Introduction
Gastric cancer (GC) is one of the most lethal
diseases in the world (1). Hypoxia
is a general problem in solid tumors, including GC. Hypoxic cells
undergo pro-survival alterations, including improved proliferation
and invasion ability and enhanced resistance to chemotherapy
(2,3). Therefore, actions to clarify the
mechanisms and restrain the malignant behavior of hypoxic GC are
not only vital but also urgent.
Stanniocalcin-1 (STC1), a glycoprotein involved in
calcium/phosphate homeostasis, was demonstrated to be elevated in
several types of carcinoma tissue compared with in corresponding
normal tissue (4–7). Notably, Fang et al (7) reported that overexpression of STC1 in
tissue from GC was associated with a poor prognosis, but the
mechanisms are unclear. Notably, the expression of STC1 could be
induced by hypoxia (8,9). Therefore, for better control of GC, it
is necessary to elucidate the roles of STC1 in hypoxic GC and
identify proper therapeutic targets.
B cell lymphoma-2 (Bcl-2), one of the key proteins
upstream of the apoptotic cascade (10), also serves roles in hypoxia
(11). Given that overexpression of
STC1 elevated the levels of Bcl-2 under normal conditions (4), it is possible that STC1 works together
with Bcl-2 in a hypoxic environment.
In the present study, the roles of STC1 in GC were
investigated. It was demonstrated that under hypoxic conditions,
STC1 probably promoted cell proliferation, chemoresistance and
metastasis in GC via positively regulating Bcl-2, suggesting that
STC1 and Bcl-2 may be potential targets in the treatment of GC.
Materials and methods
Cell lines and cell culture
GC cell lines HGC27 (#TCHu22) and NCIN87 (#SCSP-534)
were obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China); MKN28
(#3111C0001CCC000482) and MKN45 (#3111C0001CCC000229) cells were
obtained from The National Infrastructure of Cell line Resource
(Beijing, China); SNU601 cells were obtained from the Korean Cell
Line Bank (Seoul, Korea); SNU216 cells were provided by the Medical
College of Xiamen University (Xiamen, China). Retroviral packaging
cells (Phoenix amphotropic cells) were gifts from Professor Gong
Yang at Fudan University Shanghai Cancer Center (Shanghai, China).
Cells were cultured in Minimum Essential Medium (cat. no.
11095-080; HGC27) or RPMI1640 medium (cat. no. 12633-012)
containing 10% fetal bovine serum (cat. no. 10099-141; all from
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100
units/ml penicillin and 100 µg/ml streptomycin. To simulate
ahypoxic environment in vitro, cells were incubated with 100
µM cobalt chloride (CoCl2; Sigma Aldrich; Merck KGaA,
Darmstadt, Germany) for at least 48 h according to different
procedures. All cells were cultured or treated in a humid
atmosphere at 37°C with 5% CO2 and 95% air.
It has been reported that MKN28, a kind of gastric
tubular adenocarcinoma, was contaminated with MKN74, another
gastric tubular adenocarcinoma cell line (12). However, throughout the study MKN28
was used, and the results are repeatable.
Cell transfection and viral
infection
The retroviral plasmids carrying STC1 cDNA and the
empty vector (Vec), small hairpin RNA (shRNA) against the open
reading frame of STC1 mRNA and the scrambled shRNA (Scr) were
provided by Professor Gong Yang. The cells were infected as
described previously (4).
SNU601-STC1 cells were transfected with either 50 nM
Bcl-2-targeting small interfering (si)RNA (Bcl2i; targeting
sequence, CAGGACCTCGCCGCTGCAGAC) (13) or scrambled negative controls (NCi;
target sequence, AATTCTCCGAACGTGTCACGT; Shanghai GenePharma Co.,
Ltd., Shanghai, China) (13) using
the Lipofectamine 2000 transfection reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Cells were then incubated for 48 h prior to subsequent
experiments.
Immunoblotting analysis
To analyze protein levels in hypoxic or normoxic
cells, cells were treated with or without CoCl2 for 48
h, and immunoblotting analysis was performed as previously
described (14). The following
antibodies were used for immunoblotting: STC1 (cat. no. 20621-1-AP;
1:1,000), hypoxia inducible factor (HIF)-1α (cat. no. 66730-1-Ig;
1:5,000), epithelial (E)-cadherin (cat. no. 20874-1-AP; 1:1,000;
all ProteinTech Group, Inc., Chicago, IL, USA), Bcl-2 (cat. no.
sc-7382; 1:500), caspase-9 (cat. no. sc-56076; 1:500), caspase-3
(cat. no. sc-7148; 1:500), neural (N)-cadherin (cat. no. sc-1502;
1:1,000; all Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
matrix metalloproteinase (MMP)-1 (cat. no. M4427; 1:1,000;
Sigma-Aldrich; Merck KGaA), X-linked inhibitor of apoptosis protein
(XIAP; cat. no. cs-2045; 1:1,000) and β-actin (cat. no. cs-4970;
1:1,000; both Cell Signaling Technology, Inc., Danvers, MA, USA).
β-Actin was used as loading control. The secondary antibodies were
horse anti-mouse immunoglobulin (Ig)G or goat anti-rabbit IgG
conjugated to horseradish peroxidase (HRP) (cat. nos. 7076 and
7074, respectively; 1:2,000; Cell Signaling Technology, Inc.).
Immobilon Western Chemiluminescent HRP Substrate (cat. no.
WBKLS0500) was purchased from Merck KGaA. The integrated density of
each band was normalized to the density of corresponding
β-actin.
Growth curve assay
Growth curves were measured by MTT assay as
previously described (14). To
compare cell proliferation ability underhypoxia or normoxia, cells
were cultured with or without CoCl2. The medium were
refreshed at the 4th day. Data were collected at 1, 3, 5, and 7
days by measuring the absorbance at 490 nm.
Cisplatin treatment
Cells were treated with cisplatin (Hansoh Pharma,
Lianyungang, Jiangsu, China) and CoCl2 for 48 h at 37°C.
The half-maximal inhibitory concentration (IC50) was
measured by MTT assay according to procedures described previously
(15). The concentration of
cisplatin used included 1, 10, 100 and 1,000 µM for the primary
screening for approximate ranges of IC50, and 20, 30,
40, 50, 60, 70, 80, 90 and 100 µM for the final evaluation of
IC50. For apoptosis analysis, cells were treated with
cisplatin at the approximate concentration of IC50 (100
µM for SNU601 and 50 µM for MKN28) and CoCl2 for 48 h
prior to flow cytometry. Apoptosis was assessed by Annexin
V-PE-7AAD (cat. no. 559763; BD Pharmingen; BD Biosciences, San
Jose, CA, USA) following the protocol described previously
(15).
Cell migration and invasion assay
To evaluate cell migration, wound-healing assays
were performed as previously described (16). The widths of the wounds were
measured at 72 h following scratching. The 24-well Transwell
chamber with 8-µm pore and Matrigel (Corning, Inc., Corning, NY,
USA) were used for evaluating cell invasion as previously described
(16). Cells were incubated with
CoCl2 in the two assays.
Xenograft tumors in nude mice
A total of 20 mice (BALB/c athymic nude mice; age,
4–6 weeks; weight, 15–20 g; sex ratio, 1:1) were purchased from,
and also treated and maintained in Shanghai Experimental Animal
Center, Chinese Academy of Sciences (Shanghai, China). The mice
were maintained in the animal facility in individually ventilated
cages in specific pathogen-free conditions in a temperature of
18–29°C and humidity of 40–70%, with a 12-h light/dark cycle, and
received food and water ad libitum. The animal experiments
were approved by the Institutional Animal Care and Use Committee of
Chinese Academy of Sciences and performed following institutional
guidelines and protocols. To observe the effects of STC1 on tumor
growth in vivo, SNU601-STC1 and SNU601-Vec cells were used
to generate xenograft tumors on mice. A total of 8×106
cells were subcutaneously injected into each mouse (4 mice for each
cell line). The longest diameter ‘a’ and the shortest diameter ‘b’
of tumors were measured and the tumor volume was calculated with
the use of the following formula: Tumor volume (in mm3)
= axb2x0.52 (14). To
observe the effects of STC1 on peritoneal metastasis in
vivo, 1×107 cells were injected into the peritoneal
cavity of every mouse (six mice for each cell line). When the
volume of subcutaneous tumor reached 1,500 mm3, mice
were sacrificed. All the subcutaneous tumor and peritoneal
cancerous nodes were excised and weighed.
Online data analysis
Data from the large online public database
Kaplan-Meier Plotter (n=876; http://kmplot.com/analysis/) was analyzed on the
website with the three different probes (204595_s_at, 204596_s_at
and 204597_x_at) using the auto select best cut-off (17).
Immunohistochemical (IHC) staining and
analysis
IHC assay were performed on a tissue microarray
containing samples from GC patients (HStm-Ade180Sur-06; Outdo
Biotech Co., Ltd., Shanghai, China). All procedures were in
accordance with the 1964 Declaration of Helsinki and its later
amendments. All patients received surgery between August 2008 and
March 2009. Excluding patients older than 70 years, 66 patients
(28–70 years old) with follow-up information remained in the
cohort. The clinical characteristics of patients are presented in
Table I. The pathological stage was
evaluated according to 7th American Joint Committee on Cancer
staging system (18).
 | Table I.Clinical characteristics of patients
with different levels of stanniocalcin-1 protein in gastric cancer
tissue. |
Table I.
Clinical characteristics of patients
with different levels of stanniocalcin-1 protein in gastric cancer
tissue.
|
| Low | High |
|
|---|
|
|
|
|
|
|---|
| Features | Patients, n | (%) | Patients, n | (%) | P-value |
|---|
| Total | 17 |
| 49 |
|
|
| Age, years |
|
|
|
| 0.567 |
|
<60 | 8 | 12 | 27 | 41 |
|
|
≥60 | 9 | 14 | 22 | 33 |
|
| Sex |
|
|
|
| 0.842 |
|
Male | 11 | 17 | 33 | 50 |
|
|
Female | 6 | 9 | 16 | 24 |
|
| Histologic
grade |
|
|
|
| 0.030 |
|
<III | 9 | 14 | 12 | 18 |
|
|
≥III | 8 | 12 | 37 | 56 |
|
| Nerve/vascular
invasion |
|
|
|
| 0.578 |
|
Negative | 13 | 20 | 34 | 52 |
|
|
Positive | 4 | 6 | 15 | 23 |
|
| T stage |
|
|
|
| 0.958 |
|
T1/2 | 2 | 3 | 6 | 9 |
|
|
T3/4 | 15 | 23 | 43 | 65 |
|
| N stage |
|
|
|
| 0.092 |
| N0 | 7 | 11 | 10 | 15 |
|
|
N1/2/3 | 10 | 15 | 39 | 59 |
|
| M stage |
|
|
|
| 0.130 |
| M0 | 17 | 26 | 43 | 65 |
|
| M1 | 0 | 0 | 6 | 9 |
|
| TNM stage |
|
|
|
| 0.021 |
|
I/II | 11 | 17 | 16 | 24 |
|
|
III/IV | 6 | 9 | 33 | 50 |
|
STC1 protein expression was detected by IHC
staining, and the adjacent section was stained with
hematoxylin-eosin and evaluated by two pathologists. The staining
and scoring system was performed as previously described (4). The primary antibody against STC1 (N15;
sc-14346; Santa Cruz Biotechnology, Inc.) was applied at a dilution
of 1:200 and incubated in a humid chamber at 4°C overnight. The
staining intensity was evaluated as follows: Low, negative or ≤40%
weak staining; High, >40% cells weak staining, or any cells with
moderate or strong staining).
Statistical analysis
Data expressed as the mean ± standard deviation (SD)
of three independently repeated experiments. Error bars in all
graphs present the SD. Student's t-test was used to evaluate the
difference between every two different groups. A Pearson
χ2 test was used to evaluate the association between
different clinicopathological characteristics of patients. A
Kaplan-Meier log-rank test was used to analysis the survival of
patients. P<0.05 was considered to indicate a statistically
significant difference. SPSS 16.0 (SPSS, Inc., Chicago, IL, USA)
was used to perform the analysis.
Results
Elevated STC1 is associated with poor
survival in GC
To examine the potential role of STC1 in patients
with GC, we first investigated the large online database
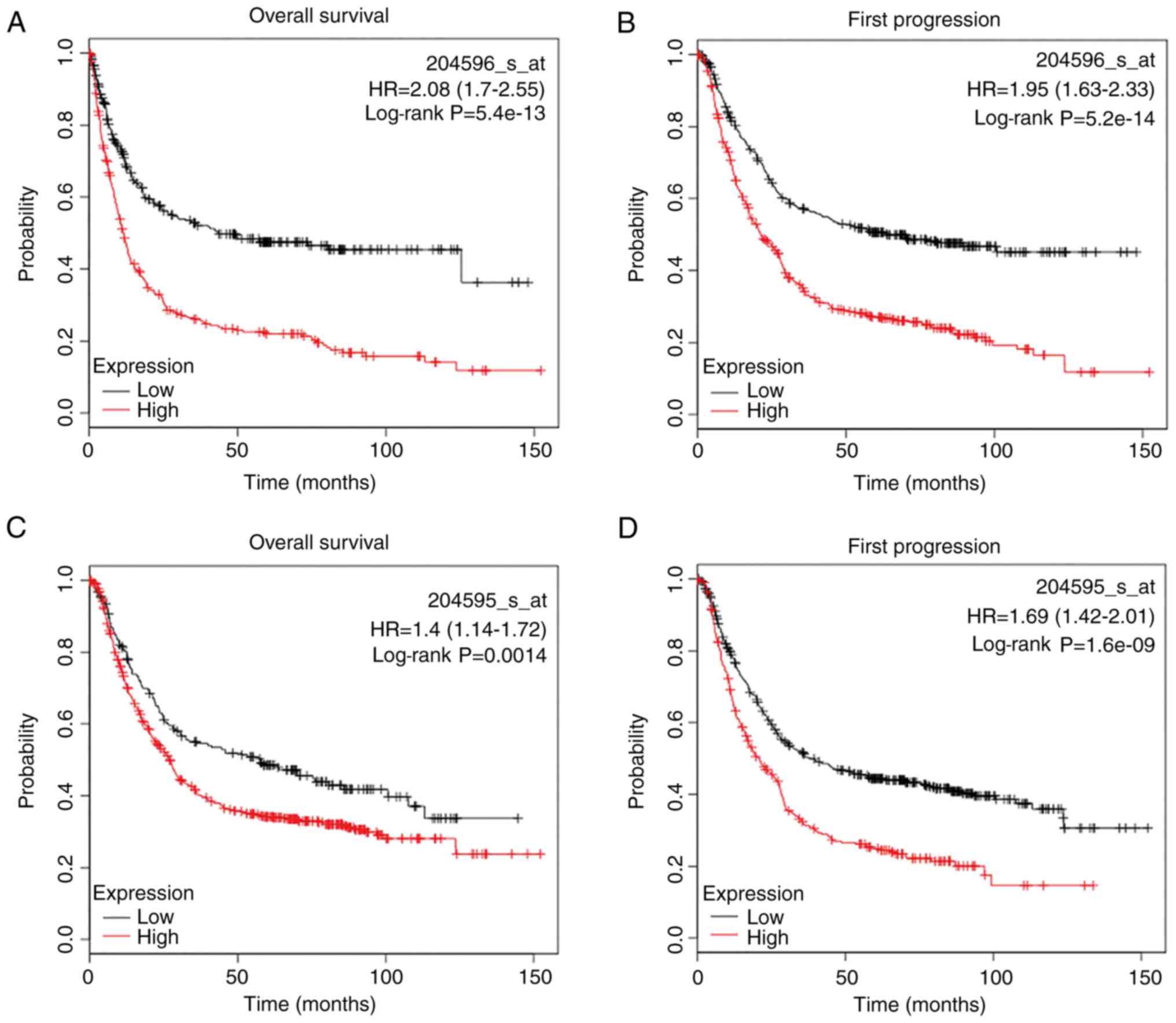
Kaplan-Meier Plotter. It was demonstrated that with different
probes, higher expression of STC1 mRNA was associated with worse
overall survival (OS) and first progression (FP) in patients with
GC (Fig. 1A-F).
To further validate the association between STC1 and
prognosis, IHC staining was used to evaluate the expression of the
STC1 protein in tissue samples from GC. The clinicopathological
characteristics of patients are summarized in Table I. Higher expression of STC1 in
cancer tissue was associated with worse OS (P=0.033; Fig. 1G and H). Additionally, correlation
analysis indicated that higher levels of STC1 in cancer tissue was
associated with poorer differentiation (P=0.030) and more advanced
tumor node and metastasis stages (P=0.021; Table I).
Collectively, these results suggested that
overexpression of STC1 was associated with poor prognosis in
patients with GC.
Expression of STC1 is enhanced in
hypoxic GC cells
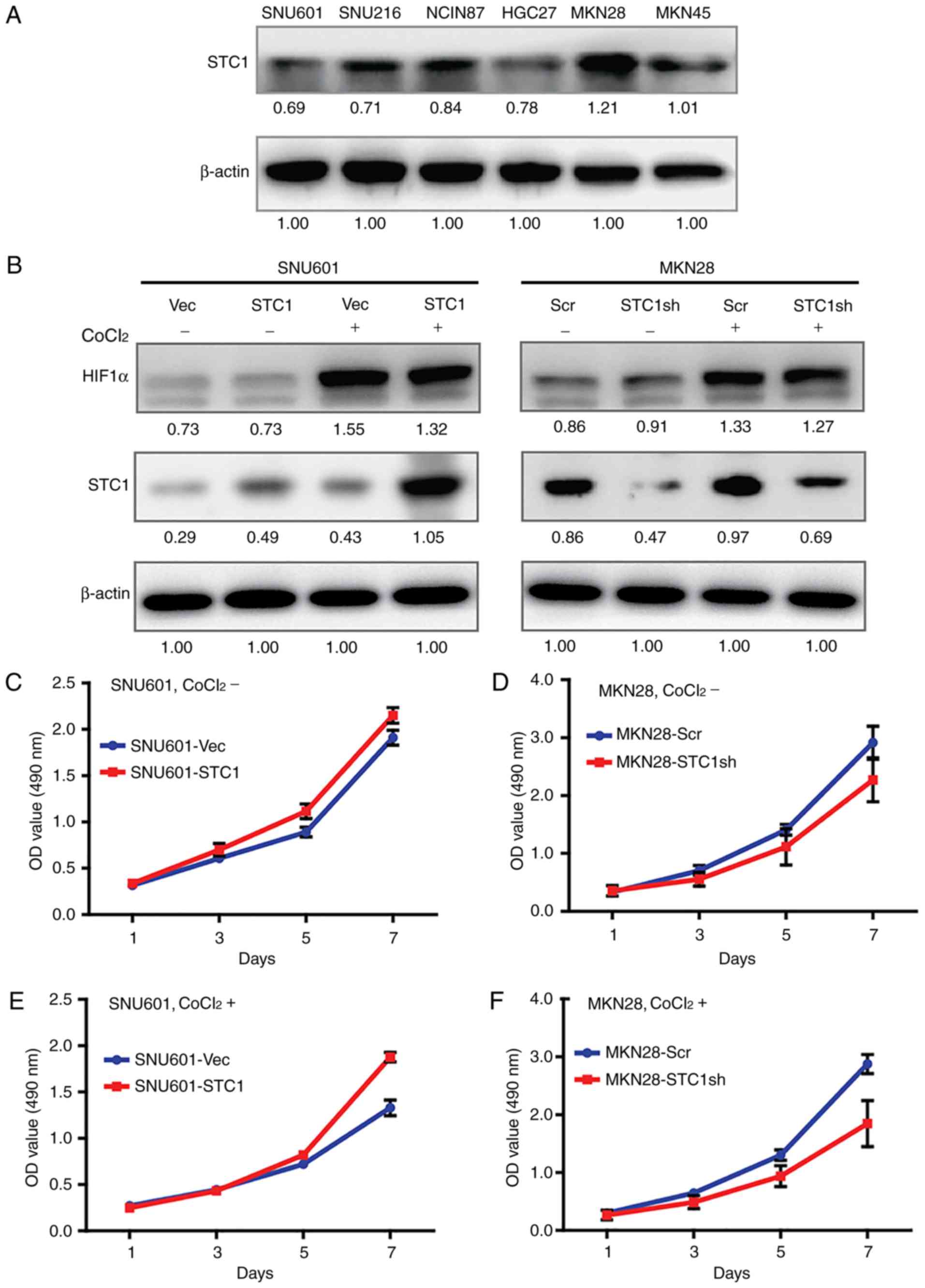
To investigate the roles of STC1 in GC in
vitro, the expression of STC1 in a set of GC cell lines was
screened (Fig. 2A). In the cells
which expressed the lowest level of STC1 (SNU601), STC1 cDNA and
the vector control was overexpressed; while in the cells which
expressed the highest level of STC1 (MKN28), the expression of STC1
was knocked down with shRNA (Fig.
2B).
CoCl2 was used, which inhibits the
activity of the key enzyme in the oxygen-sensing pathway (19), to stimulate hypoxia. Results from
immunoblotting analysis demonstrated that the expression of HIF1-α,
the well-established marker of hypoxia (20), increased in cells treated with
CoCl2, indicating that the hypoxic environment had been
successfully mimicked in vitro (Fig. 2B). Furthermore, the levels of STC1
in hypoxic cells were always increased compared with the same cell
lines in normoxic conditions (Fig.
2B), indicating that STC1 may serve certain roles in hypoxic
cells.
STC1 promotes cell proliferation and
chemoresistance in hypoxia
Previous studies demonstrated that STC1 promoted
cell proliferation in a series of cancer cells (4,6,21)..
However, the effects of STC1 on cell growth in normoxic or hypoxic
GC cells have not been reported, to the best of our knowledge. In
the present study, in cells cultured with normal medium the growth
curves of SNU601-STC1 cells and SNU601-Vec cells could not be
distinguished clearly (Fig. 2C), as
well as the curves of MKN28-STC1sh cells and MKN28-Scr cells
(Fig. 2D). However, when cells were
co-cultured with CoCl2, SNU601-STC1 cells proliferated
more compared with the SNU601-Vec cells (Fig. 2E), while MKN28-STC1sh cells grew
more slowly than MKN28-Scr cells (Fig.
2F). The results indicated that overexpression of STC1 promoted
cell proliferation in GC cells in hypoxia but not in normoxia.
The influence of STC1 on chemoresistance in GC was
measured further and the influence of STC1 on the cell response to
cisplatin in a hypoxic environment was focused on. Results from
IC50 (Fig. 2G) and flow
cytometry (Fig. 2H-I) demonstrated
that, in hypoxic SNU601-STC1 cells, the IC50 of
cisplatin was ~82 µM, and the proportions of late and total
apoptotic cells were 16.9 and 25.0%, respectively. However, in
hypoxic SNU601-Vec cells, the IC50 of cisplatin was only
71 µM, and the ratio of cells in late and total apoptosis was 38.5
and 47.4%, respectively. Furthermore, compared with MKN28-Scr
cells, the IC50 of MKN28-STC1sh cells was decreased by
16%, and the percentage of late and total apoptotic cells was
improved by 33 and 23%, respectively. Therefore, enhanced
expression of STC1 facilitated the resistance to cisplatin
treatment in hypoxic GC cells.
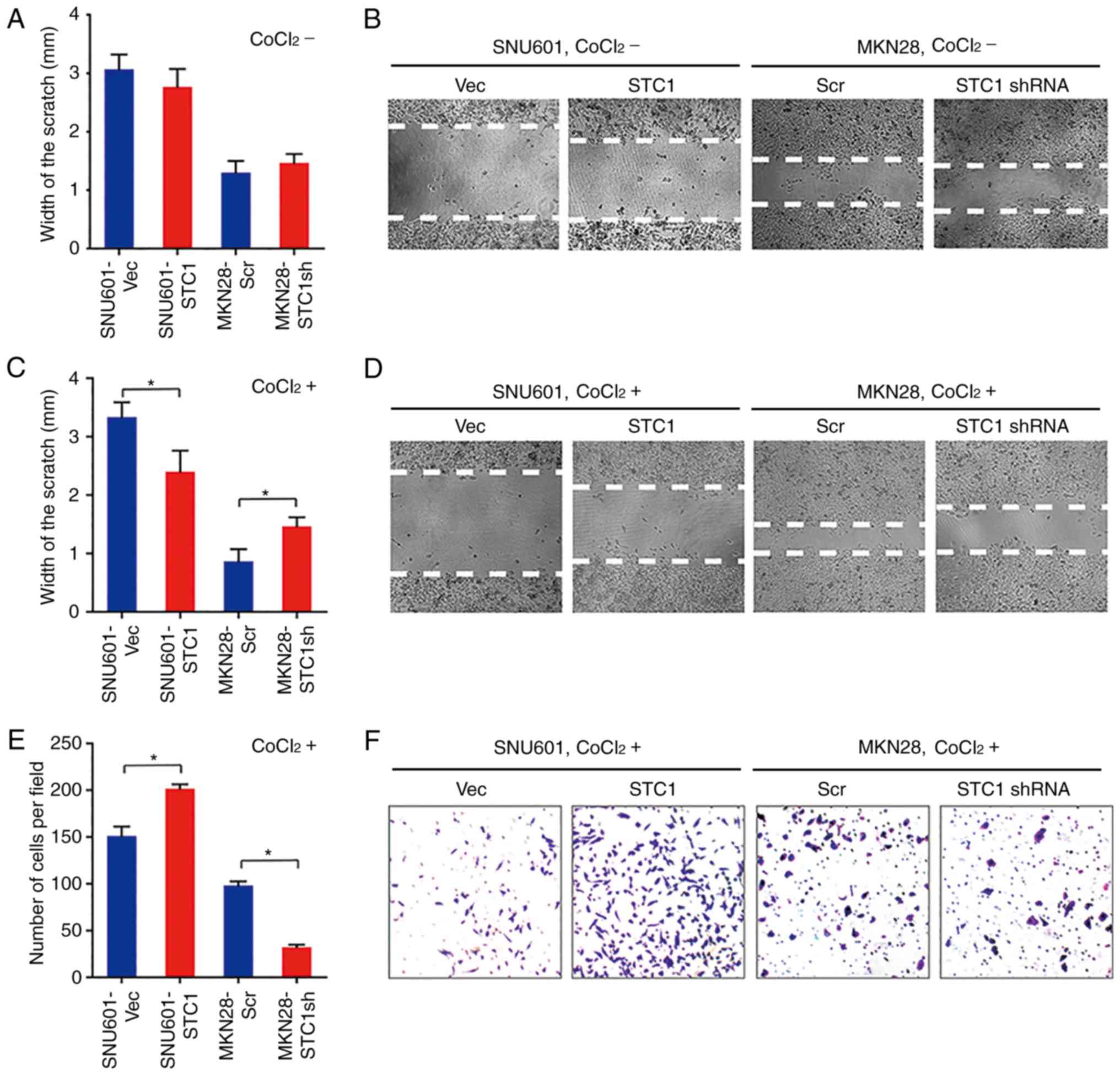
STC1 accelerates hypoxia-induced cell
migration and invasion
To elucidate the roles of STC1 in cell migration in
GC, wound-healing assays were performed in normoxia and hypoxia. At
72 h following scratching, in cells cultured without
CoCl2 the widths of wounds in SNU601-STC1 cells were
slightly narrower than in SNU601-Vec cells, and the widths in
MKN28-STC1sh cells were wider compared with the MKN28-Scr cells but
the change was not statistically significant (Fig. 3A and B). However, in cells treated
with CoCl2, the wounds in SNU601-STC1 cells were almost
healed, while those in SNU601-Vec cells remained large (Fig. 3C and D). Similarly, the gaps in
hypoxic MKN28-STC1sh cells were obviously wider compared with the
control cells (Fig. 3C and D). The
results suggested that overexpression of STC1 promotes cell
migration in hypoxia, but not in normoxia.
Therefore, the function of STC1 on cell invasion
under hypoxic conditions was further investigated. At 72 h
following seeding, there were 33% more cells invading through the
gel in SNU601-STC1 cells compared within control cells, while there
were 60% fewer cells moving into the bottom chamber in MKN28-STC1sh
cells compared with in MKN28-Scr cells (Fig. 3E and F). Collectively, in hypoxic GC
cells, higher levels of STC1 facilitated cell invasion.
STC1 inhibits apoptosis and promotes
invasion via Bcl-2
To investigate the mechanisms of STC1-promoted
malignant properties under hypoxic conditions, the expression of
key proteins in cells treated with CoCl2 was measured.
As presented in Fig. 4A, compared
with those in SNU601-Vec cells, the expression of Bcl-2 was
increased, while the levels of cytochrome c, cleaved-caspase-9 and
cleaved-caspase-3 were decreased in SNU601-STC1 cells. In contrast,
compared with the MKN28-Scr cells, the level of Bcl-2 was
decreased, but those of cytochrome c, cleaved-caspase-9 and
cleaved-caspase-3 were increased in MKN28-STC1sh cells. On the
other hand, compared with corresponding control cells, E-cadherin
was downregulated, but N-cadherin and MMP2 were upregulated in
SNU601-STC1 cells; whereas E-cadherin was upregulated, but
N-cadherin and MMP2 were downregulated in MKN28-STC1sh cells. The
expression of XIAP and MMP1 were not markedly changed according to
the different expression of STC1 (data not shown). The results
indicated that the expression of STC1 was positively associated
with that of Bcl-2, N-cadherin and MMP2, but negatively associated
with that of cytochrome c, cleaved-caspase-9, cleaved-caspase-3 and
E-cadherin.
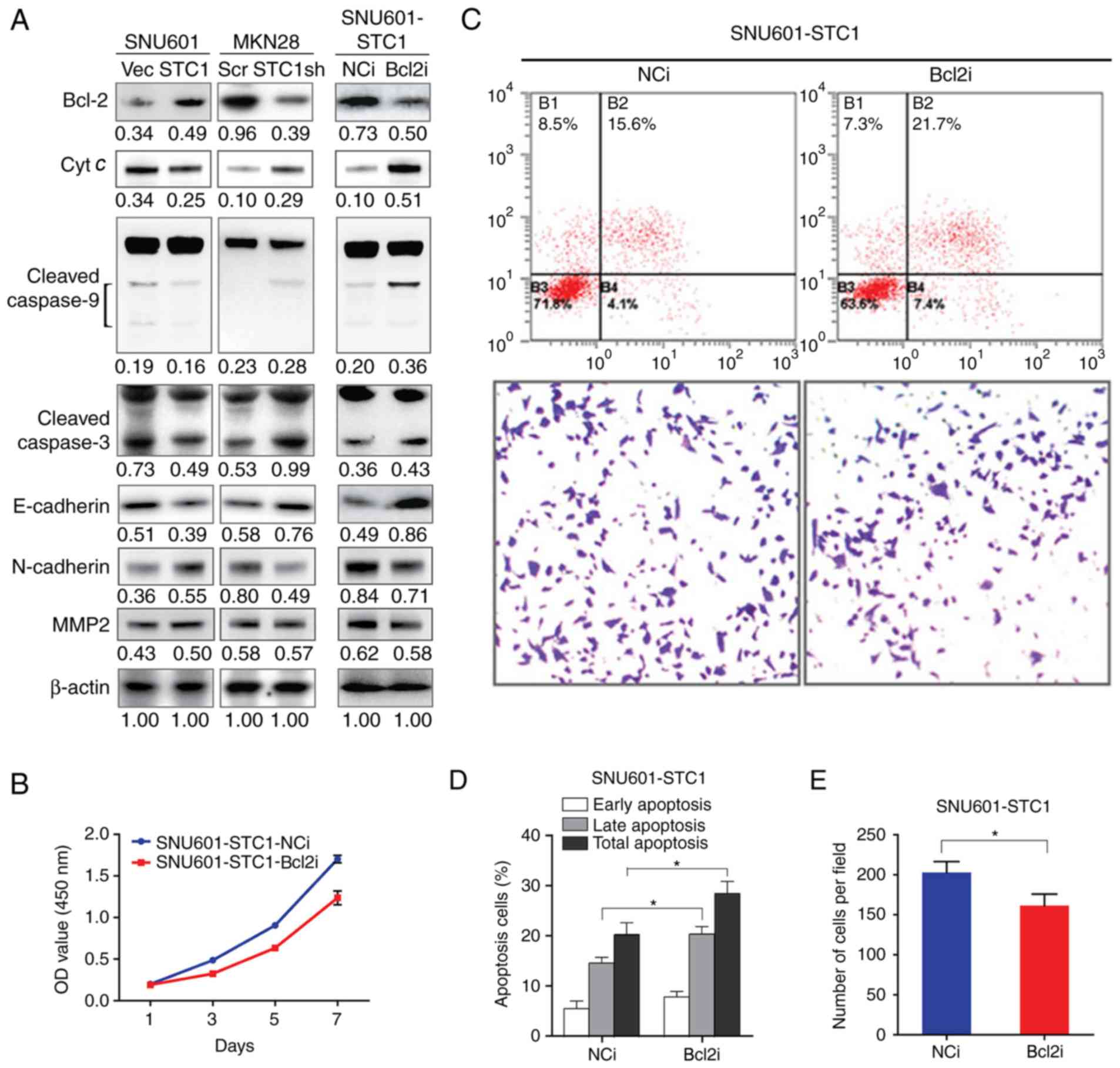 | Figure 4.STC1 inhibits apoptosis and promotes
invasion via Bcl-2. (A) Immunoblotting analysis of the protein
levels of Bcl-2, Cyt c, cleaved-caspase-9, cleaved-caspase-3,
E-cadherin, N-cadherin, and MMP2. For caspase-9 and −3, upper bands
present procaspase-9 and −3, and lower bands present
cleaved-caspase-9 and −3. β-actin was used as loading control. The
number below each band indicated the integrated density normalized
to the density of corresponding β-actin. (B) Cell proliferation,
(C) representative images of cell apoptosis tested by flow
cytometry and cell invasion tested by Transwell assays
(magnification, ×20). (D) Cell apoptosis in response to cisplatin
and (E) cell invasion in cells treated with Bcl-2 siRNA or NC
siRNA. All experiments were performed in presence of
CoCl2. *P<0.05. NC, negative control; E, epithelial;
N, neural; Bcl, B cell lymphoma; MMP, matrix metalloproteinase;
STC1, stanniocalcin-1; NC, normal control; siRNA, small interfering
RNA; Vec, empty vector; sh, small hairpin RNA; Scr, scramble shRNA;
Bcl2i, Bcl-2 siRNA; NCi, normal control siRNA; OD, optical
density. |
As Bcl-2 is a well-characterized and relatively
upstream anti-apoptotic protein, the expression of Bcl-2 was
artificially knocked-down with siRNA in SNU601-STC1 cells (Fig. 4A), and the cells' biological
behavior was tested under hypoxic conditions. As presented in
Fig. 4B-D, in SNU601-STC1- Bcl2i
cells, cell proliferation was arrested, and the ratios of apoptotic
cells treated with cisplatin were raised in SNU601-STC1-Bcl2i
cells. Furthermore, under hypoxic conditions, the expression of
cytochrome c, cleaved-caspase-9, and cleaved-caspase-3 were
upregulated in SNU601-STC1- Bcl2i cells compared with those in
control cells (Fig. 4A). As Bcl-2
was also involved in the process of epithelial-mesenchymal
transition (EMT) (13,22,23),
the invasive ability of SNU601-STC1-Bcl2i cells was also measured.
The results demonstrated that the numbers of cells invading through
the Transwell chamber were decreased, and the expression of
E-cadherin was increased, but those of N-cadherin and MMP2 were
lower in hypoxic SNU601-STC1-Bcl2i cells compared with in
SNU601-STC1-NCi cells (Fig. 4C and
E). Therefore, downregulating the expression of Bcl-2 could
significantly reverse the aggressive properties induced by
STC1.
These results suggest that, in hypoxic GC cells,
STC1 may facilitate cell proliferation, chemoresistance and
metastasis via Bcl-2.
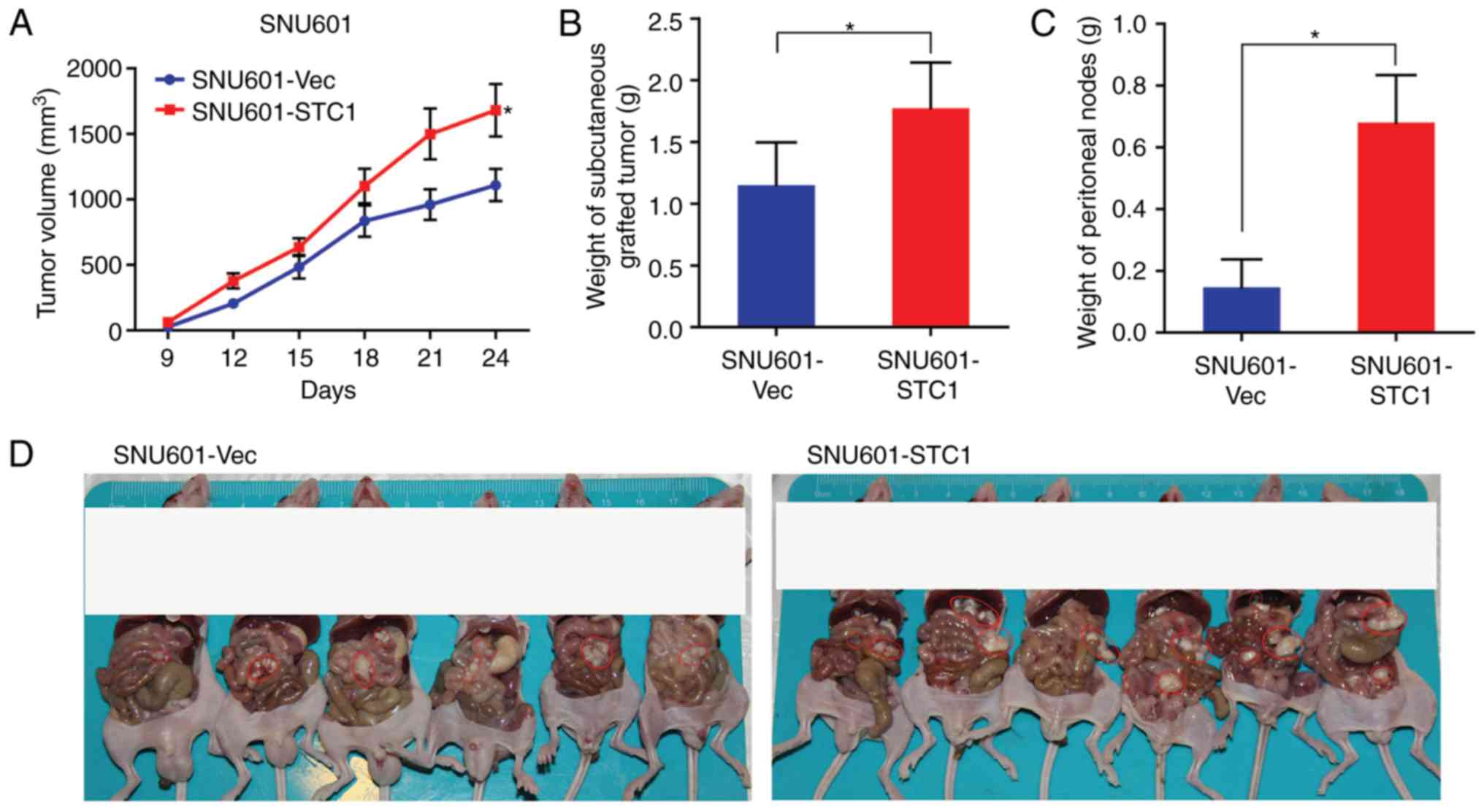
STC1 promotes tumor growth and
metastasis in vivo
To further test the influence of STC1 in
vivo, SNU601-STC1 cells and SNU601-Vec cells were injected into
nude mice.
As presented in Fig.
5A, the volumes of subcutaneous tumors withSNU601-STC1 cells
were increased compared with SNU601-Vec cells, and the disparities
increased with time. Furthermore, at the endpoint of the
experiments the weight of SNU601-STC1 tumors was increased compared
with that of SNU601-Vec tumors (Fig.
5B). Therefore, the results suggested that SNU601-STC1 tumors
grow faster than SNU601-Vec tumors.
Additionally the weight of peritoneal masses
produced with SNU601-STC1 cells was significantly heavier compared
with SNU601-Vec cells (Fig. 5C),
indicating that SNU601-STC1 cells caused increased peritoneal
metastasis compared with SNU601-Vec cells. Furthermore, the
malignant nodules in the abdominal cavity generated with
SNU601-STC1 cells were greater in number and larger compared with
SNU601-Vec cells (Fig. 5D).
In short, the results implied that overexpression of
STC1 promoted gastric tumor growth and metastasis in nude mice
in vivo.
Discussion
In the present study, the roles of STC1 in GC were
illustrated. It was demonstrated that the higher level of STC1 mRNA
and STC1 protein were associated with worse survival of patients
with GC. In vitro experiments identified that the expression
of STC1 is enhanced in hypoxic GC cells, and that overexpression of
STC1 facilitated cell proliferation in hypoxia but not in normoxia.
Furthermore, STC1 promoted chemoresistance, migration and invasion
in hypoxia. Additionally, under hypoxic conditions, the elevated
expression of STC1 was associated with an enhanced expression of
Bcl-2, N-cadherin and MMP2, and downregulated levels of cytochrome
c, cleaved-caspase-9, cleaved-caspase-3, and E-cadherin. However,
interfering with the expression of Bcl-2 increased the levels of
cytochrome c, cleaved-caspase-9, cleaved-caspase-3 and E-cadherin,
decreased the levels of N-cadherin and MMP2, and further restrained
cell proliferation, chemoresistance and cell invasion in hypoxia.
In vivo experiments also indicated that STC1 promotes
gastric tumor growth and metastasis. Collectively, STC1may promote
GC development, chemoresistance and metastasis via Bcl-2 under
hypoxic conditions (Fig. 6).
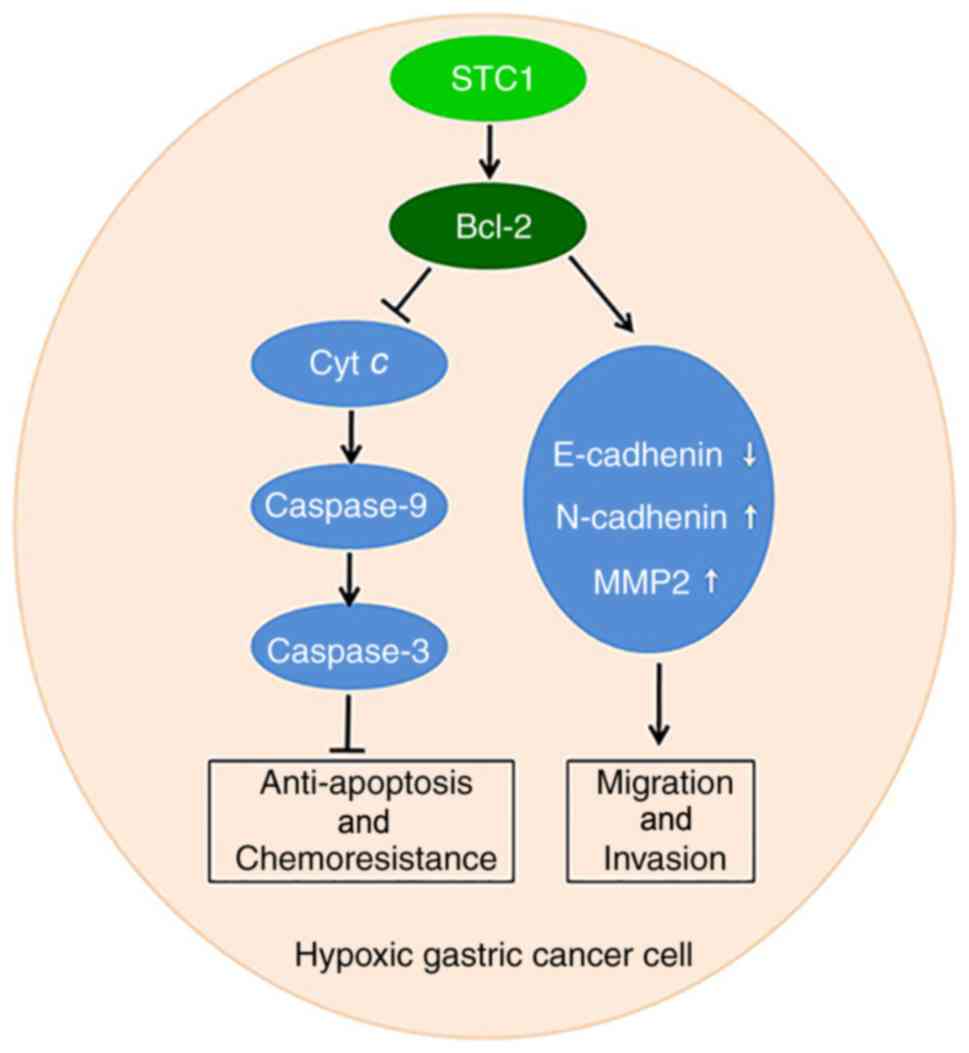 | Figure 6.Model of STC1-mediated GC progression
in hypoxia. In a hypoxic environment, elevated STC1 promotes the
expression of Bcl-2, and restrains the expression of cyt c and the
activation of caspase-9 and caspase-3 to inhibit apoptosis and
further promote cell proliferation and chemoresistance. It also
acts to restrain the expression of E-cadherin while increasing the
levels of N-cadherin and MMP2 to promote metastasis and invasion.
E, epithelial; N, neural; Bcl, B-cell lymphoma; MMP, matrix
metalloproteinase; GC, gastric cancer; STC1, stanniocalcin-1; Bcl,
B cell lymphoma; cyt c, cytochrome c. |
In patients with GC, a previous study reported that
the high/moderate level of STC1 protein was associated with poor
progression-free survival (7), but
the effects of STC1 on OS has not been illustrated. In the present
study, according to data in the Kaplan-Meier Plotter database, a
higher level of STC1 mRNA was associated with shorter OS and FP. In
another cohort or GC patients, it was demonstrated that a higher
level of STC1 protein in cancer tissues was associated with poorer
differentiation, more advanced clinical stage, and worse OS. The
results indicated that overexpression of STC1 may participate in GC
progression.
Hypoxia-induced genes make cells more tolerant to
the unfavorable conditions in certain cancer cell lines (24,25).
In accordance with studies in other cell lines (9,26–29),
it was demonstrated that STC1 could be stimulated in hypoxic
environments. However, the roles of STC1 under hypoxic conditions
are inconsistent according to different cell types. In mouse
embryonic fibroblast cells, STC1−/− cells
exhibited greater survival than STC1+/+ cells
(30). Whereas in rat
cardiomyocytes (31) and in mice
neural and renal cells (32,33),
STC1 serves protective roles. In the present study, it was
demonstrated that in GC cell lines, STC1 accelerated cell
proliferation in hypoxia but not in normoxia. Enhanced expression
of STC1 inhibited cell apoptosis in response to cisplatin treatment
in hypoxia.
According to the study by Liu et al (4) in ovarian cancer cells in normoxia,
STC1 appears to promote the expression of Bcl-2, a
well-characterized protein in apoptosis, leading to
caspase-9/-3-dependent cell death. In the present study, it was
demonstrated that in hypoxic cells the level of STC1 was positively
associated with the expression of Bcl-2, but negatively associated
with the levels of down stream cytochrome c, cleaved-caspase-9, and
the executioner cleaved-caspase-3. Downregulating the expression of
Bcl-2 upregulated the levels of cytochrome c, cleaved-caspase-9,
cleaved-caspase-3, and partly reversed the pro-growth and
anti-apoptosis roles of STC1. Furthermore, subcutaneous tumors with
enhanced expression of STC1 grew faster, and therefore were larger
and heavier compared with the control tumors. Collectively, the
results indicated that STC1 may promote the development and
chemoresistance of GC via the Bcl-2-mediated anti-apoptosis pathway
in hypoxia.
Several studies have reported that elevated STC1
promoted metastasis in various types of cancer, including GC
(7,27,34,35).
However, the studies on GC seldom involved cells in hypoxia. In
glioma cells, STC1 promoted cell migration in a hypoxia-dependent
manner, but did not exhibit pro-migration abilities under normoxic
conditions (36). However, the
mechanism was not clear. In the present study, it was demonstrated
that overexpression of STC1 facilitated both migration and invasion
in hypoxic GC cells. Previously, Bcl-2 was reported to promote EMT
and metastasis (13,37). Furthermore, Bcl-2 was also involved
in cell metastasis in hypoxia (38,39).
In the present study, it was demonstrated that the expression of
STC1 was positively associated with that of Bcl-2, N-cadherin and
MMP2, but negatively associated with the level of E-cadherin.
Furthermore, restraining the expression of Bcl-2 decreased the
level of N-cadherin and MMP2, but increased that of E-cadherin, and
partially reversed the invasive potential of STC1 overexpressing
cells in hypoxia. Furthermore, cells with elevated levels of STC1
formed an increased number and heavier metastatic nodes
inperitoneal cavities compared with control cells. Therefore, the
results suggested that under hypoxic conditions STC1 may promote
metastasis via restraining the Bcl-2-adjusted EMT process in
GC.
In conclusion, the results of the present study
indicated that STC1 positively regulated Bcl-2 to promote GC
development, chemoresistance and metastasis under hypoxia, and STC1
and Bcl-2 may be considered as potential therapeutic targets in
GC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81502049 for Yan
Wang and 81572955 for Zhen Zhang).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YW, ZQ, MZ, WY, RH and GL performed the experiments
on cells and mice, the database analysis and IHC analysis. YW, XM
and ZZ designed the experiments. YW drafted the manuscript. XM and
ZZ edited the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The animal experiments were approved by the
Institutional Animal Care and Use Committee of Chinese Academy of
Sciences (Beijing, China). The use of human tissues was approved by
the Ethics Committee of Outdo Biotech Co., and the cooperating
hospitals and written informed consent was obtained from all
patients.
Patient consent for publication
Consent for the publication of the clinical and
pathological data was obtained from all patients who were involved
in the present study.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GC
|
gastric cancer
|
|
STC1
|
stanniocalcin-1
|
|
OS
|
overall survival
|
|
FP
|
first progression
|
References
|
1
|
Khazaei S, Rezaeian S, Soheylizad M,
Khazaei S and Biderafsh A: Global incidence and mortality rates of
stomach cancer and the human development index: An ecological
study. Asian Pac J Cancer Prev. 17:1701–1704. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sullivan R and Graham CH: Hypoxia-driven
selection of the metastatic phenotype. Cancer Metastasis Rev.
26:319–331. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vaupel P and Mayer A: Hypoxia in cancer:
Significance and impact on clinical outcome. Cancer Metastasis Rev.
26:225–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu G, Yang G, Chang B, Mercado-Uribe I,
Huang M, Zheng J, Bast RC, Lin SH and Liu J: Stanniocalcin 1 and
ovarian tumorigenesis. J Natl Cancer Inst. 102:812–827. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tamura S, Oshima T, Yoshihara K, Kanazawa
A, Yamada T, Inagaki D, Sato T, Yamamoto N, Shiozawa M, Morinaga S,
et al: Clinical significance of STC1 gene expression in patients
with colorectal cancer. Anticancer Res. 31:325–329. 2011.PubMed/NCBI
|
|
6
|
Chang AC, Doherty J, Huschtscha LI,
Redvers R, Restall C, Reddel RR and Anderson RL: STC1 expression is
associated with tumor growth and metastasis in breast cancer. Clin
Exp Metastasis. 32:15–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang Z, Tian Z, Luo K, Song H and Yi J:
Clinical significance of stanniocalcin expression in tissue and
serum of gastric cancer patients. Chinese J Cancer Res. 26:602–610.
2014.
|
|
8
|
Zhang K, Lindsberg PJ, Tatlisumak T, Kaste
M, Olsen HS and Andersson LC: Stanniocalcin: A molecular guard of
neurons during cerebral ischemia. Proc Natl Acad Sci USA.
97:3637–3642. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yeung HY, Lai KP, Chan HY, Mak NK, Wagner
GF and Wong CK: Hypoxia-inducible factor-1-mediated activation of
stanniocalcin-1 in human cancer cells. Endocrinology.
146:4951–4960. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shroff EH, Snyder C and Chandel NS: Bcl-2
family members regulate anoxia-induced cell death. Antioxid Redox
Signal. 9:1405–1409. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu L, Ning X, Sun L, Zhang H, Shi Y, Guo
C, Han S, Liu J, Sun S, Han Z, et al: Hypoxia-inducible factor-1
alpha contributes to hypoxia-induced chemoresistance in gastric
cancer. Cancer Sci. 99:121–128. 2008.PubMed/NCBI
|
|
12
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, MacLeod RA, Masters JR, Nakamura Y, Reid YA,
Reddel RR, et al: Check your cultures! A list of cross-contaminated
or misidentified cell lines. Int J Cancer. 127:1–8. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun T, Sun BC, Zhao XL, Zhao N, Dong XY,
Che N, Yao Z, Ma YM, Gu Q, Zong WK, et al: Promotion of tumor cell
metastasis and vasculogenic mimicry by way of transcription
coactivation by Bcl-2 and Twist1: a study of hepatocellular
carcinoma. Hepatology. 54:1690–1706. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Wang Z, Qi Z, Yin S, Zhang N, Liu
Y, Liu M, Meng J, Zang R, Zhang Z, et al: The negative interplay
between Aurora A/B and BRCA1/2 controls cancer cell growth and
tumorigenesis via distinct regulation of cell cycle progression,
cytokinesis, and tetraploidy. Mol Cancer. 13:942014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun H, Wang Y, Wang Z, Meng J, Qi Z and
Yang G: Aurora-A controls cancer cell radio- and chemoresistance
via ATM/Chk2-mediated DNA repair networks. Biochim Biophys Acta
1843. 934–944. 2014.
|
|
16
|
Hou J, Wang Z, Xu H, Yang L, Yu X, Yang Z,
Deng Y, Meng J, Feng Y, Guo X, et al: Stanniocalicin 2 suppresses
breast cancer cell migration and invasion via the
PKC/claudin-1-mediated signaling. PloS One. 10:e01221792015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Szasz AM, Lanczky A, Nagy A, Förster S,
Hark K, Green JE, Boussioutas A, Busuttil R, Szabó A and Győrffy B:
Cross-validation of survival associated biomarkers in gastric
cancer using transcriptomic data of 1,065 patients. Oncotarget.
7:49322–49333. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang W, Sun XW, Li CF, Lv L, Li YF, Chen
YB, Xu DZ, Kesari R, Huang CY, Li W, et al: Comparison of the 6th
and 7th editions of the UICC TNM staging system for gastric cancer:
Results of a Chinese single-institution study of 1,503 patients.
Ann Surg Oncol. 18:1060–1067. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Salnikow K, Donald SP, Bruick RK,
Zhitkovich A, Phang JM and Kasprzak KS: Depletion of intracellular
ascorbate by the carcinogenic metals nickel and cobalt results in
the induction of hypoxic stress. J Biol Chem. 279:40337–40344.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rath S, Anand A, Ghosh N, Das L, Kokate
SB, Dixit P, Majhi S, Rout N, Singh SP and Bhattacharyya A: Cobalt
chloride-mediated protein kinase Calpha (PKCalpha) phosphorylation
induces hypoxia-inducible factor 1alpha (HIF1alpha) in the nucleus
of gastric cancer cell. Biochem Biophys Res Commun. 471:205–212.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bai Y, Xiao Y, Dai Y, Chen X, Li D, Tan X
and Zhang X: Stanniocalcin 1 promotes cell proliferation via cyclin
E1/cyclin-dependent kinase 2 in human prostate carcinoma. Oncol
Rep. 37:2465–2471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zuo J, Ishikawa T, Boutros S, Xiao Z,
Humtsoe JO and Kramer RH: Bcl-2 overexpression induces a partial
epithelial to mesenchymal transition and promotes squamous
carcinoma cell invasion and metastasis. Mol Cancer Res. 8:170–182.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi J, Choi K, Benveniste EN, Rho SB,
Hong YS, Lee JH, Kim J and Park K: Bcl-2 promotes invasion and lung
metastasis by inducing matrix metalloproteinase-2. Cancer Res.
65:5554–5560. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sumiyoshi Y, Kakeji Y, Egashira A,
Mizokami K, Orita H and Maehara Y: Overexpression of
hypoxia-inducible factor 1alpha and p53 is a marker for an
unfavorable prognosis in gastric cancer. Clin Cancer Res.
12:5112–5117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu X and Kang Y: Hypoxia and
hypoxia-inducible factors: Master regulators of metastasis. Clin
Cancer Res. 16:5928–5935. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lal A, Peters H, St Croix B, Haroon ZA,
Dewhirst MW, Strausberg RL, Kaanders JH, van der Kogel AJ and
Riggins GJ: Transcriptional response to hypoxia in human tumors. J
Natl Cancer Inst. 93:1337–1343. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ma X, Gu L, Li H, Gao Y, Li X, Shen D,
Gong H, Li S, Niu S, Zhang Y, et al: Hypoxia-induced overexpression
of stanniocalcin-1 is associated with the metastasis of early stage
clear cell renal cell carcinoma. J Transl Med. 13:562015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Law AY, Ching LY, Lai KP and Wong CK:
Identification and characterization of the hypoxia-responsive
element in human stanniocalcin-1 gene. Mol Cell Endocrinol.
314:118–127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cornmark L, Lonne GK, Jogi A and Larsson
C: Protein kinase Calpha suppresses the expression of STC1 in
MDA-MB-231 breast cancer cells. Tumour Biol. 32:1023–1030. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nguyen A, Chang AC and Reddel RR:
Stanniocalcin-1 acts in a negative feedback loop in the prosurvival
ERK1/2 signaling pathway during oxidative stress. Oncogene.
28:1982–1992. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shi X, Wang J and Qin Y: Recombinant
adeno-associated virus-delivered hypoxia-inducible stanniocalcin-1
expression effectively inhibits hypoxia-induced cell apoptosis in
cardiomyocytes. J Cardiovasc Pharmacol. 64:522–529. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Westberg JA, Serlachius M, Lankila P,
Penkowa M, Hidalgo J and Andersson LC: Hypoxic preconditioning
induces neuroprotective stanniocalcin-1 in brain via IL-6
signaling. Stroke. 38:1025–1030. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu D, Shang H and Liu Y: Stanniocalcin-1
protects a mouse model from renal ischemia-reperfusion injury by
affecting ROS-mediated multiple signaling pathways. Int J Mol Sci.
17:E10512016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Arigami T, Uenosono Y, Ishigami S,
Hagihara T, Haraguchi N, Matsushita D, Yanagita S, Nakajo A,
Okumura H, Hokita S, et al: Expression of stanniocalcin 1 as a
potential biomarker of gastric cancer. Oncology. 83:158–164. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He LF, Wang TT, Gao QY, Zhao GF, Huang YH,
Yu LK and Hou YY: Stanniocalcin-1 promotes tumor angiogenesis
through up-regulation of VEGF in gastric cancer cells. J Biomed
Sci. 18:392011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yoon JH, Kim J, Kim KL, Kim DH, Jung SJ,
Lee H, Ghim J, Kim D, Park JB, Ryu SH, et al: Proteomic analysis of
hypoxia-induced U373MG glioma secretome reveals novel
hypoxia-dependent migration factors. Proteomics. 14:1494–1502.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
An J, Lv J, Li A, Qiao J, Fang L, Li Z, Li
B, Zhao W, Chen H and Wang L: Constitutive expression of Bcl-2
induces epithelial-Mesenchymal transition in mammary epithelial
cells. BMC Cancer. 15:4762015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Leng J, Song Q, Zhao Y and Wang Z: miR15a
represses cancer cell migration and invasion under conditions of
hypoxia by targeting and downregulating Bcl2 expression in human
osteosarcoma cells. Int J Oncol. 52:1095–1104. 2018.PubMed/NCBI
|
|
39
|
Duan Y, He Q, Yue K, Si H, Wang J, Zhou X
and Wang X: Hypoxia induced Bcl-2/Twist1 complex promotes tumor
cell invasion in oral squamous cell carcinoma. Oncotarget.
8:7729–7739. 2017. View Article : Google Scholar : PubMed/NCBI
|