Introduction
Colorectal cancer (CRC) is one of the most common
malignant tumors in humans. When diagnosed with advanced metastatic
disease, CRC patients traditionally have a poor prognosis, with
5-year survival rates in the range of 5 to 8% (1,2).
Clinical studies have shown that the main cause of death in
patients with CRC is tumor invasion and metastasis to other tissues
and organs (3). In addition to
surgical resection of the primary tumor, the clinical key to
improving patient survival and curing CRC is to inhibit
angiogenesis, cutting off the transfer route and looking for new
targets for effective drug treatment (4). The occurrence of CRC involves a
variety of oncogenes and tumor-suppressor genes, although the
underlying molecular mechanisms remain unclear (5,6). The
study of targeted gene therapy can provide a theoretical basis for
the treatment of the invasion and metastasis of CRC.
As a tumor-suppressor gene, the liver kinase B1 gene
(LKB1, also known as serine/threonine kinase 11, STK11) is closely
related to tumor angiogenesis, invasion and metastasis (7,8). The
loss of LKB1 gene expression is related not only to the occurrence
of a variety of tumors, such as CRC and thyroid carcinoma, but also
to tumor angiogenesis and metastasis (9–11).
Knockdown of LKB1 was found to enhance cell migration in human
colon cancer HCT116 cells (12).
The molecular mechanism by which the LKB1 pathway acts on
angiogenesis and metastasis in CRC remains unclear. It is evident
that small interfering RNAs (siRNAs) play an important role in
cancer initiation and progression via the regulation of
tumor-suppressor genes or oncogenes (13–15).
In the present study, the LKB1 gene was silenced by siRNA, and the
mechanism by which LKB1 affects the downstream invasion and
metastasis of CRC cells was observed.
LKB1 has recently been identified as an upstream
activating protein kinase of AMP-activated protein kinase (AMPK)
(16). The study found that AMPK
and the development of CRC are closely related (17,18).
Our studies revealed an important role played by AMPK alpha 1
(AMPKa1) in cell biology, connecting two hallmarks of tumor cells,
namely, hyperproliferation and DNA damage, which may be due to
reduction in the amount of p21 (19). These findings have important
implications for understanding the molecular mechanisms by which
AMPK acts as a promising tumor suppressor or senescence blocker. Su
et al found that AMPK was closely related to angiogenesis in
endothelial cells (20). LKB1 is a
key regulator of the AMPK signaling pathway in some tissues,
including liver, skeletal muscle, myocardium and various cancerous
tissues (21,22). Our data indicated that protein
kinase B (AKT) is involved in the angiogenesis and metastasis of
colon cancer. Aberrant AKT activation was found to promote the
growth of a variety of LKB1-deficient tumors (23) by promoting the activation of
vascular endothelial growth factor and angiogenic factors (24). Therefore, it is important to study
the expression and regulation of LKB1 to clarify the mechanisms
underlying the invasion and metastasis of CRC.
In the present study, immunohistochemistry (IHC) was
used to detect the expression of the tumor-suppressor gene LKB1 in
different colon tissues. CRC LoVo cells were transfected with LKB1
small interfering RNA (LKB1 siRNA). Metformin has the potential
mechanism to inhibit tumor cell growth and reduce protein synthesis
in CRC. The present study used LoVo cells treated with metformin to
study the expression and biological significance of LKB1-AMPK-AKT
to clarify the metastatic mechanism of CRC. We speculated that LKB1
activates AMPK, acting as a negative regulator of the AKT pathways
and affecting the invasion and metastasis of colon cancer cells. In
the present study, we explored the possibility of the prevention
and treatment of the invasion and metastasis of colon cancer via
the LKB1 pathway.
Materials and methods
Materials
The colon cancer tissue microarray was purchased
from Fanpu Biotech, Inc. (cat. no. COC961; Guilin, China). The SP
IHC kit and primary antibody dilution buffer were purchased from
Sangon Biotech (Shanghai, China). The LoVo human CRC cell line was
obtained from CHI Scientific (cat. no. CL-0144, Wuxi, China). The
artificially synthesized LKB1 siRNA and RNase-free water were
obtained from RiboBio (Guangzhou, China). Lipofectamine™ 2000 was
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
All primary antibodies [LKB1 (dilution 1:1,000; cat. no. 3050S),
p-LKB1 (dilution 1:1,000; cat. no. 3482S), AMPKa (dilution 1:1,000;
cat. no. 2532S), p-AMPKa (Thr172, dilution 1:1,000; cat. no.
2535S), AKT (dilution 1:1,000; cat. no. 9272S), p-AKT (dilution
1:2,000; cat. no. 4060S)] were obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA). The Prestained Dual Color
Protein Molecular Weight Marker was purchased from Thermo Fisher
Scientific, Inc. The Cell Counting Kit-8 (CCK-8) was purchased from
Dojindo Laboratories, Co., Ltd. (Kyushu, Japan). Metformin was
purchased from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany). The
study was approved by the Ethics Committee of Hubei University of
Science and Technology (Xianning, China).
IHC
The rabbit anti-human LKB1 monoclonal antibody
(dilution 1:200; cat. no. ab185734) was purchased from Sangon
Biotech. The colorectal carcinoma chip was purchased from Fanpu
Biotech, Inc. This chip included samples from 65 patients with
different types of colorectal tumors and samples of normal and
inflammatory polyps from 31 patients. The samples were
adenocarcinoma and mucinous adenocarcinoma tissues from 65
patients. Of the 65 patients, 35 were male and 30 were female; 36
were younger than 60 years, while 29 were older than 60 years. The
TNM staging of colon cancer was carried out according to the 8th
edition of the AJCC Cancer Staging Manual (25). In the present study, we observed the
effect of LKB1 on metastasis of CRC. Therefore, the lymph node
metastasis status in the present study was divided into two types:
No metastasis and metastasis, and no detailed distinction was made.
The 65 colorectal tumor samples were composed of 20 cases of
adenocarcinoma with pathological grade I, 31 cases of
adenocarcinoma with pathological grade II, and 14 cases of
adenocarcinoma with pathological grade III. The tissues were cut
into 4-µm-thick slices. The IHC SP method was performed according
to the manufacturer's instructions, with known positive tissue
sections as a positive control and phosphate-buffered saline (PBS)
instead of primary antibody as a negative control. The procedure
was conducted as follows: i) Dewaxing and hydration with a graded
ethanol series moving from a high ethanol concentration to a low
ethanol concentration; ii) antigen repair; iii) incubation with
normal goat serum solution at 37°C for 10 min; iv) application of
the primary antibody LKB1; dilution 1:200) at 4°C and incubation
overnight, washing with PBS 3 times for 3 min each time, incubation
with the mouse anti-rabbit secondary antibody (dilution 1:200; cat.
no. BM2004; Wuhan Boster Biological Technology Co., Ltd., Wuhan,
China) at 37°C for 1 h and washing with PBS 3 times for 3 min each
time; v) application of the horseradish peroxidase-labeled
streptomycin-avidin working solution and washing with PBS 3 times
for 3 min each time; vi) DAB/H2O2 reactive
dyeing followed by thorough rinsing with water, hematoxylin
staining, dehydration, transparent and drying; vii) mounting with a
neutral gum with no bubbles. The sections were scanned at
magnification (×200) using an light microscopy (Olympus Corp.
Tokyo, Japan). The intensity of the staining as well as the
percentage of positive cells was recorded. Staining intensity was
scored from 0 to 3+; the intensity score was established as
follows: 0 if tumor cells had complete absence of staining or faint
staining intensity in <10%; 1+ if >10% of tumor cells had
faint staining; 2+ if tumor cells had moderate staining; and 3+ if
tumor cells had strong staining. Tumors with 1+, 2+ and 3+
expression were interpreted as positive for LKB1 antibody
expression, and tumors with no expression (0 score) were
interpreted as negative.
Cell culture
LoVo cells were maintained in RPMI-1640 culture
medium supplemented with 10% fetal bovine serum (FBS; CHI
Scientific) with 1% penicillin-streptomycin (HyClone; GE Healthcare
Life Sciences, Logan, UT, USA). The cells were grown in monolayer
cultures with 5% CO2 in a humidified 37°C incubator.
Every 2–3 days, the cells were subcultured. When the cells reached
the logarithmic growth phase, 0.25% trypsin (Gibco Invitrogen;
Thermo Fisher Scientific, Inc.) was applied for 1–3 min. The cells
were resuspended in RPMI-1640 containing 10% FBS at a cell
concentration of 1×104 cells/ml. The experimental groups
were as follows: i) blank control group, untransfected LoVo cells;
ii) metformin group, LoVo cells treated with 20 mmol/l metformin;
iii) LKB1 siRNA group, LoVo cells transfected with LKB1 siRNA; and
iv) LKB1 siRNA+metformin group, LoVo cells transfected with LKB1
siRNA were also treated with 20 mmol/l metformin.
LKB1 siRNA synthesis and
transfection
The cells were divided into the following 3 groups:
i) blank control group, untransfected LoVo cells; ii) negative
control group, LoVo cells transfected with
Lipofectamine™ 2000; iii) positive control group (LKB1
siRNA group), transfected with LKB1 siRNA. Appropriate positive and
negative controls were run simultaneously. In total, 3 groups of
cells were transfected with siRNAs the sequences of which were
specific for LKB1 (namely, siRNA-001, siRNA-002 and siRNA-003).
Another group of cells was transfected with a fluorescein amidite
(FAM)-labeled non-specific siRNA that served as the negative
control (NC) siRNA. The sequences of the siRNAs used in the present
study were as follows: siRNA-001 sense, 5′-GAAGAAGGAAAUUCAACUA-3′
and antisense, 5′-UAGUUGAAUUUCCUUCUUC-3′; siRNA-002sense,
5′-GCUGGUUCCGGAAGAAACA-3′ and antisense, 5′-UGUUUCUUCCGGAACCAGC-3′;
siRNA-003 sense, 5′-GGACUGACGUGUAGAACAA-3′ and antisense,
5′-UUGUUCUACACGUCAGUCC-3′. In the present study, the risk of this
potential off-target effect of siRNA affected the application of
siRNA in this technology gene therapy field. LKB1 siRNA was
synthesized by Guangzhou RiboBio Co., Ltd. The company is careful
to avoid the off-target effect when designing siRNA, and the
company offers siRNA modification. The company conducts product
quality testing after the product is produced. For cell
transfection, the cells were plated on 6-well plates
(2×105 cells for LoVo) in RPMI-1640 with 10% FBS. After
12 h, 10 µl of siRNA/well was incubated with 100 µl of Opti-MEM,
and 5 µl of Lipofectamine™ 2000 was diluted with 100 µl
of Opti-MEM; 200 µl of the above mixture was added/well according
to the manufacturer's instructions. The transfected cells were
incubated in a humidified 37°C incubator with 5% CO2 for
48 h.
Examination of morphological
changes
Following the application of LoVo-Con siRNA or LKB1
siRNA for 48 h, an inverted phase-contrast microscope (Nikon
Corporation, Tokyo, Japan) was utilized to observe the
morphological changes in the cells. Images were captured using a
digital camera (Nikon Corporation) at a magnification of ×200.
CCK-8 assay
The effect on cell proliferation and toxicity due to
metformin were assessed using a CCK-8 assay. Briefly,
3×104 cells/well were seeded in 96-well culture plates.
After overnight incubation, the cells were treated with varying
concentrations of metformin for 24, 48 and 72 h. The concentrations
of metformin were 0, 5, 10, 15, 20 and 25 mmol/l. A total of 20 µl
of CCK-8 working solution was added to each well, followed by
incubation for 4 h at 37°C, and the absorbance was finally measured
at 450 nm using a Model 3550 Microplate Reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Migration and invasion assays
Transwell chambers (Corning, Inc., Corning, NY, USA)
were used to perform the cell migration and invasion assays. The
cells were inoculated into 6-well plate with RPMI-1640 containing
10% FBS at a cell concentration of 2×104 cells/ml.
First, 800 µl of RPMI-1640 medium plus 10% FBS was added to a
24-well plate, and then the plate was inserted into the Transwell
chamber. After 1 h, 200 µl of cell suspension was added to the
upper Transwell chamber. The cells were grown in monolayer cultures
with 5% CO2 in a humidified 37°C incubator. After 24 h,
the Transwell chamber was washed once with sterile PBS. The cells
that had not migrated were wiped off the upper surface with a clean
cotton ball, and then a 10% methanol solution was used to fix the
migrated cells for 30 min. The film was carefully cut and a drop of
5% crystal violet dye was added to the membrane. The cells were
incubated at room temperature for 20 min, and then washed once with
PBS. The slide was fixed with neutral gum. Finally, the purple-blue
cells were observed and counted under the multifield view of the
microscope. The number of tumor cells was calculated in 5 random
fields at a magnification of ×200, using an inverted microscope
(Olympus DP7; Olympus Corp.) and expressed as the average number of
cells/field of view. The number of migrating cells present in 9
random fields was counted, and the experiment was repeated ≥3
times. Invasion assays were performed using the same protocol,
except with Matrigel (BD Biosciences, San Jose, CA, USA)-coated
Transwell chambers and incubation for 24 h at 37°C.
Western blot analysis
Protein lysates from cells were extracted in
ice-cold lysis buffer [10 µl PMSF, 10 µl NaVO3, 10 µl
phosphatase inhibitor cocktail and 10 µl protease inhibitor
cocktail (Sigma-Aldrich; Merck KGaA) in 1 ml RIPA]. Total protein
was subjected to SDS-PAGE followed by immunoblotting. The total
protein content was quantitated according to the Bradford method.
Purified protein lysates (40 µg) were separated electrophoretically
on 12% denaturing SDS polyacrylamide gels (separating gel) and 5%
denaturing SDS polyacrylamide gel (concentrated gum). Then, the
protein was transferred to nitrocellulose membranes. After blocking
with 5% non-fat dry milk, the membranes were probed with primary
antibodies [LKB1 (dilution 1:1,000); p-LKB1 (dilution 1:1,000);
AMPKa (dilution 1:1,000); p-AMPKa (Thr172, dilution 1:1,000); AKT
(dilution 1:1,000); p-AKT (dilution 1:2,000); β-actin mouse
monoclonal antibody (dilution 1:2,000; cat. no. BM0627; Wuhan
Boster Biological Technology, Co., Ltd.)] at 4°C and incubation
overnight. The immunoblotted proteins were visualized using the
goat anti-mouse secondary antibody (dilution 1:5,000; cat. no.
BA1001; Wuhan Boster Biological Technology, Co., Ltd.) or mouse
anti-rabbit secondary antibody (dilution 1:5,000; cat. no. BM2004;
Wuhan Boster Biological Technology Co., Ltd.) and detected using
enhanced chemiluminescence (ECL) substrate (Cell Signaling
Technology, Inc.). Appropriate positive and negative controls were
run simultaneously. Western blot analysis was repeated 2–4 times.
The protein bands were analyzed and quantified using a Bio-Profil
Image Analysis system (Vilber Lourmat, Marne La Vallee, France),
and the protein expression levels were expressed in relative units
(RU). The results were normalized to actin.
Statistical analysis
The statistical software package SPSS 21.0 (IBM
Corp., Armonk, NY, USA) was used for the statistical analysis. The
experimental results are presented as the mean ± standard deviation
(mean ± SD). The data analysis was performed with one-way ANOVA and
χ2 tests. One-way analysis of variance with least
significant difference (LSD) post hoc test for multiple comparisons
was performed. P-value <0.05 was considered to indicate a
statistically significant result.
Results
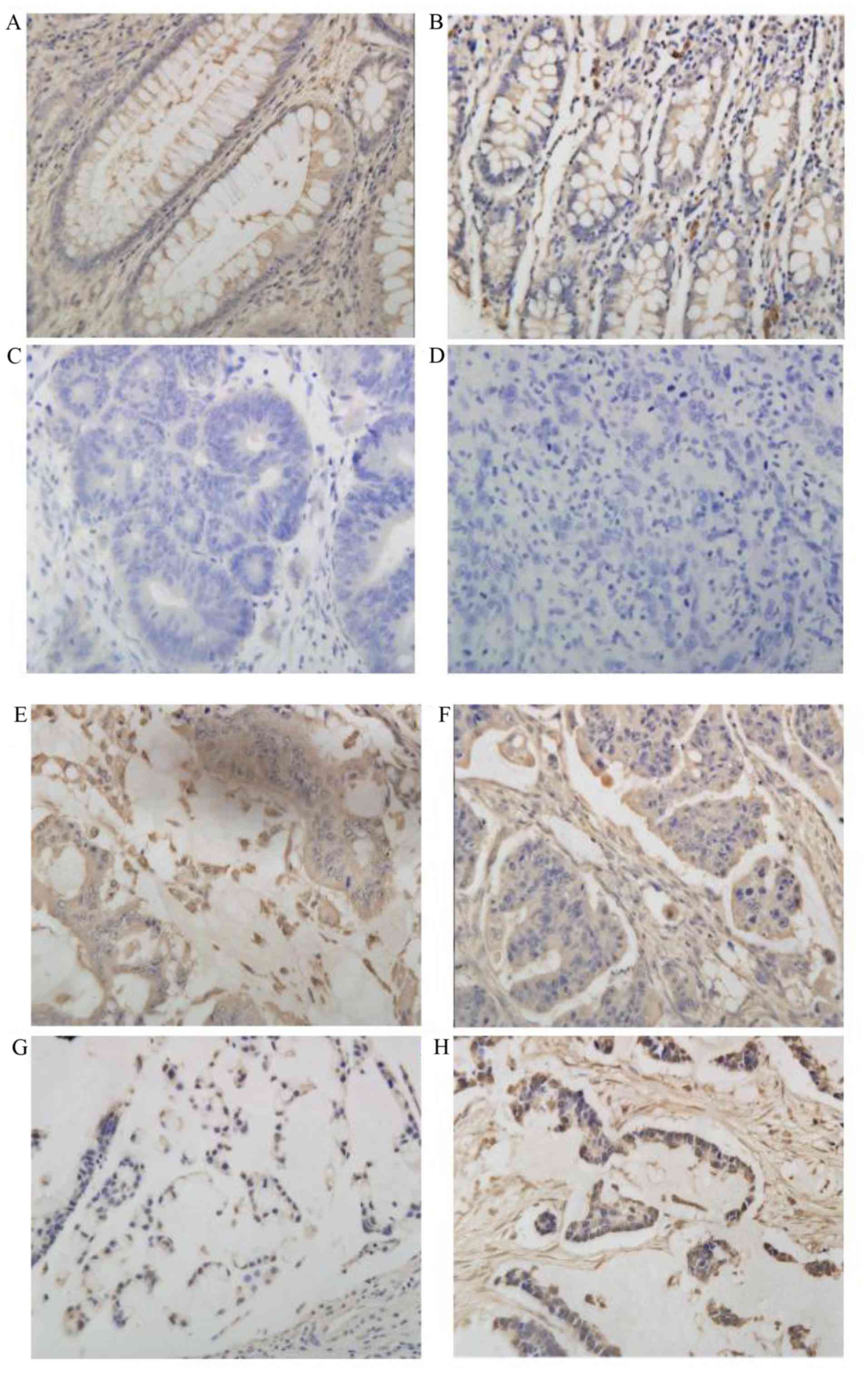
Expression of LKB1 protein in CRC and
normal colon mucosa
LKB1 protein expression was evaluated using IHC
staining in 96 cases. LKB1 protein expression and staining scores
were observed in the cytoplasm and the nucleus. The expression of
LKB1 was detected in the cytoplasm and nuclei of normal colon
tissue, and positive expression was observed in 10 cases (100%).
Positive expression of LKB1 was identified in 12 (57.1%) of the
inflammatory polyp tissue samples examined, and negative expression
was identified in 9 cases (42.9%). LKB1 expression was detected in
the cytoplasm and nucleus of the CRC cells, and positive expression
was observed in 28 cases (43.1%), while negative expression was
detected in 37 cases (56.9%). LKB1 expression in different tissues
was significantly different (P<0.01; Table I and Fig. 1).
 | Table I.Immunohistochemical staining of LKB1
in CRC and normal colon mucosa. |
Table I.
Immunohistochemical staining of LKB1
in CRC and normal colon mucosa.
| Groups | Total no. N | Strong positive
staining n (%) | Positive staining n
(%) | Negative staining n
(%) |
|---|
| Normal tissue | 10 | 10 (100.0) | 0 (0) | 0 (0) |
| Inflammatory
polyp | 21 | 0 (0) | 12 (57.1) | 9 (42.9) |
|
Adenocarcinoma/mucinous
adenocarcinoma | 65 | 0 (0) | 28 (43.1) | 37 (56.9) |
The association between LKB1 expression and
clinicopathological factors was analyzed in patients with CRC
(Table II). The positive
expression of LKB1 was significantly associated with the depth of
invasion and lymph node metastasis (χ2=4.205 and 9.802;
P<0.01). However, LKB1 expression was not associated with sex,
age or the degree of differentiation (χ2=0.215, 0.041
and 2.550; P>0.05).
| Table II.Association between
clinicopathological factors and LKB1 protein expression as
determined by IHC. |
Table II.
Association between
clinicopathological factors and LKB1 protein expression as
determined by IHC.
|
|
| LKB1 |
|
|
|---|
|
|
|
|
|
|
|---|
| Parameter | Cases | Positive | Negative | χ2 | P-value |
|---|
| Sex |
|
Male | 35 | 19 | 16 | 0.215 | 0.643 |
|
Female | 30 | 18 | 12 |
|
|
| Age (years) |
|
≤60 | 36 | 22 | 14 | 0.041 | 0.839 |
|
>60 | 29 | 17 | 12 |
|
|
| Degree of
differentiation |
| I | 20 | 15 | 5 | 2.550 | 0.279 |
| II | 31 | 18 | 13 |
|
|
|
III | 14 | 7 | 7 |
|
|
| Infiltrating
depth |
|
T1+T2 | 35 | 17 | 18 | 4.205 | 0.004 |
| T3 | 30 | 22 | 8 |
|
|
| Lymph node
m]etastasis |
| No | 37 | 15 | 22 | 9.802 | 0.002 |
|
Yes | 28 | 22 | 6 |
|
|
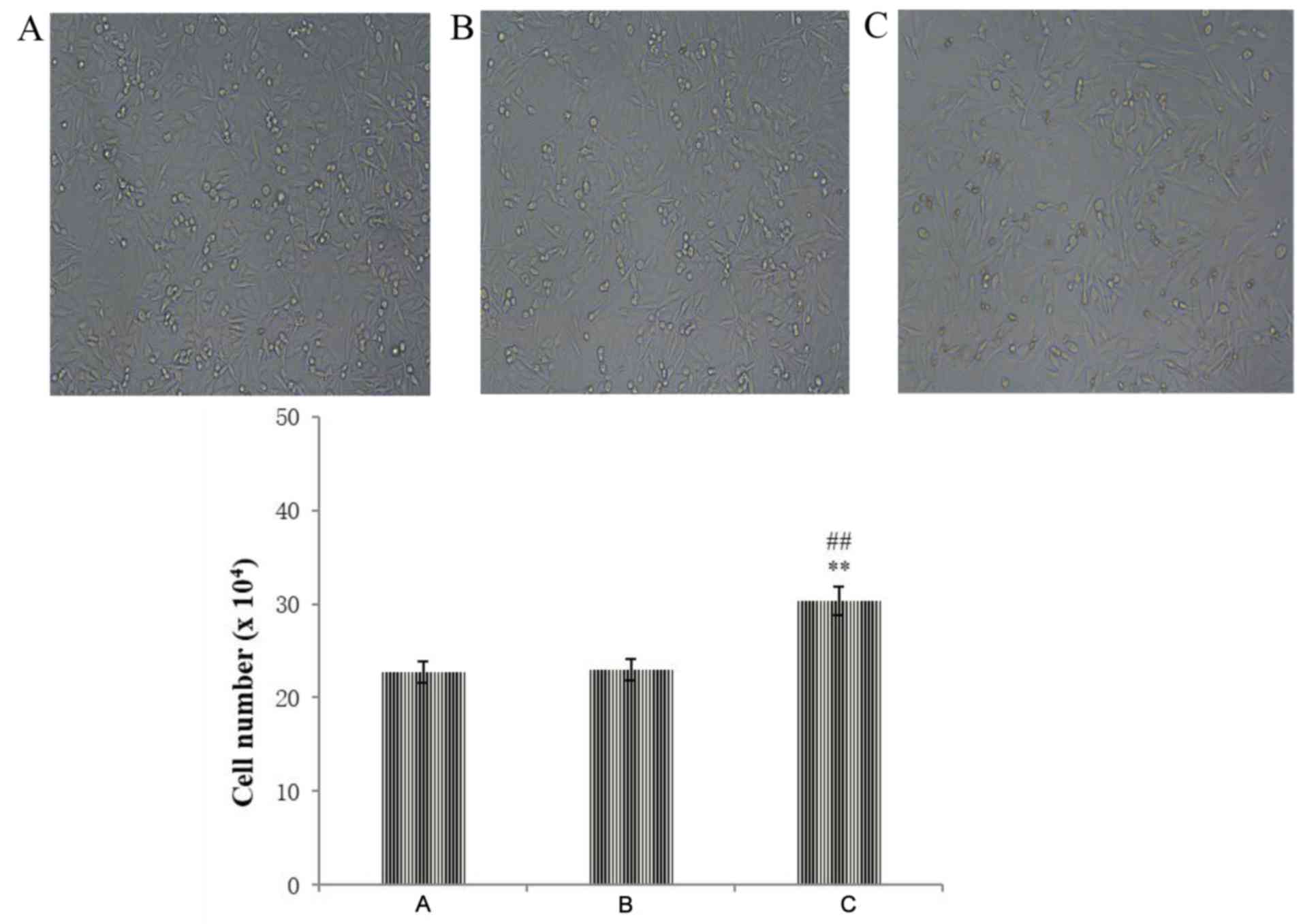
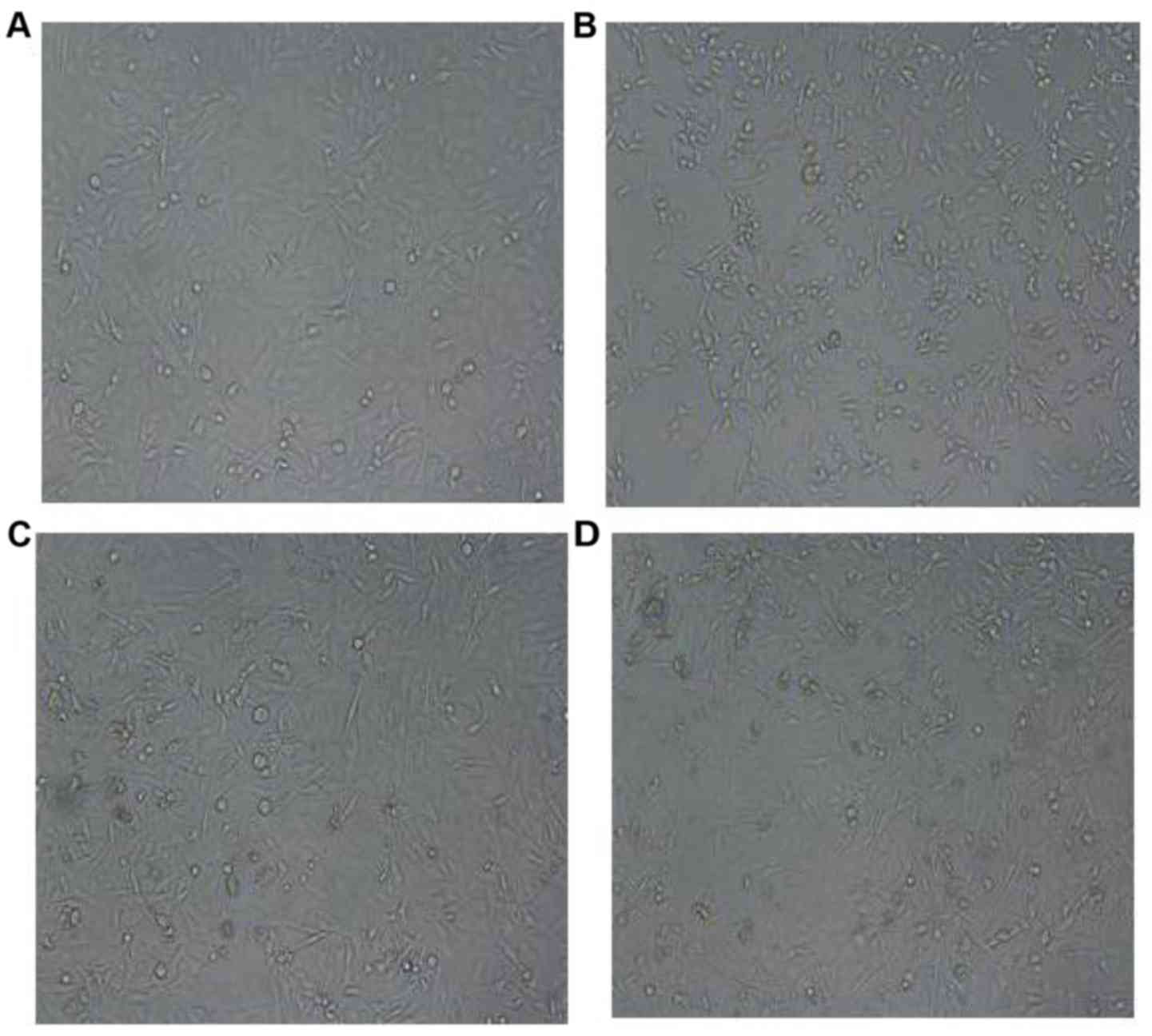
Effect of LKB1 siRNA on the morphology
of LoVo cells
After transfection, the morphological changes in the
different groups were observed by inverted microscope. There was no
significant difference between the negative control group and the
blank control group. The apoptotic cells in the present study refer
to cells that can be seen to float in the culture solution.
Compared with the negative control group, cell proliferation was
increased and the number of cells was increased in the LKB1 siRNA
group, but there was a small number of apoptotic cells, indicating
toxicity to some cells (Fig.
2).
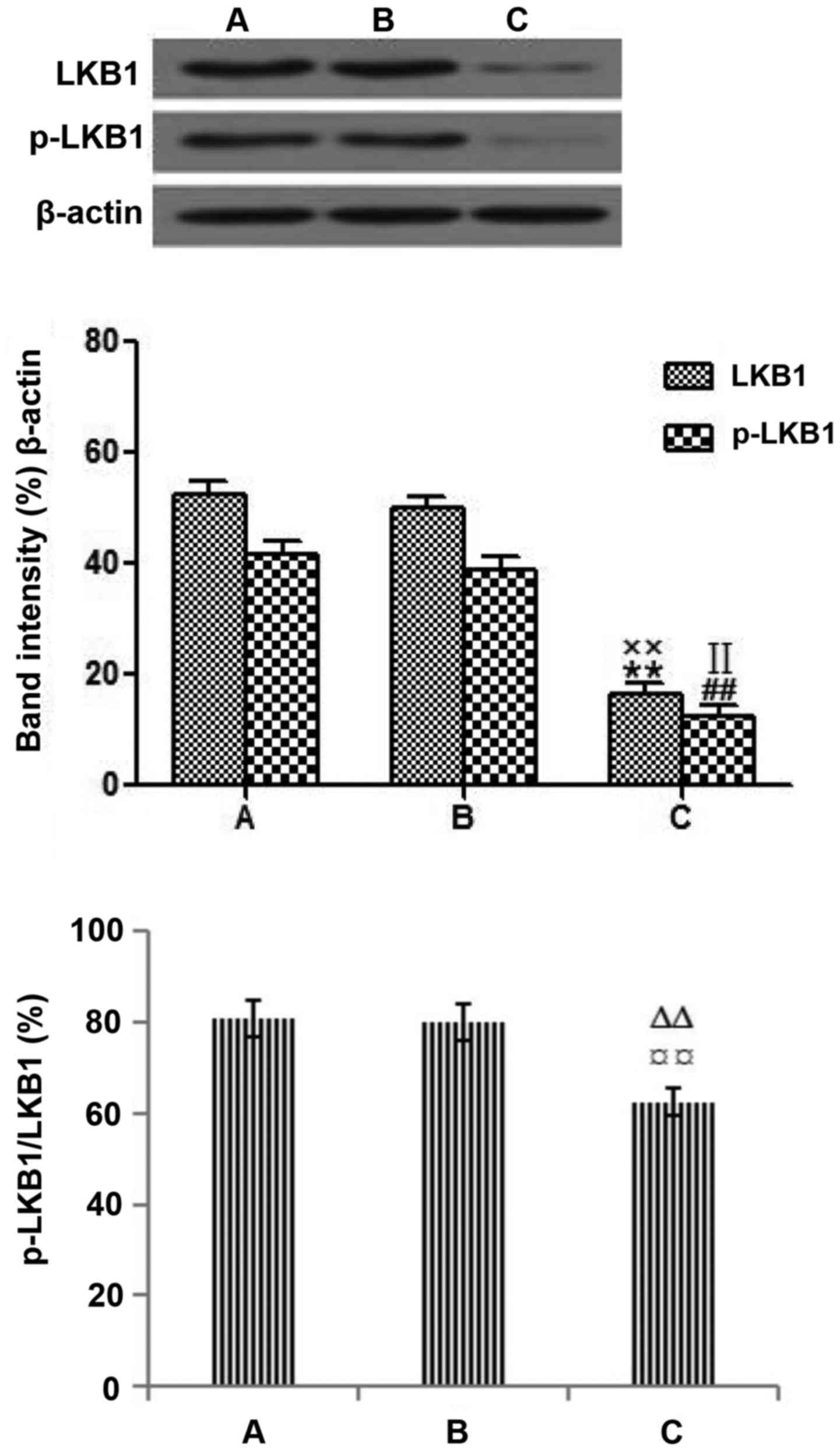
Western blotting detection of
transfected cells
Following transfection of the LoVo cells with LKB1
siRNA for 48 h, western blotting was performed. LKB1 siRNA was
synthesized by Guangzhou RiboBio Co., Ltd. The company tested that
siRNA 001 had the best transfection efficiency. According to the
company's synthetic results, the present study used siRNA-001 (data
not shown). The expression levels of LKB1 and p-LKB1 in the LKB1
siRNA group were markedly decreased compared with those in the
blank control group. Results indicate that p-LKB1/LKB1 expression
levels were lower in the LKB1 siRNA group, compare to the negative
control group (Fig. 3). The results
confirmed that LKB1 siRNA had been successfully transfected into
the LoVo cells.
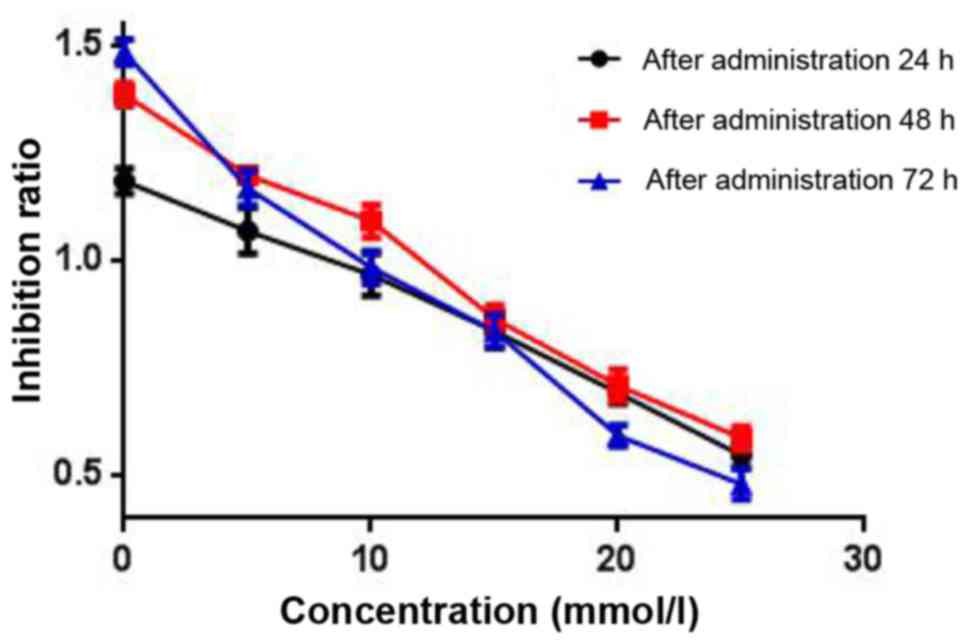
Effect of metformin on cell
proliferation as detected by CCK-8 assay
Metformin significantly inhibited the proliferation
of LoVo cells in a dose- and time-dependent manner. The
IC50 of metformin was 20 mmol/l after 48 h of treatment
(Fig. 4, Table III).
| Table III.Inhibitory rate of cell growth of
LoVo cells treated with metformin by CCK-8 assay. |
Table III.
Inhibitory rate of cell growth of
LoVo cells treated with metformin by CCK-8 assay.
| Group | 24 h | 48 h | 72 h |
|---|
| Blank control
group | 0.000+0.000 | 0.00+0.00 | 0.00+0.00 |
| Negative control
group | 1.186+0.029 | 1.388+0.027 | 1.486+0.029 |
| Metformin group
(mmol/l) |
| 5 |
1.070+0.052a |
1.199+0.099a,b |
1.169+0.040a,b |
| 10 |
0.967+0.049a,c |
1.093+0.038a–c |
0.984+0.038a–c |
| 15 |
0.084+0.038a,c |
0.086+0.031a–c |
0.084+0.038a–c |
| 20 |
0.069+0.025a,c |
0.513+0.037a–c |
0.710+0.025a–c |
| 25 |
0.547+0.027a,c |
0.587+0.028a–c |
0.480+0.035a–c |
Effect of metformin on the morphology
of LoVo cells
Compared with the blank control group, treatment
with metformin resulted in apoptotic morphological changes in LoVo
cells. Under the light microscope, we observed that the shape of
apoptotic cells became smaller, rounded and the adherent cells
shrank and desquamated. The LKB1 siRNA group indicated that
deletion of LKB1 led to cell proliferation (Fig. 5).
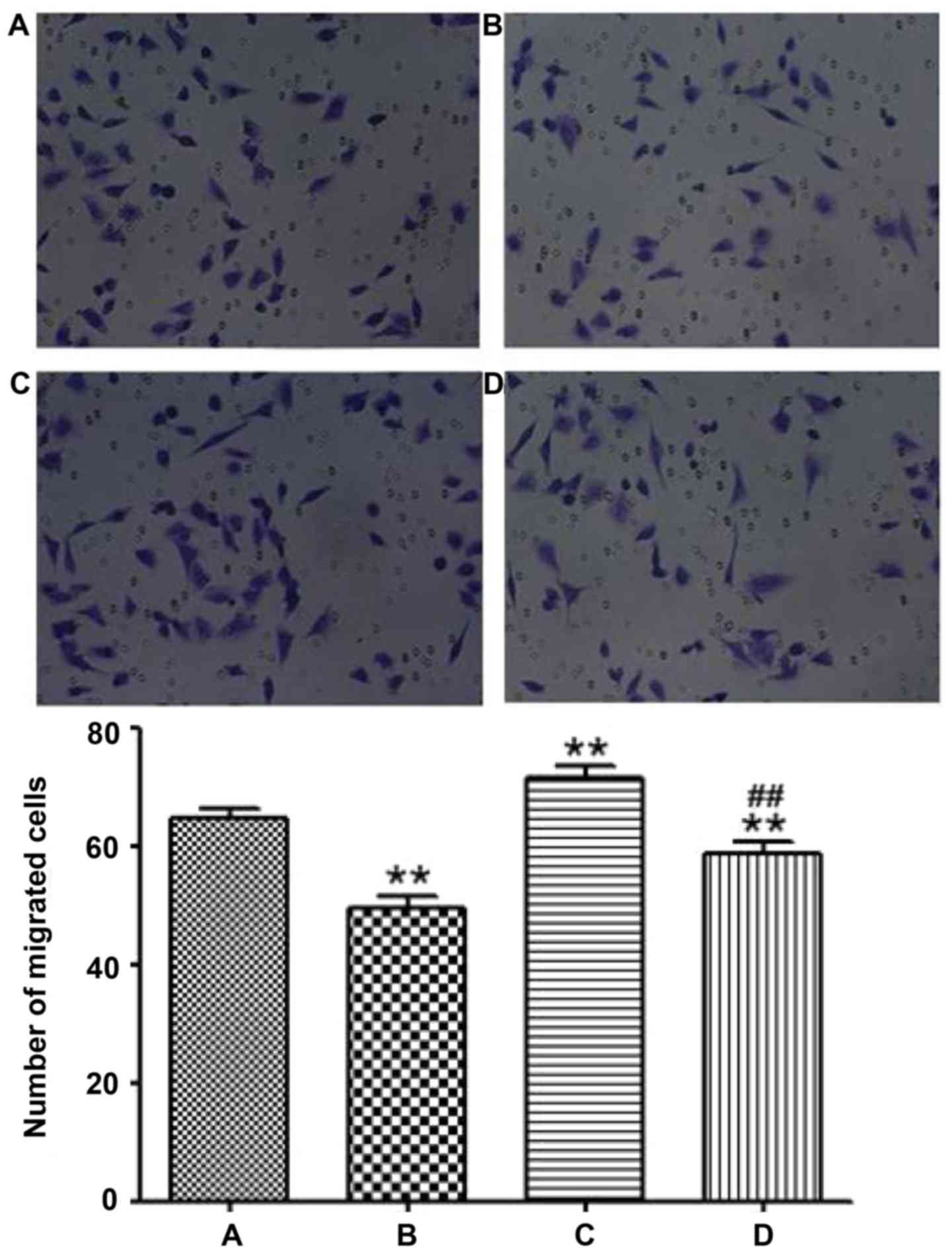
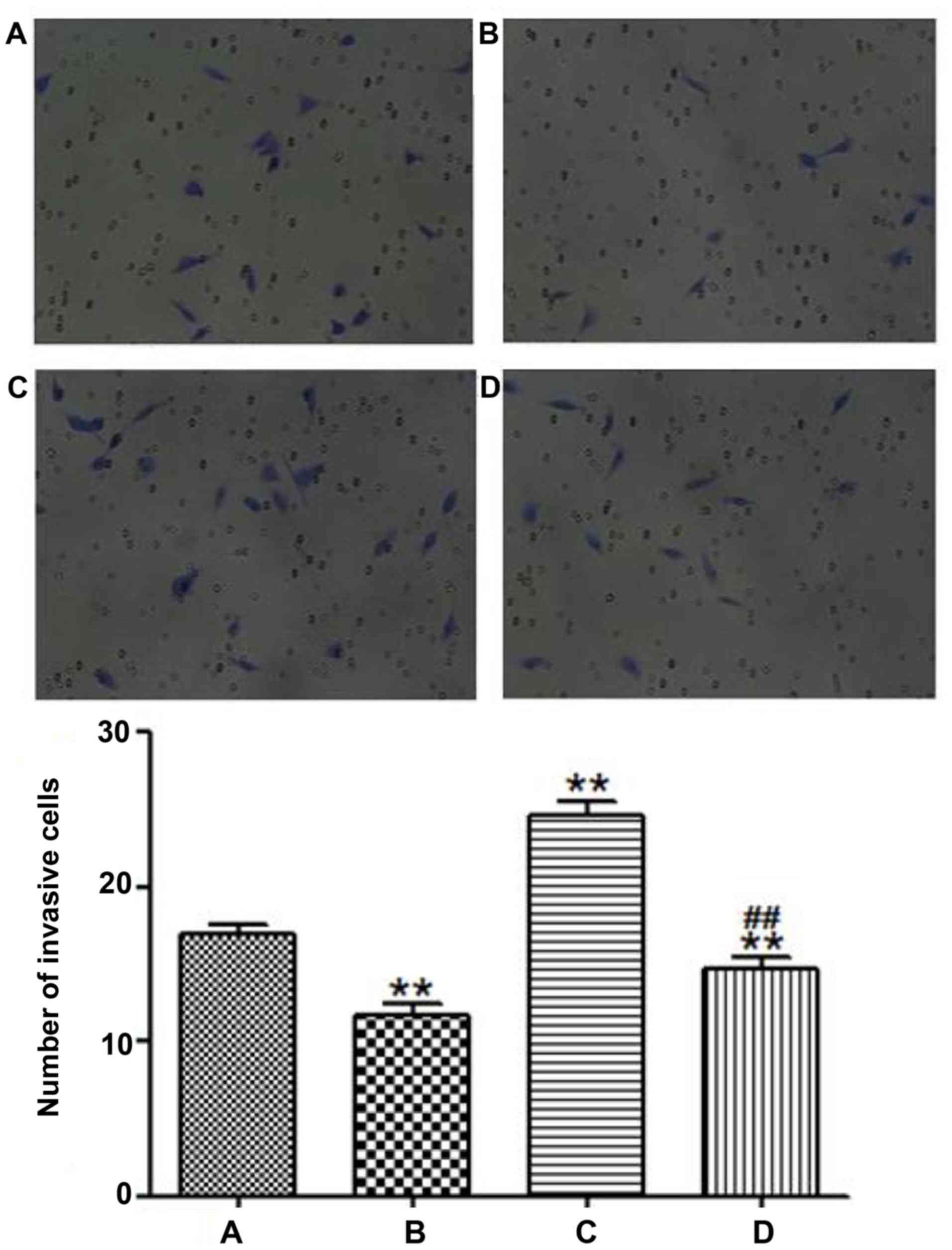
Effects of LKB1 on the migration and
invasion of LoVo cells
A Transwell chamber assay was used to determine the
migration and invasion abilities of LoVo cells. As shown in
Figs. 6 and 7, the migration and invasion capabilities
were clearly increased in the LKB1 siRNA group compared with the
blank control group (P<0.01). The results demonstrated that the
deletion of LKB1 increased the migration and invasion capacities of
CRC cells. The results of the Transwell chamber experiments in the
study indicated that the migration and invasion abilities of the
CRC cells with deletion of LKB1 were enhanced, which indicated that
the metastasis of CRC was enhanced.
To further explore whether the LKB1 signaling
pathway is associated with the migration and invasion of LoVo
cells, LoVo cells were treated with 20 mmol/l metformin for 48 h.
As shown in Figs. 6 and 7, the migration and invasion capacities
were decreased in the metformin group compared with the blank
control group. The migration and invasion capacities were decreased
in the LKB1 siRNA+metformin group compared with the LKB1 siRNA
group. The results of the present study suggest that metformin may
be associated with the inhibition of migration and
invasion-associated protein expression in LoVo cells.
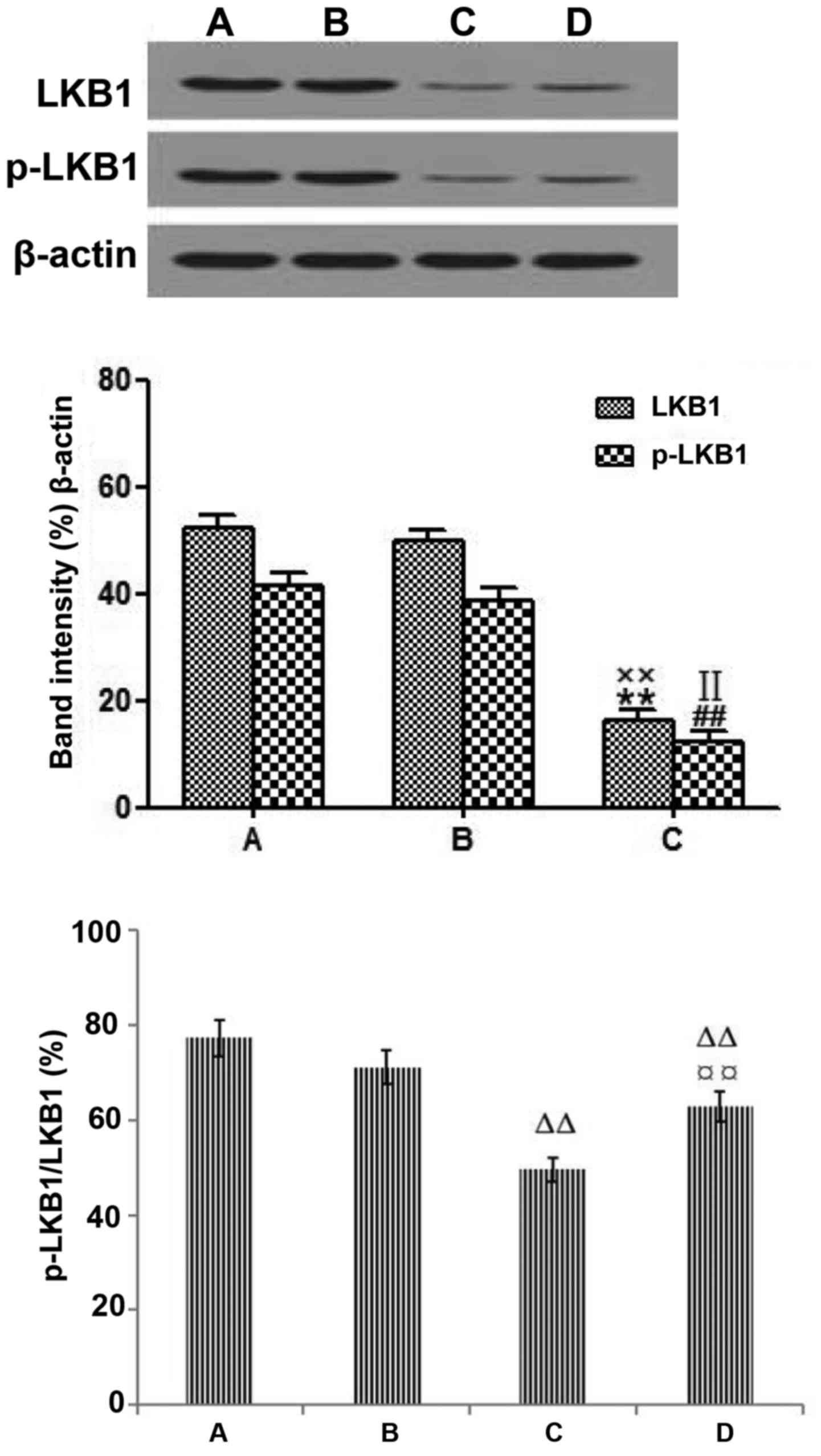
Effects of metformin on the expression
of LKB1and p-LKB1 in LoVo cells
To confirm the effect of metformin on the expression
of LKB1 and p-LKB1 in LoVo cells, western blot analysis was
performed. The protein expression levels of LKB1, p-LKB1 and
p-LKB1/LKB1 were significantly reduced in the LKB1 siRNA+metformin
group compared with the blank control group (P<0.01). The
protein expression levels of LKB1, p-LKB1 and p-LKB1/LKB1 were
significantly increased in the LKB1 siRNA+metformin group compared
with the LKB1 siRNA group (P<0.01; Fig. 8).
Effects of metformin on the expression
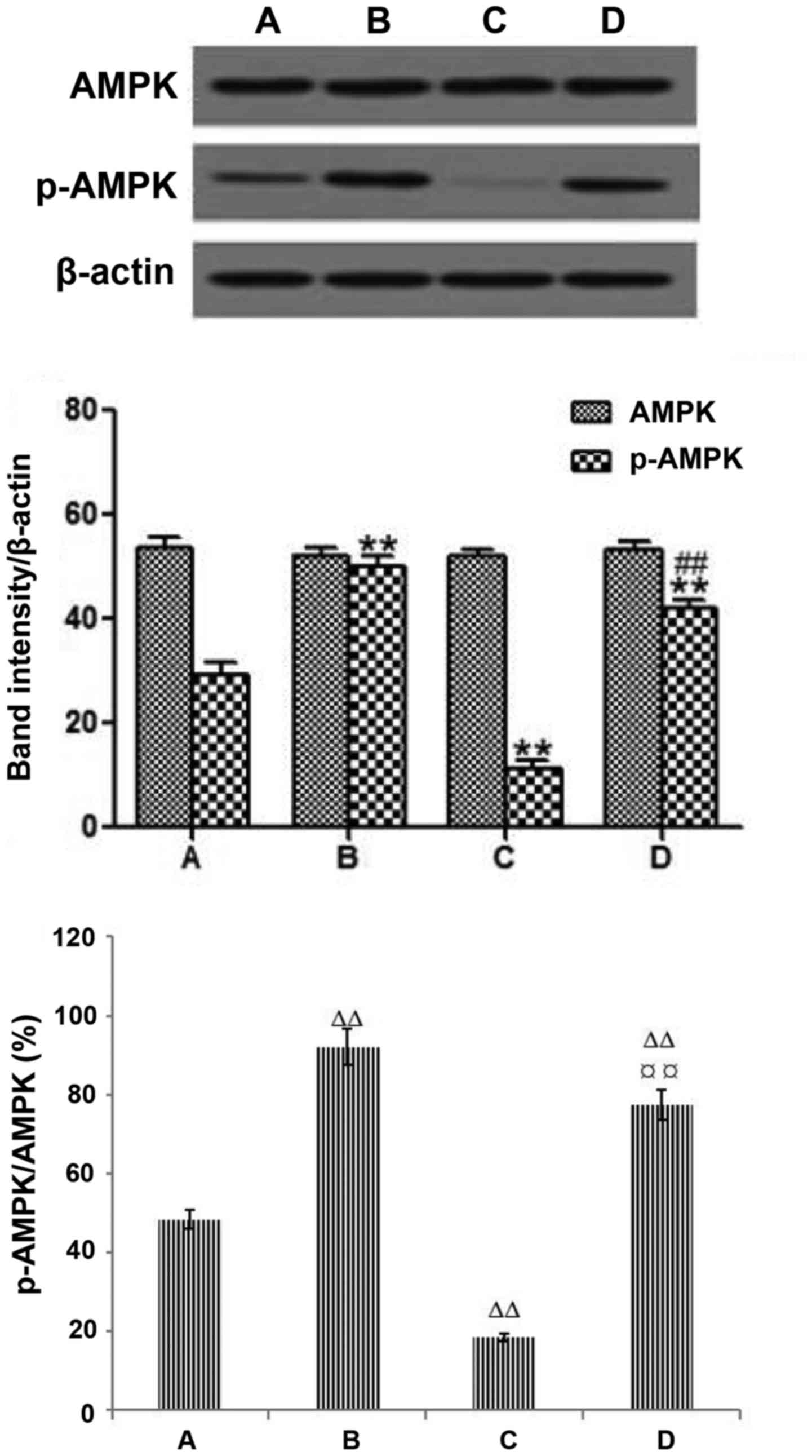
of AMPK and p-AMPK in LoVo cells
Western blot analysis showed that the expression of
AMPK was not significantly different among the 4 groups
(P>0.05). The expression level of p-AMPK was significantly
increased in the metformin group compared with the blank control
group (P<0.01). The expression of p-AMPK and p-AMPK/AMPK were
significantly decreased in the LKB1 siRNA group compared with the
blank control group (P<0.01). However, the expression level of
p-AMPK and p-AMPK/AMPK in the LKB1 siRNA+metformin group were much
higher than that in the blank control group, suggesting that the
expression of p-AMPK may play a role in CRC. Treatment with
metformin increased the protein expression of p-AMPK compared with
that of the LKB1 siRNA group, but it did not increase the
expression of AMPK in the LKB1 siRNA+metformin group, which
indicated that the effect of metformin on the p-AMPK protein
expression was dependent on LKB1 (P<0.01; Fig. 9). These results suggest that further
investigation of the biological functions of the activation of AMPK
in CRC is needed.
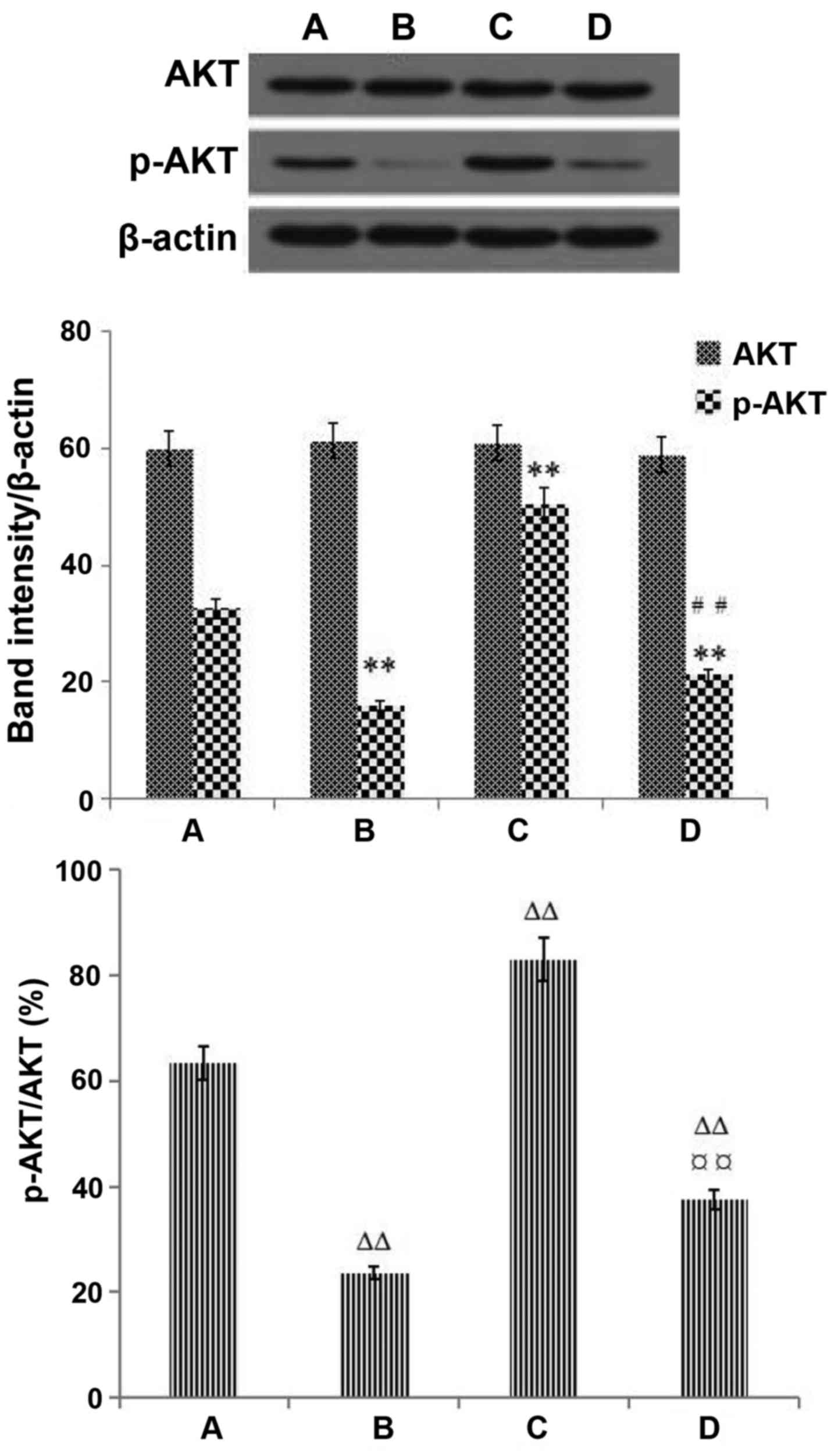
Effects of metformin on the expression
of AKT and p-AKT in LoVo cells
Western blot analysis showed that the expression of
AKT was not significantly different among the 4 groups (P>0.05).
Treatment with metformin decreased the protein expression of p-AKT
compared with that of the blank control group. The LKB1 siRNA group
had increased protein expression of p-AKT. The protein expression
of p-AKT and p-AKT/AKT were reduced in the LKB1 siRNA+metformin
group compared with the LKB1 siRNA group. These results indicated
that metformin has an effect on p-AKT protein expression and
suggested that the AKT signaling pathway may be associated with the
metformin-induced changes in the expression of migration and
invasion-associated proteins in LoVo cells (Fig. 10).
Discussion
LKB1 gene expression is closely related to the
development and metastasis of colorectal cancer (CRC). By detecting
the expression of LKB1 in CRC, we can provide a reference index for
diagnosing the depth of invasion and lymph node metastasis. It has
been reported that the mutation rate of LKB1 in sporadic left colon
cancer is 52.6% (26). In the
present study, IHC results showed that expression of LKB1 as a
tumor-suppressor gene was strongly positive in normal colorectal
tissue, indicating that it can inhibit the occurrence of CRC. The
expression of LKB1 was reduced in inflammatory polyp tissue.
However, the expression of LKB1 was significantly decreased in
adenocarcinoma and mucinous adenocarcinoma tissues, inducing the
invasion and metastasis of CRC. These results provide a reference
for further investigation of the role of LKB1 in the invasion and
metastasis of CRC cells from the perspective of cytology.
New methods based on siRNA can effectively reduce
tumor cell volume and inhibit tumor cell repair. siRNA technology
has brought new possibilities for gene therapy for CRC patients
(13). In the present study, LKB1
siRNA was transfected into LoVo cells to observe the effect of LKB1
on the invasion and metastasis of CRC and the potential underlying
mechanism of that effect. As an important tumor-suppressor gene,
LKB1 plays a key regulatory role in the LKB1/AMPK/AKT signaling
pathway. The results of the western blot analysis showed that the
deletion of LKB1 led to a decrease in p-AMPK in the downstream
pathway, which promoted p-AKT, tumor cell proliferation and
angiogenesis and enhanced the migration and invasion of CRC cells.
The present study used CRC cells to observe the effect of LKB1 on
invasion and metastasis of CRC cells. This in vitro
experiment had limitations. Further reseach will be carried out by
us in in vivo experiments to observe the effect of LKB1 on
CRC metastasis in nude mice.
Studies have shown that metformin-induced AMPK
activation can reduce protein synthesis and cell proliferation in
CRC (27,28). Metformin induces cell cycle arrest
or apoptosis, activates the immune system, improves immunity and
eliminates cancer stem cells. Increasing numbers of clinical
studies have shown that metformin as a typical adjuvant
chemotherapy can reduce the risk of invasion and metastasis in
cancer patients, resulting in more successful chemotherapy
(29). Therefore, we used metformin
treatment to observe the effect of metformin on the motility and
invasiveness of CRC cells to investigate the possible mechanism by
which LKB1 acts on the AMPK-AKT signaling pathway in CRC
metastasis. Metformin-induced AMPK activation reduced protein
synthesis and cell proliferation and inhibited p-AKT production in
CRC cells, thereby inhibiting tumor metastasis, promoting tumor
cell apoptosis and decreasing tumor cell migration and invasion.
Interestingly, our experiments showed that metformin did not
directly activate the phosphorylation of LKB1 (p-LKB1). Metformin
inhibited the invasion and metastasis of CRC by activating p-AMPK,
thereby inhibiting the activation of p-AKT.
In conclusion, our findings demonstrates that LKB1
plays an important role in the invasion and metastasis of CRC by
activating AMPK, negatively regulating the AKT signaling pathway
and regulating gene expression, which may be a new
anti-angiogenesis and antimetastasis target in CRC treatment.
Mutation or deletion of LKB1 repair is expected to be a new
therapeutic target or clinical biomarker for the treatment of the
invasion and metastasis of CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Scientific Research Staring Foundation for Returned Overseas
Chinese Scholars, the Ministry of Education of China [grant no.
(2013) 1792], the Hubei Province Health and Family Planning
Commission Research Projects (grant no. WJ2015MB236), and the
Xianning Municipal Science and Technology Research and Development
Project [grant no. (2013) 28].
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
YZ designed the experiment, analyzed and explained
the data, modified the manuscript, and approved the final version
of the manuscript. YC performed the cell experiments and data
analysis and interpretation and drafted the manuscripts. YL
conceived the experiments, obtained, compiled and analyzed
experimental data, participated in the drafting of the manuscripts
and approved the final manuscript. HY performed the
immunohistochemistry experiments and carried out the data analysis.
All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Hubei University of Science and Technology
(Xianning, China). The committee's reference no. is 201601001. The
colon cancer tissue microarray was purchased from Fanpu Biotech,
Inc. The company ensured ethical approval from the patients, and
patient consent for publication; Clinical Research Protocol
(version no. 2016LC001) and informed consent (version no.
2016TYS001).
Patient consent to publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang M, Miao F, Huang R, Liu W, Zhao Y,
Jiao T, Lu Y, Wu F, Wang X, Wang H, et al: RHBDD1 promotes
colorectal cancer metastasis through the Wnt signaling pathway and
its downstream target ZEB1. J Exp Clin Cancer Res. 37:222018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hawley SA, Pan DA, Mustard KJ, Ross L,
Bain J, Edelman AM, Frenguelli BG and Hardie DG:
Calmodulin-dependent protein kinase kinase-beta is an alternative
upstream kinase for AMP-activated protein kinase. Cell Metab.
2:9–19. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dorward HS, Du A, Bruhn MA, Wrin J, Pei
JV, Evdokiou A, Price TJ, Yool AJ and Hardingham JE:
Pharmacological blockade of aquaporin-1 water channel by AqB013
restricts migration and invasiveness of colon cancer cells and
prevents endothelial tube formation in vitro. J Exp Clin Cancer
Res. 35:362016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Slattery ML, Herrick JS, Mullany LE,
Samowitz WS, Sevens JR, Sakoda L and Wolff RK: The co-regulatory
networks of tumor suppressor genes, oncogenes, and miRNAs in
colorectal cancer. Genes Chromosomes Cancer. 56:769–787. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Eshghifar N, Farrokhi N, Naji T and Zali
M: Tumor suppressor genes in familial adenomatous polyposis.
Gastroenterol Hepatol Bed Bench. 10:3–13. 2017.PubMed/NCBI
|
|
7
|
Tiainen M, Vaahtomeri K, Ylikorkala A and
Mäkelä TP: Growth arrest by the LKB1 tumor suppressor: Induction of
p21WAF1/CIP1. Hum Mol Genet. 11:1497–1504. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ylikorkala A, Rossi DJ, Korsisaari N,
Luukko K, Alitalo K, Henkemeyer M and Mäkelä TP: Vascular
abnormalities and deregulation of VEGF in
Lkb1-deficient mice. Science. 293:1323–1326. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hardie DG and Alessi DR: LKB1 and AMPK and
the cancer-metabolism link-ten years after. BMC Biol. 11:362013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jeon SM and Hay N: The double-edged sword
of AMPK signaling in cancer and its therapeutic implications. Arch
Pharm Res. 38:346–357. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Larsen AK, de Gramont A, Poindessous V,
Bouygues A, Ayadi M and Mésange P: Functions and clinical
implications of autocrine VEGF signaling in colorectal cancer. Curr
Colorect Cancer Rep. 9:270–277. 2013. View Article : Google Scholar
|
|
12
|
Deguchi A, Miyoshi H, Kojima Y, Okawa K,
Aoki M and Taketo MM: LKB1 suppresses p21-activated kinase-1 (PAK1)
by phosphorylation of Thr109 in the p21-binding domain.
J Biol Chem. 285:18283–18290. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim SW: The role of MicroRNAs in
colorectal cancer. Korean J Gastroenterol. 69:206–211. 2017.(In
Korean). View Article : Google Scholar : PubMed/NCBI
|
|
14
|
You HX, Zhou YH, Tan SY and She TH:
Effects of silencing RIP1 with siRNA on the biological behavior of
the LoVo human colon cancer cell line. Oncol Lett. 7:2065–2072.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He G, Zou L, Zhou L, Gao P, Qian X and Cui
J: Cysteine-rich intestinal protein 1 silencing inhibits migration
and invasion in human colorectal cancer. Cell Physiol Biochem.
44:897–906. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zakikhani M, Dowling RJ, Sonenberg N and
Pollak MN: The effects of adiponectin and metformin on prostate and
colon neoplasia involve activation of AMP-activated protein kinase.
Cancer Prev Res. 1:369–375. 2008. View Article : Google Scholar
|
|
17
|
Song X, Kim SY, Zhang L, Tang D, Bartlett
DL, Kwon YT and Lee YJ: Role of AMP-activated protein kinase in
cross-talk between apoptosis and autophagy in human colon cancer.
Cell Death Dis. 5:e15042014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lipovka Y and Konhilas JP: AMP-activated
protein kinase signalling in cancer and cardiac hypertrophy.
Cardiovasc Pharm Open Access. 4:1542015.PubMed/NCBI
|
|
19
|
Xu H, Zhou Y, Coughlan KA, Ding Y, Wang S,
Wu Y, Song P and Zou MH: AMPKα1 deficiency promotes cellular
proliferation and DNA damage via p21 reduction in mouse embryonic
fibroblasts. Biochim Biophys Acta. 1853:65–73. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Su KH, Yu YB, Hou HH, Zhao JF, Kou YR,
Cheng LC, Shyue SK and Lee TS: AMP-activated protein kinase
mediates erythropoietin-induced activation of endothelial nitric
oxide synthase. J Cell Physiol. 227:3053–3062. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Monteverde T, Muthalagu N, Port J and
Murphy DJ: Evidence of cancer-promoting roles for AMPK and related
kinases. FEBS J. 282:4658–4671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Wang B, Guerram M, Sun L, Shi W,
Tian C, Zhu X, Jiang Z and Zhang L: Deoxypodophyllotoxin suppresses
tumor vasculature in HUVECs by promoting cytoskeleton remodeling
through LKB1-AMPK dependent Rho A activation. Oncotarget.
6:29497–29512. 2015.PubMed/NCBI
|
|
23
|
Altomare DA and Testa JR: Perturbations of
the AKT signaling pathway in human cancer. Oncogene. 24:7455–7464.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brugarolas J and Kaelin WG Jr:
Dysregulation of HIF and VEGF is a unifying feature of the familial
hamartoma syndromes. Cancer Cell. 6:7–10. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Amin MB, Edge SB, Greene FL, Byrd DR,
Brookland RK, Washington MK, Gershenwald JE, Compton CC, Hess KR,
Sullivan DC, et al: AJCC Cancer Staging Manual. (8th). 2018.
|
|
26
|
Dong SM, Kim KM, Kim SY, Shin MS, Na EY,
Lee SH, Park WS, Yoo NJ, Jang JJ, Yoon CY, et al: Frequent somatic
mutations in serine/threonine kinase 11/Peutz-Jeghers syndrome gene
in left-sided colon cancer. Cancer Res. 58:3787–3790.
1998.PubMed/NCBI
|
|
27
|
Li W, Wang QL, Liu X, Dong SH, Li HX, Li
CY, Guo LS, Gao JM, Berger NA, Li L, et al: Combined use of vitamin
D3 and metformin exhibits synergistic chemopreventive effects on
colorectal neoplasia in rats and mice. Cancer Prev Res. 8:139–148.
2015. View Article : Google Scholar
|
|
28
|
Zaafar DK, Zaitone SA and Moustafa YM:
Role of metformin in suppressing 1,2-dimethylhydrazine-induced
colon cancer in diabetic and non-diabetic mice: Effect on tumor
angiogenesis and cell proliferation. PLoS One. 9:e1005622014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu CY, Wang CJ, Tseng CC, Chen HP, Wu MS,
Lin JT, Inoue H and Chen GH: Helicobacter pylori promote gastric
cancer cells invasion through NF-kappaB and COX-2-mediated pathway.
World J Gastroenterol. 11:3197–3203. 2005. View Article : Google Scholar : PubMed/NCBI
|