Introduction
Rhabdomyosarcoma (RMS) constitutes ~60% of all
childhood soft tissue sarcomas. It is a small, round-cell tumor of
musculoskeletal origin that exhibits various degrees of myogenic
differentiation. Most types of RMS fall into 1 of 2 biological
distinction subgroups defined as alveolar or embryonal type
(1). Alveolar RMS (ARMS), which
accounts for 41% of all RMS, is an aggressive soft tissue sarcoma
in children, with a high invasion and metastasis at initial
diagnosis. Despite a combination therapy, which includes surgical
resection, radiation, and intensive chemotherapy, the prognosis for
patients with ARMS remains poor (2). Recently, many molecular targeted drugs
have been effectively used in patients with different types of
tumors; however, for RMS, targeted drugs are still under
investigation and have not been used clinically (3–5).
Histone deacetylase (HDAC) inhibitors are promising
drugs for the treatment of diverse cancers due to their abilities
to induce cell cycle control and apoptosis (6–9).
Numerous of HDAC inhibitors are currently being assessed in various
phases of clinical trials (10,11).
The United States Food and Drug Administration (US FDA) has already
approved the HDAC inhibitors belinostat, vorinostat, and romidepsin
for the treatment of peripheral and cutaneous T cell lymphoma
(12–14). There have been a few studies on the
tumor suppressive effects of RMS by HDAC inhibition in vitro
(15,16). A recent study in an ARMS mouse model
revealed that response to the HDAC inhibitor was different
depending on the myogenic lineage of the tumor cells (17). Presently, the role of HDAC activity
in RMS tumorigenesis and the essential mechanisms by which HDAC
inhibitors exhibit their antitumor effects remain largely
unknown.
OBP-801 (spiruchostatin A) was originally identified
as an enhancer of PAI-1 gene expression and was established as a
new HDAC inhibitor by a p21 promoter reporter screen (18). OBP-801 exerted the most potent
HDAC-inhibitory activity in our study; it was ~50 times more
effective than vorinostat (18). It
is currently under clinical trials in the United States.
The aim of this study was to evaluate the effect of
the new HDAC inhibitor, OBP-801, on the cell cycle control and on
the viability of RMS cells in vitro and in vivo using
a mouse tumor model. We also assessed the mechanism of action of
OBP-801.
Materials and methods
Cell lines and reagents
We used the human ARMS cell lines SJ-Rh30 (Rh30),
SJ-Rh41 (Rh41), SJ-Rh3 (Rh3), SJ-Rh4 (Rh4), SJ-Rh18 (Rh18) and
SJ-Rh28 (Rh28) that were kindly provided by Peter J. Houghton M,
San Antonio, TX, USA), and the embryonal RMS (ERMS) cell lines RD
that was obtained from JCRB (Japanese Collection of Research
Bioresources) Cell Bank and RMS-YM that was kindly provided by
Naoki Kakazu M.D. (Department of Environmental and Preventive
Medicine, Shimane University School of Medicine, Shimane, Japan)
(5,19). They were maintained in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% fetal bovine
serum (FBS) at 37°C in a 5% CO2 incubator. OBP-801
(Oncolys BioPharma, Inc., Tokyo, Japan) was dissolved in dimethyl
sulfoxide (DMSO) and stored as a 1-mM stock solution in 50-µl
aliquots at −20°C. The percentage of DMSO in all experiments was
<0.01%.
Western blot analysis
Cells were lysed in RIPA buffer (08714-04; Nacalai
Tesque, Kyoto, Japan). Protein concentrations in cell lysates were
determined using BCA assay. A total of 20 µg of protein were
separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) on NuPAGE Novex 4–12% Bis-Tris gels in
NuPAGE MES SDS running buffer (Life Technologies; Thermo Fisher
Scientific, Inc.). Proteins were subsequently transferred to
Immobilon-P membranes in NuPAGE Transfer buffer (Life Technologies;
Thermo Fisher Scientific, Inc). Membranes were blocked in
phosphate-buffered saline with Tween-20 (PBST) containing 5%
non-fat dry milk powder, and then incubated at 4°C overnight with
primary antibodies against the following proteins: anti-histone-H4
(dilution 1:1,000; cat. no. 13919; Cell Signaling Technology, Inc.,
Danvers, MA, USA), anti-acetyl histone-H4 (dilution 1:1,000; cat.
no. 06–866; Merck KGaA, Darmstadt, Germany), anti-Rb (dilution
1:2,000; cat. no. 554136; BD Biosciences, Franklin Lakes, NJ, USA),
anti-phospho-histone-H3 (dilution 1:2,000; cat. no. 3377; Cell
Signaling Technology, Inc.), anti-survivin (dilution 1:2,000; cat.
no. 2808; Cell Signaling Technology, Inc.) and anti-p21 Waf1/Cip1
(1:2,000; cat. no. 2946; Cell Signaling Technology, Inc.),
anti-total caspase-3 (dilution 1:2,000; cat. no. 610322; BD
Biosciences), anti-cleaved caspase-3 (dilution 1:1,000; cat. no.
9664; Cell Signaling Technology, Inc.) and anti-β-actin (dilution
1:5,000; cat. no. A2228; Merck KGaA) for the reference protein. The
membranes were then washed with PBST and incubated at room
temperature for 1 h with sheep anti-mouse secondary antibody
(dilution 1:10,000; cat. no. NA931; GE Healthcare, Little Chalfont,
UK) or donkey anti-rabbit secondary antibody (dilution 1:10,000;
cat. no. NA934; GE Healthcare). Antibody binding was detected using
the enhanced chemiluminescence detection system (ECL and ECL prime;
GE Healthcare) (20). The protein
density of western blot was measured by ImageJ 1.52a (National
Institutes of Health, Bethesda, MD, USA).
WST-8 cell viability assay
We performed WST-8 colorimetric assays with Cell
Counting Kit-8 (Nacalai Tesque, Inc.) according to the
manufacturer's instructions. We seeded the cells in 96-well plates
in 100 µl culture medium for 24 h, then added various reagents. We
determined cell viability by assessing the optical density (OD) at
450 nm with a microplate reader (Multiskan™ JX; Dainippon Sumitomo
Pharma Co., Ltd., Osaka, Japan), as previously described (20).
Cell cycle analysis
To analyze their cell cycle distribution, we
cultured the cells in the presence of OBP-801 or an equivalent
volume of DMSO for 24 h. We then isolated the cells by scraping,
washed them with phosphate-buffered saline (PBS), and incubated
them with propidium iodide at a concentration of 50 µg/ml for 30
min to stain DNA. We determined their DNA content on a FACSCalibur™
flow cytometer (BD Biosciences). We analyzed their cell cycle
status with FlowJo software 7.6.5 (Tree Star, Inc., Ashland, OR,
USA), as previously described (21).
Analysis of apoptosis by flow
cytometry
We analyzed cell death after Annexin V-FITC and
propidium iodide staining with a TACS Annexin V-FITC Apoptosis
Detection kit (R&D Systems, Inc., Minneapolis, MN, USA),
according to the manufacturer's instructions. We analyzed the data
with FlowJo software (Tree Star, Inc.) as previously described
(21).
Immunocytochemistry
We plated the cells on Falcon® 8-Well
Culture Slides (cat. no. 354118; BD Falcon; Corning, Inc., Corning,
NY, USA), then fixed with 4% paraformaldehyde, permeabilized with
0.1% Triton™ X-100, washed with PBS, and incubated with
anti-survivin (dilution 1:1,000; cat. no. 2808), anti-α-tubulin
(dilution 1:1,000; cat. no. 3873), anti-phospho-histone-H3
(dilution 1:1,000; cat. no. 3377), or anti-phospho-H2AX (dilution
1:1,000; cat. no. 9718; all from Cell Signaling Technology, Inc.)
at 4°C overnight. We then rinsed the slides with PBS and incubated
them with Alexa Fluor 488/555-conjugated anti-mouse/rabbit IgG
(dilution 1:200; cat. nos. A11008, A11001 and A21422; Cell
Signaling Technology, Inc.) at room temperature for 1 h. Finally,
we examined the slides by fluorescence microscopy with a KEYENCE
BZ-X700 instrument (Keyence Corp., Osaka, Japan).
In vivo mouse xenograft studies
Female BALB/c nu/nu nude mice (4-weeks old, total
14, 11–16 g) were purchased from Japan SLC, Inc. (Shizuoka, Japan).
All experiments and procedures were conducted in accordance with
the institutional animal care and use committee guidelines. The
present study was also approved by the Committee for Animal
Research of Kyoto Prefectural University of Medicine (permission
no. M27-477). Mice were maintained at 23±2°C under a 12-h
light/dark cycle (light period, 07:00-19:00 h). Food and water were
available ad libitum. We subcutaneously injected
1×107 luciferase-positive Rh30 cells into the dorsal
area of BALB/c nu/nu nude mice (n=3 and 4/group). We monitored
tumor growth in live mice by bioluminescent detection of the
luciferase activity of the Rh30 cells at days 3 and 52, as
previously described (22). We
assessed the tumor sizes twice per week using calipers using the
formula (a × b2)/2. The mice were euthanized by
barbiturate overdose.
Optical imaging for luminescence
We performed in vivo bioluminescence imaging
of live mice using a Xenogen IVIS®-Illumina system
(PerkinElmer, Inc., Waltham, MA, USA). The animals were maintained
under inhaled anesthesia (2% isoflurane in 100% oxygen at the rate
of 2.5 l/min). For imaging of the firefly luciferase reporter
harbored by the tumor cells, we administered a single luciferin
dose of 150 mg/kg (PerkinElmer, Inc.) via intraperitoneal injection
20 min prior to imaging. The data were acquired and analyzed using
the manufacturer's proprietary Living Image software 4.4
(PerkinElmer, Inc.).
Statistical analysis
Average values were expressed as the mean ± standard
deviation (SD). We used the 2-tailed Student's t-test for
comparison of the means between groups. Differences with a P-value
of <0.05 were considered to indicate a statistically significant
difference.
Results
OBP-801 inhibits the growth of RMS
cell lines
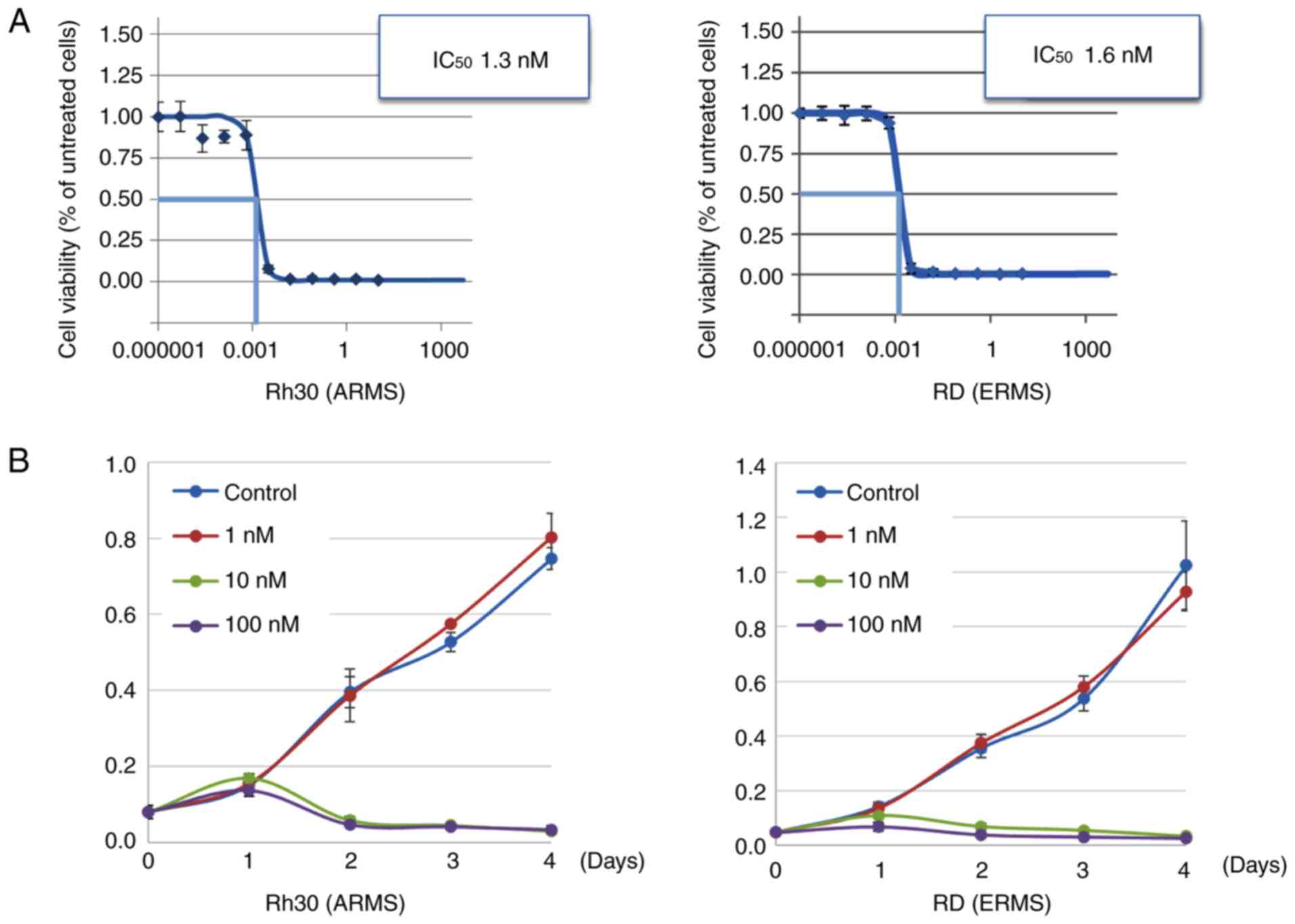
We examined the effect of the HDAC inhibitor OBP-801
on the growth of ERMS and ARMS cell lines. The mean half maximal
inhibitory concentration (IC50) values were 2.6±0.2 nM
for the ERMS cells and 1.8±0.8 nM for the ARMS cells (Table I). OBP-801 inhibited the growth of
RD and Rh30 in a concentration- and time-dependent manner (Fig. 1A and B).
 | Table I.The half maximal inhibitory
concentrations of OBP-801 in the various RMS cell lines. |
Table I.
The half maximal inhibitory
concentrations of OBP-801 in the various RMS cell lines.
| Cell line | Tumor type | IC50
(nM) |
|---|
| RD | ERMS | 2.5±1.4 |
| RMS-YM | ERMS | 2.7±0.3 |
| Rh30 | ARMS | 1.5±0.8 |
| Rh41 | ARMS | 2.5±0.9 |
| Rh3 | ARMS | 0.7±0.4 |
| Rh4 | ARMS | 2.2±1.0 |
| Rh18 | ARMS | 2.6±0.8 |
| Rh28 | ARMS | 0.9±0.7 |
OBP-801 induces cell cycle arrest and
apoptosis in RMS cell lines
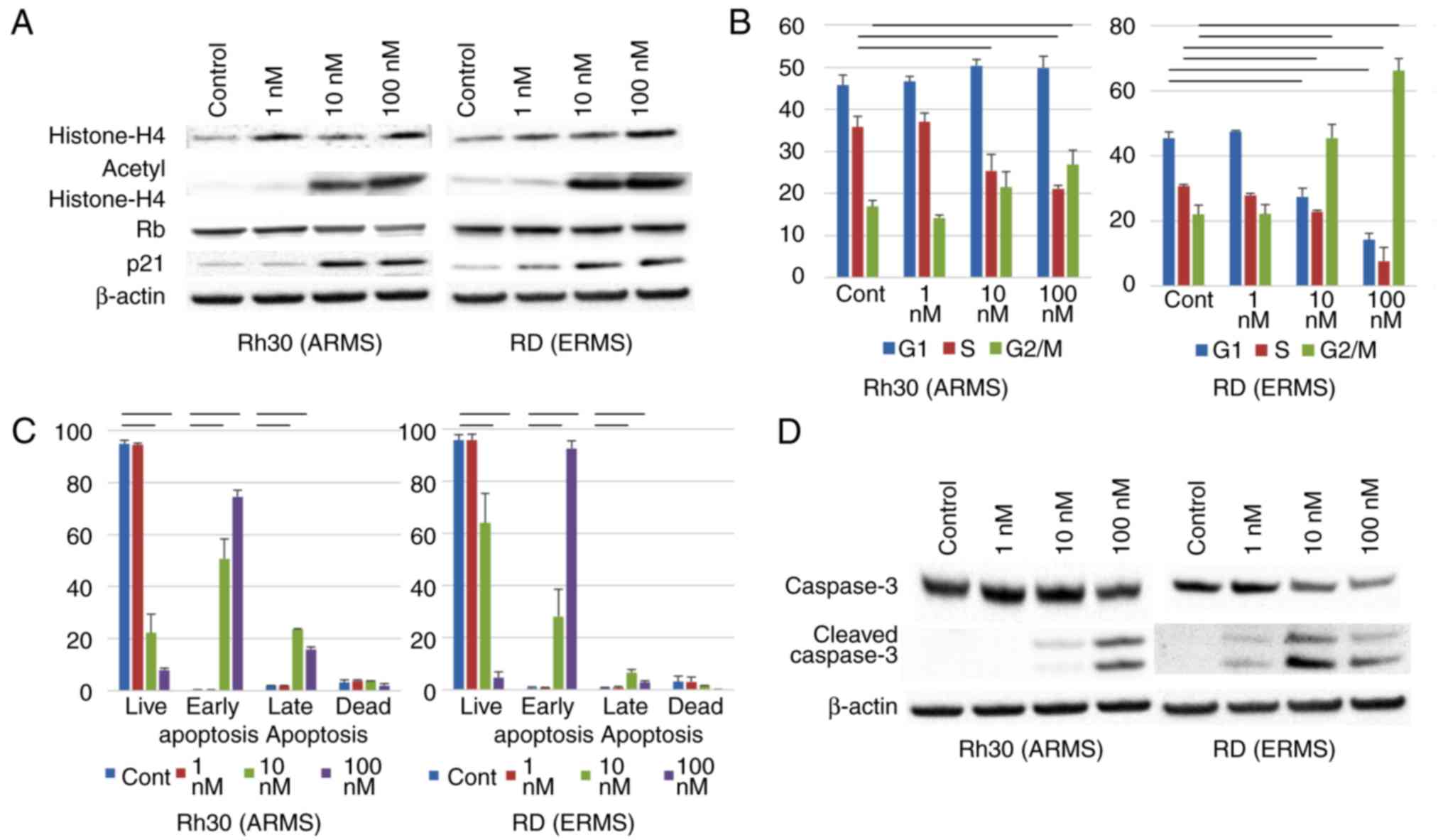
To evaluate the extent of HDAC inhibition in RMS
cells, we performed immunoblot analyses using anti-acetylated
histone antibodies. After 24 h, OBP-801 induced the accumulation of
acetylated histone H4 in a concentration-dependent fashion
(Fig. 2A). OBP-801 also induced
p21waf1/Cip1 in a concentration-dependent manner, and
hypophosphorylation of Rb in ARMS, but not in ERMS cells (Fig. 2A). We observed that OBP-801 (10 nM,
24 h) induced arrest in the G1 and G2/M-phases in ARMS cells and in
the G2/M-phase in ERMS cells (Fig.
2B). In addition, OBP-801 induced cell death in RMS cells
(Fig. 1B). OBP-801 (10 nM) induced
early and late apoptosis in RMS cells 48 h after the treatment, as
indicated by Annexin V staining assessed by flow cytometry
(Fig. 2C). Treatment with OBP-801
also led to the expression of cleaved caspase-3 in a
concentration-dependent manner (Fig.
2D).
OBP-801 causes RMS cell death via
mitotic catastrophe
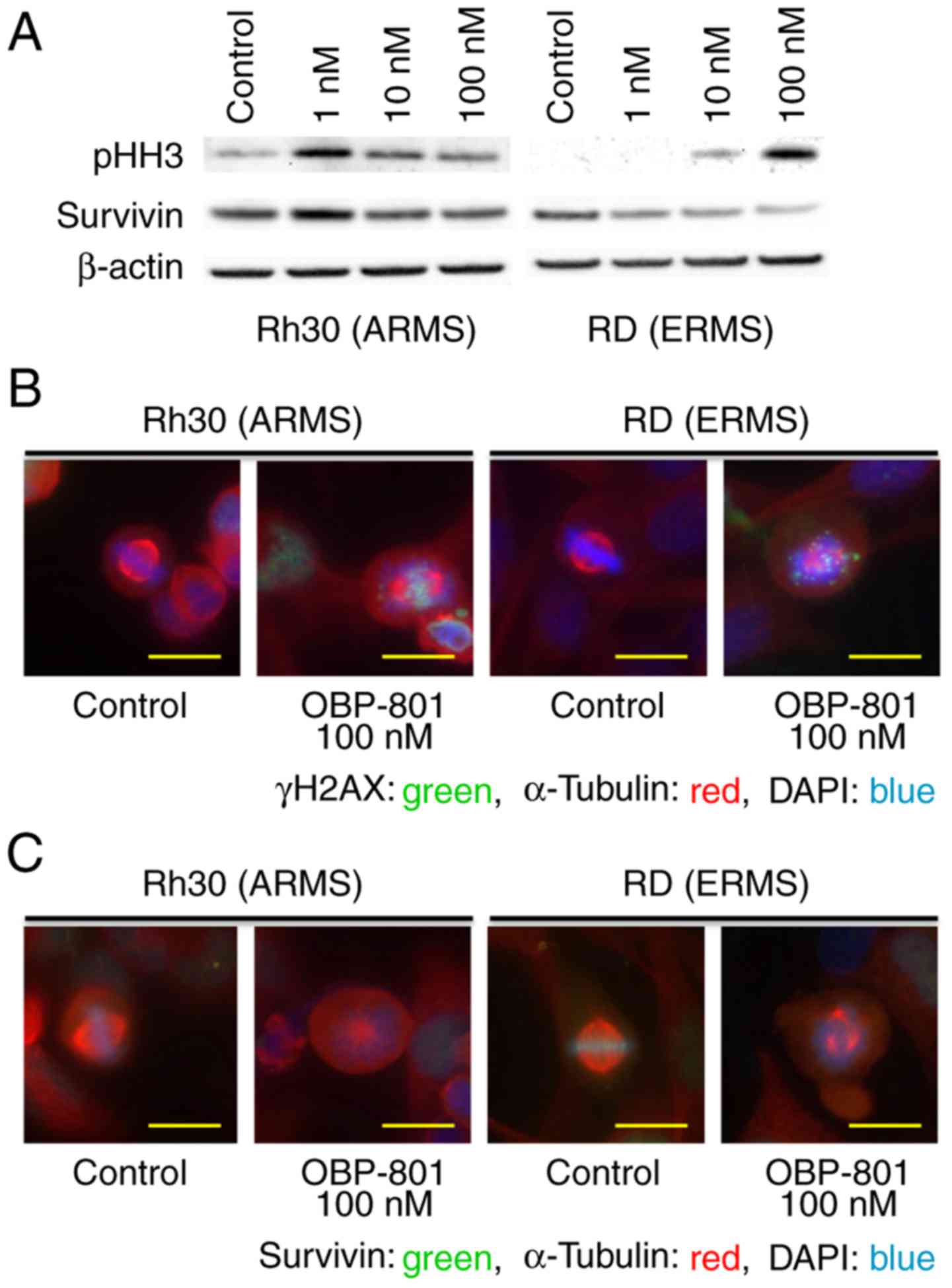
To examine if OBP-801 caused G2- or M-phase arrest,
we immunoblotted with an antibody against phospho-histone H3, a
marker of mitosis. Cells treated with OBP-801 had higher
phosphorylation levels of histone H3 than control cells 24 h after
the treatment; the effect was concentration-dependent (Fig. 3A). We next analyzed the nuclear
morphology of RMS cells with OBP-801 treatment. We found that
control cells exhibited normal chromosome distribution in
metaphase, with correctly formed mitotic spindles. However, upon
treatment with OBP-801, dividing RMS cells exhibited aberrant
metaphase morphologies: their chromosomes were not aligned at the
metaphase plate, they had an abnormal mitotic spindle distribution,
survivin was not recruited to the centromeres, and exhibited lower
levels of survivin than that in control cells (Fig. 3B and C); OBP-801 affected the
abundance of survivin in a dose-dependent manner (Fig. 3A). In addition, OBP-801-treated RMS
cells were stained with the γH2AX antibody, which indicated that
they entered mitosis with damaged DNA (Fig. 3C).
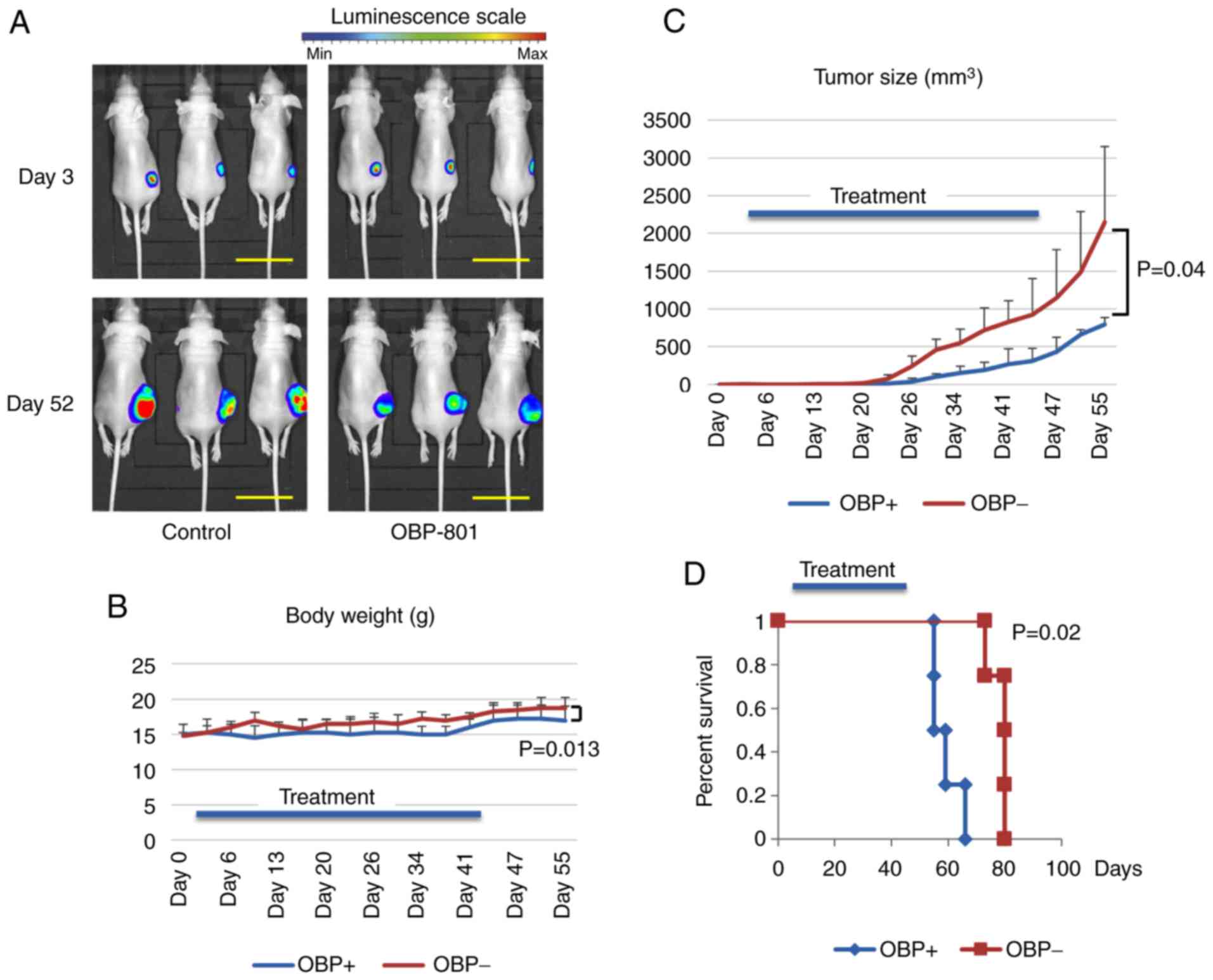
OBP-801 inhibits ARMS tumor growth in
vivo and improves the survival of tumor-bearing mice
We measured the bioluminescence from Rh30 cells in
nude mice at days 3 and 52 after injection and found that the mice
that had received OBP-801 had lower tumor-related bioluminescence
intensities at day 52 (Fig. 4A).
The mice in the drug-treated group had smaller tumors and survived
longer than those in the control group (Fig. 4C and D). We did not observe
statistically significant differences between the body weights of
the mice in the drug-treated and control groups, indicating that
the drug did not cause toxicity (Fig.
4B).
Discussion
We used a new HDAC inhibitor, OBP-801, as a
potential therapeutic drug for treating RMS. HDAC inhibitor-induced
cell death is tumor-selective, as previously described (23,24).
The IC50 values of OBP-801 that we observed for ERMS
cells were very low compared with the concentration required to
induce notable toxic effects in normal human fibroblasts (30 µM)
(18). The lack of apparent
cytotoxic effects in our mouse tumor model supports the
tumor-selectivity of OBP-801.
The cyclin-dependent kinase inhibitor, p21, a
regulator of cellular proliferation via G1 arrest, is a crucial
target for HDAC inhibitors (25,26).
In this study, we confirmed that OBP-801 increased the protein
level of p21 Waf1/Cip1 in a concentration-dependent manner in RMS
cells, possibly via the induction of p21 mRNA. Notably, however,
M-phase arrest was predominant than G1- and G2-phase arrest. These
findings indicated that OBP-801 may induce arrest of M-phase
followed by cell death in RMS cells.
Based on our observation that OBP-801 induced cell
arrest of M-phase followed by apoptosis in RMS cells, we
investigated whether apoptosis was associated with mitotic
catastrophe. Mitotic catastrophe is a regulated antitumor mechanism
that disrupts the survival and/or proliferation of cells that are
unable to undergo complete mitosis due to unrecoverable DNA damage
and that have mitotic machinery problems, and/or have a failure at
mitotic checkpoints (27–29). Recently, relationships were reported
between HDACs and mitotic arrest/catastrophe via DNA damage
(30) and between Aurora B kinase
activity (31) and Eg5 acetylation
(32). Mitotic catastrophe is
assessed based on unique morphological nuclear changes, such as
multinucleation, macronucleation, and micronucleation. We found
that OBP-801 treatment of dividing RMS cells induced γH2AX-positive
aberrant metaphase morphologies, concurrent with lower levels and
misalignment of survivin. Recently, Aljaberi et al reported
that the activity of survivin in mitosis was regulated by cyclical
acetylation and deacetylation during mitosis (33), which may explain how an HDAC
inhibitor could induce mitotic arrest via survivin in a manner
consistent with our results. These results indicated that OBP-801
may induce apoptosis in RMS cells via mitotic catastrophe.
In conclusion, to the best of our knowledge, this is
the first study revealing that treatment of RMS cells with the
novel HDAC inhibitor OBP-801 induced M-phase arrest followed by
apoptosis via mitotic catastrophe. These results indicated that
OBP-801 may be promising for the treatment of RMS. We believe that
this drug should be chosen for future clinical trials.
Acknowledgements
The authors acknowledge Peter J. Houghton M.D.
(Greehey Children's Cancer Research Institute, University of Texas
Health Science Center at San Antonio) for the cell lines (Rh30,
Rh41, Rh3, Rh4, Rh18 and Rh28) and Naoki Kakazu M.D. (Department of
Environmental and Preventive Medicine, Shimane University School of
Medicine, Shimane, Japan) for cell line RMS-YM.
Funding
The present study was supported by JSPS KAKENHI
grant no. JP26461591, JSPS KAKENHI grant no. JP25253095, and the
Health Labour Sciences Research grant no. 17ck0106333 h.
Availability of data and materials
All the data supporting the conclusions of this
article are included in the article.
Authors' contributions
CT, KK, TI and HH conceived and designed this study.
Experimental methodology was developed by KK, SY, MM, KT, TI, TS
and HH. Experimental procedures were carried out by CT, DK and KK.
All authors participated in the analysis and interpretation of
data. CT and KK performed the statistical analysis and wrote the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Committee for
Animal Research of Kyoto Prefectural University of Medicine
(permission no. M27-477).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
RMS
|
rhabdomyosarcoma
|
|
ARMS
|
alveolar RMS
|
|
HDAC
|
histone deacetylase
|
|
ERMS
|
embryonal RMS
|
|
DMSO
|
dimethyl sulfoxide
|
|
PBS
|
phosphate-buffered saline
|
|
SD
|
standard deviation
|
|
IC50
|
half maximal inhibitory
concentration
|
References
|
1
|
Merlino G and Helman LJ:
Rhabdomyosarcoma-working out the pathways. Oncogene. 18:5340–5348.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Davicioni E, Anderson MJ, Finckenstein FG,
Lynch JC, Qualman SJ, Shimada H, Schofield DE, Buckley JD, Meyer
WH, Sorensen PH, et al: Molecular classification of
rhabdomyosarcoma-genotypic and phenotypic determinants of
diagnosis: A report from the Children's Oncology Group. Am J
Pathol. 174:550–564. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kikuchi K, Soundararajan A, Zarzabal LA,
Weems CR, Nelon LD, Hampton ST, Michalek JE, Rubin BP, Fields AP
and Keller C: Protein kinase C iota as a therapeutic target in
alveolar rhabdomyosarcoma. Oncogene. 32:286–295. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miyachi M, Kakazu N, Yagyu S, Katsumi Y,
Tsubai-Shimizu S, Kikuchi K, Tsuchiya K, Iehara T and Hosoi H:
Restoration of p53 pathway by nutlin-3 induces cell cycle arrest
and apoptosis in human rhabdomyosarcoma cells. Clin Cancer Res.
15:4077–4084. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sokolowski E, Turina CB, Kikuchi K,
Langenau DM and Keller C: Proof-of-concept rare cancers in drug
development: The case for rhabdomyosarcoma. Oncogene. 33:18772014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Minucci S and Pelicci PG: Histone
deacetylase inhibitors and the promise of epigenetic (and more)
treatments for cancer. Nat Rev Cancer. 6:38–51. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bolden JE, Peart MJ and Johnstone RW:
Anticancer activities of histone deacetylase inhibitors. Nat Rev
Drug Discov. 5:769–784. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khabele D, Son DS, Parl AK, Goldberg GL,
Augenlicht LH, Mariadason JM and Montgomery RV: Drug-induced
inactivation or gene silencing of class I histone deacetylases
suppresses ovarian cancer cell growth: implications for therapy.
Cancer Biol Ther. 6:795–801. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Strait KA, Warnick CT, Ford CD, Dabbas B,
Hammond EH and Ilstrup SJ: Histone deacetylase inhibitors induce
G2-checkpoint arrest and apoptosis in
cisplatinum-resistant ovarian cancer cells associated with
overexpression of the Bcl-2-related protein Bad. Mol Cancer Ther.
4:603–611. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khan O and La Thangue NB: HDAC inhibitors
in cancer biology: Emerging mechanisms and clinical applications.
Immunol Cell Biol. 90:85–94. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
West AC and Johnstone RW: New and emerging
HDAC inhibitors for cancer treatment. J Clin Invest. 124:30–39.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Porcu P and Wong HK: We should have a
dream: Unlocking the workings of the genome in cutaneous T-cell
lymphomas. Clin Lymphoma Myeloma. 9:409–411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mann BS, Johnson JR, Cohen MH, Justice R
and Pazdur R: FDA approval summary: Vorinostat for treatment of
advanced primary cutaneous T-cell lymphoma. Oncologist.
12:1247–1252. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jain S and Zain J: Romidepsin in the
treatment of cutaneous T-cell lymphoma. J Blood Med. 2:37–47.
2011.PubMed/NCBI
|
|
15
|
Blattmann C, Oertel S, Ehemann V, Thiemann
M, Huber PE, Bischof M, Witt O, Deubzer HE, Kulozik AE, Debus J, et
al: Enhancement of radiation response in osteosarcoma and
rhabdomyosarcoma cell lines by histone deacetylase inhibition. Int
J Radiat Oncol Biol Phys. 78:237–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kutko MC, Glick RD, Butler LM, Coffey DC,
Rifkind RA, Marks PA, Richon VM and LaQuaglia MP: Histone
deacetylase inhibitors induce growth suppression and cell death in
human rhabdomyosarcoma in vitro. Clin Cancer Res. 9:5749–5755.
2003.PubMed/NCBI
|
|
17
|
Abraham J, Nuñez-Álvarez Y, Hettmer S,
Carrió E, Chen HI, Nishijo K, Huang ET, Prajapati SI, Walker RL,
Davis S, et al: Lineage of origin in rhabdomyosarcoma informs
pharmacological response. Genes Dev. 28:1578–1591. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shindoh N, Mori M, Terada Y, Oda K, Amino
N, Kita A, Taniguchi M, Sohda KY, Nagai K, Sowa Y, et al: YM753, a
novel histone deacetylase inhibitor, exhibits antitumor activity
with selective, sustained accumulation of acetylated histones in
tumors in the WiDr xenograft model. Int J Oncol. 32:545–555.
2008.PubMed/NCBI
|
|
19
|
Kubo K, Naoe T, Utsumi KR, Ishiguro Y,
Ueda K, Shiku H and Yamada K: Cytogenetic and cellular
characteristics of a human embryonal rhabdomyosarcoma cell line,
RMS-YM. Br J Cancer. 63:879–884. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kikuchi K, Tsuchiya K, Otabe O, Gotoh T,
Tamura S, Katsumi Y, Yagyu S, Tsubai-Shimizu S, Miyachi M, Iehara
T, et al: Effects of PAX3-FKHR on malignant phenotypes in alveolar
rhabdomyosarcoma. Biochem Biophys Res Commun. 365:568–574. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kikuchi K, Hettmer S, Aslam MI, Michalek
JE, Laub W, Wilky BA, Loeb DM, Rubin BP, Wagers AJ and Keller C:
Cell-cycle dependent expression of a translocation-mediated fusion
oncogene mediates checkpoint adaptation in rhabdomyosarcoma. PLoS
Genet. 10:e10041072014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nishijo K, Hosoyama T, Bjornson CR,
Schaffer BS, Prajapati SI, Bahadur AN, Hansen MS, Blandford MC,
McCleish AT, Rubin BP, et al: Biomarker system for studying muscle,
stem cells, and cancer in vivo. FASEB J. 23:2681–2690. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Marks PA, Richon VM, Miller T and Kelly
WK: Histone deacetylase inhibitors. Adv Cancer Res. 91:137–168.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ungerstedt JS, Sowa Y, Xu WS, Shao Y,
Dokmanovic M, Perez G, Ngo L, Holmgren A, Jiang X and Marks PA:
Role of thioredoxin in the response of normal and transformed cells
to histone deacetylase inhibitors. Proc Natl Acad Sci USA.
102:673–678. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zupkovitz G, Grausenburger R, Brunmeir R,
Senese S, Tischler J, Jurkin J, Rembold M, Meunier D, Egger G,
Lagger S, et al: The cyclin-dependent kinase inhibitor p21 is a
crucial target for histone deacetylase 1 as a regulator of cellular
proliferation. Mol Cell Biol. 30:1171–1181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bose P, Dai Y and Grant S: Histone
deacetylase inhibitor (HDACI) mechanisms of action: Emerging
insights. Pharmacol Ther. 143:323–336. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Galluzzi L, Vitale I, Aaronson SA, Abrams
JM, Adam D, Agostinis P, Alnemri ES, Altucci L, Amelio I, Andrews
DW, et al: Molecular mechanisms of cell death: recommendations of
the Nomenclature Committee on Cell Death 2018. Cell Death Differ.
25:486–551. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Castedo M, Perfettini JL, Roumier T,
Andreau K, Medema R and Kroemer G: Cell death by mitotic
catastrophe: A molecular definition. Oncogene. 23:2825–2837. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vitale I, Galluzzi L, Castedo M and
Kroemer G: Mitotic catastrophe: a mechanism for avoiding genomic
instability. Nat Rev Mol Cell Biol. 12:385–392. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cornago M, Garcia-Alberich C,
Blasco-Angulo N, Vall-Llaura N, Nager M, Herreros J, Comella JX,
Sanchis D and Llovera M: Histone deacetylase inhibitors promote
glioma cell death by G2 checkpoint abrogation leading to mitotic
catastrophe. Cell Death Dis. 5:e14352014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Kao GD, Garcia BA, Shabanowitz J,
Hunt DF, Qin J, Phelan C and Lazar MA: A novel histone deacetylase
pathway regulates mitosis by modulating Aurora B kinase activity.
Genes Dev. 20:2566–2579. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nalawansha DA, Gomes ID, Wambua MK and
Pflum MKH: HDAC inhibitor-induced mitotic arrest ss mediated by
Eg5/KIF11 acetylation. Cell Chem Biol. 24:481–492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Aljaberi AM, Webster JR and Wheatley SP:
Mitotic activity of survivin is regulated by acetylation at K129.
Cell Cycle. 14:1738–1747. 2015. View Article : Google Scholar : PubMed/NCBI
|