Introduction
Pancreatic cancer is an aggressive tumor type, with
the highest mortality rate and poorest long-term prognosis of all
cancer types. Since the pancreas is located in a deep
retroperitoneal site and no specific symptoms are observable at
early stages of pancreatic cancer, the diagnosis at a surgically
resectable stage is difficult (1,2).
Pancreatic cancer is a highly lethal disease for which limited
therapeutic options are available, which often combines gemcitabine
with other chemotherapeutics (3,4).
However, these chemotherapeutic agents are highly toxic and lack
therapeutic efficacy. In particular, clinical beneficial responses
to gemcitabine are only observed in approximately 25% of cases,
and, owing to long-term tolerance, this limited therapeutic
efficacy rapidly declines resulting in a median overall survival of
6 months (5,6). Therefore, a novel strategy is needed
to optimize the efficacy of gemcitabine in treating pancreatic
cancer.
Oncolytic viruses (OVs) are native or recombinant
viruses which can selectively kill cancer cells and associated
stromal cells directly by oncolysis, indirectly by immune mediated
clearance of cancer cells, or targeting of tumor vasculature.
Vaccinia virus is highly immunogenic and has properties that make
it an ideal oncolytic immunotherapy vector (7). Preclinical murine studies have
demonstrated significant antitumor efficacy and systemic antitumor
immunity, using a tumor-selective oncolytic vaccinia virus
expressing immunogenic transgenes (8–11). An
oncolytic vaccinia virus armed with GM-CSF (Pexa-Vec) was
associated with a 15% objective response rate in patients with
advanced hepatocellular carcinoma in a randomized phase II clinical
trial (11). The oncolytic vaccinia
virus (oVV), which selectively replicates in cancer cells, is a
promising alternative to conventional therapy for cancer treatment
and has been extensively examined in clinical trials (11–13).
In addition to its safety profile, one of the most important
advantages is that the ~200 kb genome of vaccinia virus enables the
insertion of foreign genes up to 25 kb in length. Furthermore, the
oVV has a broad range of host cells, high capacity of transgene
expression, and its activity is unrestrained by hypoxia (12,14).
Indeed, oVV-based gene therapy has been investigated in a range of
tumor types, such as myeloma, pancreatic carcinoma, hepatocellular
carcinoma, and gastric carcinoma, and has been reported to induce
significant growth suppression with relatively limited side
effects, indicating that oVV is a promising vector for cancer gene
therapy (15–17).
The second mitochondrial-derived activator of
caspase (Smac) is released from mitochondria into the cytosol
during the process of apoptosis (18,19).
It has been previously demonstrated that the release of Smac is
critical for apoptosis induced by anti-myeloma agents (20). Furthermore, alterations in Smac
release contribute to drug resistance in cancer cells. Our previous
study demonstrated the role of Smac in promoting apoptosis in
pancreatic cancer cells by decreasing the expression of the
inhibitor of apoptosis proteins (IAPs) (21). Based on these previous findings, the
regulation of Smac expression seems a promising therapy in
pancreatic cancer. We previously demonstrated that oVV-expressing
Smac exerted potent antitumor efficacy in hepatocellular
carcinoma (22) and
Smac-armed oncolytic adenovirus significantly inhibited
pancreatic cancer and multiple myeloma growth (23,24).
However, the therapeutic efficacy of oVV-mediated Smac gene
therapy in human pancreatic cancer is yet to be elucidated.
This present study aimed at investigating the
antitumor properties of oVV-Smac both alone or combined with
gemcitabine in pancreatic cancer cells and xenograft mouse
models.
Materials and methods
Cell lines and virus
The human pancreatic cancer cell lines SW1990,
BXPC-3 and PANC-1 were purchased from the Cell Bank of the Type
Culture Collection of the Chinese Academy of Sciences (Shanghai,
China). SW1990/GEM is gemcitabine-resistant SW1990 cell line, which
was retained in our laboratory. All the cell lines were
authenticated by short-tandem repeat profiling and cultured in
Gibco™ Dulbecco's modified Eagle's medium (DMEM) Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% heat
inactivated fetal bovine serum (FBS; Thermo Fisher Scientific,
Inc.). Cells were incubated in a 5% CO2 humidified
incubator at 37°C. Construction and production of recombinant
oncolytic vaccinia virus oVV-Smac and oVV were previously described
(24). The Smac gene was inserted
into the thymidine kinase (TK) region, disrupting the function of
TK. Deletion of the TK gene inhibits viral replication in normal,
non-dividing cells (25). However,
cancer cells have a high concentration of functional nucleotides
that enables oVV replication to occur in the absence of viral TK.
Therefore, disruption of TK results in selective replication of the
oVV in tumor cells. The T7 promoter was inserted before the
exogenous genes to initiate their expression, and the gpt gene
works as a screen gene engineered behind the exogenous genes. The
whole expression cassette was constructed into the pCB vector,
which is a shuttle plasmid for vaccinia virus packaging kindly
provided by academician Xinyuan Liu (Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences, Shanghai, China).
Each recombinant vaccinia virus was isolated through three rounds
of plaque purification in 293 cells and purified by
ultracentrifugation in a cesium chloride gradient. Moreover, virus
titers were determined by TCID50 assay in 293 cells. Cells were
infected with vaccinia virus at different doses at 37°C in a
humidified atmosphere containing 5% CO2.
Cell viability assay
PANC-1, SW1990 and BxPC-3 cells were dispensed in
96-well culture plates at a density of 5×103 cells/well.
After attachment, cells were infected with oVV, oVV-Smac with or
without gemcitabine at given concentrations and times. The medium
added together with PBS was used as a blank control. The cell
survival rate was evaluated by a standard
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), Medium was
removed and fresh medium containing MTT (5 mg/ml) was added to each
well. The cells were incubated at 37°C for 4 h, and after the
supernatant was drawn off of each well carefully and an equal
volume (150 µl) of DMSO was added to each well and mixed thoroughly
on a concentrating table for 10 min. The absorbance of the plates
was read at 595 nm with a GENios model DNA Expert Microplate Reader
(Tecan Group, Ltd., Mannedorf, Switzerland). For combination index
plots, CI is expressed as the log10(CI) ± 1.96 SD, and
the 95% confidence intervals (CIs) are shown where estimable, with
the use of the algebraic approximation algorithm of the CalcuSyn
program (Biosoft, Cambridge, UK). In the present study, CI values
were calculated over a scope of levels of growth inhibition (GI)
from 20 to 80% of the fraction affected.
Western blot analysis
Cells were harvested in lysis buffer (Beyotime
Institute of Biotehnology, Jiangsu, China) containing 1% Complete
Mini-Protease Inhibitor Cocktail (Roche Diagnostics, Basel,
Switzerland) and 5 mM NaF. Protein extractions were quantified
using the BCA kit (Thermo Fisher Scientific, Inc.) and heated for
10 min at 100°C. Protein (30 µg) was resolved on 12% SDS-PAGE and
transferred to nitrocellulose membranes (Merk Millipore, Darmstadt,
Germany). After blocking for 1 h at 37°C, the membranes were
immunoblotted with different antibodies overnight at 4°C.
Antibodies against Smac (dilution 1:1,000; cat. no. ab8114),
vaccinia virus (dilution 1:500; cat. no. ab19970), GAPDH (dilution
1:2,000; cat. no. ab128915) were purchased from Abcam (Shanghai,
China). Antibodies against caspase-8 (dilution 1:1,000; cat. no.
MABC1606), caspase-9 (dilution 1:1,000; cat. no. MAB4709),
caspase-3 (dilution 1:1,000; cat. no. AB3623), PARP (dilution
1:500; cat. no. AB16661), XIAP (dilution 1:1,000; cat. no. 07735),
cIAP-1 (dilution 1:1,000; cat. no. ABC448), cIAP-2 (dilution
1:1,000; cat. no. AB3615), survivin (dilution 1:1,000; cat. no.
MAB4617), livin (dilution 1:1,000; cat. no. ABC97), P-gp (dilution
1:500; cat. no. ABN455) and MDR1 (dilution 1:1,000; cat. no.
MAB4162) were purchased from EMD Millipore Corp. (Billerica, MA,
USA). Membranes were then washed with TBST and incubated with
HRP-conjugated goat anti-rabbit (dilution 1:5,000; cat. no. HA1001)
or anti-mouse antibody (dilution 1:5,000; cat. no. HA1006; both
from for Huabio, Hangzhou, China) for 1 h at room temperature.
Finally, blots were detected using ChemiDoc™ MP Imaging System
(Bio-Rad Laboratories) with a Super Enhanced chemiluminescence
detection kit (Applygen Technologies, Inc., Beijing, China).
Flow cytometric analysis
Cells infected with oncolytic vaccinia viruses
and/or gemcitabine were trypsinized and washed once with complete
medium. Aliquots of cells (5×105) were resuspended in
500 ml of binding buffer and stained with fluorescein
isothiocyanate (FITC)-labeled Annexin V and propidium iodide (PI)
(BioVision, Inc., Palo Alto, CA, USA) according to the
manufacturer's instructions. Cell apoptosis and cell cycle were
examined using FACS (FACStar cytofluorometer; BD Biosciences, San
Jose, CA, USA).
Real-time quantitative PCR
RNA was extracted with Invitrogen™ Trizol reagent
(Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions. cDNA was generated using the PrimeScript RT reagent
kit (Takara Bio, Inc., Tokyo, Japan). The qPCR reactions were
conducted in a total volume of 20 µl by using the following
procedure: 1 cycle at 95°C (10 min), then 60°C (30 sec), followed
by 39 cycles at 95°C (10 sec) and 60°C (30 sec). PCR amplicons were
determined based on SYBR-Green I detection (Roche Diagnostic,
Indianapolis, IN, USA), and the authenticity was certified by
melting curve analysis. Quantitative PCR was operated using the
CFX-96 qPCR system and iQ SYBR-Green SuperMix (Bio-Rad
Laboratories). Relative gene expression was determined via the
2ΔΔCt method. The primers used are as follows: Smac,
5′-GGAAGATCTCCTCCTGCATCC-3′ (forward) and
5′-CCGCTTAAGATACCGCTCGAG-3′ (reverse); GAPDH,
5′-CTTTGGTATCGTGGAAGGACTC-3′ (forward) and
5′-GTAGAGGCAGGGGATGATGTTCT-3′ (reverse).
Animal experiments
All animal experiments were approved by the
Institutional Animal Care and Use Committee, Zhejiang Provincial
People's Hospital and all procedures were in accordance to the
Guide for the Care and Use of Laboratory Animals (National
Academies Press, Washington, DC). One hundred female BALB/c nude
mice (4- to 5-weeks old, 20 g) were purchased from the Shanghai
Experimental Animal Center (Shanghai, China). All 100 mice were
housed in a specific pathogen-free environment, in which the
temperature was maintained at 26–28°C, the humidity was 40–60%, and
the daily light was maintained for 10 h (14 h without light).
Ventilation was ensured 10 to 15 times per hour. When the mouse
tumor reached a diameter of 2.0 cm or a volume of 2.5
cm3, or all mice in the PBS group died, we stopped the
mouse experiment. The sensitive SW1990 cells or SW1990/GEM cells
were injected subcutaneously into the lower right flank of female
nude mice and the tumor xenograft model was established. Each group
was composed of at least 8 animals and tumor growth was monitored
and measured for every 4 days with a Vernier caliper. Tumor volume
(V) was calculated according to the formula: V (mm3) =
1/2 × length (mm) × width (mm)2. Once the subcutaneous
tumors reached ~100 mm3, the nude mice were divided into
5 groups (8 mice in each group) randomly. Subsequently, mice were
injected with gemcitabine, oVV, oVV-Smac, gemcitabine plus
oVV-Smac, or PBS. oVV and oVV-Smac (2×107 plaque forming
unit/mouse) was injected once every day for a total of 4 times
through intratumoral injection; gemcitabine was injected
intraperitoneally at a total dose of 30 mg/kg body weight; and PBS
100 µl as control for a total of 4 times once every day.
Mice were sacrificed at 2 weeks post-injection
according to ethical instructions by carbon dioxide. Tumors were
separated, fixed using 4% paraformaldehyde, embedded in paraffin,
finally cut into 4-µm sections for hematoxylin and eosin staining,
immunohistochemical analysis and TUNEL assay according to the
manufacturers' instructions. For immunohistochemical analysis,
slides were incubated with primary antibody anti-Smac (dilution
1:100; cat. no. ab8114; Abcam, Shanghai, China) overnight at 4°C,
and then incubated with biotinylated secondary antibody (dilution
1:1,000; cat. no. B2763; Thermo Fisher Scientific, Inc.) and
further visualized using a diaminobenzidine (DAB) kit (Thermo
Scientific, Inc.).
Statistical analysis
For comparison between 2 groups, significant
differences were determined using the Student's t-test.
Analysis of variance (ANOVA) followed by a Bonferroni
multiple-group comparison test was applied for comparison of 3 or
more groups. The analysis of the combined effects was performed
with CalcuSyn software 2.0 (Biosoft, Cambridge, UK). Data are
expressed as the mean ± SD. Statistical analysis was performed with
IBM SPSS Statistics software version 20 (IBM Corp., Armonk, NY,
USA). Statistical significance was defined at P<0.05.
Results
Characterization of oVV-Smac in
vitro
The generation of oVV-Smac was performed by
homologous recombination as described in our previous study
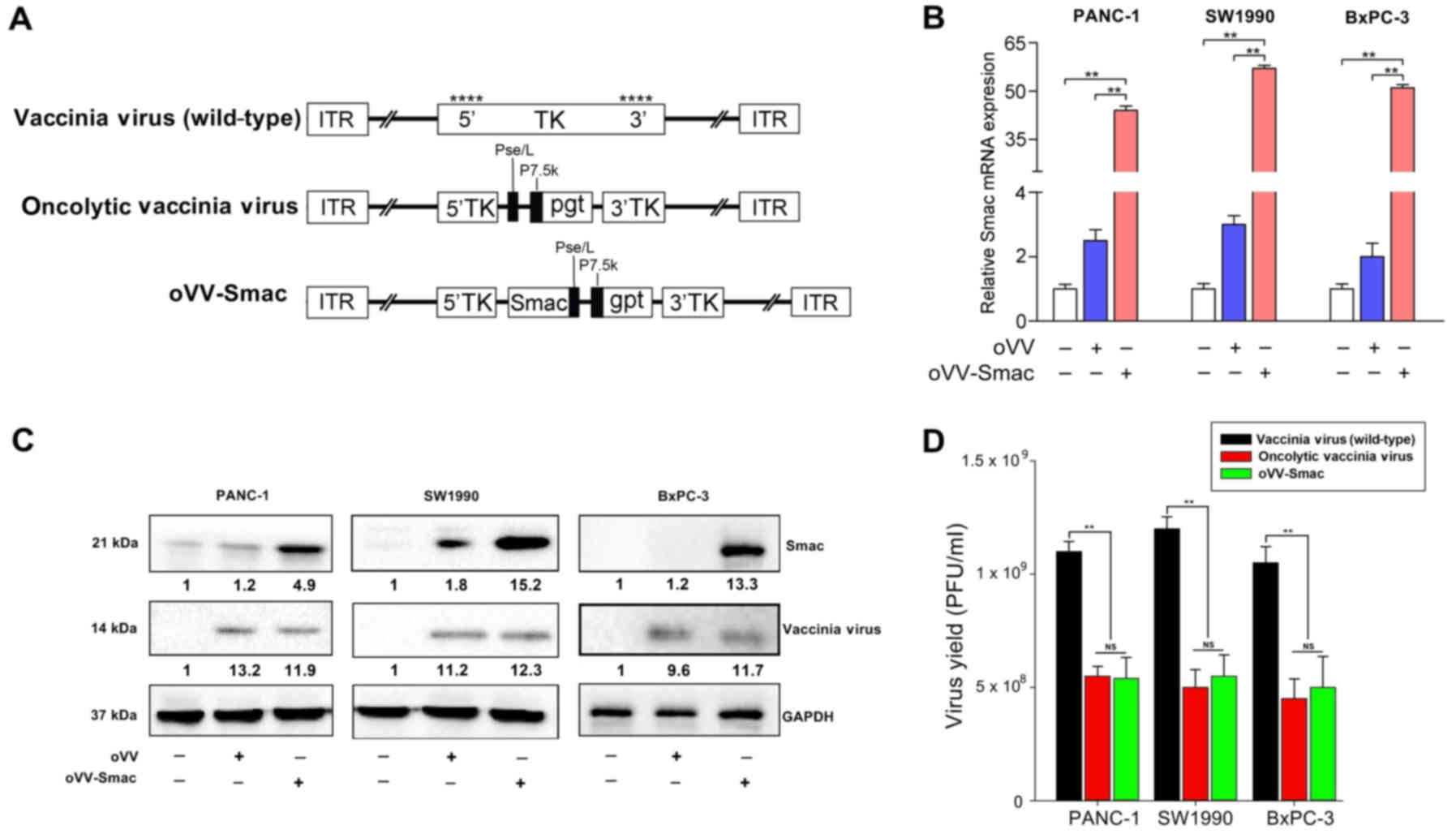
(24). Fig. 1A depicts the construction scheme of
oVV and oVV-Smac. Real-time quantitative polymerase chain
reaction (qPCR) confirmed exogenous Smac expression. Three
pancreatic cancer cell lines, namely PANC-1, SW1990 and BxPC-3,
were infected with oVV and oVV-Smac at multiplicities of
infection (MOI) of 10 for 24 h. As expected, a significant amount
of Smac was observed in all oVV-Smac-transfected
pancreatic cancer cell lines, but not in the oVV- or
phosphase-buffered saline (PBS)-treated group (Fig. 1B). Similar results were obtained
when the expression of Smac and vaccinia virus A27L was
determined at the protein level by western blot analysis (Fig. 1C), suggesting that Smac was
overexpressed in oVV-Smac-transfected cells both at the
transcriptional and translational levels.
To control for any interference of the transgene and
modified genome of vaccinia virus with the selective replicative
ability of the recombinant oVV in different cell lines, a progeny
assay was performed by infecting the three pancreatic cancer cells
with various constructs including vaccinia virus (wild-type), oVV
and oVV-Smac. The results indicated that both oncolytic
viruses oVV and oVV-Smac replicated easily in the infected
pancreatic cancer cells and yielded a high virus progeny (Fig. 1D). Thus, the selective replicative
ability of oVV in cancer cells was not affected by the insertion of
Smac and thymidine kinase (TK) deletion.
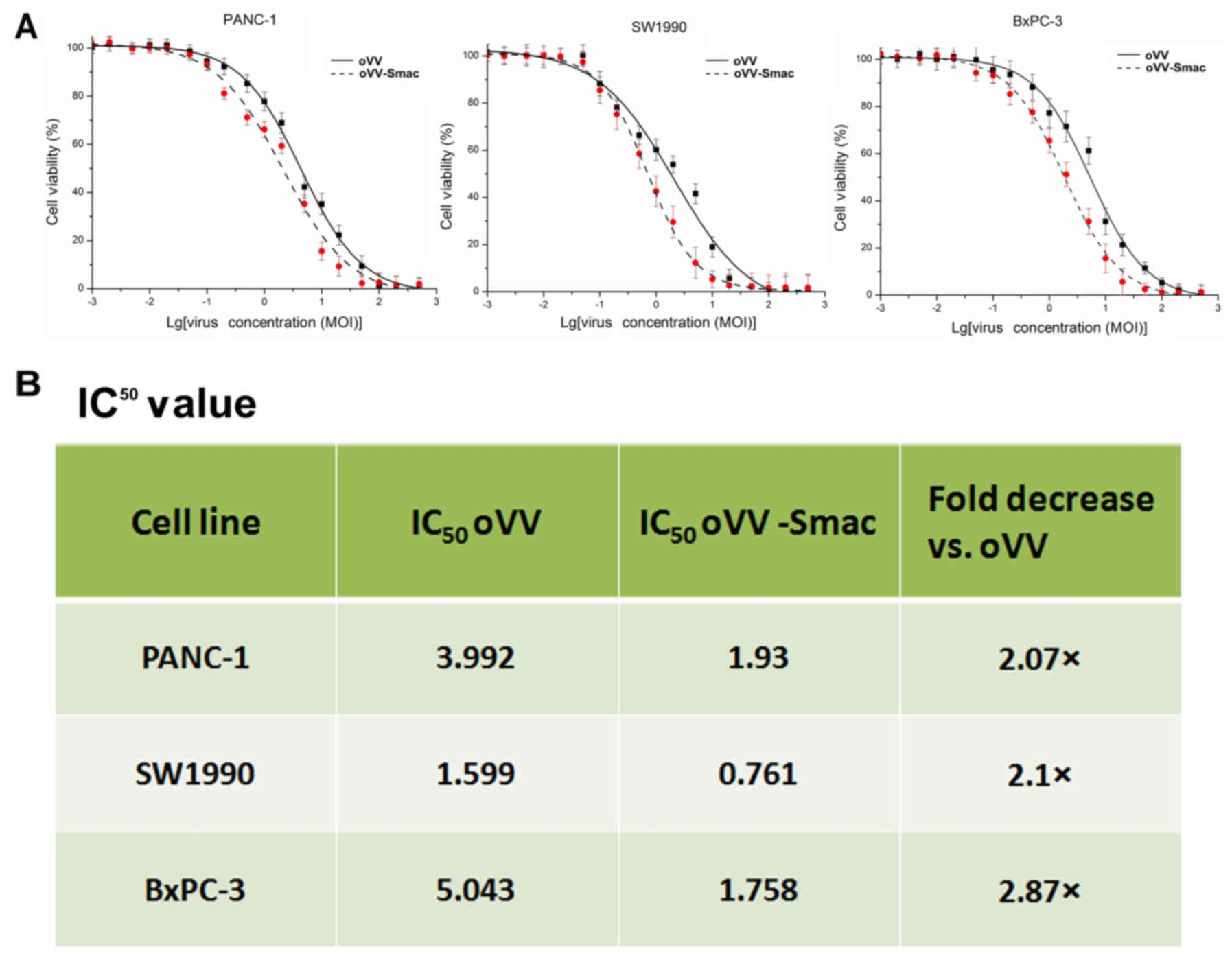
In vitro antitumor activity of
oVV-Smac
The
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay was performed 48 h post-infection to evaluate the
cytotoxicity of oVV-Smac in the PANC-1, SW1990 and BxPC-3
cell lines. The results indicated a significantly higher inhibition
of cell growth with oVV-Smac than with oVV (Fig. 2A and B); these effects were
dose-dependent. Taken together, these findings demonstrated
selective inhibitory effects of oVV-Smac on cancer cell
growth in vitro.
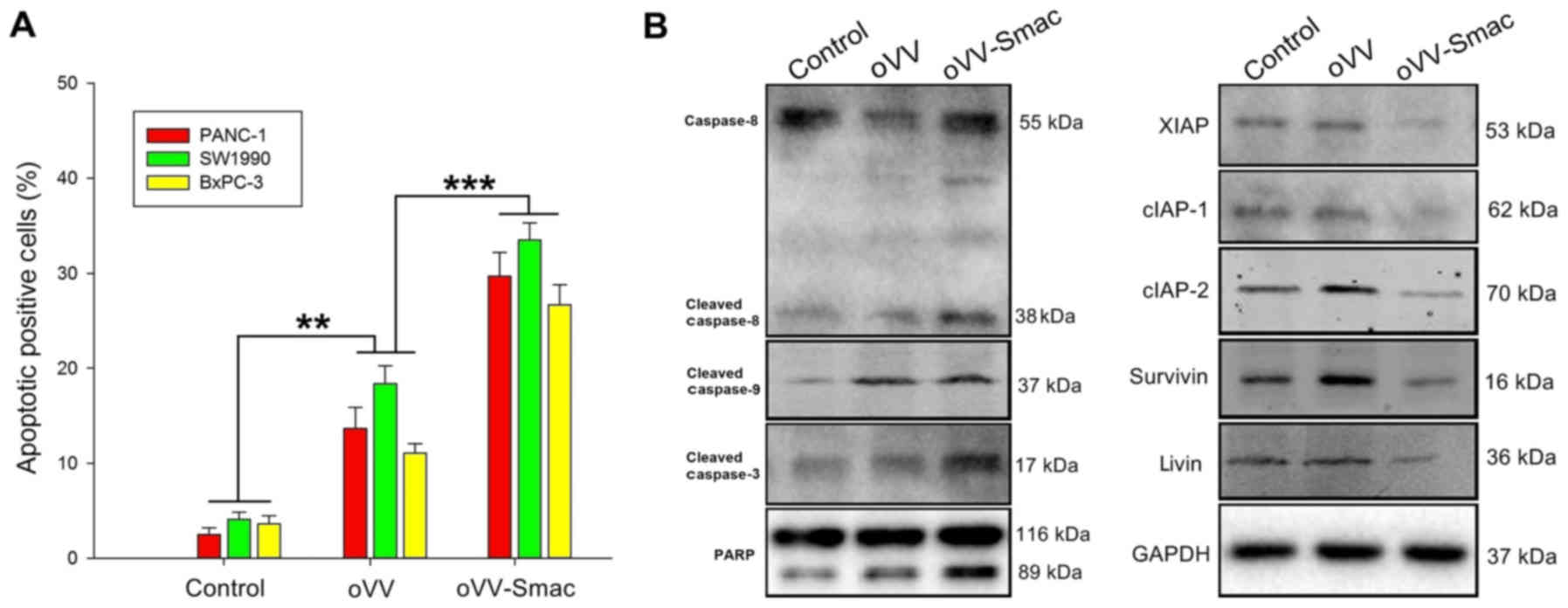
Infection with oVV-Smac induces
apoptosis in vitro
To address the underlying mechanism of
oVV-Smac-induced cytotoxicity, we evaluated
oVV-Smac-associated apoptosis in vitro using flow
cytometric analysis. The results indicated significant apoptosis in
the PANC-1, SW1990 and BxPC-3 cell lines transfected with
oVV-Smac compared with the percentage of apoptosis in the
oVV- or PBS-treated cells (Fig.
3A).
We further evaluated apoptosis by assessing the
expression of apoptosis-related proteins in SW1990 cells at 48 h
post-infection using western blotting analysis. The results
indicated a significant activation of caspase-3, −8 and −9, and
increased poly(ADP-ribose) polymerase (PARP) cleavage in the
oVV-Smac-treated cells (Fig.
3B; left blot). Additionally, the levels of X-linked IAP
(XIAP), cellular IAP-1 (cIAP-1), cIAP-2, survivin and livin were
also decreased in the oVV-Smac cells (Fig. 3B; right blot). Taken together, these
findings indicated that oVV-Smac effectively induced
apoptosis through the caspase and IAP pathways.
Combined treatment with gemcitabine
and oVV-Smac results in synergistic effects
To determine whether oVV-Smac enhances the
cytotoxic effect of gemcitabine, we analyzed the viability of
pancreatic cancer cells using the MTT assay after co-treatment with
oVV-Smac and gemcitabine. The PANC-1, SW1990 and BxPC-3
cells were treated with gemcitabine (1 or 5 µM) with or without
oVV-Smac (0.1 or 0.5 MOI). The combination of
oVV-Smac with gemcitabine significantly inhibited cell
growth compared with treatment with gemcitabine or oVV-Smac
alone (Fig. 4A upper panels). Next,
the synergistic effect of gemcitabine combined with oVV-Smac
on pancreatic cancer cells was quantified using the combination
index (CI) analysis and expressed as CI vs. the fractional affect
(Fig. 4A lower panels). In PANC-1
cells, at all the fractions considered, the Chou-Talalay CI was
lower than one (log10(CI) <0), indicating a
potentiation effect of oVV-Smac when combined with gemcitabine, and
vice versa. Additionally, investigation with SW1990 and BxPC-3
cells presented similar results (log10(CI) <0). These
results showed that the combination of gemcitabine and
oVV-Smac had a synergistic tumor killing effect.
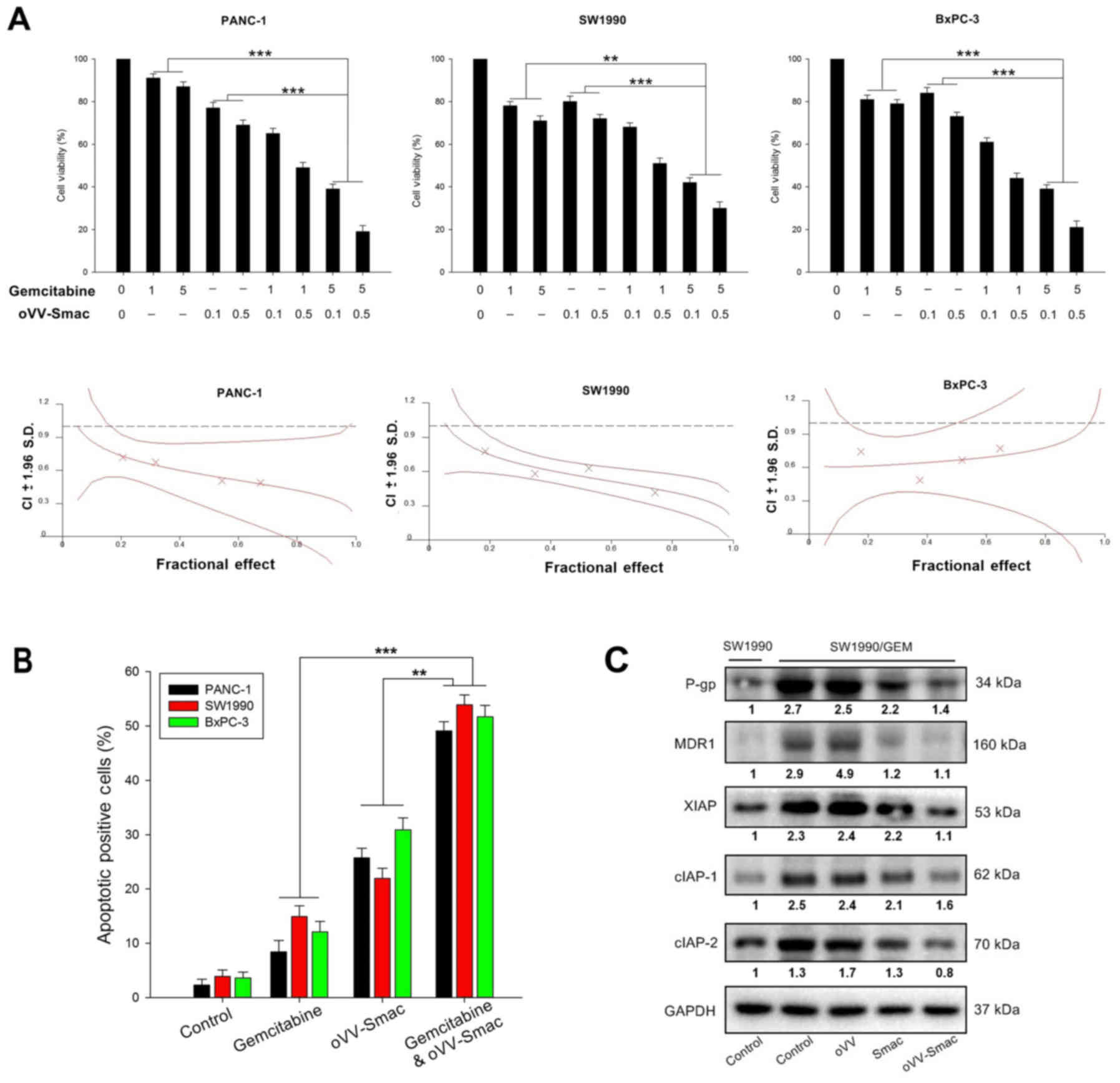 | Figure 4.oVV-Smac enhances
gemcitabine-mediated growth inhibition and apoptosis in pancreatic
cancer cells. (A) Cells were treated with gemcitabine and/or
oVV-Smac for 48 h, and cell viability was determined by MTT assay.
The potential synergistic effect of gemcitabine combined with
oVV-Smac or gemcitabine alone on pancreatic cancer cells. It was
assessed by Chou-Talalay Combination Index (CI) analysis using
CalcuSyn software. The middle curve line stands for the simulated
combination index values, which was expressed as the
log10(CI) ± 1.96 SD, encircled by two lines of algebraic
evaluation of the 95% confidence intervals. The
log10(CI) values attained at the given fractional
effects represent an antagonism between the treatments when >0,
an additive efficiency when equal to 0 and asynergism when <0.
It was quantified by CI analysis and expressed as CI vs. the
fraction affected. Where calculable, 95% confidence intervals are
shown. (B) Gemcitabine (5 µg/ml), oVV-Smac (0.1 MOI), or
gemcitabine (5 µg/ml) plus oVV-Smac (0.1 MOI) was used to treat
PANC-1, SW1990 and BxPC-3 cells. Uninfected cells served as the
control. Forty-eight hours later, apoptosis was determined by flow
cytometry. (C) Gemcitabine-resistant SW1990 cell line (SW1990/GEM)
was used to elucidate the mechanism by which oVV-Smac may influence
gemcitabine-resistant pancreatic cancer. oVV-Smac (10 MOI), oVV (10
MOI), or Smac (5 µg) was used to treat SW1990/GEM cells. Uninfected
cells served as the control. Forty-eight hours later, whole cell
extracts were prepared and immunoblotted. GAPDH was used as a
loading control. (**P<0.01 and ***P<0.001, one-way analysis
of variance (ANOVA) and multiple comparisons). oVV, oncolytic
vaccinia virus; oVV-Smac, Smac-armed oncolytic vaccinia virus. |
We further evaluated apoptosis using
Annexin-V-FITC/PI double staining to evaluate the effect of
oVV-Smac on gemcitabine-induced apoptosis (Fig. 4B). The apoptotic rate in the
pancreatic cancer cells co-treated with gemcitabine and
oVV-Smac was significantly higher than that in the cells
treated with gemcitabine or oVV-Smac alone.
The question remains as to the mechanism by which
oVV-Smac may influence gemcitabine-resistant pancreatic
cancer cell lines. Therefore, we constructed a
gemcitabine-resistant SW1990 cell line (SW1990/GEM). Subsequently,
the expression of multidrug resistance-related proteins and IAPs in
SW1990/GEM cells treated with oVV-Smac, Smac, or oVV was
assessed by western blot analysis. Compared with the sensitive
SW1990 cells, SW1990/GEM cells produced marked multidrug
resistance-related proteins (P-gp and MDR1) and IAPs (XIAP, cIAP-1
and cIAP-2). Based on these results, Smac gene transfection
has the capacity to reduce multidrug resistance-related proteins
and IAPs, which could be further potentiated by Smac-armed
oncolytic vaccinia virus (Fig.
4C).
Taken together, these findings indicated a
synergistic repressive effect of the combined gemcitabine and
oVV-Smac treatment on pancreatic cancer cell
proliferation.
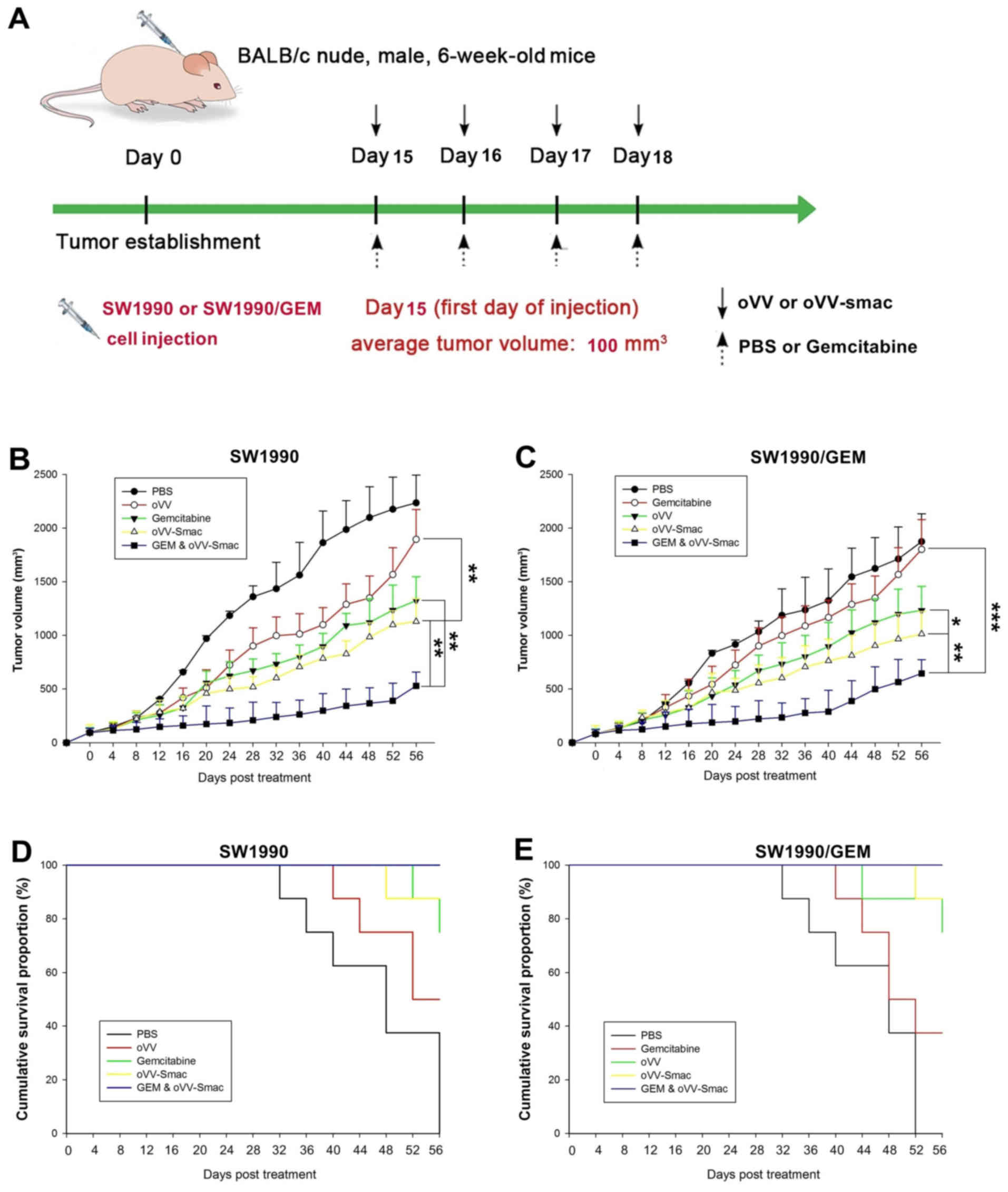
Enhanced cytotoxic effect of
co-treatment with gemcitabine and oVV-Smac in vivo
We developed two pancreatic tumor xenograft mouse
models with sensitive SW1990 cells and SW1990/GEM cells using
BALB/c athymic nude mice to evaluate the effect of co-treatment
with gemcitabine and oVV-Smac in vivo (Fig. 5A). Antitumor efficacy was evaluated
by plotting tumor growth curves over a 56-day observation period.
The mean tumor volume was significantly decreased in mice injected
with gemcitabine, oVV-Smac, and both compared with those
injected with PBS (Fig. 5B and C).
Furthermore, co-treatment of sensitive SW1990 and SW1990/GEM cells
with gemcitabine and oVV-Smac was more effective than
gemcitabine (P=0.001 and 0.002, respectively) and oVV-Smac
alone (P=0.001 and 0.003, respectively). Co-treatment with
gemcitabine and oVV-Smac was also associated with a higher
survival rate when compared with treatment with PBS, gemcitabine,
or oVV-Smac (Fig. 5D and
E).
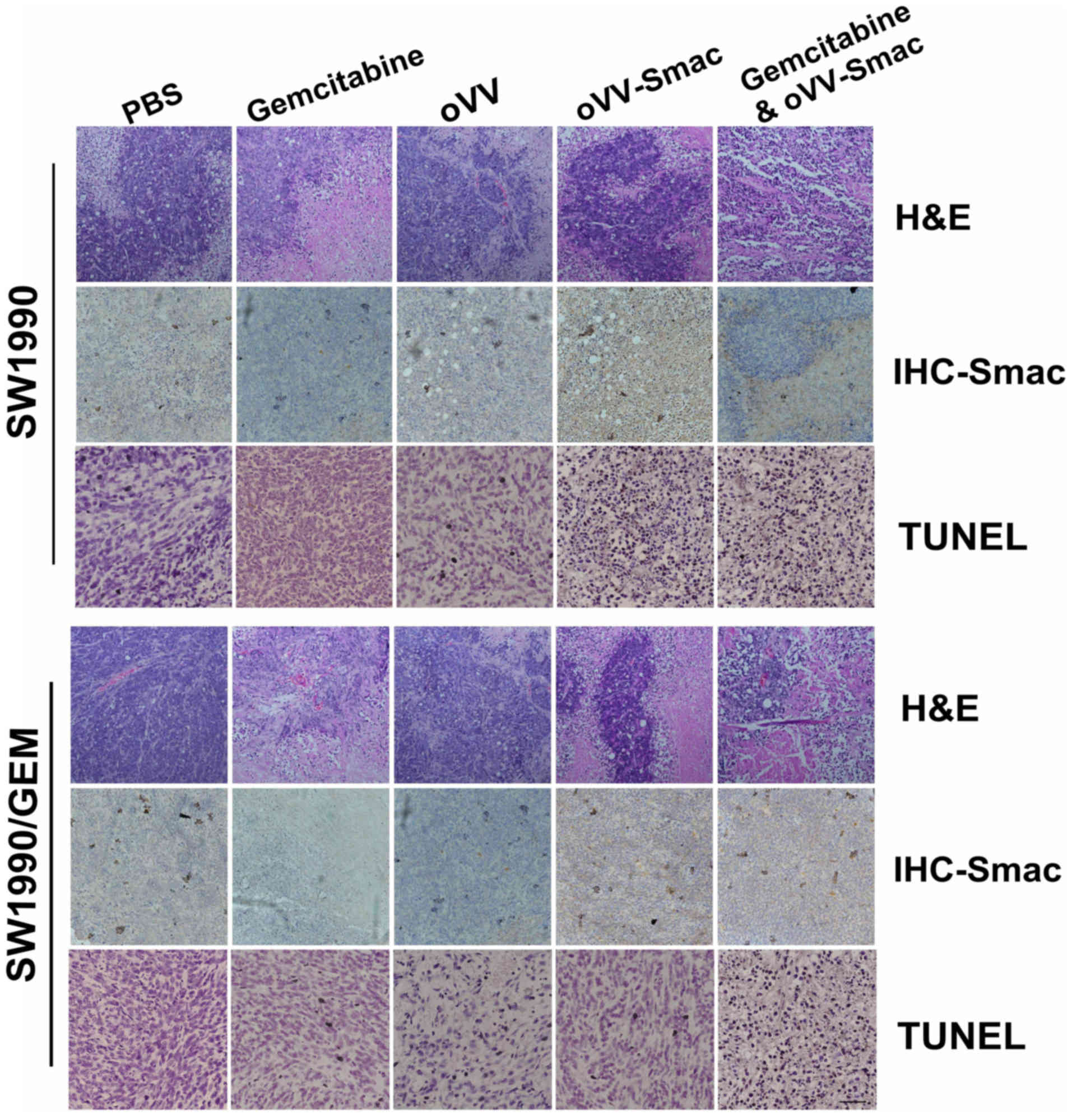
The tumor histopathological changes were further
evaluated by hematoxylin and eosin (H&E) staining,
immunohistochemistry (IHC) and TUNEL assay. The combined treatment
with gemcitabine and oVV-Smac resulted in a higher
cytotoxicity than single treatment as evidenced by H&E
staining. Moreover, an intense expression of Smac in the
tumor tissues was associated with the combined treatment as
evidenced by IHC with anti-Smac (Fig.
6). Apoptosis was further examined using the terminal
deoxynucleotidyl transferase-mediated dUTP nick end labeling
(TUNEL) assay. Results from this experiment showed significantly
higher apoptosis rates in the combination treatment when compared
with either individual treatment (Fig.
6).
Discussion
Considering the genetic diversity of most tumors and
the development of drug-resistance, monotherapy has been associated
with limited success in various types of cancer (26). Therefore, the identification of
therapeutic agents that act synergistically when combined through
different mechanisms without inducing adverse effects is critical
in cancer therapy (27). Many
cancer cells express elevated levels of inhibitor of apoptosis
proteins (IAPs) and escape apoptosis through the activity of IAPs
(28). IAPs prevent the activation
of caspases and, as such, block the extrinsic and intrinsic
apoptotic cascades. XIAP is one of the best characterized IAPs and
is expressed at a higher level in pancreatic cancer cell lines and
pancreatic tumors compared with normal pancreas (29). The mitochondrial protein Smac
inhibits IAPs, including XIAP, thus promoting caspase activation
and subsequent cell death. Smac has been shown to bind to XIAP,
cIAP-1 and cIAP-2, and Smac mimetics sensitize tumors to programmed
cell death. In the present study, Smac exhibited low expression in
our pancreatic cancer cell lines (PANC-1, SW1990 and BxPC-3), which
was confirmed in our previous study (21). In addition, Smac is expressed at a
lower level in pancreatic tumors (10 of 10) compared with normal
pancreas (data not shown). Several clinical studies have reported
synergistic antitumor activity of the co-treatment with oVV and
chemotherapy or radiation therapy (30,31).
Additionally, it has been suggested that the route of combination
of oVV with cytotoxic chemotherapies is another factor in
potentiating the effects of oVV and optimizing the therapy
(32).
Pancreatic cancer is invariably aggressive with high
mortality rates, while the acquired drug resistance makes it
intransigent (33). Thus, new
effective therapeutic approaches are urgently required. Gene
therapy has now become an innovative approach for cancer treatment.
Thus, we developed and evaluated the effect of oVV-based gene
therapy in vitro and in vivo. The results indicated
significant growth inhibition with relatively limited side
effects.
Interestingly, the mitochondrial protein Smac is
released into the cytosol during the process of apoptosis.
Recently, Smac has been reported to exhibit significant cytotoxic
effects on different tumor types (34). In addition, Smac was found to
potentiate the sensitivity to chemotherapeutic drugs in patients
with cancer, indicating that the targeted delivery of Smac may be
an auspicious gene therapy in pancreatic cancer (35). However, no studies have reported on
oVV-mediated Smac gene therapy in pancreatic cancer. We
developed a novel oVV (TK deletion) that expresses Smac
(oVV-Smac), and assessed its antitumor effect when applied
alone or combined with gemcitabine.
Based on our results, oVV-Smac potentiated
apoptosis in human pancreatic cancer cells. The IAPs bind to
caspases and inhibit their activity through their baculovirus IAP
repeat domains (36). Furthermore,
IAPs may ubiquitinate themselves and their interacting proteins
through the ubiquitin-protein isopeptide ligase activity of their
Really interesting new gene (RING) finger domain (37). On the other hand, Smac serves as a
key molecule in reducing the protein levels of XIAP, c-IAPs,
survivin and livin both in vitro and in vivo through
the ubiquitin/proteasome pathway. In addition, Smac is implicated
in mitochondrial apoptosis pathways and promotes
chemotherapy-induced apoptosis (38,39).
Our present findings demonstrated a significant reduction of IAPs
and activation of caspase-3, −8, −9 and PARP in SW1990 pancreatic
cancer cells transfected with oVV-Smac. These results
suggested the implication of caspase-dependent apoptosis and IAPs
in oVV-Smac-induced cytotoxicity.
Our results also demonstrated synergistic cytotoxic
effects of co-treatment with oVV-Smac and gemcitabine both
in vitro and in vivo. Several mechanisms may account
for this synergistic effect (40).
First, gemcitabine, which is a first-line intervention in
pancreatic cancer, bears the limitation of weak penetration into
the tumor parenchyma. In contrast, oVV-Smac selectively
replicates in tumor cells, thus resulting in their lysis, which
disrupts the tumor's architecture and facilitates the penetration
of gemcitabine. Thus, gemcitabine combined with oVV-Smac
result in a synergistic effect. Second, gemcitabine was found to
enhance vaccine efficacy by eliminating
CD11b+/Gr-1+ myeloid-derived suppressor cells
(MDSCs) in a murine model of pancreatic carcinoma, which may be
another mechanism for the synergy (41,42).
Third, suboptimal doses of gemcitabine have been reported to
stimulate the viral uptake in pancreatic cancer cells, which may
explain the observed synergistic effects. Fourth, both
oVV-Smac and gemcitabine induce apoptosis, which may explain
their synergistic effects (43).
Finally, unblocking host pathways, transporting viruses with
greater efficiency, and/or increasing viral replication at the
tumor site may also account for the synergistic effects of
oVV-Smac and gemcitabine when combined.
To the best of our knowledge, this is the first
report concerning the cytotoxic effects of oVV-Smac on
pancreatic cancer cells and the synergistic effects of co-treatment
with oVV-Smac and gemcitabine. Undoubtedly, the optimization
of oVV-Smac application in clinical practice is warranted,
particularly in relation to increasing the delivery and expression
of Smac, viral vector cytotoxicity, and immune response to
viral antigens. Thus, additional research aimed at promoting the
transfection efficiency of oVV-Smac and reducing its
potential toxicity is required.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Science Foundation of China (nos. 81602706 and 81570198), the Funds
of Science Technology Department of Zhejiang Province (no.
2017C33116), the Zhejiang Medical Technology Plan Project (no.
2016KYA018) and State Administration of Traditional Chinese
Medicine of Zhejiang (no. 2017ZB006).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
SW, GR and XM conceived and designed the study. WC,
WF, XZ, FH, XL and SW performed the experiments. SW, WC and GR
wrote the paper. SW, XL, XZ and WC reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All animal experiments were approved by the
Institutional Animal Care and Use Committee, Zhejiang Provincial
People's Hospital and all procedures were in according to the Guide
for the Care and Use of Laboratory Animals (National Academies
Press, Washington, D.C.).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ryan DP, Hong TS and Bardeesy N:
Pancreatic adenocarcinoma. N Engl J Med. 371:1039–1049. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bednar F and Simeone DM: Recent advances
in pancreatic surgery. Current Opin Gastroenterol. 30:518–523.
2014. View Article : Google Scholar
|
|
3
|
Ghaneh P, Smith R, Tudor-Smith C, Raraty M
and Neoptolemos JP: Neoadjuvant and adjuvant strategies for
pancreatic cancer. Eur J Surg Oncol. 34:297–305. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vaccaro V, Sperduti I and Milella M:
FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N
Engl J Med. 365:768–769. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kristensen A, Vagnildhaug OM, Gronberg BH,
Kaasa S, Laird B and Solheim TS: Does chemotherapy improve
health-related quality of life in advanced pancreatic cancer? A
systematic review. Crit Rev Oncol Hematol. 99:286–298. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sultana A, Smith CT, Cunningham D,
Starling N, Neoptolemos JP and Ghaneh P: Meta-analyses of
chemotherapy for locally advanced and metastatic pancreatic cancer.
J Clin Oncol. 25:2607–2615. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kirn DH and Thorne SH: Targeted and armed
oncolytic poxviruses: A novel multi-mechanistic therapeutic class
for cancer. Nat Rev Cancer. 9:64–71. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaufman HL, Deraffele G, Mitcham J,
Moroziewicz D, Cohen SM, Hurst-Wicker KS, Cheung K, Lee DS, Divito
J, Voulo M, et al: Targeting the local tumor microenvironment with
vaccinia virus expressing B7.1 for the treatment of melanoma. J
Clin Invest. 115:1903–1912. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chard LS, Lemoine NR and Wang Y: New role
of Interleukin-10 in enhancing the antitumor efficacy of oncolytic
vaccinia virus for treatment of pancreatic cancer. Oncoimmunology.
4:e10386892015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Z, Ravindranathan R, Li J, Kalinski P,
Guo ZS and Bartlett DL: CXCL11-armed oncolytic poxvirus elicits
potent antitumor immunity and shows enhanced therapeutic efficacy.
Oncoimmunology. 5:e10915542015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Heo J, Reid T, Ruo L, Breitbach CJ, Rose
S, Bloomston M, Cho M, Lim HY, Chung HC, Kim CW, et al: Randomized
dose-finding clinical trial of oncolytic immunotherapeutic vaccinia
JX-594 in liver cancer. Nat Med. 19:329–336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Breitbach CJ, Thorne SH, Bell JC and Kirn
DH: Targeted and armed oncolytic poxviruses for cancer: The lead
example of JX-594. Curr Pharm Biotechnol. 13:1768–1772. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deng L, Fan J, Ding Y, Zhang J, Zhou B,
Zhang Y and Huang B: Oncolytic efficacy of thymidine kinase-deleted
vaccinia virus strain Guang9. Oncotarget. 8:40533–40543.
2017.PubMed/NCBI
|
|
14
|
Zhang Q, Yu YA, Wang E, Chen N, Danner RL,
Munson PJ, Marincola FM and Szalay AA: Eradication of solid human
breast tumors in nude mice with an intravenously injected
light-emitting oncolytic vaccinia virus. Cancer Res.
67:10038–10046. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park SH, Breitbach CJ, Lee J, Park JO, Lim
HY, Kang WK, Moon A, Mun JH, Sommermann EM, Maruri Avidal L, et al:
Phase 1b trial of biweekly intravenous Pexa-Vec (JX-594), an
oncolytic and immunotherapeutic vaccinia virus in colorectal
cancer. Mol Ther. 23:1532–1540. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kochneva G, Zonov E, Grazhdantseva A,
Yunusova A, Sibolobova G, Popov E, Taranov O, Netesov S, Chumakov P
and Ryabchikova E: Apoptin enhances the oncolytic properties of
vaccinia virus and modifies mechanisms of tumor regression.
Oncotarget. 5:11269–11282. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Timiryasova TM, Li J, Chen B, Chong D,
Langridge WH, Gridley DS and Fodor I: Antitumor effect of vaccinia
virus in glioma model. Oncol Res. 11:133–144. 1999.PubMed/NCBI
|
|
18
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chauhan D and Anderson KC: Mechanisms of
cell death and survival in multiple myeloma (MM): Therapeutic
implications. Apoptosis. 8:337–343. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chauhan D, Neri P, Velankar M, Podar K,
Hideshima T, Fulciniti M, Tassone P, Raje N, Mitsiades C, Mitsiades
N, et al: Targeting mitochondrial factor Smac/DIABLO as therapy for
multiple myeloma (MM). Blood. 109:1220–1227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ge Y, Lei W, Ma Y, Wang Y, Wei B, Chen X,
Ru G, He X, Mou X and Wang S: Synergistic antitumor effects of CDK
inhibitor SNS032 and an oncolytic adenovirus co-expressing TRAIL
and Smac in pancreatic cancer. Mol Med Rep. 15:3521–3528. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pan Q, Huang Y, Chen L, Gu J and Zhou X:
SMAC-armed vaccinia virus induces both apoptosis and necroptosis
and synergizes the efficiency of vinblastine in HCC. Human Cell.
27:162–171. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang S, Shu J, Chen L, Chen X, Zhao J, Li
S, Mou X and Tong X: Synergistic suppression effect on tumor growth
of ovarian cancer by combining cisplatin with a manganese
superoxide dismutase-armed oncolytic adenovirus. Onco Targets Ther.
9:6381–6388. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lei W, Wang S, Yang C, Huang X, Chen Z, He
W, Shen J, Liu X and Qian W: Combined expression of miR-34a and
Smac mediated by oncolytic vaccinia virus synergistically promote
anti-tumor effects in Multiple Myeloma. Sci Rep. 6:321742016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Buller RM, Smith GL, Cremer K, Notkins AL
and Moss B: Decreased virulence of recombinant vaccinia virus
expression vectors is associated with a thymidine kinase-negative
phenotype. Nature. 317:813–815. 1895. View Article : Google Scholar
|
|
26
|
Levinson AD: Cancer therapy reform.
Science. 328:1372010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kaiser J: Combining targeted drugs to stop
resistant tumors. Science. 331:1542–1545. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Salvesen GS and Duckett CS: IAP proteins:
Blocking the road to death's door. Nat Rev Mol Cell Biol.
3:401–410. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lopes RB, Gangeswaran R, Mcneish IA, Wang
Y and Lemoine NR: Expression of the IAP protein family is
dysregulated in pancreatic cancer cells and is important for
resistance to chemotherapy. Int J Cancer. 120:2344–2352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wilkinson MJ, Smith HG, McEntee G,
Kyula-Currie J, Pencavel TD, Mansfield DC, Khan AA, Roulstone V,
Hayes AJ and Harrington KJ: Oncolytic vaccinia virus combined with
radiotherapy induces apoptotic cell death in sarcoma cells by
down-regulating the inhibitors of apoptosis. Oncotarget.
7:81208–81222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu Y, Mou X, Wang S, Liu XE and Sun X:
ING4 expressing oncolytic vaccinia virus promotes anti-tumor
efficiency and synergizes with gemcitabine in pancreatic cancer.
Oncotarget. 8:82782–82739. 2017.
|
|
32
|
Liu Z, Ravindranathan R, Kalinski P, Guo
ZS and Bartlett DL: Rational combination of oncolytic vaccinia
virus and PD-L1 blockade works synergistically to enhance
therapeutic efficacy. Nat Commun. 8:147542017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martinez-Ruiz G, Maldonado V,
Ceballos-Cancino G, Grajeda JP and Melendez-Zajgla J: Role of
Smac/DIABLO in cancer progression. J Exp Clin Cancer Res. 27:1–7.
2008. View Article : Google Scholar :
|
|
35
|
Fulda S, Wick W, Weller M and Debatin KM:
Smac agonists sensitize for Apo2L/TRAIL- or anticancer drug-induced
apoptosis and induce regression of malignant glioma in vivo. Nat
Med. 8:808–815. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shiozaki EN and Shi Y: Caspases, IAPs and
Smac/DIABLO: Mechanisms from structural biology. Trends Biochem
Sci. 29:486–494. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bader M and Steller H: Regulation of cell
death by the ubiquitin-proteasome system. Curr Opin Cell Biol.
21:878–884. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xu Y, Zhou L, Huang J, Liu F, Yu J, Zhan
Q, Zhang L and Zhao X: Role of Smac in determining the
chemotherapeutic response of esophageal squamous cell carcinoma.
Clin Cancer Res. 17:5412–5422. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tirrò E, Consoli ML, Massimino M, Manzella
L, Frasca F, Sciacca L, Vicari L, Stassi G, Messina L, Messina A
and Vigneri P: Altered expression of c-IAP1, survivin, and Smac
contributes to chemotherapy resistance in thyroid cancer cells.
Cancer Res. 66:4263–4272. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Metwalli AR, Khanbolooki S, Jinesh G,
Sundi D, Shah JB, Shrader M, Choi W, Lashinger LM, Chunduru S,
Mcconkey DJ, et al: Smac mimetic reverses resistance to TRAIL and
chemotherapy in human urothelial cancer cells. Cancer Biol Ther.
10:885–892. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ishizaki H, Manuel ER, Song GY, Srivastava
T, Sun S, Diamond DJ and Ellenhorn JD: Modified vaccinia Ankara
expressing survivin combined with gemcitabine generates specific
antitumor effects in a murine pancreatic carcinoma model. Cancer
Immunol Immunother. 60:99–109. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Al Yaghchi C, Zhang Z, Alusi G, Lemoine NR
and Wang Y: Vaccinia virus, a promising new therapeutic agent for
pancreatic cancer. Immunotherapy. 7:1249–1258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nguyen TL, Tumilasci VF, Singhroy D,
Arguello M and Hiscott J: The emergence of combinatorial strategies
in the development of RNA oncolytic virus therapies. Cell
Microbiol. 11:889–897. 2009. View Article : Google Scholar : PubMed/NCBI
|