Introduction
Oncolytic viruses (OVs) are genetically engineered
viruses that selectively replicate in and kill cancer cells, and
represent a novel type of antitumor therapy (1–4). This
approach has numerous advantages as a cancer therapeutic agent due
to its mechanism-based selectivity, potential for mediating tumor
cell death and possibility of expressing additional therapeutic
transgenes at the tumor site (5,6). Since
OVs are designed for intratumoral injection, this unique cancer
therapeutic is often coupled with the antitumor immunity
(immunovirotherapy) of the host. The herpes simplex virus type 1
(HSV-1)-based oncolytic HSV (oHSV) talimogene laherparepvec (T-VEC,
Imlygic) was the first US Food and Drug Administration-approved OV
(7). As with other OV candidates,
the host response against oHSV is complex, multifaceted, and
modulated by host immunity and the tumor microenvironment. These
various immune and inflammatory responses may be beneficial and
detrimental (8,9), among which, the induction of cytokine
storm by OV delivery to particular organs (such as lung) is one of
the increasingly recognized impediments. The term cytokine storm
was first used to describe influenza-induced cytokine
overproduction in a short time, and is associated with uncontrolled
pro-inflammatory responses and significant immunopathology and
severe disease outcomes (10). The
development of cytokine storm with attendant pulmonary damage has
subsequently been reported in various viral, bacterial and fungal
infections (11). Consistent
observations indicated that the concept of cytokine storm was much
more complicated; however, our understanding of the mechanisms that
promote cytokine storm remains limited, and countermeasures to
control the balance between appropriate cytokine release and
cytokine overproduction are relatively unexplored. The challenge of
oHSV as an efficacious oncolytic agent is the potential tissue
damage that is induced by a range of cytokine production following
oHSV infection. Furthermore, the weakened immunity and unbalanced
immune homeostasis of patients with tumors would exacerbate the
situation. Therefore, understanding the molecular mechanism of
cytokine storm caused by oHSV injection and developing a strategy
to control the hazards of cytokine storm following oHSV treatment
become critically urgent.
The signaling cascade of a number of cytokines pass
through the Janus kinase (JAK)/signal transducer and activator of
transcription (STAT) pathway, and the suppressors of cytokine
signaling (SOCS) proteins are pivotal negative regulators of the
JAK/STAT signaling pathway that are part of cytokine networks
(12). It has been identified that
SOCS4-deficient mice exhibited increased lethality, which is
associated with increased levels of pro-inflammatory cytokines in
the lungs following influenza A virus infection (13). Accordingly, SOCS4 was postulated to
be a critical regulator of antiviral immunity. However, very little
is known regarding its function during oHSV infection.
In the present study, an HSV strain was constructed
with a SOCS4 protein insert (HSV-SOCS4) to investigate its
suppression of cytokine storm. The present study focused on several
representative cytokines that serve key functions, including
monocyte chemoattractant protein 1 (MCP-1), interleukin (IL)-1β,
tumor necrosis factor α (TNF-α), IL-6 and interferon γ (IFN-γ), and
identified that their concentrations were decreased in
HSV-SOCS4-infected mice compared with those in HSV-1(F)-infected
mice, and HSV-SOCS4-infected mice exhibited slight lung damage,
decreased weight loss and a 100% survival rate. The results of the
present study indicate that HSV-SOCS4 may provide a possible
solution to control the cytokine storm induced by oncolytic HSV
therapy.
Materials and methods
Animals
A total of 60 female BALB/c mice, aged 6 weeks, were
purchased from the Experiment Animal Center of Guangdong
(Guangzhou, China) and were housed at 22±2°C, ammonia ≤14 ppm, 12-h
light/12-h dark cycle, and free access to food and water, free from
microbial pathogens at the Animal Center of Guangzhou Medical
University (Guangzhou, China). All procedures involving mice were
approved by the Institutional Animal Care and Use Committee of
Guangzhou Medical University.
Cells and virus strains
Vero cells were obtained from the American Type
Culture Collection (Manassas, VA, USA) and were cultured in
Dulbecco's modified Eagle's medium (high glucose) supplemented with
5% (v/v) fetal bovine serum or 5% (v/v) newborn calf serum (NBCS),
respectively. All media were from HyClone; GE Healthcare (Logan,
UT, USA). HSV-1(F), the prototype HSV-1 strain used in our
laboratory (14), was propagated
and titrated on Vero cells. pReceiver-M02 with human SOCS4 mRNA was
purchased from GeneCopoeia, Inc. (Rockville, MD, USA).
Construction of HSV recombinant virus
with SOCS4 expression
Two oligonucleotide primers were designed according
to the human SOCS4 gene sequence: Forward,
5′-GTCGACATGTGGTGGCGCCTGTGGTGGCTCTGCTGCTGTGGCCCATGGTGTGGGCCGCAGAAAATAATGAAAATATTAG-3′
(with an AccI site and underlined signal peptide Hmm38);
reverse, 5′-GCGGCCGCCTAGTGATGGTGATGGTGATGGCATTGCTGTTCTGGTGCATC-3′
(with a NotI site and underlined His tag). The polymerase
chain reaction (PCR) was performed in a total reaction volume of 50
µl for 30 cycles consisting of a denaturation step at 95°C for 1
min, a primer annealing step at 58°C for 30 sec and a primer
extension step at 72°C for 2 min with Taq DNA polymerase (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). The PCR product of
SOCS4 was ligated into T-easy plasmid and then was
transformed into XL-1blue cells (Tiangen Biotech Co., Ltd.,
Beijing, China) and sent to IGE Biotech Ltd. (Guangzhou, China) for
sequencing confirmation. The confirmed SOCS4 gene was
excised from the T-easy plasmid and ligated into the carrier pNEWUL
backbone site at ClaI/AccI and NotI (Fig. 1A). Next, the sequence between
BglII and PacI (including UL3, UL4 and
SOCS4) was cleaved from pNEWUL and cloned into plasmid
pKO5.1 at the same sites (Fig. 1B).
The recombinant plasmid was transformed into a bacterial artificial
chromosome (BAC) and cultured on a Luria-Bertani (LB) plate with
chloramphenicol and zeocin at 43°C overnight. Next, the bacterial
colony was selected and enriched on an LB plate with
chloramphenicol and sucrose at 30°C overnight to excise plasmid
pKO5.1. Following identification by PCR amplification of the
SOCS4 gene (as aforementioned) and agarose (1%) gel
electrophoresis with ethidium bromide for visualization under UV,
and analysis using Image Lab software (version 2.1; MCM Design,
Hercules, CA, USA), the positive recombinant BAC was cultured to
proliferate and was transformed into Vero cells using
Lipofectamine® LTX with PLUS Reagent (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
cells were incubated until a viral cytopathic effect was observed.
Viral plaques were collected in 1 ml milk and then underwent three
freeze-thaw (−80°C for 10 min and 37°C for 2 min) cycles to release
virus. The virus was inoculated and cultured into Vero cells in
25-cm2 flasks, and DNA was extracted for PCR for final
confirmation (including the UL3, UL4 and SOCS4
genes). Oligonucleotide primers for UL3 were
5′-GGATAGCAGATGTGAGGAAGTC-3′ (forward) and
5′-ATGACACAGGCGCTCGGCATCG-3′ (reverse) and for UL4 were
5′-GTATACCACCACCGTCGACATT-3′ (forward) and
5′-GCGTTAAGGGGTCCGTTGTGTT-3′ (reverse). PCR of UL3 and
UL4 was performed in a total reaction volume of 50 µl for 30
cycles consisting of a denaturation step at 95°C for 1 min, a
primer annealing step at 55°C for 30 sec and a primer extension
step at 72°C for 2 min with Taq DNA polymerase (Thermo Fisher
Scientific, Inc.), and PCR amplification of the SOCS4 gene
was as aforementioned. All PCR products were identified with 1.0%
agarose gel electrophoresis with ethidium bromide for visualization
under UV and Image Lab software (version 2.1). Expression of SOCS4
protein was determined by western blot analysis. Briefly, the
recombinant virus-infected Vero cells were collected and
centrifuged at 150 × g for 20 min at 4°C. The cell pellet was
resuspended in 200 µl cell lysis buffer (cat. no. P0013;
Beyotime Institute of Biotechnology, Haimen, China) for 30
min on ice, and then the supernatant was collected and proteins
were quantified using a Bicinchoninic Acid kit (cat no. P0010;
Beyotime Institute of Biotechnology), according to the
manufacturer's protocol. Proteins (60 µg/lane) were separated by
SDS-PAGE (10% gel). Subsequently, proteins were transferred onto a
polyvinylidene difluoride (PVDF) membrane (cat. no. ISEQ00010; EMD
Millipore, Billerica, MA, USA). The PVDF membrane was incubated for
2 h at room temperature in blocking solution (5% non-fat milk in
PBS containing 0.05% Tween-20). Following washing, the PVDF
membrane was incubated overnight at 4°C in solution [PBS containing
0.05% Tween-20 and 1.0% bovine serum albumin (BSA; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany)] containing 1:1,000 diluted
anti-His-tag antibody (cat. no. 66005; ProteinTech Group, Inc.,
Chicago, IL, USA). The PVDF membrane was washed and incubated for 1
h at room temperature in the aforementioned solution containing
1:10,000 diluted horseradish peroxidase-conjugated goat anti-mouse
immunoglobulin G secondary antibody(cat. no. 31430; Thermo Fisher
Scientific, Inc.). Following washing, the result was determined
using an Enlight™ Western kit (cat. no. 29050; Engreen Biosystem
Co., Ltd., Auckland, New Zealand), according to the manufacturer's
protocol. The desired recombinant virus (named HSV-SOCS4) was
multiplied by culture, collected and stored at −80°C until further
use.
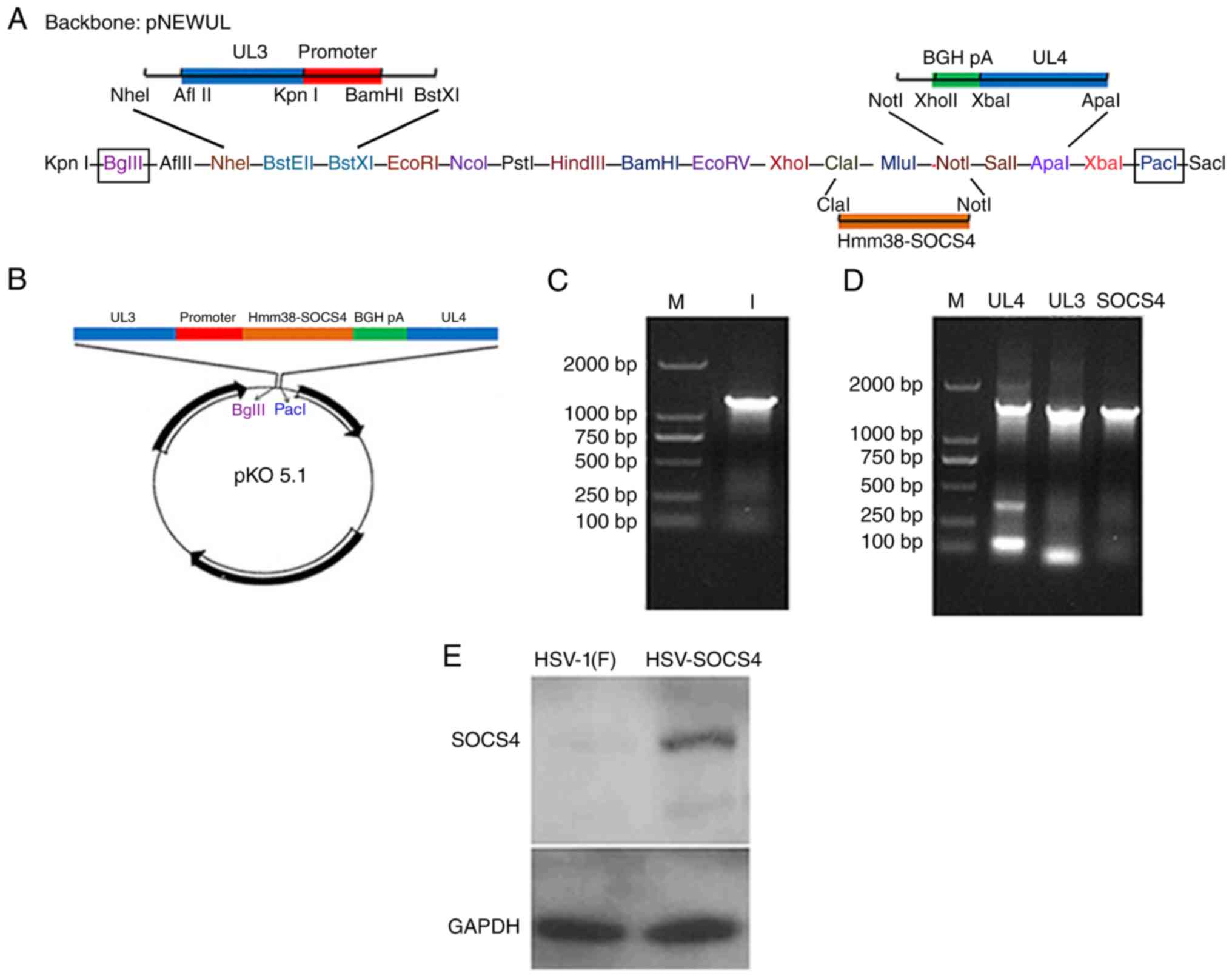 | Figure 1.Schematic representation, PCR
confirmation and western blot analysis of the HSV-SOCS4
recombinant. (A) The confirmed SOCS4 gene was ligated into
the pNEWUL backbone at ClaI/AccI and NotI
sites. (B) The sequence between BglII and PacI at the
pNEWUL backbone (including UL3, UL4 and SOCS4) was
cleaved from pNEWUL and cloned into plasmid pKO5.1 at the same
site. (C) The band in lane 1 is the PCR product of SOCS4 (1,397 bp)
from pReceiver-M02. (D) DNA was extracted from the reconstructed
virus to perform PCR for the final confirmation, and bands between
1,000 and 2,000 bp were identified as: UL4 (1,492 bp),
UL3 (1,319 bp) and SOCS4 (1,397 bp). (E) A specific
band of recombinant HSV-SOCS4 was observed, but not of HSV-1(F),
from western blot analysis. PCR, polymerase chain reaction; HSV,
herpes simplex virus; SOCS4, suppressor of cytokine signaling 4; M,
molecular mass markers. |
Infection of mice
BALB/c mice were divided randomly into three groups:
One group was infected with HSV-1(F), one group was infected with
HSV-SOCS4 and one group was treated with PBS as mock infection. In
detail, each mouse was lightly anaesthetized and then was infected
via the intranasal route with 106 plaque-forming units
of HSV-1(F) or HSV-SOCS4 in 30 µl PBS or treated with 30 µl PBS
only. Following infection, the mice were weighed and monitored for
morbidity and mortality every day for 12 days.
Serum and bronchoalveolar lavage fluid
samples collection and double sandwich ELISA
Orbital blood from each mouse was collected on days
1, 3 and 7 following infection, and the serum was separated and
then stored at −20°C for further ELISA. Following bleeding, the
mouse was sacrificed and the lung was flushed three times with 1 ml
PBS through a blunted needle inserted into the trachea to collect
bronchoalveolar lavage fluid (BALF). The samples were centrifuged
at 100 × g for 20 min at 4°C, the supernatant was removed for
ELISA, and the cells were collected for flow cytometric analysis.
For the BALF and serum samples, levels of a panel of cytokines,
including MCP-1 (cat. no. 1217392), IL-1β (cat. no. 1210122), IL-6
(cat. no. 1210602), TNF-α (cat. no. 1217202) and IFN-γ (cat. no.
1210002), were determined using a double sandwich ELISA (Dakewe
Biotech Co., Ltd., Shenzhen, China), and the concentration of each
cytokine was determined relative to the standard curve, according
to the manufacturer's protocol.
Cell isolation and flow cytometric
analysis
Cells harvested from BALF were treated with
Tris/NH4Cl to lyse erythrocytes, washed twice and
resuspended in ice-cold RPMI-1640 medium (Thermo Fisher Scientific,
Inc.). The spleen from each sacrificed mouse was eviscerated and
fully ground, and the tissue was rinsed through a sterile wire
screen. Spleen cell suspensions were collected, red blood cells
inside were lysed, and the remaining cells were washed and cell
pellet was resuspended in RPMI-1640 medium containing 1% NBCS. BALF
cells and spleen cells were counted and adjusted to
2×106 cells/ml. The cells were washed and stained for
the following surface markers: Allophycocyanin (APC)-cluster of
differentiation (CD)4 [APC-conjugated anti-mouse CD4 monoclonal
antibody (mAb), clone GK1.5; cat. no. 104412], fluorescein
isothiocyanate (FITC)-CD8a (FITC-conjugated anti-mouse CD8 mAb,
clone 53-6.7; cat. no. 100706) and phycoerythrin (PE)-CD62L
(PE-conjugated anti-mouse CD62L mAb, clone MEL-14; cat. no. 104408)
for spleen cells and Pacific Blue (PB)-CD11b (PB anti-mouse CD11b
mAb, clone M1/70; cat. no. 101224) for BALF cells. All antibodies
were purchased from BioLegend, Inc. (San Diego, CA, USA). Cells
were stained with 1:20 diluted antibody at 37°C for 10 min, and the
stained cells were washed with PBS containing 1% BSA and
resuspended in 2% formaldehyde in PBS for flow cytometric analysis
using a CytoFLEX flow cytometer (Beckman Coulter, Inc., Brea, CA,
USA) and CytExpert software (version 2.0; Beckman Coulter,
Inc.).
Lung samples for viral titration
analysis and pathological analysis
Following bleeding and sacrifice, the lungs of two
mice from each group were directly removed and minced completely
with cell culture medium; the tissue homogenate was collected and
centrifuged at 100 × g for 20 min at 4°C, and the supernatant was
collected for viral titration analysis. Vero cells were cultured on
6-well plates at 2×105 cells/well until 90% of cells
formed a monolayer, and the supernatant sample was added. After 24
h of incubation at 37°C, cells were removed carefully, centrifuged
as aforementioned and the cell pellet was resuspended in 1 ml milk,
and stored at −80°C. Following three freeze-thaw cycles as
aforementioned, the released virus sample was diluted 1:100,
1:1,000 and 1:10,000 with 1% NBCS. Next, 100 µl each dilution was
added to Vero cell monolayers on 6-well plates followed by culture
for 2 h, the medium was replaced with basal medium Eagle (Thermo
Fisher Scientific, Inc.) containing 1% NBCS and 0.5% immunoglobulin
G (cat. no. S20013001; Taibang Biological Products Co, Ltd.,
Shandong, China), and the cells were incubated for an additional 72
h. Subsequently, the cells were collected and stained with 0.03%
methylene blue to quantify the plaques.
Pathological analysis of the mouse lung (n=2,
without BALF collection) from HSV-1(F)-infected and
HSV-SOCS4-infected mice was performed by the Department of
Pathology at Guangzhou Medical University. The results were
observed using an Eclipse 80i confocal microscope (Nikon
Corporation, Tokyo, Japan).
Statistical analysis
Statistical analyses were performed using analysis
of variance and Tukey's honestly significant difference test using
SPSS statistical software (version 16.0; SPSS, Inc., Chicago, IL,
USA) for multiple comparisons. P<0.05 was considered to indicate
a statistically significant difference.
Results
Construction of HSV-SOCS4 recombinant
virus
The SOCS4 gene was inserted into a BAC to
construct the HSV-SOCS4. The SOCS4 gene PCR product and
sequencing identification confirmed the recombination. Fig. 1C presents the PCR product of
SOCS4 (1,397 bp) from pReceiver-M02, and the final PCR
confirmation of the fragments of UL4 (1,492 bp), UL3
(1,319 bp) and SOCS4 (1,397 bp) is presented in Fig. 1D. A specific band was observed for
HSV-SOCS4, but not for HSV-1(F), using western blot analysis
(Fig. 1E), suggesting that SOCS4
protein was expressed.
Cytokine production in BALF and serum
following virus infection
To analyze the effect of SOCS4 protein on cytokine
secretion, the profiles of the major inflammatory cytokines MCP-1,
IL-1β, IL-6, TNF-α and IFN-γ were determined in BALF and serum from
mice infected with PBS or HSV-1(F) or HSV-SOCS4 on days 1, 3 and 7
post-infection. Mock-infected mice did not induce appreciable
amounts of cytokines in either BALF or serum at any time point
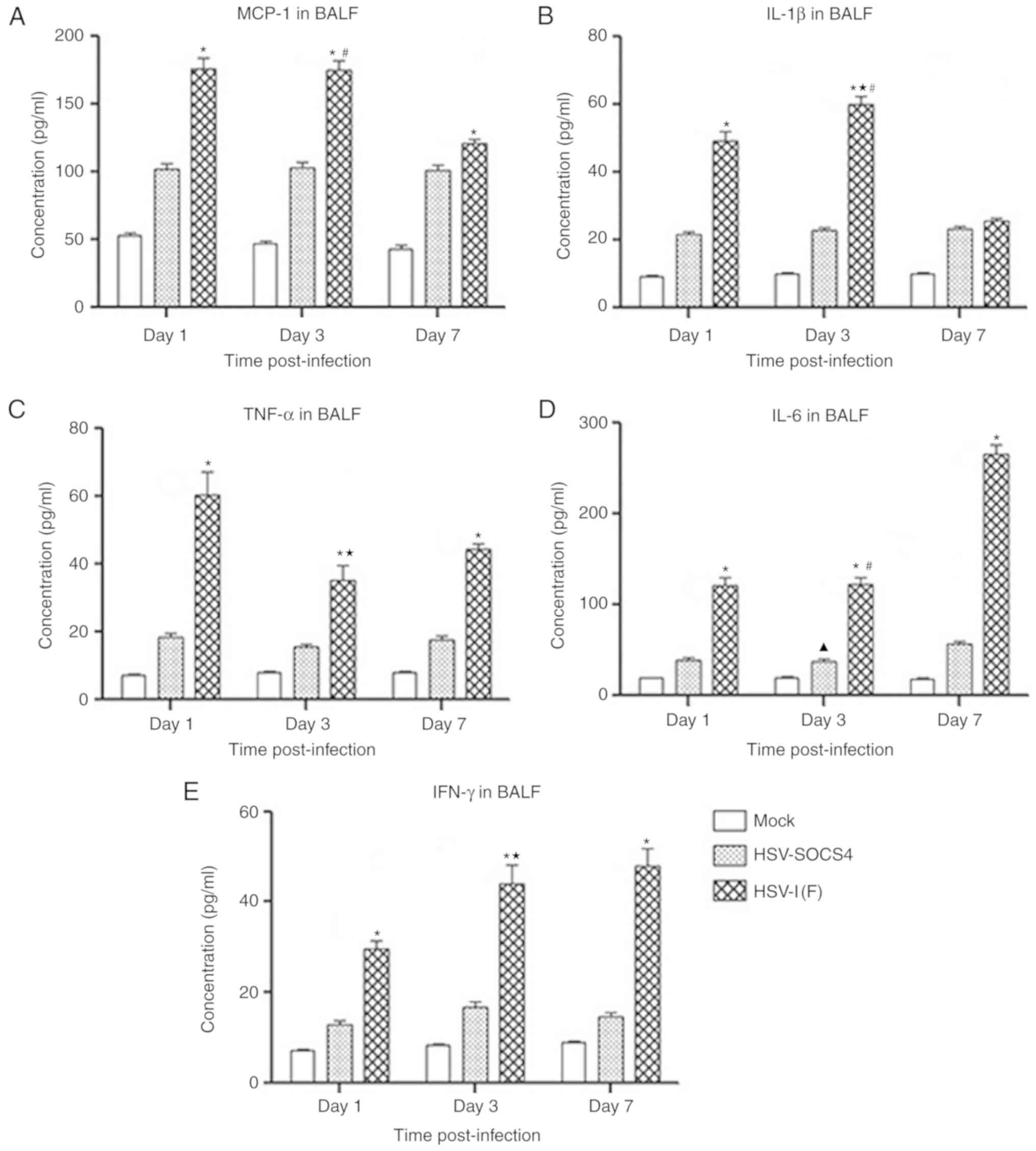
investigated. The outcomes of cytokine production in BALF samples
are presented in Fig. 2. A
significantly increased level of all five cytokines was observed in
HSV-1(F)-infected mice compared with that in HSV-SOCS4-infected
mice on days 1, 3 and 7, except for IL-1β production on day 7 for
which a negligible difference between the two groups of mice was
observed. IL-1β production increased on day 3 compared with day 1
in HSV-1(F)-infected mice, but decreased by ~50% on day 7 (Fig. 2B). IL-6 and IFN-γ production in
HSV-1(F)-infected mice was increased, but the greatest increase in
IL-6 was observed on day 7, whereas that of IFN-γ was observed on
day 3. The maximum level of MCP-1 in HSV-1(F)-infected mice was
detected on day 1 and day 3, but levels had decreased by day 7. The
highest TNF-α level was also observed on day 1, but it had
decreased when determined on day 3. The cytokine secretion of BALF
from HSV-SOCS4-infected mice was consistent at the time points
investigated with a slight difference, except for a significant
increase in IL-6 observed on day 7.
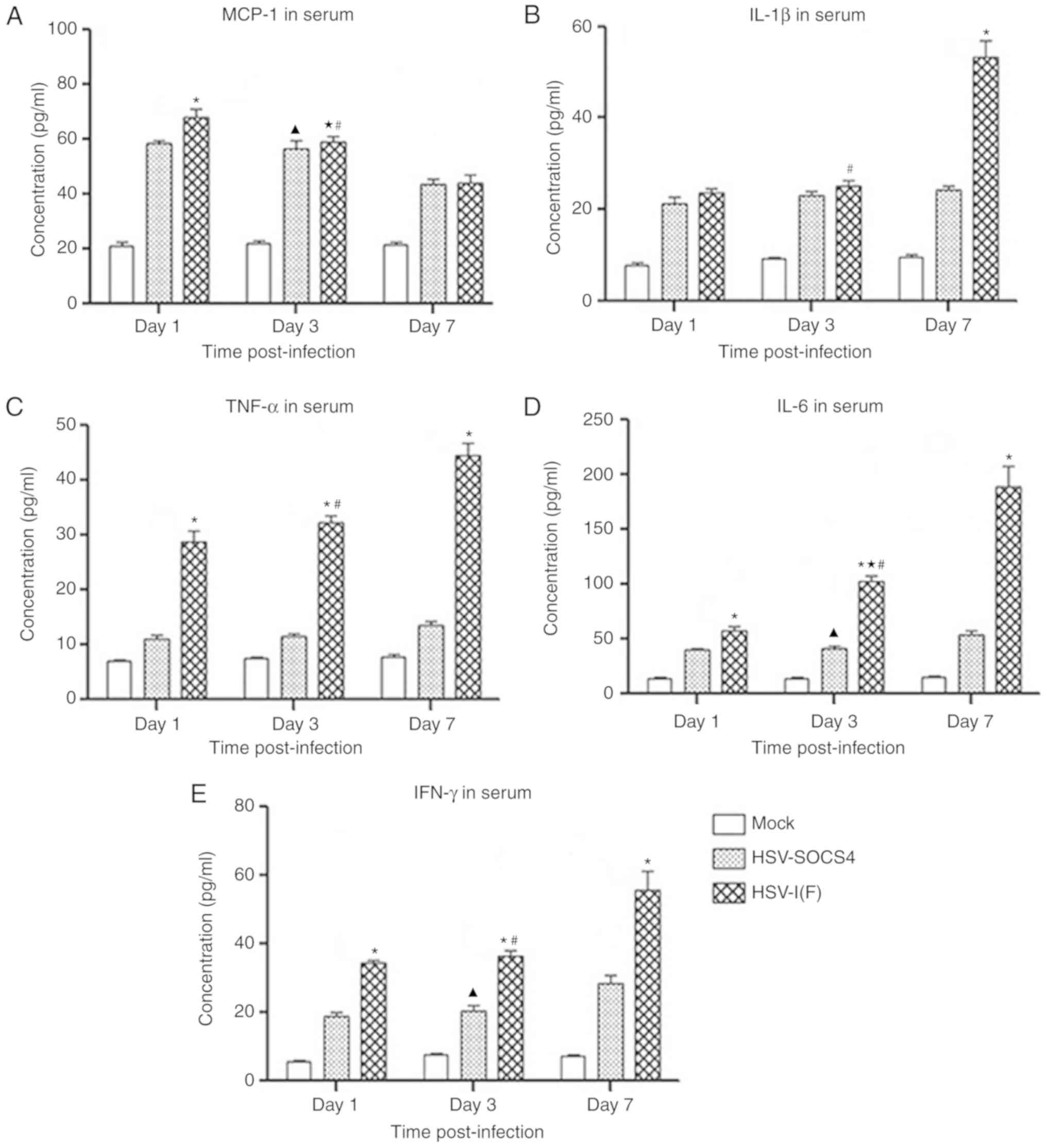
To determine the effect of SOCS4 protein on cytokine
production in systemic circulation, mouse sera were collected for
analysis by ELISA, and results are presented in Fig. 3. A significantly increased
concentration of MCP-1 was detected in HSV-1(F)-infected mice
compared with that in HSV-SOCS4-infected mice on day 1, and MCP-1
production in HSV-1(F)-infected mice decreased significantly at
each time point, but was significantly decreased only on day 7 in
HSV-SOCS4-infected mice. Additionally, the MCP-1 level in BALF was
markedly higher compared with that in serum at every day measured
in the two groups. IL-1β levels in HSV-1(F)- and HSV-SOCS4-infected
mice were similar on day 1 and 3, but a significant increase was
observed on day 7 in HSV-1(F)-infected mice. For TNF-α production,
levels in HSV-SOCS4 were consistent over time, but TNF-α levels in
HSV-1(F)-infected mice were increased compared with those in
HSV-SOCS4-infected mice on days 1 and 3. The level of IL-6 was
significantly increased in HSV-1(F)-infected mice over time, but a
significantly increased level in HSV-SOCS4-infected mice was
observed only on day 7. Additionally, the IL-6 level in
HSV-1(F)-infected mice was significantly increased compared with in
HSV-SOCS4-infected mice on every day analyzed. For IFN-γ, the
increases on day 1 and 3 were slight, but were significantly by day
7 in HSV-1(F)-infected mice and HSV-SOCS4-infected mice. The
concentration of IFN-γ in serum was increased compared with that in
BALF on day 7, confirming that activated T cells became the primary
source of IFN-γ production. To further determine cytokine
production in serum of HSV-SOCS4-infected mice, cytokine levels
were determined on day 12, and no notable differences were observed
(data not shown). In summary, the cytokine levels in BALF of
HSV-SOCS4-infected mice were more consistent compared with those in
serum.
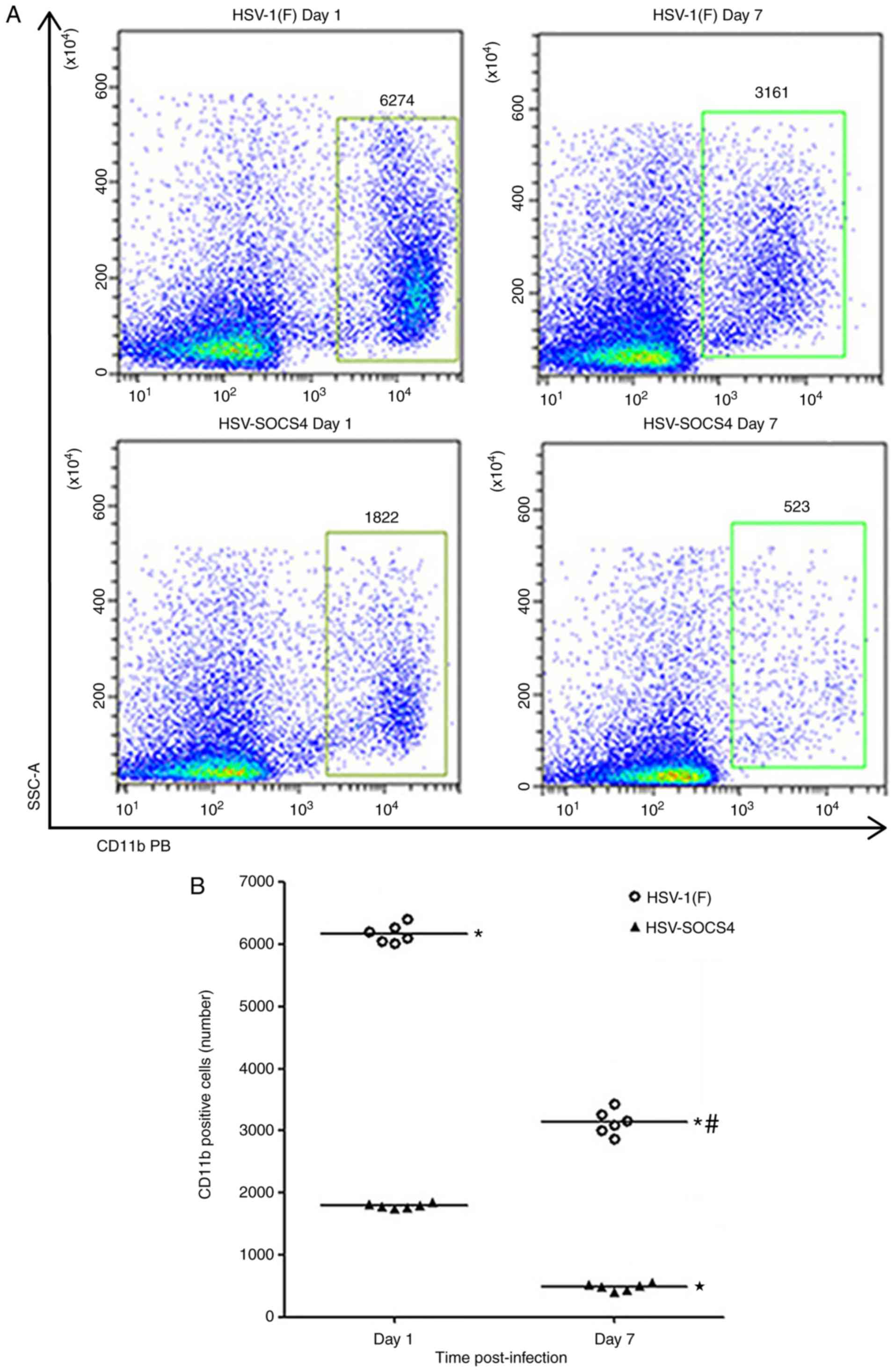
Cell analysis of BALF and spleen
following virus infection
To determine whether the diverse cytokine levels in
BALF and serum were associated with the quantity of immune cells,
cells from BALF and spleen of infected mice were collected for flow
cytometric analysis. Since the difference in cytokine production
was evident on days 1 and 7, cells were collected on these days to
make the comparison. Considering that CD11b+ cells,
including macrophages, neutrophils and natural killer (NK) cells,
constitute the main cell population present in BALF, the variation
in CD11b+ cells was quantified between HSV-1(F)- and
HSV-SOCS4-infected mice. It was observed that CD11b+
cells from HSV-1(F)-infected mice were more prevalent compared with
those from HSV-SOCS4-infected mice, and there was a significant
decrease in positively stained cells on day 7 compared with on day
1 in the two groups of infected mice (Fig. 4).
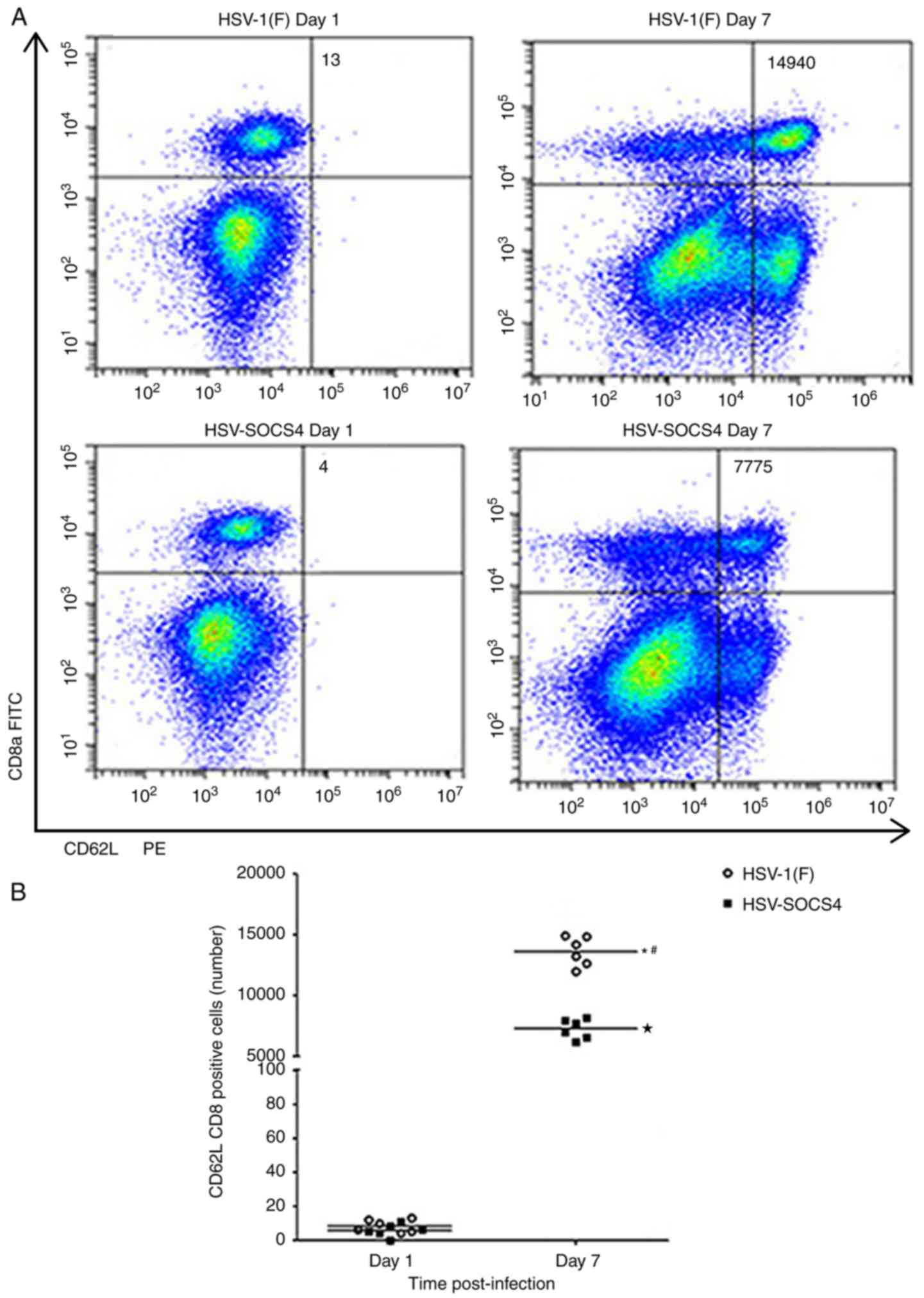
CD4+ and CD8+ cells from
spleen were stained and CD62L was used as an activated marker. As
presented in Fig. 5A, few
double-positive cells were evident on day 1 in either group of
infected mice, but an increased number of
CD8+CD62L+ T cells was observed on day 7.
These differences were determined to be significant (Fig. 5B). A significant difference in the
number between HSV-1(F) and HSV-SOCS4-infected mice was also
observed (Fig. 5B). A similar
pattern was also observed for CD4+CD62L+ T
cells (Fig. 5C and D).
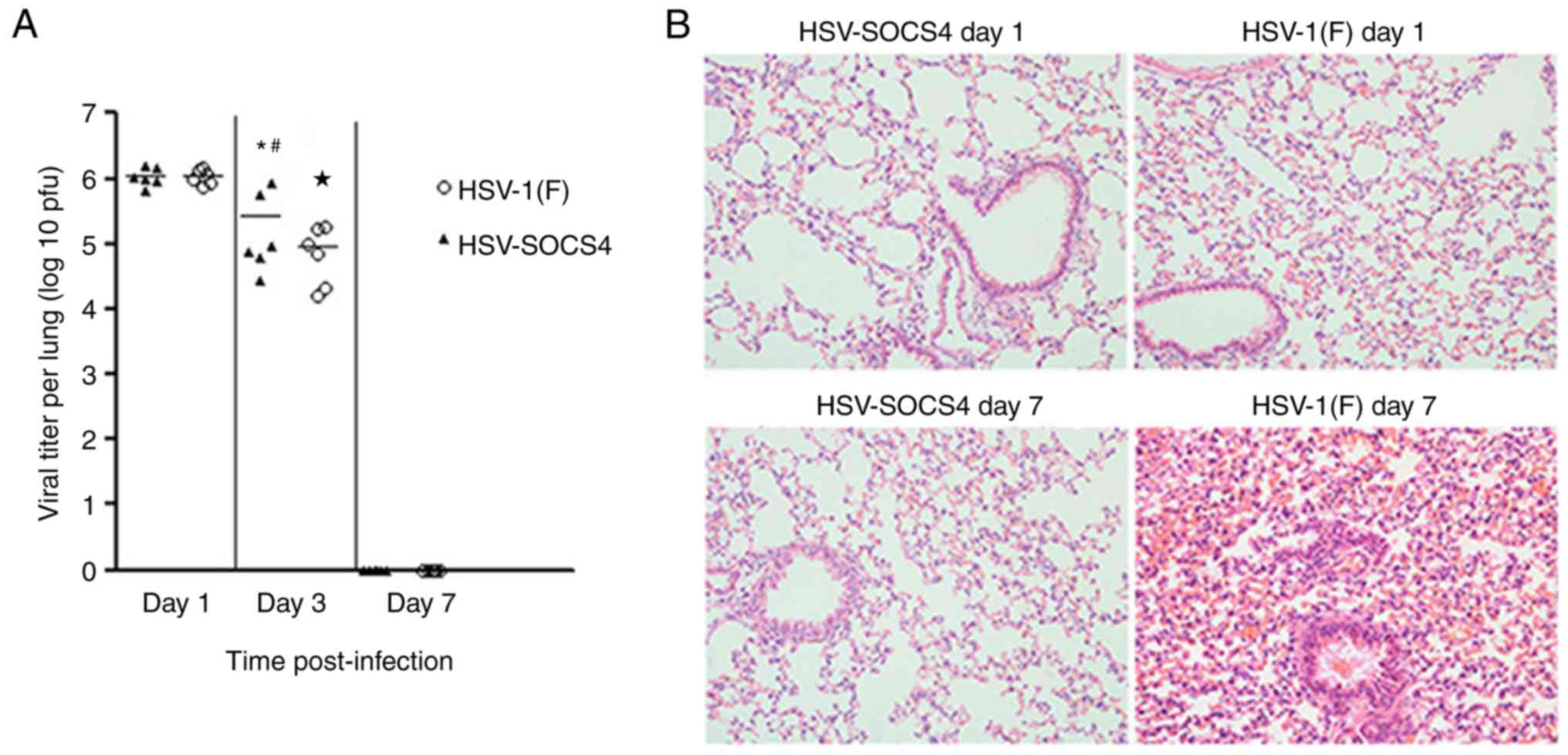
Virus titers and pathological changes
in the lung following virus infection
To determine the association of cytokine production
with virus replication/clearance, the virus titer from infected
mouse lung was quantified. The maximum virus titer was observed on
day 1, and evidently decreased by day 3, and no virus was detected
on day 7. Furthermore, the virus titer was significantly different
between HSV-1(F)-infected mice and HSV-SOCS4-infected mice on day 3
(Fig. 6A).
The lungs of mice with no BALF collection underwent
histopathological analysis, and representative images (at ×200
magnification) are presented in Fig.
6B. On day 1, the lung of HSV-SOCS4-infected mouse barely
exhibited any pathological changes, but evident cell infiltration
with slight dilatation and hyperemia of local capillary was
exhibited in the HSV-1(F)-infected mouse lung. On day 7,
infiltration of a number of immune cells and mild-to-moderate
dilatation and hyperemia of the capillary were observed in
HSV-SOCS4-infected mouse lung, but the architecture of the lung
alveolar wall was undisrupted. Additionally, a severe pathological
change appeared in HSV-1(F)-infected mouse lung: A thickened and
disrupted alveolar wall with severe surrounding hyperemia,
accompanied by congested immune cells.
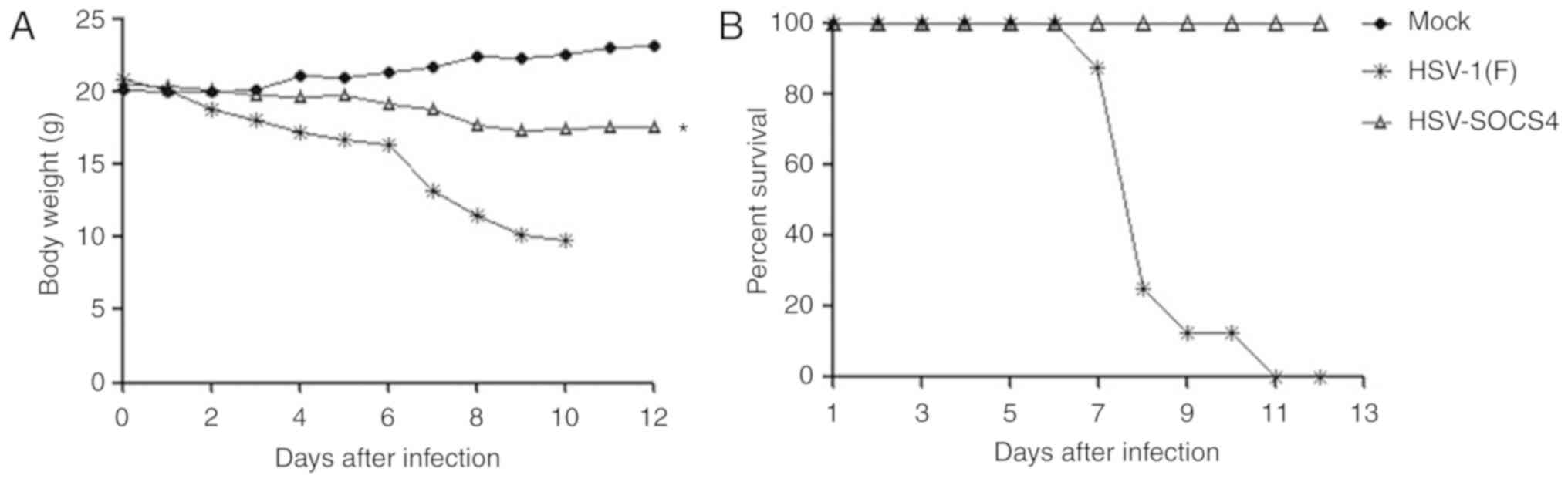
Body weight and mortality of mice
following virus infection
Following infection, all mice were monitored twice
daily for a period of 12 days to determine the body weight and
onset of mortality. HSV-1(F)-infected mice started to lose their
body weight gradually on day 2, with a marked loss on day 7, and
the mice remaining alive had lost 50% of the body weight on day 10
(Fig. 7A). Consistently, the
survival rate started to decline on day 7 (Fig. 7B), and then, the mice succumbed
rapidly; no mouse from the HSV-1(F) group survived by day 11. The
mice in the HSV-SOCS4 group lost weight slightly and generally kept
80% of their weight on day 12. The survival rate of
HSV-SOCS4-infected mice was maintained at 100%, which was
significantly different from that of the HSV-1(F) group.
Mock-infected mice exhibited no weight loss and no mortality.
Discussion
HSV-1 infects various mucosal tissues, including the
respiratory tract, and it was identified previously that
HSV-1-inducted pneumonia was due to the inflammatory response
rather than direct cytopathic effects of the virus itself (15), indicating the potential risk of
using oHSV as antitumor therapy, particularly because patients with
cancer are generally in disordered immuno-homeostasis. This
uncontrolled inflammatory response is the consequence of an
excessive release of pro-inflammatory cytokines, known as cytokine
storm. Unfortunately, the understanding of molecules involved in
cytokine storm, contribution of cytokines to pathogenesis, and
therapeutic strategies to prevent or alleviate symptoms remain
insufficient. We hypothesized that increased negative feedback to
the immune system or inhibiting the overproduction of inflammatory
cytokines may be a prospective method to prevent tissue and organ
damage caused by cytokine storm.
Unlike SOCS1, SOCS3 and other members of the SOCS
family that have been well-described (16–18),
studies concerning SOCS4 protein are relatively scarce. SOCS4 is
known to be responsible for the inhibition of immune signaling by
STAT3, which is associated with human cancer therapy (19,20).
Mice lacking functional SOCS4 were hypersusceptible to primary
infection with influenza A virus, and displayed dysregulated
pro-inflammatory cytokine production in the lungs, delayed viral
clearance and impaired trafficking of influenza-specific
CD8+ T cells to the site of infection (13). Therefore, SOCS4 may be a promising
protein to control the excessive release of pro-inflammatory
cytokines induced by oHSV treatment. To validate our hypothesis, a
novel HSV-1 strain with an inserted SOCS4 gene named
HSV-SOCS4 was constructed, and it was used to infect mice
intranasally to evaluate whether cytokine storm could be
inhibited.
Since cytokines instilled into the lungs could pass
into the bloodstream, providing direct communication between local
and systemic response, BALF and serum samples were collected for
the determination of several cytokines that are involved in
cytokine storm on days 1, 3 and 7 post-infection. Produced
primarily by dendritic cells (DCs) and macrophages, IL-1β is a key
cytokine driving pro-inflammatory activity. It promotes recovery
when present early in infection, but is associated with a damaging
inflammatory response, leading to severe pathogenesis and mortality
when present at late stages of infection (21). It was identified that IL-1β remained
at a high level at the early stages (days 1 and 3) in BALF samples
and was degraded on day 7. By contrast, its production in serum was
increased on day 7 in HSV-1(F)-infected mice. BALF and serum IL-1β
maintained corresponding low levels during all time points in
HSV-SOCS4-infected mice. These data suggested that SOCS4 protein
may inhibit early IL-1β production in BALF and later production in
serum. IL-1 and IL-6 are synergistic, but IL-1 is expressed
principally in the early stages of infection, followed by an
increase in the expression of IL-6 (22). The production of IL-6 in BALF
following HSV infection has been reported (23), and the results of the present study
were consistent: Strong upregulation of IL-6 levels in BALF of
HSV-1(F)-infected mice was evident, followed by a sustained
increase, particularly on day 7; and similar results were also
evident in serum with a relatively lower concentration. Restricted
levels of IL-6 were apparent in BALF and serum in
HSV-SOCS4-infected mice and exhibited an increase on day 7. The
marked later production of IL-6 was independent of the presence of
virus (24) and may be associated
with promotion of Th2 responses (25). TNF-α, another prominent
acute-response cytokine, is primarily produced by macrophages, lung
epithelial cells and helper T cells, and may appear at early times
following infection. TNF-α contributes to the symptoms of severe
disease following H5N1 virus infection and represents the
quintessential features of cytokine storm (26,27);
it is also involved in immunopathology associated with HSV
infection (28). TNF-α was released
from the innate immune system through virus interaction with
macrophages and NK cells at early times following infection (a
notably high level of TNF-α was observed in BALF on day 1 in the
present study), and the adaptive immune system via activation of
virus-specific CD4+ or CD8+ T cells [an
increased level of TNF-α was observed in serum on day 7 from
HSV-1(F)-infected mice], but variations in the TNF-α level in
HSV-SOCS4-infected mice were indistinguishable at every time point
in BALF and serum samples. It was reported that anti-TNF-α
treatment decreased the severity of weight loss and illness
following H3N2 virus challenge, indicating that it may be a
promising therapeutic target (29),
and we hypothesized that restrained TNF-α production may also
decrease the symptoms caused by HSV infection. MCP-1 is rapidly
produced by various cell types, primarily monocytes, macrophages,
epithelial cells and endothelial cells, following inflammatory
stimuli and tissue damage. It recruits monocytes, memory T cells,
NK cells and DCs to the sites of tissue injury and infection
(30,31). A markedly higher level of MCP-1 in
BALF was observed at early stages from HSV-1(F)-infected mice,
consistent with the observation that more CD11b+ cells
were present in HSV-1(F)-infected mouse BALF samples. IFN-γ is a
potent cytokine with numerous functions, including promoting the
activation of DCs and macrophages, enhancing the cytotoxicity of NK
cells and inducing antibody production of B cells (32). The increased IFN-γ level of
HSV-1(F)-infected mice on day 3 in BALF was produced primarily by
NK cells at the early stage of infection and it helped to control
viral replication (33); the
increased level in serum from the two groups of infected mice at
the later stage (day 7) was because T cells became the principal
source of IFN-γ. Certain activities of IFN-γ in the later response
had been associated with inflammation and lung injury (34), possibly explaining the lung damage
in HSV-1(F)-infected mice. Prolonged and indiscriminate IFN-γ
production was observed in the serum samples of HSV-SOCS4-infected
mice on day 12 when the virus could not be identified in lung
tissue, confirming that IFN-γ could be induced downstream by other
cytokines or features of the immune response (35), and SOCS4 protein may inhibit the
prolonged production in serum.
Pro-inflammatory cytokines are responsible for cell
activation and tissue damage; additionally, the release of one
cytokine may induce new cytokine production, which will in turn
further cause cell and organ necrosis. The consistently low level
of pro-inflammatory cytokine production in BALF and serum of
HSV-SOCS4-infected mice is thought to be associated with the
amelioration of mice lung damage compared with that of
HSV-1(F)-infected mice.
A marked number of CD11b+ cells was
present in the HSV-1(F)-infected mouse BALF on day 1, which was the
consequence of the high level of MCP-1 in BALF. It was reported
that macrophages serve an essential function in the first line of
defense against HSV within the lung by rapidly secreting a primary
wave of pro-inflammatory cytokines (36), explaining the increased production
of TNF-α, IL-1β and IL-6 in BALF of HSV-1(F)-infected mice on day
1, because macrophages and NK cells are the primary source of those
cytokines in the initial response. Recruitment of macrophages into
the lung and alveolar spaces is a hallmark of the initial immune
response, and CD4+ and CD8+ T cells in the
spleen may reflect the activity of the adaptive immune response.
CD62L is generally used as an activation marker of T cells, and it
serves a major function in directing lymphocytes to the site of
infection and inflammation (37).
No activated T cells were identified in the spleen on day 1 from
either group of infected mice, but marked activation of cells was
evident on day 7, and the CD4+CD62L+ T cell
number and the CD8+CD62L+ T cell number in
HSV-1(F)-infected mice were increased 2-fold compared with those of
HSV-SOCS4-infected mice, a result that was associated with the
increased level of TNF-α, IL-1β, IL-6 and IFN-γ in serum of
HSV-1(F)-infected mice on day 7. Effective Th cells and CTLs are
critical for the efficient resolution of viral infection through
the production of cytokines and/or direct lysis of infected cells;
however, these same mechanisms also contribute to pulmonary damage
(38).
The virus titer in the lung was similar between
HSV-1(F)- and HSV-SOCS4-infected mice on day 1, indicating that the
two groups of mice were equally infected. However, the levels
declined on day 3, and an evident difference was detected between
the two groups. This rapid virus clearance was consistent with
innate immunity mechanisms, and, due to the activation of a larger
quantity of immune cells (as observed by flow cytometric analysis
and pathological analysis on day 1), a decreased virus load was
detected in HSV-1(F)-infected mouse lung. Typically, lytic
infection is shut off on day 7; therefore, no virus was tested.
Following HSV-1(F) infection, serious pathological changes in the
mouse lung were observed on day 7, consistent with the increased
cytokine levels and T cells of the mice. Owing to lung damage
(possibly involving other organ lesions), excessive weight loss of
HSV-1(F)-infected mice started on day 7, and the mortality rate
reached 75% on day 8 and 100% on day 11. Conversely,
HSV-SOCS4-infected mice exhibited only slight weight loss and no
mortality on day 12. Those results indicated that controlling
cytokine over-release could maintain the body weight, health and
survival of infected mice. The results of the present study
indicated that HSV with SOCS4 protein insertion inhibited cytokine
overproduction, excessive infiltration of immune cells, alleviated
lung pathological damage and decreased the mortality rate of
mice.
Immunity to virus infection is multifaceted and
highly complex, and cytokine induction is a series of sprawling
network with redundancy and amplified cascades. Thus, intervention
strategies should target multiple cytokine pathways. The attempt in
the present study to create a recombinant HSV-1 strain with SOCS4
expression was to provide a valuable tool to suppress the cytokine
storm and its disastrous consequence, which may improve oHSV
clinical treatment. The molecular mechanisms of SOCS4 in the
cytokine pathway and effects of the HSV-SOCS4 strain in mice with
tumor and other animals requires further investigation.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Nature
Science Foundation of China (grant no. NSFC 81472826), the Shenzhen
Science and Innovation Commission Project (grant no.
CYZZ20150831144745459), and the National Major Scientific and
Technological Special Project for Significant New Drugs Development
during the Thirteenth Five-year Plan Period (grant no.
2018ZX09733002).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SR performed all sample collection, double sandwich
ELISA, and flow cytometric analysis. XC recombined the HSV-SOCS4
and cell culture. RH performed the mice infection and pathological
analysis of mouse lung. GGZ contributed to the study design and
wrote the manuscript. ZY designed the study, analyzed the data and
drafted the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study does not contain any studies with
human participants performed by any of the authors. All procedures
involving mice were approved by the Institutional Animal Care and
Use Committee of Guangzhou Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Russell SJ, Peng KW and Bell JC: Oncolytic
virotherapy. Nat Biotechnol. 30:658–670. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jhawar SR, Thandoni A, Bommareddy PK,
Hassan S, Kohlhapp FJ, Goyal S, Schenkel JM, Silk AW and Zloza A:
Oncolytic viruses-natural and genetically engineered cancer
immunotherapies. Front Oncol. 7:2022017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maroun J, Muñoz-Alía M, Ammayappan A,
Schulze A, Peng KW and Russell S: Designing and building oncolytic
viruses. Future Virol. 12:193–213. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lin CZ, Xiang GL, Zhu XH, Xiu LL, Sun JX
and Zhang XY: Advances in the mechanisms of action of
cancer-targeting oncolytic viruses. Oncol Lett. 15:4053–4060.
2018.PubMed/NCBI
|
|
5
|
Kaufman HL, Kohlhapp FJ and Zloza A:
Oncolytic viruses: A new class of immunotherapy drugs. Nat Rev Drug
Discov. 14:642–662. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Loskog A: Immunostimulatory gene therapy
using oncolytic viruses as vehicles. Viruses. 7:5780–5791. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peters C and Rabkin SD: Designing herpes
viruses as oncolytics. Mol Ther Oncolytics. 2(pii): 150102015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saha D, Wakimoto H and Rabkin SD:
Oncolytic herpes simplex virus interactions with the host immune
system. Curr Opin Virol. 21:26–34. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chaurasiya S, Chen NG and Fong Y:
Oncolytic viruses and immunity. Curr Opin Immunol. 51:83–90. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tisoncik JR, Korth MJ, Simmons CP, Farrar
J, Martin TR and Katze MG: Into the eye of the cytokine storm.
Microbiol Mol Biol Rev. 76:16–32. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chousterman BG, Swirski FK and Weber GF:
Cytokine storm and sepsis disease pathogenesis. Semin Immunopathol.
39:517–528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alexander WS: Suppressors of cytokine
signalling (SOCS) in the immune system. Nat Rev Immunol. 2:410–416.
2002. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kedzierski L, Linossi EM, Kolesnik TB, Day
EB, Bird NL, Kile BT, Belz GT, Metcalf D, Nicola NA, Kedzierska K,
et al: Suppressor of cytokine signaling 4 (SOCS4) protects against
severe cytokine storm and enhances viral clearance during influenza
infection. PLoS Pathog. 10:e10041342014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Y, Qu L, Liu Y, Roizman B and Zhou GG:
PUM1 is a biphasic negative regulator of innate immunity genes by
suppressing LGP2. Proc Natl Acad Sci USA. 114:E6902–E6911. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Adler H, Beland JL, Del-Pan NC, Kobzik L,
Brewer JP, Martin TR and Rimm IJ: Suppression of herpes simplex
virus type 1 (HSV-1)-induced pneumonia in mice by inhibition of
inducible nitric oxide synthase (iNOS, NOS2). J Exp Med.
185:1533–1540. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liau NPD, Laktyushin A, Lucet IS, Murphy
JM, Yao S, Whitlock E, Callaghan K, Nicola NA, Kershaw NJ and Babon
JJ: The molecular basis of JAK/STAT inhibition by SOCS1. Nat
Commun. 9:15582018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mahony R, Ahmed S, Diskin C and Stevenson
NJ: SOCS3 revisited: A broad regulator of disease, now ready for
therapeutic use? Cell Mol Life Sci. 73:3323–3336. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Linossi EM, Babon JJ, Hilton DJ and
Nicholson SE: Suppression of cytokine signaling: The SOCS
perspective. Cytokine Growth Factor Rev. 24:241–248. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Trengove MC and Ward AC: SOCS proteins in
development and disease. Am J Clin Exp Immunol. 2:1–29.
2013.PubMed/NCBI
|
|
20
|
Wang X, Crowe PJ, Goldstein D and Yang JL:
STAT3 inhibition, a novel approach to enhancing targeted therapy in
human cancers (Review). Int J Oncol. 41:1181–1191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tate MD, Ong JD, Dowling JK, McAuley JL,
Robertson AB, Latz E, Drummond GR, Cooper MA, Hertzog PJ and
Mansell A: Reassessing the role of the NLRP3 inflammasome during
pathogenic influenza A virus infection via temporal inhibition. Sci
Rep. 6:279122016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kaiser L, Fritz RS, Straus SE, Gubareva L
and Hayden FG: Symptom pathogenesis during acute influenza:
Interleukin-6 and other cytokine responses. J Med Virol.
64:262–268. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Murphy EA, Davis JM, Brown AS, Carmichael
MD, Ghaffar A and Mayer EP: Effect of IL-6 deficiency on
susceptibility to HSV-1 respiratory infection and intrinsic
macrophage antiviral resistance. J Interferon Cytokine Res.
28:589–595. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tengku-Muhammad TS, Hughes TR, Ranki H,
Cryer A and Ramji DP: Differential regulation of macrophage
CCAAT-enhancer binding protein isoforms by lipopolysaccharide and
cytokines. Cytokine. 12:1430–1436. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hunter CA and Jones SA: IL-6 as a keystone
cytokine in health and disease. Nat Immunol. 16:448–457. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peiris JS, Cheung CY, Leung CY and
Nicholls JM: Innate immune responses to influenza A H5N1: Friend or
foe? Trends Immunol. 30:574–584. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Perrone LA, Szretter KJ, Katz JM, Mizgerd
JP and Tumpey TM: Mice lacking both TNF and IL-1 receptors exhibit
reduced lung inflammation and delay in onset of death following
infection with a highly virulent H5N1 virus. J Infect Dis.
202:1161–1170. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fujii S, Akaike T and Maeda H: Role of
nitric oxide in pathogenesis of herpes simplex virus encephalitis
in rats. Virology. 256:203–212. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hussell T, Pennycook A and Openshaw PJ:
Inhibition of tumor necrosis factor reduces the severity of
virus-specific lung immunopathology. Eur J Immunol. 31:2566–2573.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Deshmane SL, Kremlev S, Amini S and Sawaya
BE: Monocyte chemoattractant protein-1 (MCP-1): An overview. J
Interferon Cytokine Res. 29:313–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rose CJ, Sung SS and Fu SM: Significant
involvement of CCL2 (MCP-1) in inflammatory disorders of the lung.
Microcirculation. 10:273–288. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Billiau A and Matthys P: Interferon-gamma:
A historical perspective. Cytokine Growth Factor Rev. 20:97–113.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reading PC, Whitney PG, Barr DP, Smyth MJ
and Brooks AG: NK cells contribute to the early clearance of HSV-1
from the lung but cannot control replication in the central nervous
system following intranasal infection. Eur J Immunol. 36:897–905.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ramana CV, DeBerge MP, Kumar A, Alia CS,
Durbin JE and Enelow RI: Inflammatory impact of IFN-γ in
CD8+ T cell-mediated lung injury is mediated by both
Stat1-dependent and -independent pathways. Am J Physiol Lung Cell
Mol Physiol. 308:L650–L657. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guo XJ and Thomas PG: New fronts emerge in
the influenza cytokine storm. Semin Immunopathol. 39:541–550. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Holt PG, Strickland DH, Wikstrom ME and
Jahnsen FL: Regulation of immunological homeostasis in the
respiratory tract. Nat Rev Immunol. 8:142–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rainer TH: L-selectin in health and
disease. Resuscitation. 52:127–141. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu Q, Zhou YH and Yang ZQ: The cytokine
storm of severe influenza and development of immunomodulatory
therapy. Cell Mol Immunol. 13:3–10. 2016. View Article : Google Scholar : PubMed/NCBI
|