Introduction
For the past few years, lung cancer has gradually
become one of the most common solid and malignant tumours that
threaten the health of human beings. There are 1.8 million new
patients each year that suffer from lung cancer (1). Approximately 80% of lung cancer cases
are non-small cell lung cancer (NSCLC), which consists of lung
adenocarcinoma (LAD), squamous cell carcinoma, large cell lung
cancer and adenosquamous cell carcinoma (2). Since early symptoms are not obvious,
early diagnosis is more difficult. Most lung cancer patients have
lost their opportunity to undergo surgery by the time they are
diagnosed, making chemotherapy the main treatment for NSCLC
(3). Docetaxel is a semi-synthetic
taxane antitumour agent that is utilized extensively in the
clinical therapy of advanced NSCLC. Mechanistically, it induces
abnormalities in microtubule dynamics by promoting microtubule
polymerization. Abnormal microtubule dynamics activate spindle
assembly checkpoint proteins to cause cell cycle arrest (4). In addition, taxanes can affect
mitochondrial membrane permeability, resulting in the production
and release of pro-apoptotic substances and the induction of
apoptosis (5). Nonetheless,
chemoresistance is the most important reason for the failure of
docetaxel treatment (6).
The development of docetaxel resistance in tumour
cells is a multi-step, multi-factor process involving the genomic
and epigenetic abnormalities of key genes related to drug efflux,
signal transduction, DNA damage repair, the cell cycle and
apoptosis (7). Based on data from
previous studies, our laboratory has confirmed that Aurora A
promotes the resistance of primary hepatocarcinoma cells by
regulating the NF-κB/miRNA-21/PTEN signalling pathway (8). Aurora A is a member of the mitotic
serine/threonine kinase family that is encoded by the AURKA gene.
Aurora A plays important roles in mitosis, which is associated with
central body maturation and separation, spindle assembly and
stability, and chromosome pairing (9). The expression and activation peak of
Aurora A appears in the late stage of cell mitosis during the G2 to
M phase transition, and its localization changes with cell cycle
evolution (10). In NSCLC, Aurora A
expression is upregulated, causing cell aneuploidy formation and
promoting malignant transformation (11). Aurora A gene expression
abnormalities are also very common in the formation of
drug-resistant phenotypes, but the role Aurora A plays remains
unclear in the process of docetaxel resistance in human lung
adenocarcinoma.
MicroRNAs (miRNAs) are small non-coding RNA
molecules that can be found in plants, animals and some viruses.
miRNAs contain approximately 22 non-coding nucleotides that bind to
the 3′-untranslated region (3′-UTR) of the target gene mRNA and
inhibit its expression. miRNAs play an important role in epigenetic
regulation. miRNAs have a small number of bases that bind to the
site of its target genes, and complete pairing is not required;
thus, one miRNA can target multiple genes, and the target genes
also have multiple miRNA binding sites that result in a complex
regulatory network. miRNAs play an important role in cell
proliferation and apoptosis, blood cell differentiation, insulin
secretion, late embryonic development, tumour development and many
other important physiological and pathological processes. Current
studies have demonstrated that miRNA expression abnormalities are
very common in the development of drug-resistance in tumour cells,
and the possible mechanisms include the abnormal modulation of cell
proliferation, apoptosis, the cell cycle and signalling pathways.
miRNA-mediated chemotherapy resistance is one of the key research
areas of chemotherapy resistance.
According to high throughput miRNA chip data from
our previous research, miRNA-885-3p (miR-885-3p) was identified as
one of the most downregulated miRNAs in docetaxel-resistant
SPC-A1/DTX cells (human lung adenocarcinoma) compared with those in
parental SPC-A1 cells (12). Since
miR-885-3p is involved in cell apoptosis in human cancer cells
(13,14), we hypothesized that miR-885-3p
downregulation may be related to chemoresistance in SPC-A1/DTX
cells. In the present study, we explored the function of miR-885-3p
and Aurora A in chemoresistance in human LAD cells. Our results
indicated that miR-885-3p could act as a chemosensitizer to
docetaxel in human LAD cells by targeting Aurora A.
Materials and methods
Cell culture
Human lung adenocarcinoma cell lines (SPC-A1 and
NCI-H1299) were purchased from the Tumor Cell Bank of the Chinese
Academy of Medical Sciences (Shanghai, China). The
docetaxel-resistant SPC-A1 and NCI-H1299 cell lines (SPC-A1/DTX and
H1299/DTX) were established and maintained in our laboratory. All
the cell lines were cultured in RPMI-1640 medium supplemented with
10% fetal bovine serum (FBS) and ampicillin (100 U/ml)-streptomycin
(100 µg/ml) at 37°C and 5% CO2.
Plasmids, shRNA, miRNA mimics and
miRNA inhibitors
miR-885-3p mimics and inhibitors (anti-miR-885-3p)
were synthesized by Shanghai GenePharma Co., Ltd. (Shanghai,
China), as well as the negative controls (miR-NC mimics or
anti-miR-NC). Short hairpin RNAs (shRNAs) specifically targeting
the human Aurora A gene (GenBank no. NM_003600) were designed to
knock down Aurora A expression. The shRNA sequences targeting
Aurora A were as follows: Aurora A-shRNA sense,
5′-GATCCATGCCCTGTCTTAACTGTCATTCAAGAGATGACAGTAAGACAGGGCATAGA-3′;
negative control (NC) shRNA sense,
5′-GATCCAAGCTGAAGTACAACCTTCTTCAAGAGAGAAGGTTGTACTTCAGCTTAGA-3′. The
aforementioned sequences were inserted into the pSilencer4.1-CMVneo
vector (between the BglII(A-GATCT) and
HindIII(A-AGCTT) enzyme sites). The constructed plasmids
were named pSil/shAuro and pSil/NC, respectively. All newly
constructed plasmids were confirmed by DNA sequencing. A plasmid
vector (pMD18/Auro) expressing Aurora A was purchased from Sino
Biological, Inc. (Beijing, China). Lipofectamine PLUS (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) was used for cell
transfection. Ampicillin-streptomycin-free medium was used for
transfection. All the procedures were carried out according to the
manufacturer's protocols. Stably transfected cell lines were
selected with G418 (400 µg/ml) 48 h after transfection, and
individual clones were isolated and maintained in a medium
containing G418 (100 µg/ml).
RNA extraction and real-time
quantitative RT-PCR (RT-qPCR) assay
Total RNA was extracted from cultured cells using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. miRNA expression was determined by
stem-loop reverse-transcription (RT) and real-time quantitative
PCR. The primers were designed as follows: miR-885-3p,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTATCCA-3′; and U6,
5′-CGCTTCACGAATTTGCGTGTCA-3′. RT was performed using a PrimeScript
RT reagent kit (Takara Bui Inc., Otsu, Japan) according to the
manufacturer's instructions. Reverse transcriptase reactions were
performed in a Mastercycler Thermocycler (Eppendorf AG, Hamburg,
Germany) at 16 °C for 30 min, 42 °C for 30 min and 85 °C for 5 min.
The qRT-PCR primers were designed as follows: Aurora A sense,
5′-AATGCCCTGTCTTACTGTCATTC-3′ and antisense,
5′-TCCAGAGATCCACCTTC-TCATC-3′; GAPDH sense,
5′-GACTCATGACCACAGTCCATGC-3′ and antisense,
5′-AGAGGCAGGGATGATGTTCTG-3′; miR-885-3p sense,
5′-CGTTAGGCAGCGGGGTGTAG-3′, and antisense,
5′-ATCCAGTGCAGGGTCCGAGG-3′; U6 sense,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and antisense,
5′-CGCTTCACGAATTTGCGTGTCAT-3′. Real-time PCR was performed using a
MicroRNA assay kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The reaction
conditions were 50°C for 2 min, 95°C for 10 min, 40 cycles of 95°C
for 15 sec and 60°C for 1 min. Reactions were performed in
triplicate using an ABI StepOne RT-PCR system.
In vitro chemosensitivity assay
Single-cell suspensions were prepared and seeded in
96-well plates (2×103 cells/well) and cultured
overnight. After incubation for 48 h with freshly prepared
docetaxel, a
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide
(MTT; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) solution (0.5
mg/ml) was added. After incubation for 4 h, the culture medium was
removed and replaced with 100 µl of DMSO in each well. The
absorbance at 490 nm was measured using a microplate reader (model
680; Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
concentration gradient was designed to cover the killing efficiency
of the docetaxel against SPC-A1 or H1299 cells from 0 to 100%: 1,
2, 4, 8, 16, 32, 64 and 128 µg/l for the parental strains. For the
drug-resistant strains, docetaxel was designed to have a
concentration gradient of 4, 8, 16, 32, 64, 128, 256 and 512 µg/l.
The cell survival curve was plotted using GraphPad Prism 5
(GraphPad Software, Inc., La Jolla, CA, USA) and the
IC50 values were calculated. The relative survival rate
was calculated as: Relative survival rate = (OD experiment group -
OD blank control group)/(OD negative control group - OD blank
control group) × 100%. The blank control group referred to the
control well containing the culture medium only, and the negative
control group contained the cells and the culture solution without
the addition of docetaxel. All assays were performed in
sextuplicate and repeated at least 3 times.
Colony formation assay
Cells were trypsinized into single cell suspensions
and added to 6-well plates at 500 cells/well. Following 14 days of
culture, the RPMI-1640 medium was removed, and the colonies were
fixed with methanol. All the cell colonies were then stained with a
0.1% crystal violet solution and counted manually. Each experiment
was performed in triplicate.
Apoptosis analysis
An Annexin V-fluorescein isothiocyanate (FITC)
apoptosis detection kit (KeyGen Biotech, Co., Ltd., Nanjing, China)
was used to detect apoptosis in cancer cells according to the
manufacturer's instructions. All the samples were assayed in
triplicate.
Cell cycle analysis
Cells were collected and washed with ice-cold
phosphate-buffered saline (PBS), and then fixed in 70% ethanol
overnight at −20°C. The fixed cells were rehydrated using PBS for
10 min and subjected to PI/RNase staining, followed by flow
cytometric analysis using a FACScan flow cytometer and CellQuest
software (both from BD Biosciences, San Jose, CA, USA).
Western blotting assay
Cells were harvested by suspension in lysis buffer
(1 mM dithiothreitol, 0.125 mM EDTA, 5% glycerol, 1.0 mM
phenylmethylsulfonylfluoride, 1.0 mg/ml leupeptin, 1.0 mg/ml
pepstatin, 1.0 mg/ml aprotinin, 1% Triton X-100 in 12.5 mM TRIS-HCl
buffer, pH 7.0) on ice. The protein concentration was measured
using Pierce™ BCA Protein Assay kit (Thermo Fisher Scientific,
Inc., Rockford, IL, USA). Then, equal weights (50 µg) of cell
protein lysates were separated on 10% SDS polyacrylamide gels,
followed by being electrophoretically transferred onto polyvinyl
difluoride membranes (PVDF) (Roche Diagnostics, Basel,
Switzerland). Next, the membranes were blocked in PBS containing 5%
non-fat milk and 0.2% Tween-20 for 1 h at room temperature and
incubated overnight at 4°C rabbit anti-Aurora A (dilution 1:100;
cat. no. 4718T), p-Aurora A (dilution 1:100; cat. no. 3079T),
cleaved caspase-3 (dilution 1:200; cat. no. 9654S), NF-κB (dilution
1:200; cat. no. 4764T), Bcl-2 (dilution 1:50; cat. no. 4223T), Bax
(dilution 1:50; cat. no. 5023T), E-cadherin (dilution 1:150; cat.
no. 3195T) and vimentin (dilution 1:150; cat. no. 5741T) proteins
in PBS containing 0.1% Tween-20 followed by incubation with
horseradish peroxidase-conjugated anti-mouse IgG (dilution 1:1,000;
cat. no. 58802S) at 37°C for 1 h. An anti-GAPDH monoclonal antibody
(dilution 1:500; cat. no. 97166T) was used as a control. ECL
detection reagents (EMD Millipore, Billerica, MA, USA) were added
on the membranes for 1 min and were immediately exposed to X-ray
film (Kodak, Rochester, NY, USA). ImageJ analysis software
(https://imagej.nih.gov/ij/) was used to
quantify the band intensities. All antibodies were purchased from
Univ-bio Inc (Shanghai, China).
Luciferase activity
Human docetaxel-resistant LAD cells (SPC-A1/DTX)
grown in a 48-well plate were co-transfected with miRNA
mimics/inhibitors and pLUC firefly luciferase vectors containing
empty, wild-type or mutant Aurora A 3′-UTR sequences using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
The cells were collected and lysed at 48 h post-transfection to
assess the luciferase activities using a dual-luciferase assay kit
(Promega Corporation, Madison, WI, USA). The relative luciferase
activities were calculated by normalizing the ratio of
Firefly/Renilla luciferase to that of the negative
control-transfected cells.
Wound healing assay
Wound healing assays were used to assess cell
migration. In brief, cells were seeded in 12-well plates and
cultured to confluence. Wounds of 1.0-mm width were created with a
plastic scriber, and the cells were washed with PBS and cultured in
serum-free medium. At 24 h after wounding, cell migration was
observed under a light microscope. The distance of cell migration
under five randomly chosen areas was measured to determine the
wound area.
Transwell assay
Cells in serum-free medium were seeded into inserts
(800 cells/each insert, pore size 8 µm; Corning Inc., Corning, NY,
USA), which were then transferred into wells with medium containing
10% FBS and cultured for an additional 24 h. Thereafter, the
non-invading cells on the top of the membrane were scraped, and the
invaded cells on the bottom of the membrane were fixed in methanol
and stained with a 0.05% crystal violet solution. The number of
invaded cells on the membrane was then counted manually under an
inverted microscope. Each experiment was performed in
triplicate.
Bioinformatics and statistical
analyses
Online miRNA databases (TargetScan, miRBase, and
PicTarget) were used to predict the target genes of miR-885-3p.
Statistical analyses were conducted using SPSS software v16.0
(SPSS, Inc., Chicago, IL, USA). The experimental data were
expressed as the mean ± SEM. For the comparison of means between
two groups, a two-tailed t-test was conducted, and for the
comparison of means among three or more groups, one-way ANOVA and
LSD were used. Differences were considered significant when
P<0.05.
Results
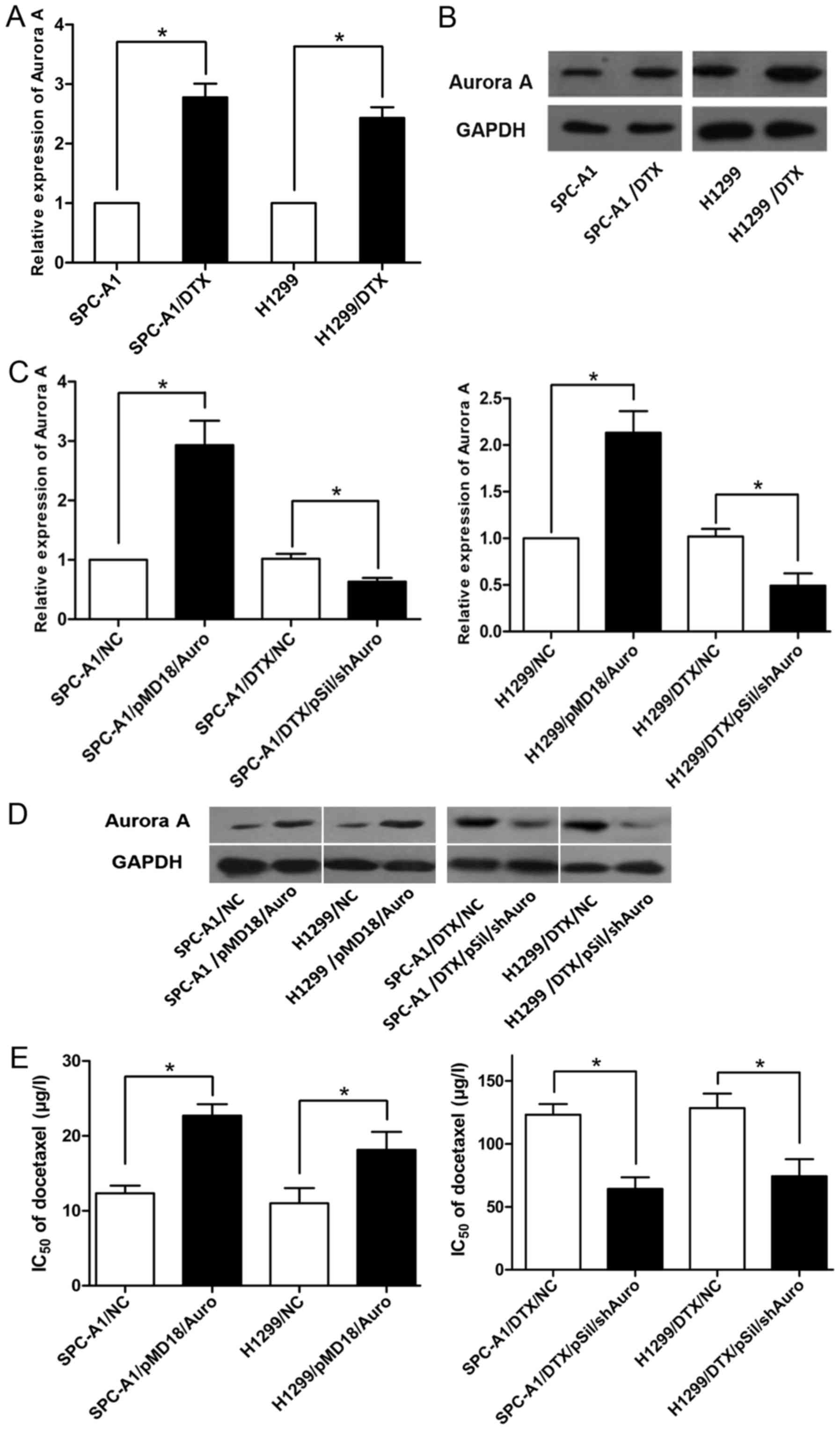
Aurora A is significantly upregulated
in docetaxel-resistant LAD cells
To detect the role of Aurora A in lung
adenocarcinoma resistance, we first analysed the expression of
Aurora A in docetaxel-resistant LAD cells by qRT-PCR
(P<0.05, Fig. 1A).
Compared with that in the parental human lung adenocarcinoma SPC-A1
and NCI-H1299 cell lines, Aurora A was significantly upregulated by
(2.78±0.16)-fold and (2.43±0.12)-fold in the docetaxel-resistant
LAD cell lines SPC-A1/DTX and H1299/DTX. Additionally, Aurora A
protein levels were significantly higher in SPC-A1/DTX and
H1299/DTX cells than in parental SPC-A1 and H1299 cells
(P<0.05, Fig. 1B).
Therefore, these data indicated that the upregulation of Aurora A
may play a role in the development of docetaxel resistance in LAD
cells.
Expression of Aurora A is positively
correlated with the resistance of LAD cells to docetaxel
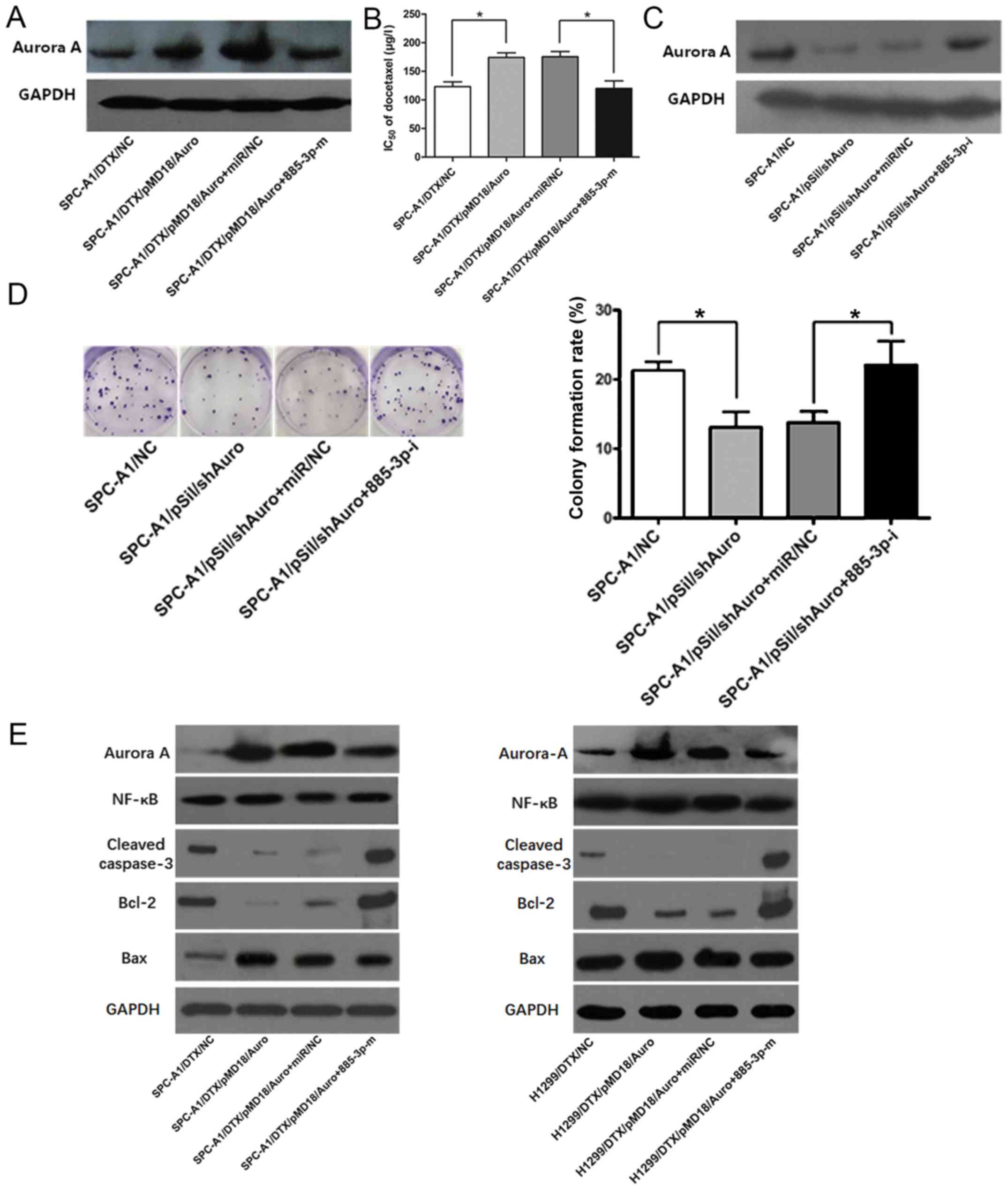
After confirming that Aurora A was amplified in the
drug-resistant cells, we regulated Aurora A expression in parental
and resistant SPC-A1 and H1299 cells by using the plasmid vectors
pMD18/Auro and pSil/shAuro to explore the relationship between
Aurora A expression and the degree of docetaxel resistance.
Transfection efficiency was validated by RT-qPCR and western
blotting (P<0.05, Fig. 1C and
D), and the Aurora A expression levels were increased in the
pMD/Auro-transfected SPC-A1 and H1299 cells and decreased in the
pSil/shAuro-transfected SPC-A1/DTX and H1299/DTX cells. MTT assays
were used to calculate the chemosensitivity (IC50 of
docetaxel) of various cell lines (SPC-A1, H1299, SPC-A1/DTX and
H1299/DTX cells) with different expression levels of Aurora A.
Higher expression of Aurora A promoted docetaxel resistance in
SPC-A1 or H1299 cells, and the chemosensitivity of SPC-A1/DTX and
H1299/DTX cells transfected with pSil/shAuro was significantly
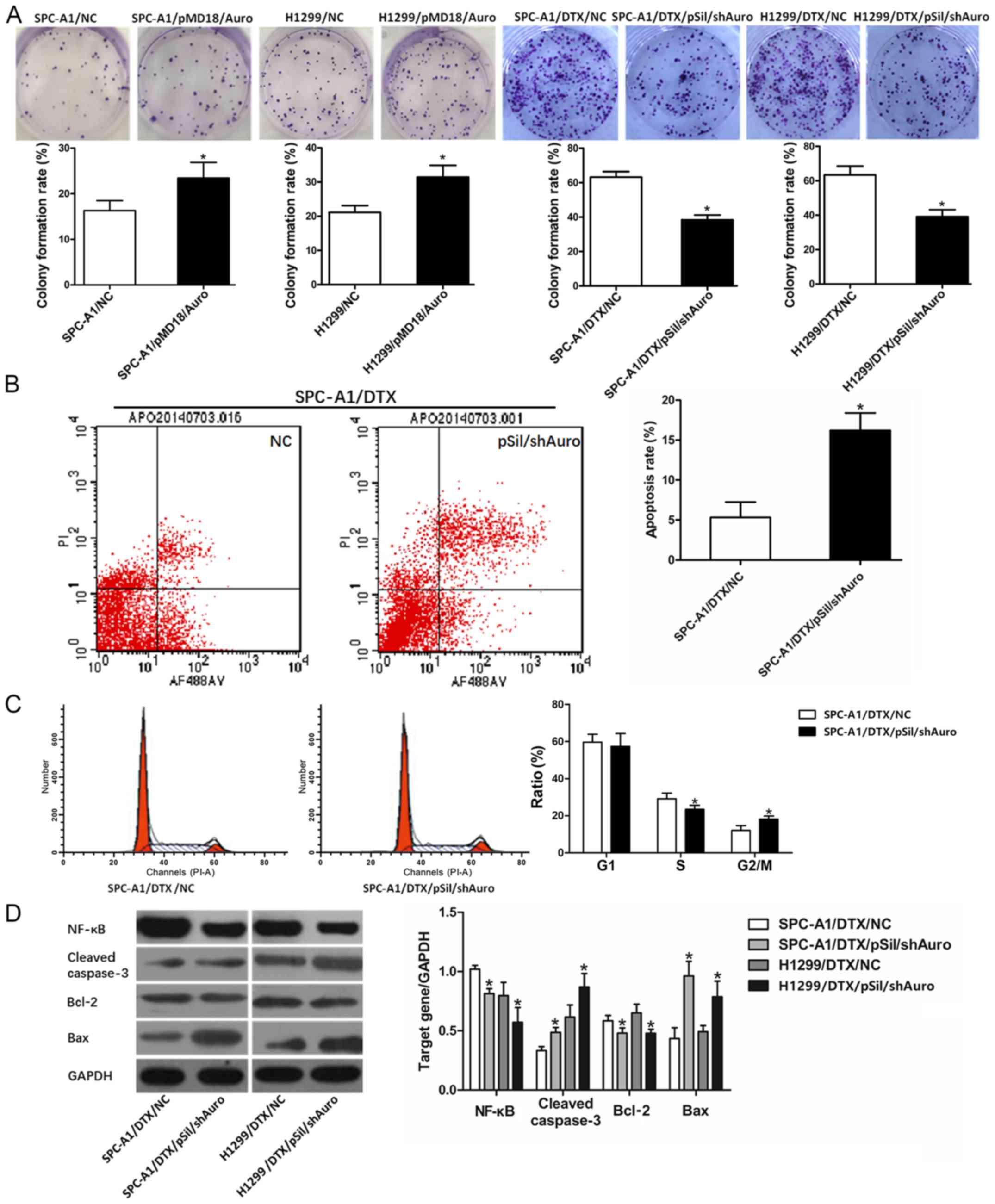
lower than that of the control cells (P<0.05, Fig. 1E). Similarly, the colony formation
viability of SPC-A1 and H1299 cells transfected with pMD/Auro was
enhanced, and pSil/shAuro transfection decreased the colony
formation rate of SPC-A1/DTX and H1299/DTX cells (P<0.05,
Fig. 2A). The MTT and colony
formation assay results demonstrated that Aurora A may be a key
induction factor for docetaxel resistance in LAD cells.
Silencing Aurora A affects
chemotherapy-induced apoptosis and cell cycle distribution in
SPC-A1/DTX cells
Next, we aimed to determine the relationship between
Aurora A and chemotherapy-induced apoptosis and cell cycle
distribution. pSil/shAuro was transfected into SPC-A1/DTX cells,
and the effects of Aurora A inhibition on SPC-A1/DTX cell apoptosis
were determined. The apoptosis rate was significantly higher in
SPC-A1/DTX/pSilAuro cells than in SPC-A1/DTX/NC cells treated with
the same docetaxel concentration (half of the IC50
concentration for 48 h) (P<0.05, Fig. 2B). To analyse the mechanisms by
which Aurora A expression affect cell proliferation, flow
cytometric analyses of the cell cycle were performed. As shown in
Fig. 2C, Aurora A inhibition
resulted in an increased proportion of cells in the G2/M phase and
a decreased proportion of cells in the S phase (P<0.05,
Fig. 2C).
Silencing Aurora A affects apoptosis
by regulating NF-κB and Bcl-2/Bax
According to the function of Aurora A, we analysed
the expression changes of molecules related to cell apoptosis and
proliferation. Western blot assays revealed that pSil/shAuro
decreased NF-κB protein levels in SPC-A1/DTX and H1299/DTX cells
(P<0.05, Fig. 2D).
Moreover, the expression levels of cleaved-caspase-3 were increased
when SPC-A1/DTX and H1299/DTX cells were transfected with
pSil/shAuro (P<0.05, Fig.
2D). Additionally, Bcl-2 downregulation and Bax upregulation
were observed in pSil/shAuro-transfected SPC-A1/DTX and H1299/DTX
cells, which reflected a decrease in the anti-apoptosis ability of
the cells (Bcl-2/Bax ratio).
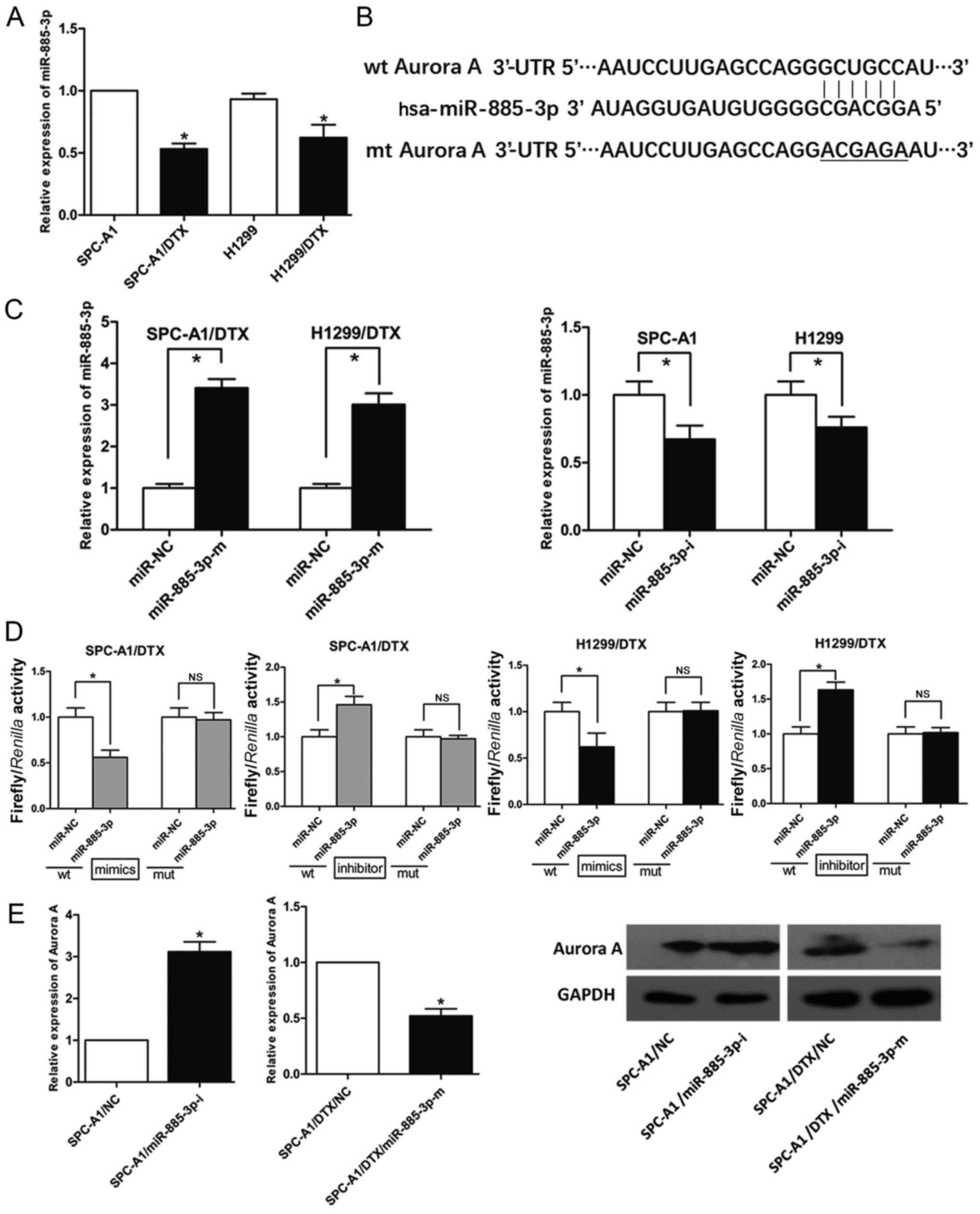
miR-885-3p targets Aurora A
directly
By using open access databases (PicTarget and
miRBase) and studying our previous microRNA profiles of
docetaxel-resistant human LADs, we found miR-885-3p to be a
preferred upstream candidate miRNA for controlling Aurora A due to
the putative binding site within its 3′-UTR (Fig. 3B); miR-885-3p was significantly
downregulated in SPC-A1/DTX and H1299/DTX cells (P<0.05,
Fig. 3A). Then, we designed mimics
and an inhibitor of miR-885-3p and validated their transfection
efficiency (P<0.05, Fig.
3C). To determine whether Aurora A is a direct downstream
target of miR-885-3p, luciferase reporter assays were performed. A
fragment of the Aurora A 3′-UTR containing the putative miR-885-3p
binding site was cloned into a luciferase reporter vector. As shown
in Fig. 3D, the luciferase reporter
assays indicated that the luciferase activities of LAD cells
transfected with the Aurora A-wt construct were significantly
suppressed after the transfection of miR-885-3p mimics and were
significantly increased after the transfection of the miR-885-3p
inhibitor; however, there was no significant difference found in
the cells transfected with the Aurora A-mut construct. Then, we
determined the effect of miR-885-3p on Aurora A protein expression
and found that miR-885-3p mimics decreased Aurora A protein
expression in SPC-A1/DTX cells, while the miR-885-3p inhibitor
increased Aurora A protein expression (Fig. 3E). Due to these results, we
concluded that miR-885-3p downregulated Aurora A expression by
targeting its 3′-UTR directly.
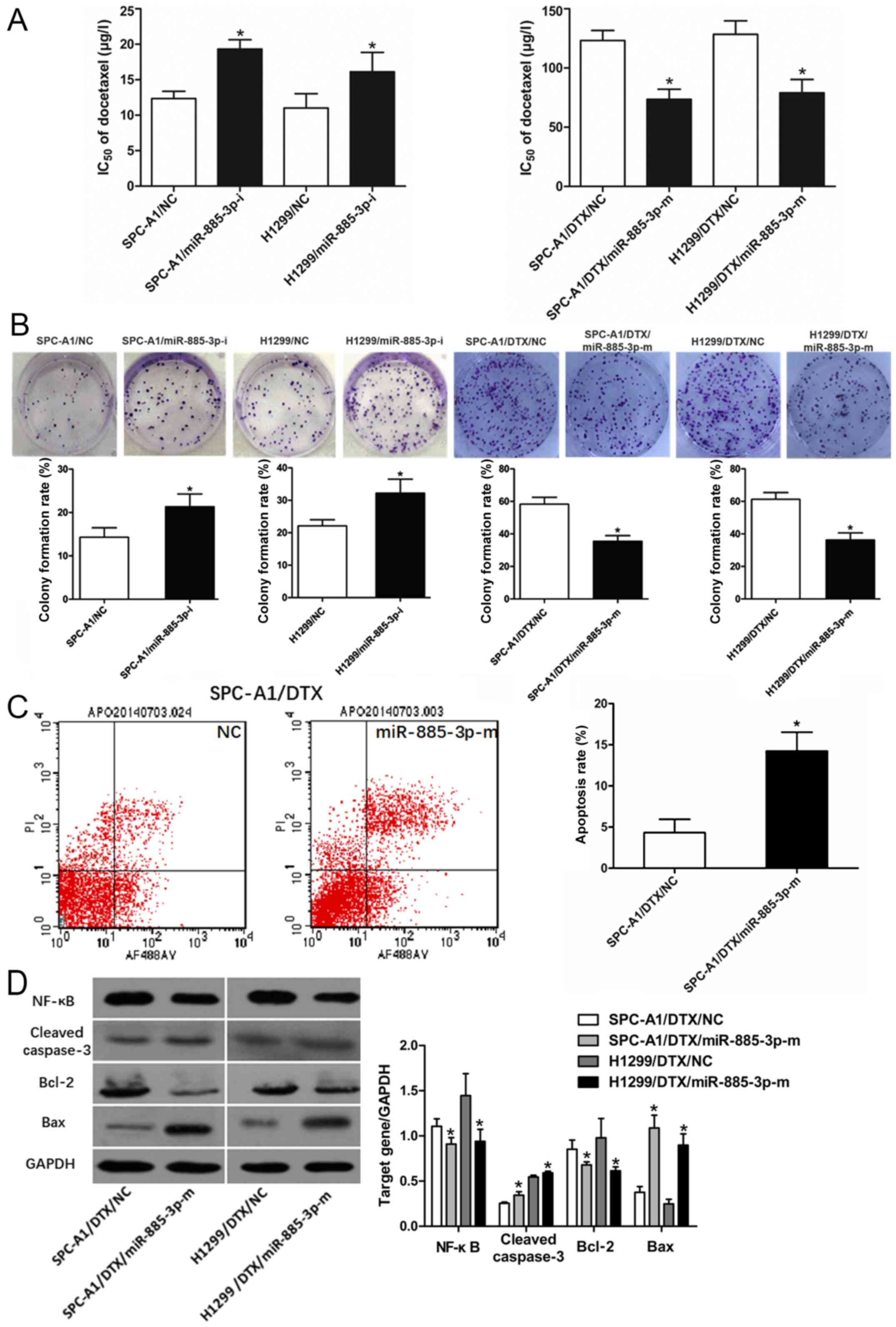
miR-885-3p is involved in docetaxel
resistance in lung adenocarcinoma
To gain more insight into how miR-885-3p is involved
in Aurora A-induced chemoresistance, we used gain-of-function and
loss-of-function experiments. First, SPC-A1 and H1299 cells were
transfected with a miR-885-3p inhibitor or negative control, and
SPC-A1/DTX and H1299/DTX cells were transfected with miR-885-3p
mimics or a negative control. MTT assays revealed that the
IC50 values of docetaxel in SPC-A1 and H1299 cells were
significantly increased, and the IC50 values of
docetaxel in SPC-A1/DTX and H1299/DTX cells were decreased
(P<0.05, Fig. 4A). Then,
the growth rates of SPC-A1 and H1299 cells transfected with the
miR-885-3p inhibitor were increased, but the colony formation
efficiency of SPC-A1/DTX and H1299/DTX cells was decreased by
miR-885-3p mimics (P<0.05, Fig. 4B).
miR-885-3p affects
chemotherapy-induced apoptosis by regulating NF-κB and
Bcl-2/Bax
To further examine whether miR-885-3p participated
in cell apoptosis and the molecular mechanisms of miR-885-3p in
apoptotic regulation, flow cytometric analyses of SPC-A1/DTX
apoptosis were performed. As expected, miR-885-3p overexpression
significantly increased apoptosis in SPC-A1/DTX cells exposed to
docetaxel (P<0.05, Fig.
4C). Western blots indicated that the expression of NF-κB was
significantly lower in SPC-A1/DTX and H1299/DTX cells than in cells
transfected with the negative control. Additionally,
cleaved-caspase-3 levels were increased in SPC-A1/DTX and H1299/DTX
cells with miR-885-3p overexpression (P<0.05, Fig. 4D). Bcl-2 downregulation and Bax
upregulation were also detected in miR-885-3p mimic-transfected
SPC-A1/DTX and H1299/DTX cells (P<0.05, Fig. 4D). These data revealed that
miR-885-3p may regulate the response to docetaxel by promoting cell
apoptosis.
High miR-885-3p expression in lung
adenocarcinoma is associated with decreased Aurora A expression and
chemotherapeutic resistance
To further explore whether Aurora A was involved in
the function of miR-885-3p in response to docetaxel, we then
performed rescue experiments and evaluated LAD cell proliferation
and apoptosis. After transfection with pMD18/Auro, SPC/DTX cells
were co-transfected with miR-885-3p mimics, which could partially
rescue the Aurora A expression upregulation and docetaxel
IC50 increase in SPC-A1/DTX cells (P<0.05, Fig. 5A and B). Co-transfection of
pSil/shAuro and a miR-885-3p inhibitor could partially rescue the
Aurora A expression downregulation and the colony formation arrest
in SPC-A1 cells (P<0.05, Fig. 5C and
D). Furthermore, co-transfection could partially rescue the
decreased expression of cleaved caspase-3 protein in SPC-A1/DTX and
H1299/DTX cells induced by the upregulation of Aurora A (Fig. 5E). As expected, co-transfection
could partially rescue the Bcl-2 protein overexpression and
decreased Bax protein expression in SPC-A1/DTX and H1299/DTX cells
induced by miR-885-3p inhibition. NF-κB protein analyses revealed
similar results. Collectively, these data indicated that miR-885-3p
may regulate the chemosensitivity of LAD cells in part by targeting
Aurora A.
miR-885-3p/Aurora A is involved in
epithelial-mesenchymal transition (EMT) in LAD cells
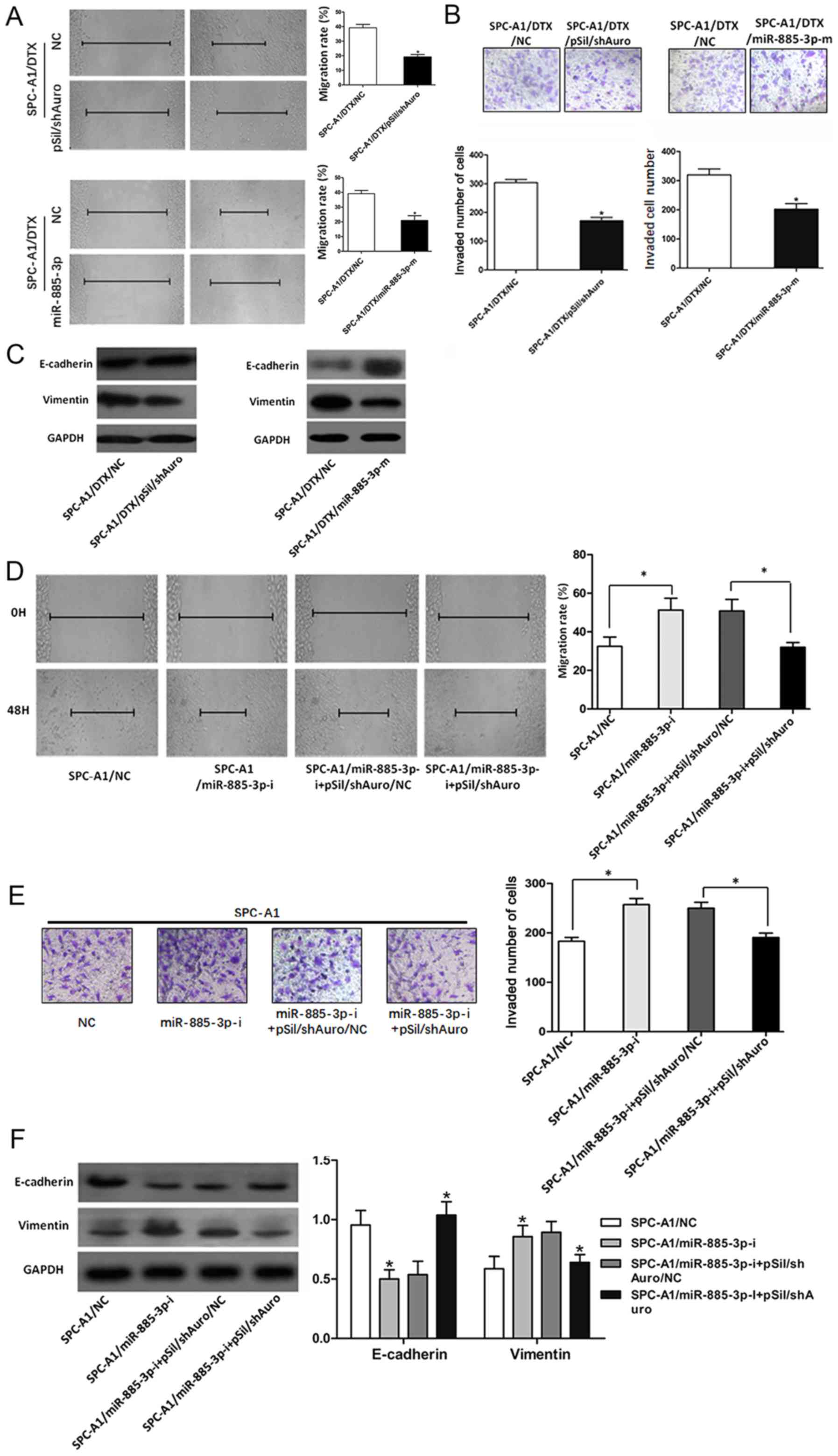
To determine the association between
miR-885-3p/Aurora A expression and EMT in LAD cells, SPC-A1/DTX
cells were transfected with miR-885-3p mimics and pSil/shAuro. Cell
wound scratch assays and Transwell invasion assays revealed
significant inhibition of the migration and invasion abilities of
SPC-A1/DTX cells (P<0.05, Fig. 6A and B). SPC-A1/DTX cells were
transfected with pSil/shAuro or miR-885-3p mimics. As shown in
Fig. 6C, pSil/shAuro and miR-885-3p
mimics could increase E-cadherin and downregulate vimentin
expression in SPC-A1/DTX cells. After transfection with the
miR-885-3p inhibitor, SPC/DTX cells were co-transfected with
pSil/shAuro. The co-transfection partially rescued the cell
migration and invasion and E-cadherin/vimentin changes induced by
the miR-885-3p inhibitor (Fig.
6D-F).
Discussion
Aurora A is a member of a new serine/threonine
kinase family that plays an important role in mitosis (15). In the process of forming the
spindle, Aurora A inhibition can cause the formation of the
single-pole spindle and influence the stability of the spindle. In
addition, Aurora A also participates in the regulation of the G2/M
phase cell cycle checkpoint; Aurora A overexpression can counter
the checkpoint activation induced by DNA damage in the G2/M phase,
resulting in genome instability and tumour formation. Increasing
evidence has revealed that aberrant Aurora A expression was found
in lung adenocarcinoma, liver, colon and breast cancer, and many
other malignant tumours (16,17).
The overexpression of Aurora A in cells can cause aneuploidy
formation and promote cell malignant transformation. It has been
reported that the overexpression of Aurora A could promote growth
and inhibit apoptosis in tumour cells (18). For instance, it was found that
Aurora A overexpression promoted human embryonic kidney HEK293T
cell proliferation and cell migration via the activation of cyclin
E/CDK2 and cyclin B1 (19). In
ovarian cancer, the combination of the Aurora A kinase inhibitor
alisertib and a CHEK1 inhibitor triggered apoptosis, reduced the
population of stem cells and increased the effect of taxanes and
platinum compounds (20). Aurora A
is also a good therapeutic target for inhibiting cancer cell growth
in gastric carcinoma, glioblastoma and small cell lung cancer
(21–23). Furthermore, the relationship between
Aurora A expression and tumour invasion and metastasis has been
recently revealed. Kozyreva et al revealed that the
combination of an Aurora A inhibitor with eribulin could lead to a
synergistic increase in apoptosis in mammary tumours, as well as
cytotoxic autophagy in metastases (24). Maimaiti et al reported that
the simultaneous overexpression of Aurora A and CFL-1 associated
with lymph node metastasis in thyroid cancer (25). The inhibition of Aurora A suppressed
thyroid cancer cell migration in vitro and decreased lymph
node metastasis in nude mice (25).
Notably, the roles of Aurora A overexpression in drug-resistance in
tumour cells remain unclear. Sun et al reported that the
inhibition of Aurora A promotes chemosensitivity via induction of
cell cycle arrest and apoptosis in cervical cancer cells (26). De Bacco et al revealed that
Aurora A inhibitors can radio-sensitize tumours and convert
GSC-positive selection (27). These
data indicated that Aurora A may play an important role in chemo-
or radio-resistance in human cancers. In our previous study, Aurora
A promoted the phosphorylation of the nuclear Ikappaβ-alpha (Iκβα)
protein and increased NF-kappa B (NFκB) activity, thus promoting
chemoresistance in HCC cells (8).
However, the association of Aurora A expression with
chemoresistance in LAD cells remains unclear. To investigate this
relationship, the associations between Aurora A protein expression
levels and docetaxel sensitivity in SPC-A1 and H1299 cells were
analysed in the present study. It was found that the expression
level of Aurora A was negatively associated with chemosensitivity
in LAD cells. Additionally, Aurora A overexpression increased the
IC50 values of docetaxel and cell growth in both the
SPC-A1 and H1299 cell lines. Then, we further investigated the
effect of Aurora A expression on chemosensitivity in LAD cells. At
the same concentration of docetaxel, silencing Aurora A increased
apoptosis compared to that in the control cells by enhancing
caspase-3-dependent apoptosis. In addition, Aurora A inhibition
increased the proportion of cells in the G2/M phase and decreased
the proportion of cells in the S phase. Moreover, western blot
assays revealed that pSil/shAuro decreased NF-κB, Bcl-2 levels, as
well as Bax protein levels, in SPC-A1/DTX cells. Therefore, the
overexpression of Aurora A promoted the formation of chemoresistant
LAD cells.
Chemoresistance is an important reason for the
therapeutic failure of lung adenocarcinoma. The mechanism of
chemotherapy resistance is very complicated and includes ATP
binding box transporters, drug metabolism enzymes,
hyperfunction-induced drug metabolism changes, drug target changes,
apoptotic protein level changes (such as Bcl-2 and Bax) and DNA
damage repair abnormalities (28).
Increasing evidence has shown that miRNAs may play a cursory role
in the development of drug resistance (29). miRNAs can reduce target gene
expression at the transcriptional level, affect multiple molecular
signalling pathways concurrently, and regulate key pathways in cell
apoptosis, the cell cycle and drug metabolism (30). In the present study, it was
concluded that the 3′-UTR of Aurora A is a direct target of
miR-885-3p. This conclusion was confirmed by four levels of
evidence. First, bioinformation prediction was conducted using the
Pictarget and miRBase platforms; second, there was a negative
association between the expression of miR-885-3p and Aurora A in
drug-resistant cells; third, luciferase reporter assays confirmed
that Aurora A-wt can block the effects of miR-885-3p on the
expression of Aurora A in drug-resistant cells; and fourth,
subsequent rescue experiments revealed that miR-885-3p-regulated
Aurora A expression was associated with docetaxel resistance
development. Recently, miR-885-3p was found to have binding sites
in the 3′-UTR regions of the Mdm4, Akt1 and Bcl-2 genes; these
binding sites affect protein expression and apoptosis-related
proteins. In a study of epithelial cell carcinoma, miR-885-3p acted
as a negative regulator of the AKT1 gene to affect cell metabolism
and chemotherapy drug sensitization (31). To reveal the relationship between
miR-885-3p and docetaxel resistance development, SPC-A1 and H1299
cells were transfected with a miR-885-3p inhibitor, and SPC-A1/DTX
and H1299/DTX cells were transfected with miR-885-3p mimics. The
IC50 values of docetaxel and cell proliferation rates in
SPC-A1 and H1299 cells were significantly increased, while the
IC50 values of docetaxel and cell proliferation rates in
SPC-A1/DTX and H1299/DTX cells were decreased. Then, we further
investigated the effect of miR-885-3p on chemosensitivity in LAD
cells. SPC-A1/DTX and H1299/DTX cells were stably transfected with
miR-885-3p mimics, and it was observed that miR-885-3p
overexpression increased apoptosis by enhancing caspase-3-dependent
apoptosis. Additionally, western blot assays revealed that
miR-885-3p mimics decreased NF-κB and Bcl-2 and upregulated Bax
protein levels in SPC-A1/DTX and H1299/DTX cells. Moreover, the
effects of Aurora A inhibition on chemoresistance in
docetaxel-resistant LAD cells were similar to the effects of the
miR-885-3p mimics. The miR-885-3p inhibitor could partially rescue
the effects of Aurora A silencing on chemosensitivity in
docetaxel-resistant LAD cells. In conclusion, miR-885-3p may confer
docetaxel chemoresistance in LAD cells partially by targeting
Aurora A and then regulating NF-κB and Bcl-2/Bax expression.
Epithelial-mesenchymal transition (EMT) includes
mainly the process of cell polarity loss; the surrounding cells and
matrix gradually disintegrate, and the abilities of migration and
movement become abnormal (32).
Research have shown that EMT is also closely connected with
chemoresistance in tumour cells (33). In a study on miR-885-3p, increased
miR-885-3p levels could block the BMP/Smad/Id1 signalling pathway
and regulate the function of TGF-β, which induced
epithelial-mesenchymal transition (13). Additionally, D'Assoro et al
found that Aurora A kinase could activate the EMT pathway, which is
responsible for the development of distant metastases in breast
cancer cells (34). In our previous
study, it was revealed that the methylation-associated silencing of
miR-129-3p promoted the epithelial-mesenchymal transition, invasion
and metastasis of hepatocellular cancer by targeting Aurora A
(35). To determine the connection
between miR-885-3p/Aurora A expression and EMT in LAD cells,
SPC-A1/DTX cells were transfected with miR-885-3p mimics and
pSil/shAuro. The migration and invasion abilities of SPC-A1/DTX
cells were significantly suppressed. Additionally, increased
E-cadherin and decreased vimentin expression levels in SPC-A1/DTX
cells were observed after transfection with miR-885-3p mimics or
pSil/shAuro. These data indicated that Aurora A overexpression or
miR-885-3p inhibition is associated with higher aggressive tumour
behaviour; thus, we further established that the miR-885-3p-Aurora
Axis may be partially responsible for EMT in human LAD cells.
In conclusion, the present study revealed that
Aurora A expression was significantly upregulated in
docetaxel-resistant LAD cells. Silencing Aurora A expression could
significantly increase chemosensitivity in LAD cells. Moreover,
miR-885-3p could affect the chemosensitivity of LAD cells to
docetaxel by targeting Aurora A and then regulating NF-κB and
Bcl-2/Bax expression. Further investigations using animal
experiments and human LAD tissue samples are required to confirm
the correlation between the function of miR-885-3p and its target
Aurora A and the responses to docetaxel-based chemotherapy in LAD
patients.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (no. 81472668) and the Young
Key Talents of Medicine Project in Jiangsu Province
(QNRC2016887).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
JC, JG, XC and RW contributed to the design of the
experiments, the acquisition, analysis and the interpretation of
the data. GH and LC conceived the study and contributed to the
interpretation of the data. JC and GH wrote the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All experiments were performed according to the
guidelines of the Ethics Committee of Jingling Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tunali I, Stringfield O, Guvenis A, Wang
H, Liu Y, Balagurunathan Y, Lambin P, Gillies RJ and Schabath MB:
Radial gradient and radial deviation radiomic features from
pre-surgical CT scans are associated with survival among lung
adenocarcinoma patients. Oncotarget. 8:96013–96026. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Domagala-Kulawik J and Raniszewska A: How
to evaluate the immune status of lung cancer patients before
immunotherapy. Breathe. 13:291–296. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xu-Welliver M and Carbone DP: Blood-based
biomarkers in lung cancer: Prognosis and treatment decisions.
Transl Lung Cancer Res. 6:708–712. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Torigoe H, Soh J, Tomida S, Namba K, Sato
H, Katsui K, Hotta K, Shien K, Yamamoto H, Yamane M, et al:
Induction chemoradiotherapy using docetaxel and cisplatin with
definitive-dose radiation followed by surgery for locally advanced
non-small cell lung cancer. J Thorac Dis. 9:3076–3086. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim ST, Kyung EJ, Suh JS, Lee HS, Lee JH,
Chae SI, Park ES, Chung YH, Bae J, Lee TJ, et al:
Phosphatidylcholine attenuated docetaxel-induced peripheral
neurotoxicity in rats. Drug Chem Toxicol. 41:476–485. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matsunaga T, Saito H, Endo S, Iguchi K,
Soda M, El-Kabbani O, Hara A and Ikari A: Roles of aldo-keto
reductases 1B10 and 1C3 and ATP-binding cassette transporter in
docetaxel tolerance. Free Radic Res. 50:1296–1308. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fennell DA, Summers Y, Cadranel J, Benepal
T, Christoph DC, Lal R, Das M, Maxwell F, Visseren-Grul C and Ferry
D: Cisplatin in the modern era: The backbone of first-line
chemotherapy for non-small cell lung cancer. Cancer Treat Rev.
44:42–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang K, Chen J, Chen D, Huang J, Feng B,
Han S, Chen Y, Song H, De W, Zhu Z, et al: Aurora A promotes
chemoresistance in hepatocelluar carcinoma by targeting
NF-kappaB/microRNA-21/PTEN signaling pathway. Oncotarget.
5:12916–12935. 2014.PubMed/NCBI
|
|
9
|
Lykkesfeldt AE, Iversen BR, Jensen MB,
Ejlertsen B, Giobbie-Hurder A, Reiter BE, Kirkegaard T and
Rasmussen BB: Aurora kinase A as a possible marker for endocrine
resistance in early estrogen receptor positive breast cancer. Acta
Oncol. 57:67–73. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee DH, Kim CG, Lim Y and Shin SY: Aurora
kinase A inhibitor TCS7010 demonstrates pro-apoptotic effect
through the unfolded protein response pathway in HCT116 colon
cancer cells. Oncol Lett. 14:6571–6577. 2017.PubMed/NCBI
|
|
11
|
Tagal V, Wei S, Zhang W, Brekken RA,
Posner BA, Peyton M, Girard L, Hwang T, Wheeler DA, Minna JD, et
al: SMARCA4-inactivating mutations increase sensitivity to Aurora
kinase A inhibitor VX-680 in non-small cell lung cancers. Nat
Commun. 8:140982017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rui W, Bing F, Hai-Zhu S, Wei D and
Long-Bang C: Identification of microRNA profiles in
docetaxel-resistant human non-small cell lung carcinoma cells
(SPC-A1). J Cell Mol Med. 14:206–214. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiao F, Qiu H, Cui H, Ni X, Li J, Liao W,
Lu L and Ding K: MicroRNA-885-3p inhibits the growth of HT-29 colon
cancer cell xenografts by disrupting angiogenesis via targeting
BMPR1A and blocking BMP/Smad/Id1 signaling. Oncogene. 34:1968–1978.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Summerer I, Niyazi M, Unger K, Pitea A,
Zangen V, Hess J, Atkinson MJ, Belka C, Moertl S and Zitzelsberger
H: Changes in circulating microRNAs after radiochemotherapy in head
and neck cancer patients. Radiat Oncol. 8:2962013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bury L, Coelho PA, Simeone A, Ferries S,
Eyers CE, Eyers PA, Zernicka-Goetz M and Glover DM: Plk4 and Aurora
A cooperate in the initiation of acentriolar spindle assembly in
mammalian oocytes. J Cell Boil. 216:3571–3590. 2017. View Article : Google Scholar
|
|
16
|
Martin D, Fallaha S, Proctor M, Stevenson
A, Perrin L, McMillan N and Gabrielli B: Inhibition of Aurora A and
Aurora B is required for the sensitivity of HPV-Driven cervical
cancers to Aurora kinase inhibitors. Mol Cancer Ther. 16:1934–1941.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Simpson PV, Casari I, Paternoster S,
Skelton BW, Falasca M and Massi M: Defining the anti-cancer
activity of tricarbonyl rhenium complexes: Induction of G2/M cell
cycle arrest and blockade of Aurora A kinase phosphorylation.
Chemistry. 23:6518–6521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Asteriti IA, De Mattia F and Guarguaglini
G: Cross-talk between AURKA and Plk1 in mitotic entry and spindle
assembly. Front Oncol. 5:2832015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vo TT, Park JH, Seo JH, Lee EJ, Choi H,
Bae SJ, Le H, An S, Lee HS, Wee HJ, et al: ARD1-mediated aurora
kinase A acetylation promotes cell proliferation and migration.
Oncotarget. 8:57216–57230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Alcaraz-Sanabria A, Nieto-Jimenez C,
Corrales-Sanchez V, Serrano-Oviedo L, Andrés-Pretel F, Montero JC,
Burgos M, Llopis J, Galán-Moya EM, Pandiella A, et al: Synthetic
lethality interaction between aurora kinases and CHEK1 inhibitors
in ovarian cancer. Mol Cancer Ther. 16:2552–2562. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qiao W, Guo B, Zhou H, Xu W, Chen Y, Liang
Y and Dong B: miR-124 suppresses glioblastoma growth and
potentiates chemosensitivity by inhibiting AURKA. Biochem Biophys
Res Commun. 486:43–48. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mesic A, Markocic E, Rogar M, Juvan R,
Hudler P and Komel R: Single nucleotide polymorphisms rs911160 in
AURKA and rs2289590 in AURKB mitotic checkpoint genes
contribute to gastric cancer susceptibility. Environ Mol Mutagen.
58:701–711. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu Y, Liu Y, Jiang J, Xi Z, Zhong N, Shi
S, Wang J and Wei X: Knocking down the expression of Aurora A gene
inhibits cell proliferation and induces G2/M phase arrest in human
small cell lung cancer cells. Oncol Rep. 32:243–249. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kozyreva VK, Kiseleva AA, Ice RJ, Jones
BC, Loskutov YV, Matalkah F, Smolkin MB, Marinak K, Livengood RH,
Salkeni MA, et al: Combination of eribulin and Aurora A inhibitor
MLN8237 prevents metastatic colonization and induces cytotoxic
autophagy in breast cancer. Mol Cancer Ther. 15:1809–1822. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Maimaiti Y, Jie T, Jing Z, Changwen W, Pan
Y, Chen C and Tao H: Aurora kinase A induces papillary thyroid
cancer lymph node metastasis by promoting cofilin-1 activity.
Biochem Biophys Res Commun. 473:212–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun JM, Yang LN, Xu H, Chang B, Wang HY
and Yang G: Inhibition of Aurora A promotes chemosensitivity via
inducing cell cycle arrest and apoptosis in cervical cancer cells.
Am J Cancer Res. 5:1133–1145. 2015.PubMed/NCBI
|
|
27
|
De Bacco F, D'Ambrosio A, Casanova E,
Orzan F, Neggia R, Albano R, Verginelli F, Cominelli M, Poliani PL,
Luraghi P, et al: MET inhibition overcomes radiation resistance of
glioblastoma stem-like cells. EMBO Mol Med. 8:550–568. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bhat P, Kriel J, Shubha Priya B, Basappa,
Shivananju NS and Loos B: Modulating autophagy in cancer therapy:
Advancements and challenges for cancer cell death sensitization.
Biochem Pharmacol. 147:170–182. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Caliskan M, Guler H and Bozok Cetintas V:
Current updates on microRNAs as regulators of chemoresistance.
Biomed Pharmacother. 95:1000–1012. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bourguignon LY, Earle C and Shiina M:
Activation of matrix hyaluronan-mediated CD44 signaling, epigenetic
regulation and chemoresistance in head and neck cancer stem cells.
Int J Mol Sci. 18:E18492017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang Y, Chuang AY and Ratovitski EA:
Phospho-ΔNp63α/miR-885-3p axis in tumor cell life and cell death
upon cisplatin exposure. Cell Cycle. 10:3938–3947. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lo UG, Lee CF, Lee MS and Hsieh JT: The
role and mechanism of epithelial-to-mesenchymal transition in
prostate cancer progression. Int J Mol Sci. 18:E20792017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng HC: The molecular mechanisms of
chemoresistance in cancers. Oncotarget. 8:59950–59964.
2017.PubMed/NCBI
|
|
34
|
D'Assoro AB, Liu T, Quatraro C, Amato A,
Opyrchal M, Leontovich A, Ikeda Y, Ohmine S, Lingle W, Suman V, et
al: The mitotic kinase Aurora - a promotes distant metastases by
inducing epithelial-to-mesenchymal transition in ERalpha(+) breast
cancer cells. Oncogene. 33:599–610. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cui S, Zhang K, Li C, Chen J, Pan Y, Feng
B, Lu L, Zhu Z, Wang R and Chen L: Methylation-associated silencing
of microRNA-129-3p promotes epithelial-mesenchymal transition,
invasion and metastasis of hepatocelluar cancer by targeting Aurora
A. Oncotarget. 7:78009–78028. 2016. View Article : Google Scholar : PubMed/NCBI
|