of ATM in DSB repair also
requires the phosphorylation of γ-H2AX and 53BP1 (50). It was observed that there was
decreased phosphorylation of γ-H2AX in U-87 MG and U-118 MG cells
treated with mangiferin. The formation of γ-H2AX foci was observed
in GBM cells treated with either mangiferin or control solvent, and
it was identified that mangiferin affected γ-H2AX phosphorylation
and subcellular localization. It may be possible that mangiferin
has a phosphatase inhibitor role and directly inhibits the
phosphorylation of ATM. It was reported that the SQ/TQ domain is
the principal ATM kinase phosphorylation region (51). If mangiferin directly inhibits ATM
phosphorylation, it may be possible that mangiferin binds to the
SQ/TQ domain and that this binding inhibits ATM phosphorylation.
There also exists the possibility that mangiferin indirectly
inhibits the phosphorylation of ATM by the inhibition of upstream
genes of ATM. Thus, further studies examining the molecular
mechanism of mangiferin-inhibited ATM phosphorylation are required.
It has been reported that ATM phosphorylates a broad range of
substrates, including 53BP1 and γ-H2AX (52), thus it may be possible that
mangiferin phosphorylates ATM at the SQ/TQ domain, and the p-ATM
further phosphorylated 53BP1 and γ-H2AX. Therefore, mangiferin may
also directly phosphorylate 53BP1 at the SQ/TQ domain. In order to
unravel the molecular mechanisms of the mangiferin-induced
phosphorylation of ATM, 53BP1 and γ-H2AX, further experiments are
required. To confirm the role of NHEJ repair in mangiferin-mediated
inhibition of DSB repair following radiation, an in vivo
assay system was generated, measuring NHEJ repair using a
linearized plasmid (53). By using
the NHEJ-I SceI assay system, a markedly decreased NHEJ percentage
was observed in U-87 MG cells treated with mangiferin. To evaluate
whether the inhibition of DSB repair by mangiferin was mediated
only by the inhibition of NHEJ, an HR-I SceI assay system was also
generated, and a significant difference in HR repair percentages
between mangiferin-treated and DMSO-treated cells was not observed.
The present data supported an inhibitory role of mangiferin in NHEJ
repair following radiation. The present study subsequently assessed
whether mangiferin was able to mediate neuronal protection from IR.
Data from rat immortalized Schwann cells revealed that treatment
with mangiferin did not result in any alteration in DSB repair.
Prior to this, to the best of our knowledge, there had been no
direct evidence of the mechanisms underlying the different effects
of mangiferin on radiosensitivity between Schwann and GBM cells. A
previous study indicated that 5–25 µg/ml mangiferin protected
against γ-radiation-induced DNA damage in human lymphocytes
(40). It has also been reported
that mangiferin reduces etoposide-induced DNA damage in human
umbilical cord blood cells, by inducing the nuclear accumulation of
nuclear factor erythroid 2-related factor 2 (Nrf2) and increasing
the expression of NAD(P)H dehydrogenase [quinone] 1, a downstream
target gene of the Nrf2 pathway (41). Furthermore, the in vitro
experiments by Lei et al (54) demonstrated that pretreatment with
either mangiferin aglycone or mangiferin was able to inhibit DNA
damage by IR in human intestinal epithelial cells. Therefore, it
was hypothesized that for normal cells, which are less
proliferative compared with tumor cells, mangiferin serves a DNA
damage-protective role via its antioxidant function. However, for
proliferative tumor cells, mangiferin may increase the
radiosensitivity. Another reason may be the crosstalk between
metabolism and DNA damage repair. According to the Warburg effect,
the majority of cancer cells predominantly produce their energy
through aerobic glycolysis; whereas, normal cells primarily produce
energy through mitochondrial oxidative phosphorylation (43). Pyruvate kinase PKM (PKM2) has been
reported to be the key enzyme in the Warburg effect in tumor cells,
and it is not expressed in healthy tissues (44). However, PKM2 also interacts with DNA
damage-associated proteins and regulates the DSB repair pathway
(45,46). Recent studies revealed a modulating
function of mangiferin on the metabolism of carbohydrates and
lipids (47). Therefore, it was
postulated that by regulating tumor metabolism enzyme PKM2, which
exhibits crosstalk with DNA repair pathways, mangiferin may have
different DNA damage repair functions in tumor cells and normal
cells. However, these assumptions require further examination in
future studies. Therefore, mangiferin may possess dual roles by
increasing the radiosensitivity of glioblastoma cells and enhancing
neuronal protection for normal neuronal cells.
Subsequently, the present study examined the in
vivo radiosensitivity of mangiferin in tumor-bearing nude mice.
Although complete tumor regression did not occur in all mice, mice
treated with mangiferin exhibited significantly smaller tumors with
lower tumor weights. As the primary purpose of this study was to
examine whether mangiferin was able to prolong the life span of
xenograft mice, the survival study was of critical importance. As
predicted, mice treated with mangiferin exhibited increased
survival following radiation, compared with mice treated with
radiation alone. Therefore, mangiferin increased the sensitivity of
glioblastoma cells to IR in vivo. Furthermore, no adverse
effects were observed, including diarrhea or symptoms of decreased
food intake. It has previously been reported that there is no
clinical evidence of adverse effects of mangiferin (55). In summary, the present data
demonstrated that the radioresistance of GBM cells may be reversed
by mangiferin. Therefore, with the marked radiosensitization of GBM
cells at low concentrations and neuroprotection of neuronal cells
during cranial IR, mangiferin has potential as a novel drug for the
treatment of human glioblastoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Sichuan
Health and Family Planning Commission Funding (grant no. 16ZD0253),
the Sichuan Science and Technology Funding (grant no. 2018JY0645),
the Funding from Sichuan Academy of Medical Science & Sichuan
Provincial People's Hospital, and Sichuan Scientific Research
Foundation of the Returned Overseas Chinese Scholars and the
National Science Funding of China (no. 81802504) for YW. The
present study was also supported by Sichuan National Science
Research Funding (grant no. 2014FZ0126) for RT. The present study
was also supported by the National Key Specialty Construction
Project of Clinical Pharmacy (grant no. 30305030698).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW, SD and RT designed the study. FM, TLi and HZ
performed the molecular and cell experiments. XX performed the
animal experiments. TLe generated the plasmids. XH performed the
statistical analysis. FM and SD were principal contributors to the
production of the figures and writing the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Animal handling was approved by the Ethics Committee
of Sichuan Academy of Medical Science and Sichuan Provincial
People's Hospital (Chengdu, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Killock D: CNS cancer: Molecular
classification of glioma. Nat Rev Clin Oncol. 12:5022015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Walker MD, Alexander E Jr, Hunt WE,
MacCarty CS, Mahaley MS Jr, Mealey J Jr, Norrell HA, Owens G,
Ransohoff J, Wilson CB, et al: Evaluation of BCNU and/or
radiotherapy in the treatment of anaplastic gliomas. A cooperative
clinical trial. J Neurosurg. 49:333–343. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Debus J and Abdollahi A: For the next
trick: New discoveries in radiobiology applied to glioblastoma. Am
Soc Clin Oncol Educ Book. e95–e99. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fan QW and Weiss WA: Targeting the
RTK-PI3K-mTOR axis in malignant glioma: Overcoming resistance. Curr
Top Microbiol Immunol. 347:279–296. 2010.PubMed/NCBI
|
|
5
|
El-Deiry WS: The role of p53 in
chemosensitivity and radiosensitivity. Oncogene. 22:7486–7495.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bao S, Wu Q, McLendon RE, Hao Y, Shi Q,
Hjelmeland AB, Dewhirst MW, Bigner DD and Rich JN: Glioma stem
cells promote radioresistance by preferential activation of the DNA
damage response. Nature. 444:756–760. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
King HO, Brend T, Payne HL, Wright A, Ward
TA, Patel K, Egnuni T, Stead LF, Patel A, Wurdak H, et al: RAD51 is
a selective DNA repair target to radiosensitize glioma stem cells.
Stem Cell Reports. 8:125–139. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
van Gent DC, Hoeijmakers JH and Kanaar R:
Chromosomal stability and the DNA double-stranded break connection.
Nat Rev Genet. 2:196–206. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dobbs TA, Tainer JA and Lees-Miller SP: A
structural model for regulation of NHEJ by DNA-PKcs
autophosphorylation. DNA Repair. 9:1307–1314. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang H, Perrault AR, Takeda Y, Qin W, Wang
H and Iliakis G: Biochemical evidence for Ku-independent backup
pathways of NHEJ. Nucleic Acids Res. 31:5377–5388. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dimitrova N, Chen YC, Spector DL and de
Lange T: 53BP1 promotes non-homologous end joining of telomeres by
increasing chromatin mobility. Nature. 456:524–528. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
DiTullio RA Jr, Mochan TA, Venere M,
Bartkova J, Sehested M, Bartek J and Halazonetis TD: 53BP1
functions in an ATM-dependent checkpoint pathway that is
constitutively activated in human cancer. Nat Cell Biol.
4:998–1002. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lieber MR, Ma Y, Pannicke U and Schwarz K:
Mechanism and regulation of human non-homologous DNA end-joining.
Nat Rev Mol Cell Biol. 4:712–720. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sánchez GM, Re L, Giuliani A, Núñez-Sellés
AJ, Davison GP and León-Fernández OS: Protective effects of
Mangifera indica L. extract, mangiferin and selected antioxidants
against TPA-induced biomolecules oxidation and peritoneal
macrophage activation in mice. Pharmacol Res. 42:565–573. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guha S, Ghosal S and Chattopadhyay U:
Antitumor, immunomodulatory and anti-HIV effect of mangiferin, a
naturally occurring glucosylxanthone. Chemotherapy. 42:443–451.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Duang XY, Wang Q, Zhou XD and Huang DM:
Mangiferin: A possible strategy for periodontal disease to therapy.
Med Hypotheses. 76:486–488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang HL, Li CY, Zhang B, Liu YD, Lu BM,
Shi Z, An N, Zhao LK, Zhang JJ, Bao JK, et al: Mangiferin
facilitates islet regeneration and β-cell proliferation through
upregulation of cell cycle and β-cell regeneration regulators. Int
J Mol Sci. 15:9016–9035. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang H, He X, Lei T, Liu Y, Huai G, Sun M,
Deng S, Yang H, Tong R and Wang Y: Mangiferin induces islet
regeneration in aged mice through regulating p16INK4a.
Int J Mol Med. 41:3231–3242. 2018.PubMed/NCBI
|
|
19
|
Garcia-Rivera D, Delgado R, Bougarne N,
Haegeman G and Berghe WV: Gallic acid indanone and mangiferin
xanthone are strong determinants of immunosuppressive anti-tumour
effects of Mangifera indica L. bark in MDA-MB231 breast cancer
cells. Cancer Lett. 305:21–31. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Noratto GD, Bertoldi MC, Krenek K, Talcott
ST, Stringheta PC and Mertens-Talcott SU: Anticarcinogenic effects
of polyphenolics from mango (Mangifera indica) varieties. J Agric
Food Chem. 58:4104–4112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shi W, Deng J, Tong R, Yang Y, He X, Lv J,
Wang H, Deng S, Qi P, Zhang D, et al: Molecular mechanisms
underlying mangiferin-induced apoptosis and cell cycle arrest in
A549 human lung carcinoma cells. Mol Med Rep. 13:3423–3432. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chieli E, Romiti N, Rodeiro I and Garrido
G: In vitro effects of Mangifera indica and polyphenols derived on
ABCB1/P-glycoprotein activity. Food Chem Toxicol. 47:2703–2710.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng P, Peng ZG, Yang J and Song SJ: The
effect of mangiferin on telomerase activity and apoptosis in
leukemic K562 cells. Zhong Yao Cai. 30:306–309. 2007.(In Chinese).
PubMed/NCBI
|
|
24
|
Peng ZG, Luo J, Xia LH, Chen Y and Song
SJ: CML cell line K562 cell apoptosis induced by mangiferin.
Zhongguo Shi Yan Xue Ye Xue Za Zhi. 12:590–594. 2004.(In Chinese).
PubMed/NCBI
|
|
25
|
Percival SS, Talcott ST, Chin ST, Mallak
AC, Lounds-Singleton A and Pettit-Moore J: Neoplastic
transformation of BALB/3T3 cells and cell cycle of HL-60 cells are
inhibited by mango (Mangifera indica L.) juice and mango juice
extracts. J Nutri. 136:1300–1304. 2006. View Article : Google Scholar
|
|
26
|
Chari NS, Pinaire NL, Thorpe L, Medeiros
LJ, Routbort MJ and McDonnell TJ: The p53 tumor suppressor network
in cancer and the therapeutic modulation of cell death. Apoptosis.
14:336–347. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zou B, Wang H, Liu Y, Qi P, Lei T, Sun M
and Wang Y: Mangiferin induces apoptosis in human ovarian
adenocarcinoma OVCAR3 cells via the regulation of Notch3. Oncol
Rep. 38:1431–1441. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiao J, Liu L, Zhong Z, Xiao C and Zhang
J: Mangiferin regulates proliferation and apoptosis in glioma cells
by induction of microRNA-15b and inhibition of MMP-9 expression.
Oncol Rep. 33:2815–2820. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jung JS, Jung K, Kim DH and Kim HS:
Selective inhibition of MMP-9 gene expression by mangiferin in
PMA-stimulated human astroglioma cells: Involvement of PI3K/Akt and
MAPK signaling pathways. Pharmacol Res. 66:95–103. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Andreu Pardo GL, Maurmann N, Reolon GK, de
Farias CB, Schwartsmann G, Delgado R and Roesler R: Mangiferin, a
naturally occurring glucoxilxanthone improves long-term object
recognition memory in rats. Eur J Pharmacol. 635:124–128. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, MacLeod RA, Masters JR, Nakamura Y, Reid YA,
Reddel RR, et al: Check your cultures! A list of cross-contaminated
or misidentified cell lines. Int J Cancer. 127:1–8. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boyé K, Pujol N, Alves D I, Chen YP,
Daubon T, Lee YZ, Dedieu S, Constantin M, Bello L, Rossi M, et al:
The role of CXCR3/LRP1 cross-talk in the invasion of primary brain
tumors. Nat Commun. 8:15712017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Volpin F, Casaos J, Sesen J, Mangraviti A,
Choi J, Gorelick N, Frikeche J, Lott T, Felder R, Scotland SJ, et
al: Use of an anti-viral drug, Ribavirin, as an anti-glioblastoma
therapeutic. Oncogene. 36:3037–3047. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang Y, Lei T, Du S, Tong R, Wang H, Yang
J, Huang J, Sun M, Wang Y and Dong Z: Nuclear GSK3β induces DNA
double-strand break repair by phosphorylating 53BP1 in
glioblastoma. Int J Oncol. 52:709–720. 2018.PubMed/NCBI
|
|
36
|
Valavanidis A, Vlachogianni T and Fiotakis
C: 8-hydroxy-2′ -deoxyguanosine (8-OHdG): A critical biomarker of
oxidative stress and carcinogenesis. J Environ Sci Health C,
Environ Carcinog Ecotoxicol Rev. 27:120–139. 2009. View Article : Google Scholar
|
|
37
|
Zhang Q, Karnak D, Tan M, Lawrence TS,
Morgan MA and Sun Y: FBXW7 facilitates nonhomologous end-joining
via K63-linked polyubiquitylation of XRCC4. Mol Cell. 61:419–433.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mao Z, Jiang Y, Liu X, Seluanov A and
Gorbunova V: DNA repair by homologous recombination, but not by
nonhomologous end joining, is elevated in breast cancer cells.
Neoplasia. 11:683–691. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shrivastav M, De Haro LP and Nickoloff JA:
Regulation of DNA double-strand break repair pathway choice. Cell
Res. 18:134–147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ricard D, Idbaih A, Ducray F, Lahutte M,
Hoang-Xuan K and Delattre JY: Primary brain tumours in adults.
Lancet. 379:1984–1996. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
del Alcazar Gil CR, Hardebeck MC,
Mukherjee B, Tomimatsu N, Gao X, Yan J, Xie XJ, Bachoo R, Li L,
Habib AA and Burma S: Inhibition of DNA double-strand break repair
by the dual PI3K/mTOR inhibitor NVP-BEZ235 as a strategy for
radiosensitization of glioblastoma. Clin Cancer Res. 20:1235–1248.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gursoy-Yuzugullu O, Carman C, Serafim RB,
Myronakis M, Valente V and Price BD: Epigenetic therapy with
inhibitors of histone methylation suppresses DNA damage signaling
and increases glioma cell radiosensitivity. Oncotarget.
8:24518–24532. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Maachani UB, Kramp T, Hanson R, Zhao S,
Celiku O, Shankavaram U, Colombo R, Caplen NJ, Camphausen K and
Tandle A: Targeting MPS1 enhances radiosensitization of human
glioblastoma by modulating DNA repair proteins. Mol Cancer Res.
13:852–862. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sarkar A, Sreenivasan Y, Ramesh GT and
Manna SK: beta-D-Glucoside suppresses tumor necrosis factor-induced
activation of nuclear transcription factor kappaB but potentiates
apoptosis. J Biol Chem. 279:33768–33781. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rajendran P, Ekambaram G and Sakthisekaran
D: Protective role of mangiferin against Benzo(a)pyrene induced
lung carcinogenesis in experimental animals. Biol Pharm Bull.
31:1053–1058. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kass EM and Jasin M: Collaboration and
competition between DNA double-strand break repair pathways. FEBS
Lett. 584:3703–3708. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Abraham RT: Checkpoint signalling:
Focusing on 53BP1. Nat Cell Biol. 4:E277–E279. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ward IM, Minn K, van Deursen J and Chen J:
p53 Binding protein 53BP1 is required for DNA damage responses and
tumor suppression in mice. Mol Cell Biol. 23:2556–2563. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kurz EU and Lees-Miller SP: DNA
damage-induced activation of ATM and ATM-dependent signaling
pathways. DNA Repair. 3:889–900. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Riballo E, Kühne M, Rief N, Doherty A,
Smith GC, Recio MJ, Reis C, Dahm K, Fricke A, Krempler A, et al: A
pathway of double-strand break rejoining dependent upon ATM,
Artemis, and proteins locating to gamma-H2AX foci. Mol Cell.
16:715–724. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Traven A and Heierhorst J: SQ/TQ cluster
domains: Concentrated ATM/ATR kinase phosphorylation site regions
in DNA-damage-response proteins. Bioessays. 27:397–407. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fernandez-Capetillo O, Chen HT, Celeste A,
Ward I, Romanienko PJ, Morales JC, Naka K, Xia Z, Camerini-Otero
RD, Motoyama N, et al: DNA damage-induced G2-M
checkpoint activation by histone H2AX and 53BP1. Nat Cell Biol.
4:993–997. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mukherjee B, Tomimatsu N, Amancherla K,
Camacho CV, Pichamoorthy N and Burma S: The dual PI3K/mTOR
inhibitor NVP-BEZ235 is a potent inhibitor of ATM- and
DNA-PKCs-mediated DNA damage responses. Neoplasia. 14:34–43. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lei J, Zhou C, Hu H, Hu L, Zhao M, Yang Y,
Chuai Y, Ni J and Cai J: Mangiferin aglycone attenuates
radiation-induced damage on human intestinal epithelial cells. J
Cell Biochem. 113:2633–2642. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Guo HW, Yun CX, Hou GH, Du J, Huang X, Lu
Y, Keller ET, Zhang J and Deng JG: Mangiferin attenuates TH1/TH2
cytokine imbalance in an ovalbumin-induced asthmatic mouse model.
PLoS One. 9:e1003942014. View Article : Google Scholar : PubMed/NCBI
|