Introduction
Prostate cancer (PC) is the most common cancer
diagnosed in males in more than one-half of all countries and is
the main cause of cancer-related death in males in 46 countries
(1). At diagnosis, in approximately
90% of cases, PC is organ-confined or only locally advanced
(2). Treatment is dependent on
multiple parameters including clinical stage and prostate-specific
antigen (PSA) levels; the options for localized PC are active
surveillance, local radiotherapy or prostatectomy (3). With the spread of the disease outside
the prostate, androgen deprivation therapy (ADT) by surgical or
chemical castration is the recommended therapeutic strategy
(3,4). Unfortunately, the response is only
transient and most patients will develop resistance to ADT and
progress towards castration-resistant prostate cancer (CRPC) in
~18–36 months (5). The androgen
receptor (AR) axis is still active during PC progression and is an
essential player in CRPC. Other options to achieve maximal androgen
deprivation are the direct blockage of AR function with competitive
antagonists or reduction of intra-tumoral androgen synthesis
(3,4). Moreover, taxanes appear to be the only
class of chemotherapeutic agents capable of acting on aggressive
castration-resistant clones which emerge during ADT (6,7)
besides being the first therapeutic option to provide a significant
overall survival (OS) advantage and improvement of the quality of
life in the setting of metastatic CRPC (mCRPC) (6). Despite these benefits, development of
resistance to docetaxel is inevitable, and PC will eventually
progress, and the treatment for patients progressing during or
after docetaxel is limited and the prognosis is poor. Resistance to
taxanes involves a multidrug resistance (MDR) phenotype with cross
reactivity to unrelated drugs, alterations in the apoptotic
signaling pathway and increased expression of transport proteins of
the ABC family of transporters (8,9).
Some new agents have been evaluated for mCRPC in
phase III clinical trials in the last 5 years. Although some
improvement in OS and symptomatic benefits have been obtained, none
of these agents is considered curative (10). Therefore, there is an urgent need
for ongoing research to accomplish new approaches able to bypass
the resistance mechanisms evolving in CRPC or, to block signaling
pathways that lead to CRPC (11,12).
Recently, pentacyclic triterpenes are emerging as important
anti-neoplastic molecules. Increasing evidence shows that
triterpenes target important biological events in cancer including
cell proliferation, migration, invasion, and alterations of
mechanisms involved in apoptotic control (13–15).
Moreover, they are also effective against neoplastic cells whose
resistance is mediated by ABC transporters (16–18).
In addition, triterpenes were shown to inhibit certain signaling
pathways important for the development, invasion/progression and
metastasis in PC cell lines (19,20)
and in experimental models of cancer (21).
The present study examined the effects of the
pentacyclic triterpene pomolic acid (PA) on docetaxel-resistant PC3
cells. Herein, we showed that PA strongly reduced the viability and
induced apoptosis in both parental (PC3) and docetaxel-resistant
(PC3R) cell lines. We also showed that PA bypasses the mechanisms
of resistance mediated by P-gp/ABCB1 and MRP1/ABCC1 activity. In
addition to this effect, PA reverted the alteration of various
markers of EMT in docetaxel-resistant PC3 cells.
Materials and methods
Reagents and cell lines
Docetaxel (Taxotere®) and MK571 were
purchased from Enzo Life Sciences, Inc. (Farmingdale, NY, USA).
Pomolic acid (PA), provided by BioBioPha Co. Ltd. (Yunnan, China),
was dissolved in dimethyl sulfoxide (DMSO) to a concentration of 10
mM, stored at −20°C and diluted in culture medium for use.
Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum
(FBS), penicillin and streptomycin were purchased from Life
Technologies, Inc. (Carlsbad, CA, USA). DMSO,
3-(4,5-dimethylthiozol-2-yl)-2,5-diphenyl-tetrazolium bromide
(MTT), propidium iodide (PI) and KO 143 were obtained from Sigma
Chemical Co./Merck KGaA (Darmstadt, Germany). Rhodamine 123
(Rho123) and 5-carboxyfluorescein diacetate (5-CFDA) were obtained
from Calbiochem/EMD/Merck KGaA. Doxorubicin (cat. no. ab120629),
vincristin (cat. no. ab120226), and anti-N-cadherin antibody
(polyclonal rabbit, cat. no. ab12221) were from Abcam (Cambridge,
MA, USA). Verapamil was obtained from Alexis Biochemicals (San
Diego, CA, USA). Mitoxantrone (cat. no. sc-207888) and PE-labeled
anti-MRP1 (QCRL-2) were obtained from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). Anti-E-cadherin/CDH1 antibody (HECD-1)
(monoclonal mouse, cat. no. 13-1700) was obtained from Thermo
Fisher Scientific, Inc. (Waltham, MA, USA). Anti-vimentin antibody,
clone SP20 (rabbit monoclonal, cat. no. VPRM17) and Vectashield
mounting medium (cat. no. H-1000) were from Vector Laboratories
(Burlingame, CA, USA). Anti-N-cadherin, clone SP90 (rabbit
monoclonal) was obtained from Sigma-Aldrich/Merck KGaA. Goat
anti-rabbit IgG, F(ab′)2 fragment conjugated to cyanine
Cy™3 (111-166-047), and goat anti-mouse IgG, F(ab′)2
conjugated to cyanine Cy™3 (111-586-072) were obtained from Jackson
ImmunoResearch Laboratories (West Grove, PA, USA).
The human prostate cell line PC3 (ATCC,
CRL-1435TM) was grown in DMEM, supplemented with 10%
heat-inactivated FBS, 100 U penicillin and 100 mg/ml streptomycin
in disposable plastic bottles at 37°C in 5% CO2. Cells
were sub-cultured using trypsin-EDTA every 3–4 days. The
docetaxel-resistant PC-3 cell line was developed by stepwise
increased concentrations of docetaxel, essentially as previously
described (22). PC3 cells were
continuously maintained in docetaxel, with treatments beginning at
4 nM. After treatment, the surviving cells were re-seeded into new
flasks and allowed to recover. The procedure was repeated until the
cells displayed resistance to treatment as evaluated by cellular
viability (MTT assay). New cycles were performed, in which the
docetaxel concentration was subsequently increased up to a final
concentration of 20 nM. Surviving cells were termed PC3R.
Cell viability assay
Cell viability was assessed using the MTT assay as
previously described (15).
Briefly, 180 ml of a PC3 or PC3R cell suspension (104
cells/well) was distributed in 96-well plates and incubated for 24
h at 37°C/5% CO2 to allow the culture to stabilize. The
cells were then treated with 20 ml of medium, various
concentrations of docetaxel (0.5, 1, 2, 4, 8, 12, 16, 20, 100, 500
and 1,000 nM); PA (1, 2.5, 5, 7.5, 10, 12.5, 15 µg/ml equivalent to
5.29, 10.57, 15.86, 21.15, 26.44, 31.73 µM); DMSO concentrations
for each dose were used as controls. After 72 h of incubation, the
culture was treated with 20 µl MTT (2.5 mg/ml) and maintained for 4
h at 37°C in the dark before the supernatant being discarded. The
formazan produced by reduction of the MTT by viable cells was
dissolved in DMSO and the optical density was measured in a
multi-mode microplate reader (Synergy™ HT; BioTek Instruments,
Inc., Winooski, VT, USA) at 570 nm (reference filter 630 nm).
Experiments were repeated at least 3 times. The results are
expressed as the mean ± SD of percent inhibition of cell
viability.
DNA fragmentation assay
Apoptosis was assessed by cell cycle analysis using
flow cytometry as previously described (15). After 24 h of resting, the plated
prostate cells (2×104/well) were treated with media or
various concentrations of pomolic acid (PA) (2.5, 5.0, 7.5, 10.0,
12.5 and 15.0 µg/ml) and incubated for another 48 h. After this
time, cells were harvested and re-suspended in a hypotonic
fluorescent solution (50 µg/ml PI and 0.1% Triton X-100 in 0.1% Na
citrate buffer) for 1 h, at 4°C in the dark. The cell cycle was
analyzed by flow cytometry (FL-2) (FACSCalibur; Becton Dickinson,
San Jose, CA, USA) to determine the sub-G0/G1 DNA content.
Sub-diploid populations were considered to be apoptotic. Data
acquisition and analysis were carried out by BD CellQuest software,
version 3.1f (BD Biosciences, San Jose, CA, USA). Each experiment
was repeated at least 3 times. The results are presented as
representative histograms and as mean ± SD of the percentage of the
DNA that was fragmented.
Activity of ABC transporter
proteins
The functional activity of the MDR proteins was
determined based on the intracellular accumulation of specific
substrates as previously described (15). For each experiment, cells
(1×105/well) were seeded into 24-well plates and
pre-incubated for 24 h at 37°C/5% CO2 to allow
stabilization of the culture. Plated cells were then incubated for
30 min with substrates specific for P-gp/ABCB1 (200 ng/ml Rho123),
MRP1/ABCC1 (5 µM CFDA) or BCRP/ABCG2 (3 µM mitoxantrone) in the
presence of medium or the conventional inhibitor of these proteins,
verapamil (50 µM), MK571 (50 µM) and KO143 (40 µM). Then the cells
were washed in phosphate-buffered saline (PBS), harvested and kept
on ice until flow cytometric analysis (FACSCalibur;
Beckton-Dickinson cytometer). In this condition, an increase in
cellular fluorescence correlates with transport activity. Results
are presented as representative histograms or as the mean ± SD of
arbitrary units of mean fluorescence intensity (MFI).
To assess the effect of PA on the activity of
P-gp/ABCB1 and MRP1/ABCC1, plated cells were incubated for 30 min
with medium (autofluorescence), with substrates specific for each
protein in the presence or absence of specific inhibitors or, with
substrate plus the desired concentration of PA. After harvesting,
the cells were analyzed as described above.
Immunocytochemical assays
PC3 or PC3R cells (2×104/well) were
seeded on coverslips in 24-well plates, left to rest for 24 h and
then treated with medium or 5 µg/ml of PA. After 48 h of
incubation, the cells were washed twice with PBS, pH 7.4, and fixed
with a 4% buffered paraformaldehyde solution containing 4% sucrose
for 40 min at 4°C. After washing in PBS, the coverslips were
permeabilized with PBS-0.5% Triton, pH 7.0 for 15 min, and then
blocked with PBS-5% BSA-0.1% Triton-Tween 0.05% for 1 h. After this
time, coverslips were incubated with the diluted primary antibodies
(E-cadherin 1:50; N-cadherin: 1:100; vimentin 1:50) at 4°C
overnight. Then the coverslips were washed in PBS-0.25% Tween and
incubated with secondary antibodies [goat anti-rabbit IgG,
F(ab′)2 fragment-Cy™3 (1:200), and goat anti-mouse IgG,
F(ab′)2 -Cy™3 (1:200)] diluted in PBS pH 7.4 for 1 h.
Next, the coverslips were washed 2 times with PBS-0.25% Tween and
then stained with 0.5 mg/ml DAPI for 5 min. Following washes with
PBS and distilled water, the coverslips were mounted with
Vectashield medium and observed in an epifluorescence microscope
(Eclipse E-800; Nikon Corp., Tokyo, Japan). The quantitative
analysis was performed using an image analysis system (Image-Pro
Plus 4.5; Media Cybernetics, Inc., Rockville, MD, USA) composed of
a digital camera (Evolution; Media Cybernetics, Inc.) coupled to a
fluorescence microscope. High quality images of cells were captured
(2048×1536 pixels buffer), using a 40× objective lens. At least 20
fields/coverslip were captured. For vimentin quantification, since
all cells were labelled, the percentage of immunoreactive cells was
calculated from the DAPI-positive cells.
Statistical analysis
The results are presented as mean ± standard
deviation (SD) of at least 3 independent experiments. Student's
t-test was used to analyze comparisons. One-way analysis of
variance (ANOVA) and Dunnett and Tukey multiple comparison test
were used to analyze differences among groups. P<0.05 was
considered to refer to statistically significant differences.
Statistical analysis was performed using GraphPad Prism 5 software
(version 5.01; GraphPad Software, Inc., La Jolla, CA, USA).
Results
Sensitivity of the cell lines to
docetaxel (DTX)
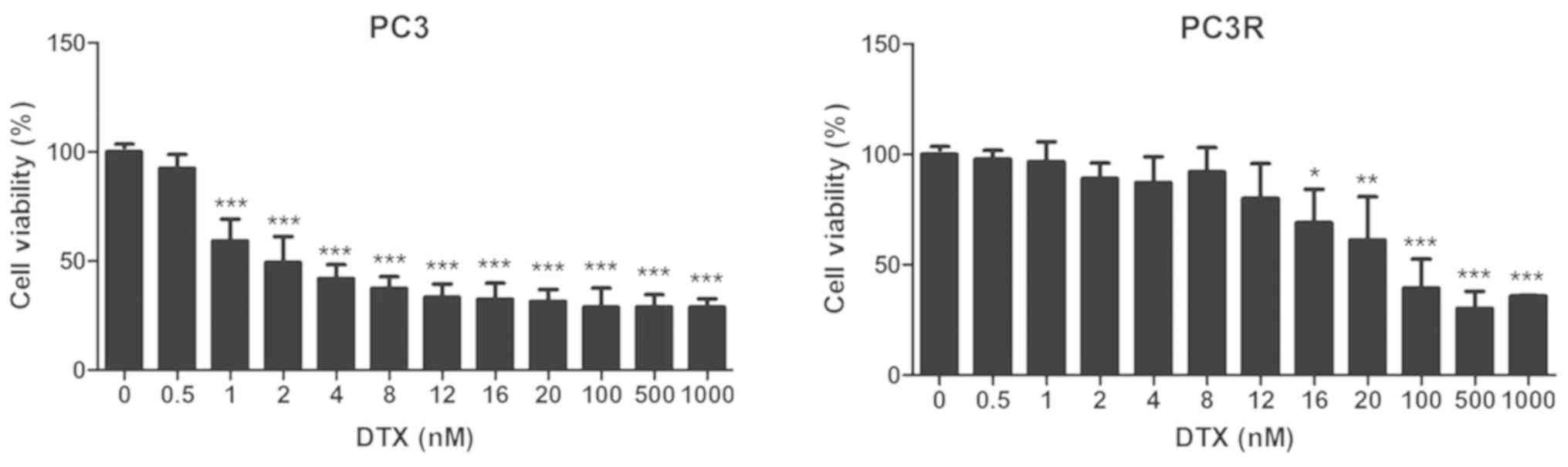
The MTT assay was used to evaluate the response of
PC3 cells to docetaxel (DTX). Cells were treated with different
concentrations of DTX and viability was assessed 72 h later.
Fig. 1 (left panel) shows that 2 nM
of DTX reduced PC3 viability by approximately 50%, demonstrating
the sensitivity of the cell line to DTX. Establishment of a
docetaxel-resistant PC3 cell line, PC3R, was accomplished by 4–5
cycles of treatment with stepwise increasing concentrations of the
drug from 4 to 20 nM, as described in Materials and methods. Higher
concentrations were avoided due to their toxicity. At this point,
cells were treated with DTX in the same conditions used for the
parental line and the cell viability was measured by MTT assay.
Fig. 1 (right panel) shows that the
treated cells (PC3R) were resistant to low concentrations of DTX,
being responsive only to higher concentrations of the drug.
DTX-resistant cell line PC3R displays
a multidrug resistance phenotype
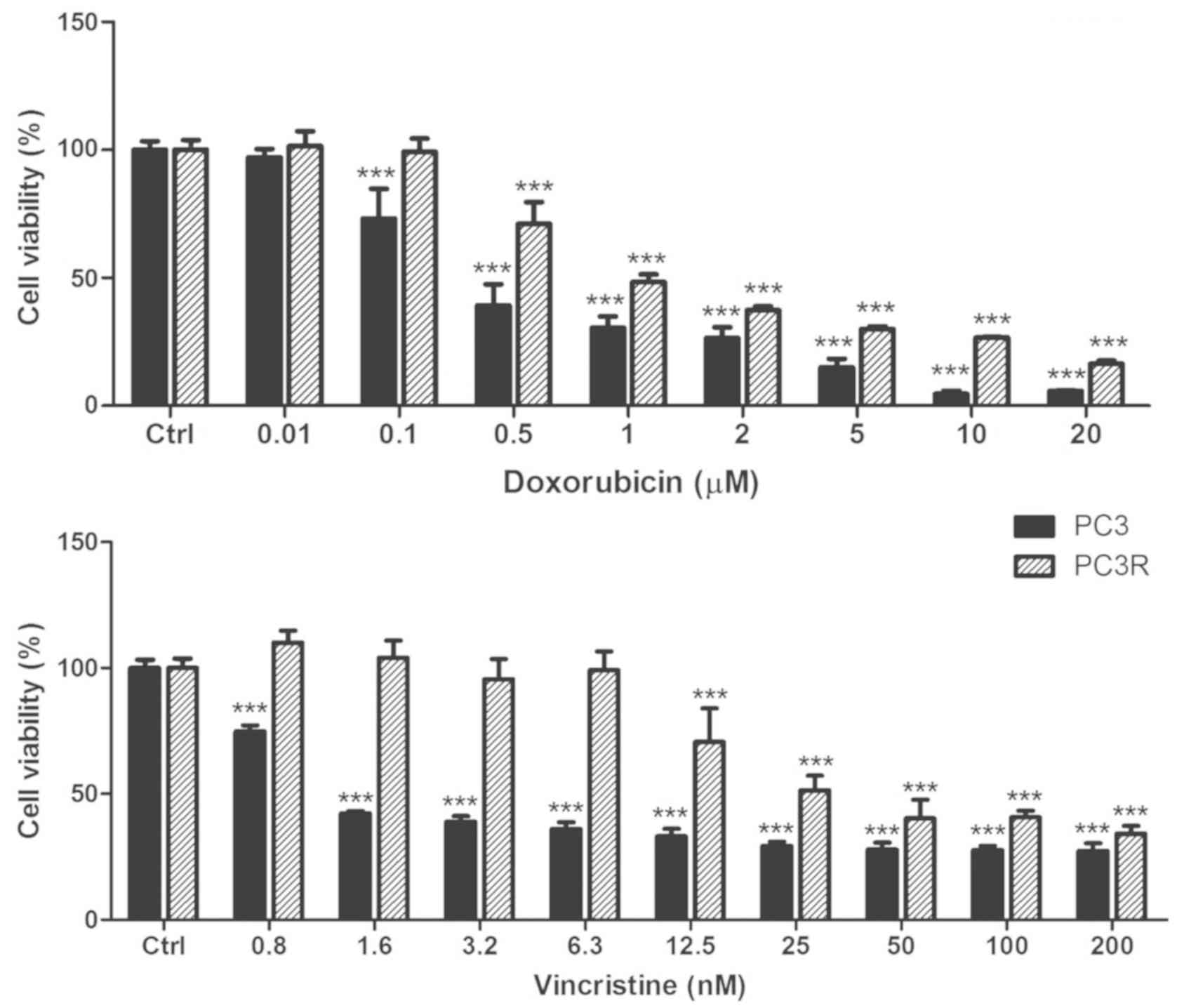
To ascertain whether the PC3R cell line presents
with a multidrug resistance phenotype, parental and resistant cells
were treated with doxorubicin or vincristin (Fig. 2) and the cell viability was assessed
by MTT assay, 72 h later. The results showed an increase in
resistance of the PC3R cells to both drugs corroborating the
multidrug resistance phenotype of this cell line.
Activity of ABC transporters on PC3
lineages
Extrusion of drugs by transporter proteins of the
ABC family is one of the main mechanisms of drug resistance. In
order to investigate the involvement of the transporter proteins in
DTX resistance, we analyzed the activity of P-gp/ABCB1, MRP1/ABCC1
and BCRP/ABCG2 on both parental and resistant cells. For this,
cells were incubated for 30 min with substrates for BCRP/ABCG2,
MRP1/ABCC1 and P-gp/ABCB1 (mitoxantrone, CFDA or Rho123) in the
presence or absence of inhibitors specific for each transporter
(KO143, MK571 or verapamil), and the cell fluorescence was
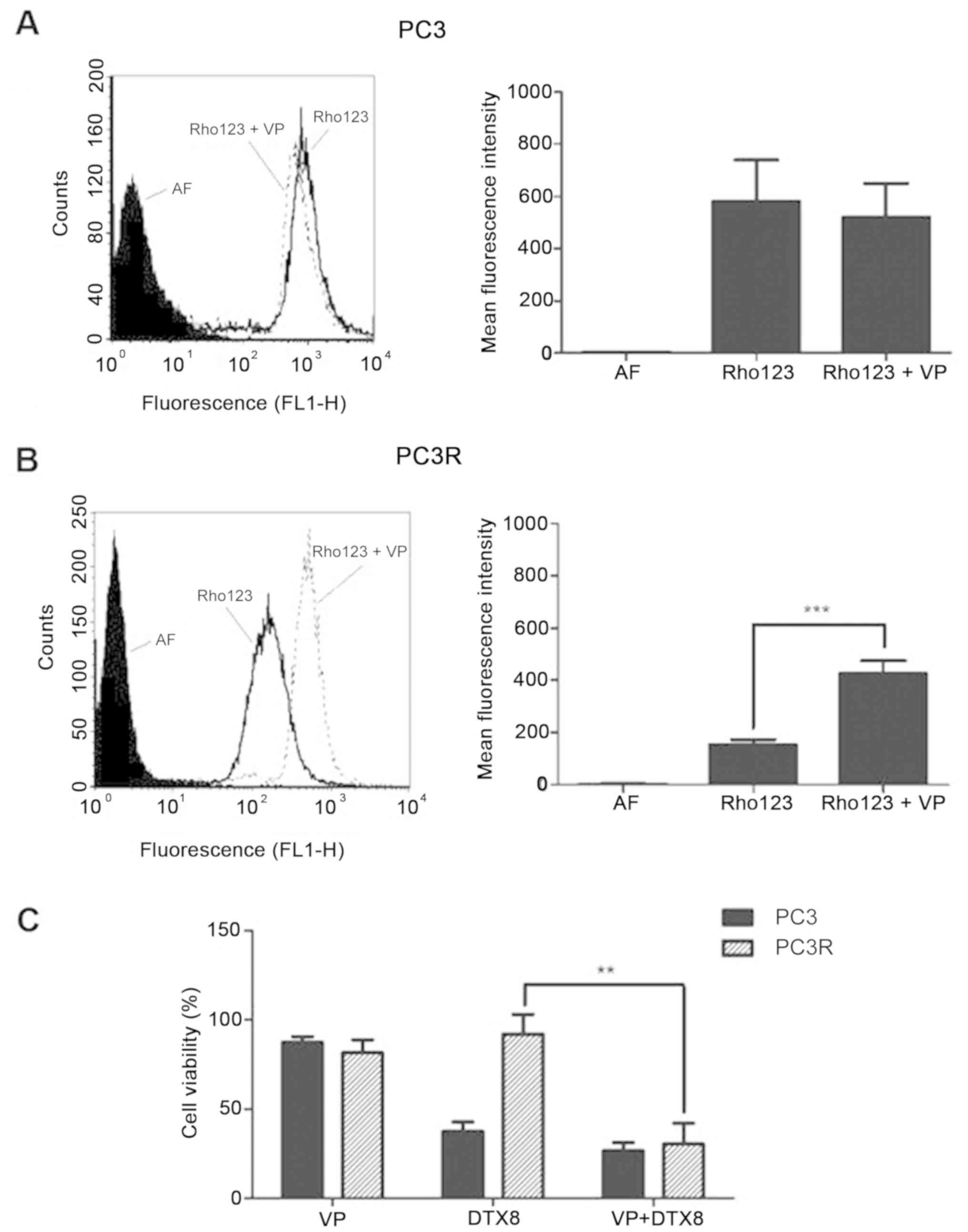
evaluated by flow cytometry. As shown in Fig. 3, while no P-gp/ABCB1 activity was
observed in the parental cell line (Fig. 3A), a significant increase in
activity (P<0.001) was observed in the DTX-resistant PC3R cell
line (Fig. 3B). To evaluate the
involvement of P-gp/ABCB1 activity in DTX resistance, cells were
treated with 8 nM DTX in the presence or absence of the P-gp
inhibitor verapamil (VP) and cell viability was measured by MTT
assay 72 h later. The results (Fig.
3C) suggest that inhibition of P-gp activity reverts the
resistance of the PC3R cell line to DTX.
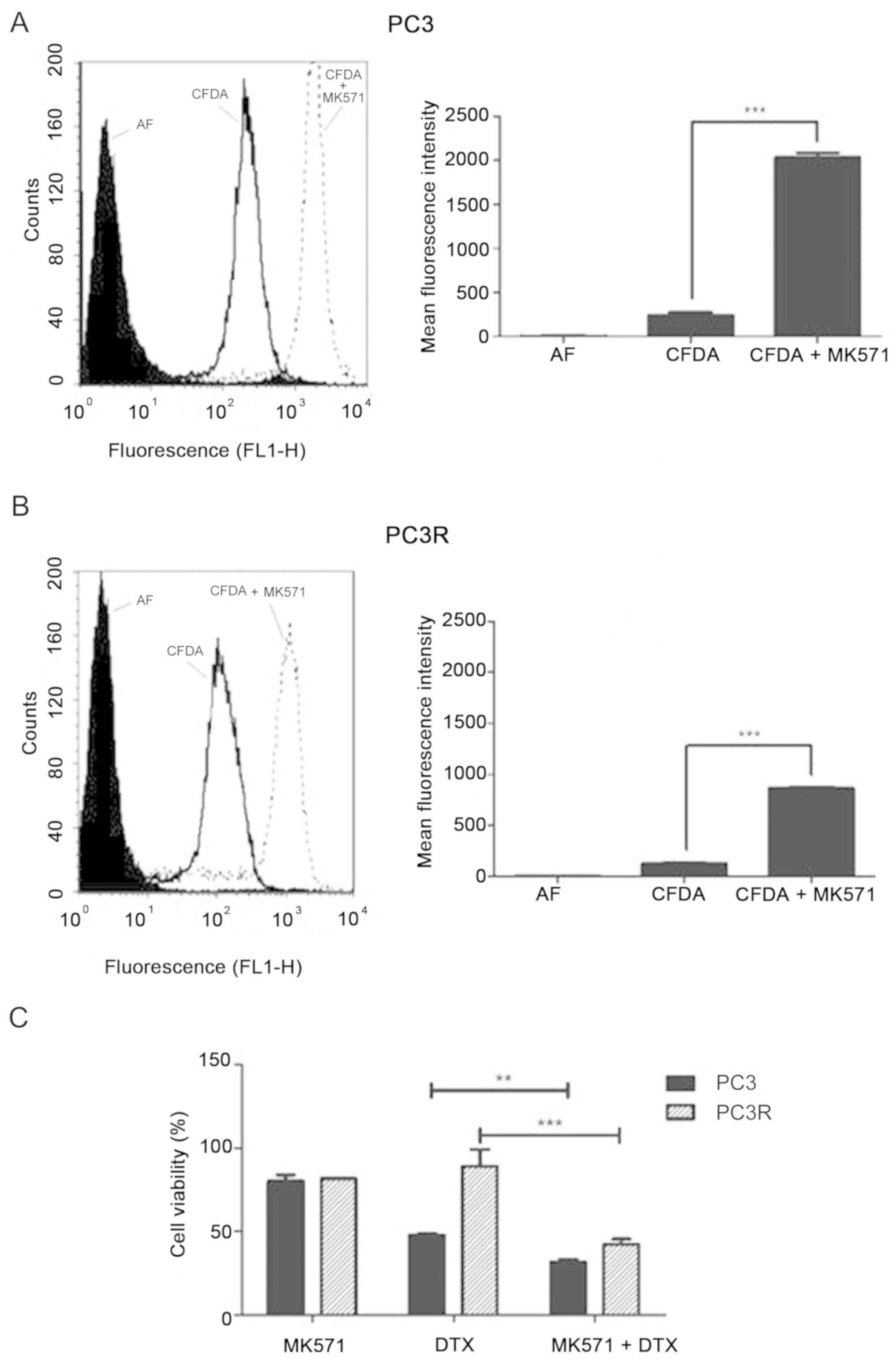
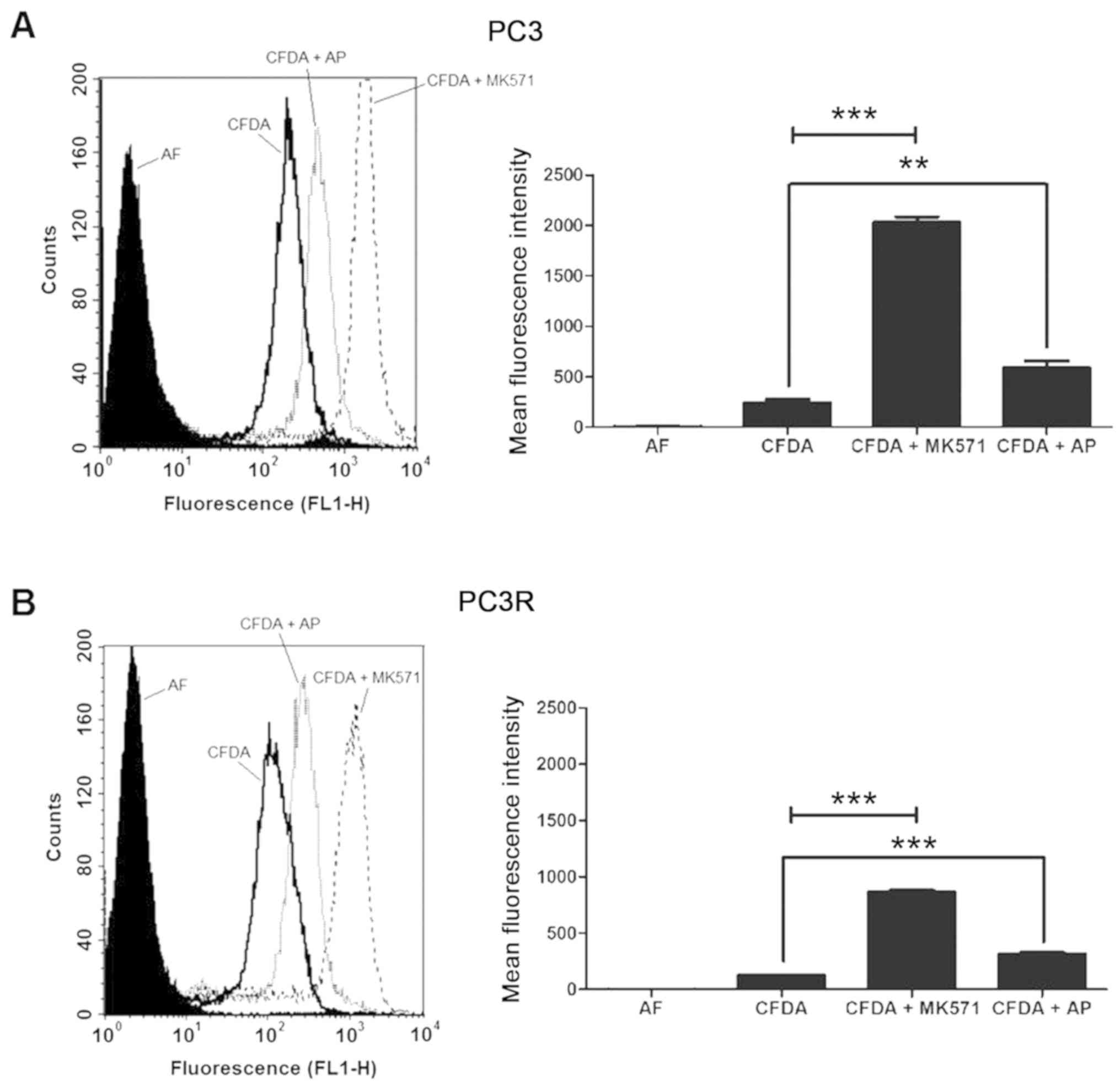
Different from P-gp, both PC3 and PC3R cell lines
displayed MRP1/ABCC1 activity (Fig. 4A
and B). To test the involvement of this protein in DTX
response, we measured the viability of the cells after 72 h
treatment with 8 nM DTX in the presence or absence of the
MRP1/ABCC1 inhibitor MK571. Although the co-treatment induced a
significant decrease in cell viability of both lineages this
response was more evident in the PC3R cells (Fig. 4C).
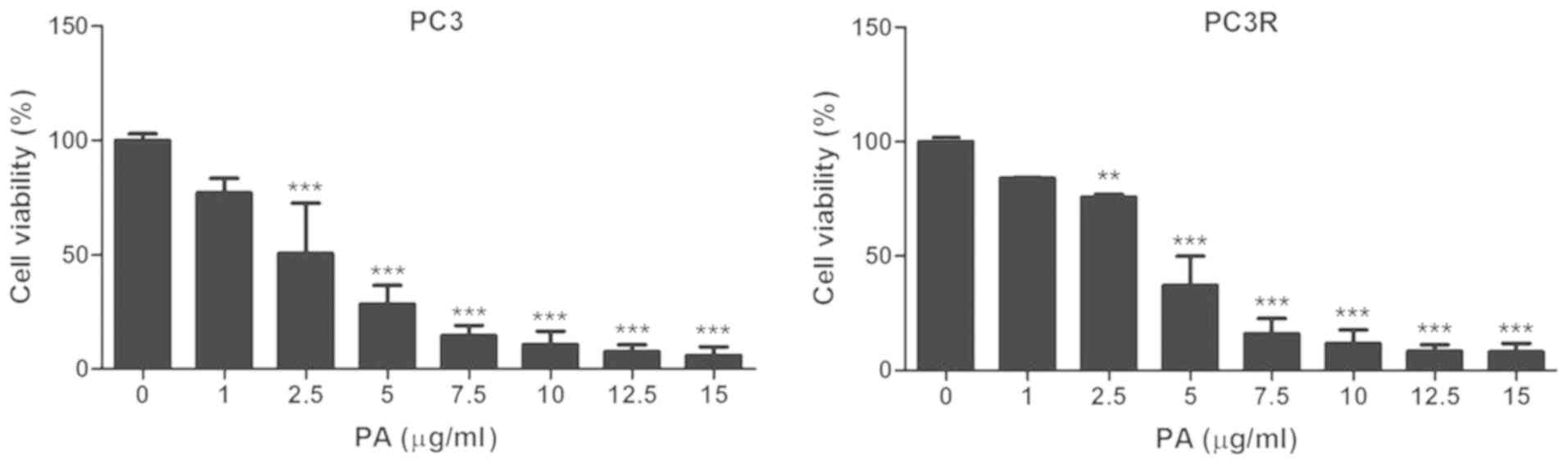
Pomolic acid induces cell death of the
PC3 lineages
However, even when these transporters were
inhibited, approximately 40% of the cells remained alive suggesting
the involvement of other resistance mechanisms in PC3 and PC3R
cells. These mechanisms are not mediated by BCRP/ABCG2 as a
negligible activity of this transporter was observed in the
parental and DTX-resistant cell lines (data not shown).
In an attempt to identify options for CRPC
treatment, we evaluated the effects of PA on the PC3R cell line.
For this, parental and resistant cell lines were incubated with
different concentrations of the triterpene and the cell viability
was assessed by MTT assay 72 h later. PA appeared to be more active
then DTX against PC as it acted on the two lineages in a similar
way decreasing their viability up to 90% (Fig. 5).
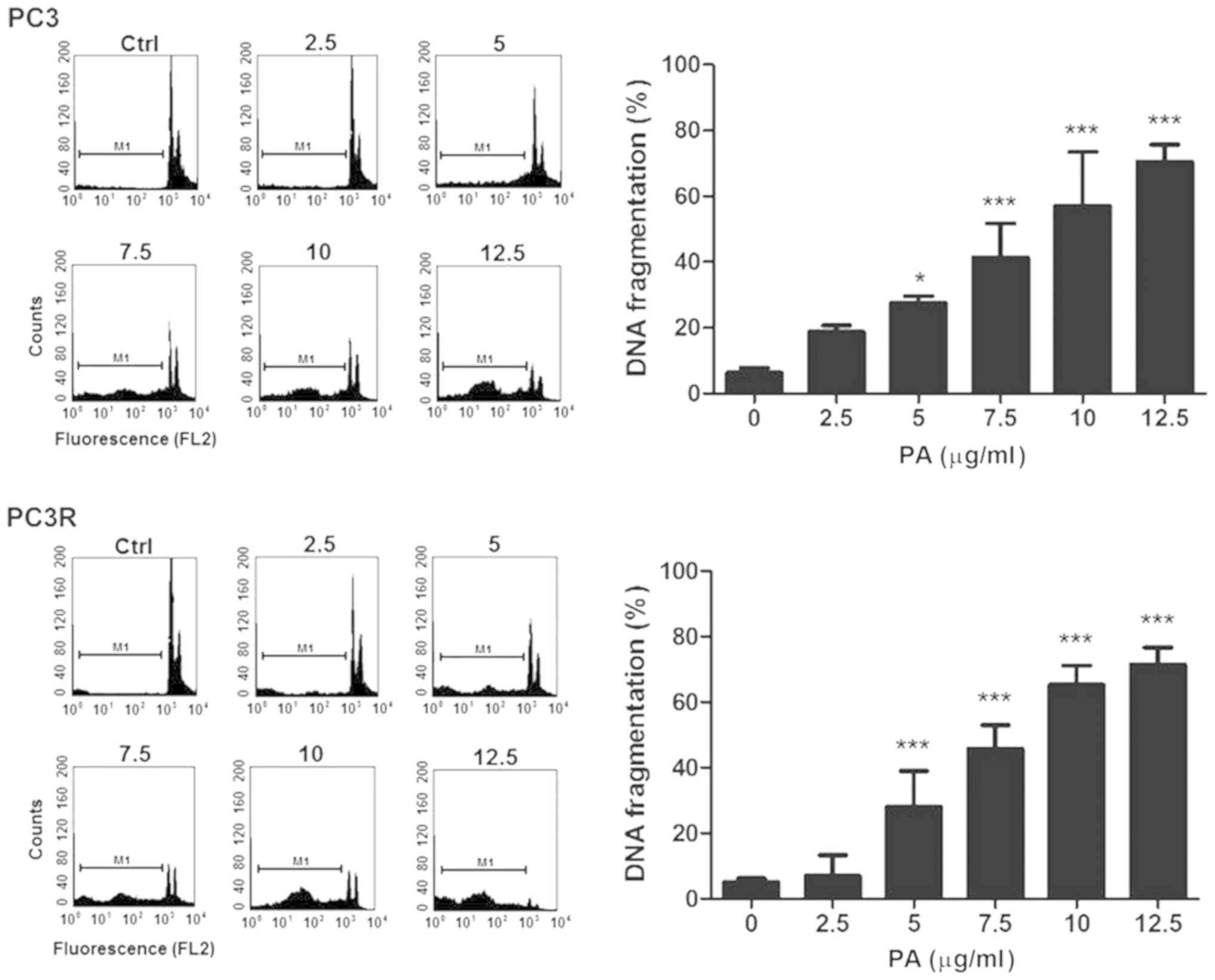
In order to investigate whether the decrease in cell
viability was due to apoptosis, we analyzed the DNA content of the
sub-G0/G1 population in the cell cycle after 48 h of PA treatment.
As shown in Fig. 6, PA increased
the DNA fragmentation of both lineages in a dose-dependent manner
indicating that death was mediated by apoptosis.
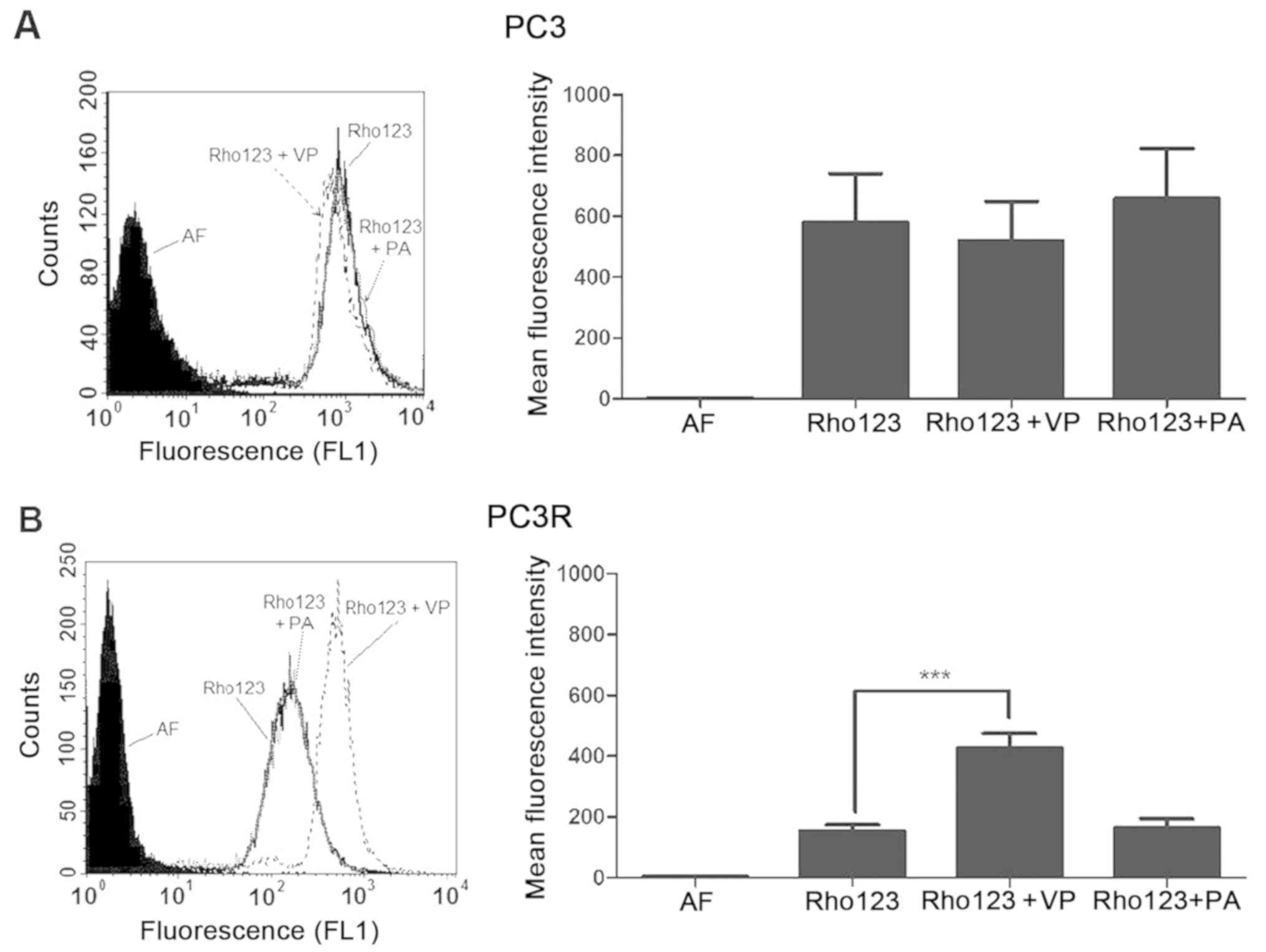
Pomolic acid has no effect on
P-gp/ABCB1 but downmodulates MRP1/ABCC1 activity
Since inhibition of P-gp/ABCB1 or MRP1/ABCC1
increases the susceptibility of the resistant cell line to DTX, we
analyzed whether the enhanced cytotoxic effect of PA would be due
to an effect on these proteins. PC3 and PC3R cells were incubated
with substrates of each protein in the presence or absence of
inhibitors or 15 µg/ml PA and accumulation of the substrate was
evaluated by cytometry. The results obtained demonstrated that
PA-induced death in PC3R is independent of the increase in
P-gp/ABCB1 since it does not interfere with the protein activity of
the cell line (Fig. 7). Moreover,
PA was able to modulate MRP1/ABCB1 activity in both parental and
PC3R cell lines (Fig. 8), although
less efficiently then the commercial inhibitor MK571.
Pomolic acid partially reverts EMT
changes induced by DTX treatment
Progression from the localized to invasive and
metastatic tumors is partially dependent on a process called
epithelial-mesenchymal-transition (EMT) in which the tumor
epithelial cells loose expression of adhesion molecules such as
E-cadherin and express proteins characteristic of mesenchymal cells
such as vimentin (23). This
process is also involved in the deregulation of androgenic
signalization that occurs during the development of CRPC and
confers resistance to apoptosis and stem cell like properties to
tumoral cells (12).
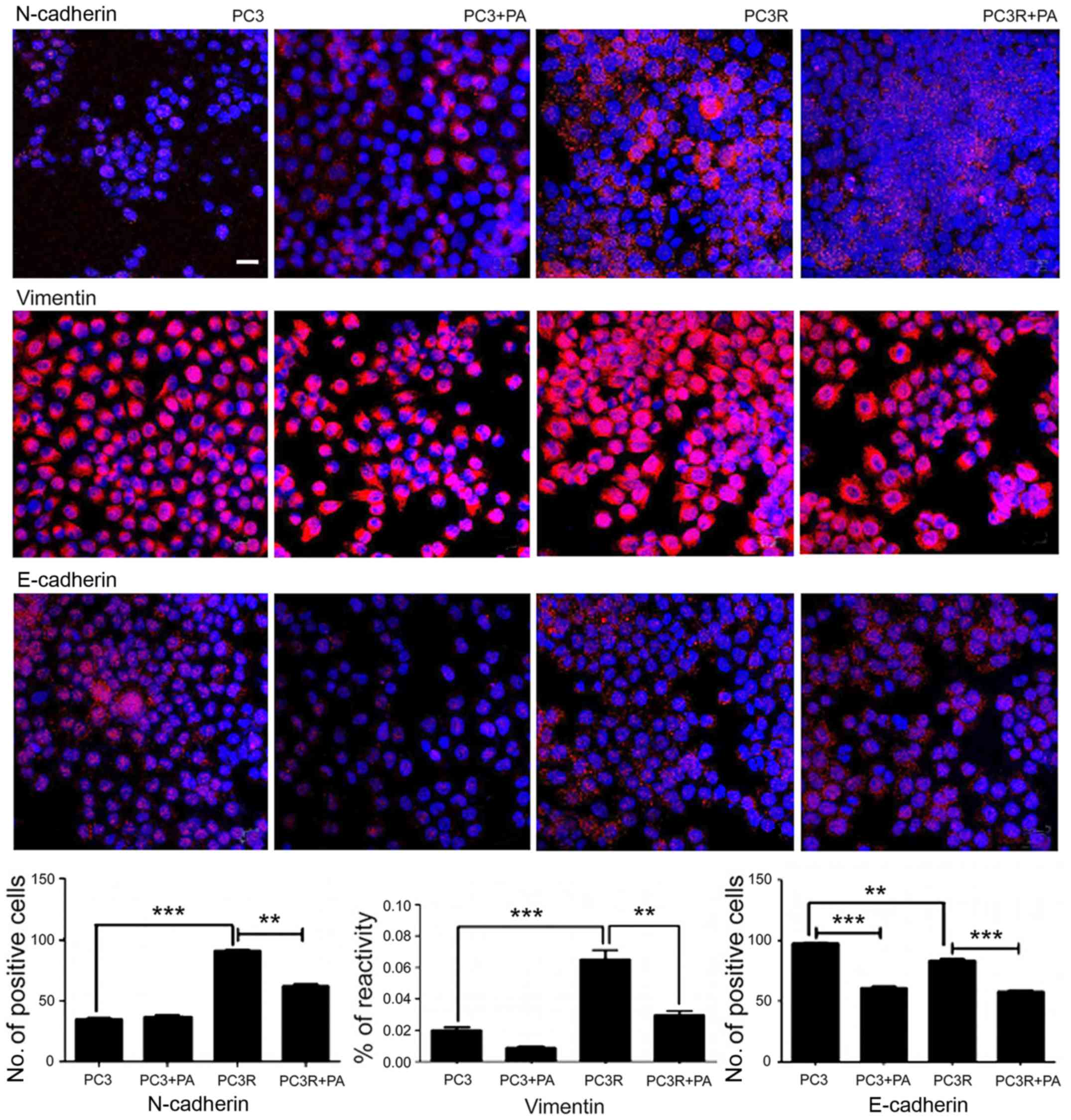
Morphological analyses of the cells showed that DTX
treatment changed the shape of the parental cells to a more
elongated, fibroblastic appearance suggestive of EMT. Indeed, PC3R
cells showed an increased amount of N-cadherin and vimentin
(P<0.001), and a slight decreased amount of E-cadherin
(P<0.01) compared with the PC3 parental cells suggesting an EMT
phenotype in PC3R cells. Since EMT is a process involved in the
progression of PC to CRPC and therefore, a possible target for the
treatment of this tumor, to explore the mechanism of action of PA
on PC, we evaluate the effects of this compound on various markers
of EMT. The treatment of PC3R cells with 5 µg/ml PA for 48 h,
decreased N-cadherin, vimentin, and E-cadherin levels in PC3R cells
compared with non-treated PC3R cells (Fig. 9) indicating that the triterpene
partially reverts the EMT phenotype observed in the resistant cell
line.
Discussion
Prostate cancer (PC) is one of the leading causes of
cancer-related death in the world in the male population (1). The mortality of PC is dependent on
tumor recurrence and its progression to metastatic disease. To
date, there is no effective treatment for metastatic disease which
remains incurable (24).
Resistant PC strains are an important tool for
understanding castration-resistant prostate cancer (CRPC) and for
the search for alternatives to its treatment (22,25,26).
The present study showed that the continuous treatment of PC3 with
docetaxel (DTX) generated a resistant cell lineage (PC3R) as
demonstrated by its lower sensitivity to the drug (Fig. 1). Thus, while 8 nM DTX was able to
reduce the cell viability of the parental line (PC3) by 60%, ~100
nM of the drug was required to attain the same effect on PC3R
cells. These data have been corroborated by the work of other
researches describing PC3 cell lines resistant to DTX (22,26)
and to paclitaxel (25). The
DTX-resistant lineage (PC3R) also exhibited cross-resistance to
doxorubicin and vincristine, characterizing the MDR phenotype of
this lineage.
Transporter proteins of the ABC family are one of
the main mechanisms of MD, and differential expression of these
pumps has been demonstrated in several tumor types including PC. In
fact, an increase in P-gp/ABCB1 and MRP1/ABCC1 expression in PC
samples in comparison with normal prostate tissue has already been
observed (27).
Functional analysis of the major ABC transporters
showed P-gp/ABCB1 activity in the resistant (PC3R) cell line but
not in the parental cell line (PC3) (Fig. 3A and B). Expression of P-gp/ABCB1 in
DTX-resistant PC lines (22) and
correlation of this expression with DTX resistance has been
previously reported (28,29). In the TaxR line, the deletion
(knockout) of P-gp/ABCB1 expression by shRNA reversed the response
to DTX (30), indicating that
increased expression and/or activity of this protein could be
responsible for the increased resistance to DTX noted in the PC3R
cells. In fact, our results demonstrating that treatment with
verapamil (VP), a specific inhibitor of P-gp/ABCB1, increased the
sensitivity to DTX (Fig. 3C),
corroborates this hypothesis.
Different form P-gp/ABCB1, MRP1/ABCC1 transporter
activity was detected in both the resistant and parental PC3 cells
(Fig. 4A and B). The presence of
active MRP1/ABCC1 has also been described in PC cell lines
(31) and increased expression of
this protein was found in doxorubicin-resistant prostate lines
(32). The presence of an active
MRP1 in the studied lineages suggests a possible contribution of
this transporter in the intrinsic resistance of PC. Moreover,
treatment of PC3R with MK57, a specific inhibitor of MRP1/ABCC1
activity, only partially reversed the response to DTX (Fig. 4C) indicating that other mechanisms
are also involved in the intrinsic resistance of these cells to
DTX. Absence of BCRP/ABCG2 activity (data not shown), indicates
that this resistance was not mediated by this transporter.
Several chemotherapeutic drugs have been used for
the treatment of PC. However, in spite of several new therapies,
they generally induce resistance, leading to treatment failure,
disease progression and patient death (12,13).
New approaches able to bypass the resistance mechanisms and to
increase the survival of patients with CRPC are urgently needed in
the clinic. With the objective of searching for new alternatives
for CRPC, we evaluated the effect of the triterpene pomolic acid
(PA) on the PC3 and PC3R cell lines. The antitumor activity of
triterpenes in several cancer lineages, including PC, is well known
(13,15,21,33,34).
Similar to other triterpenes, PA shows potent antitumor activity,
reducing the viability and inducing apoptosis of different tumor
cell lines (17,35,36)
including cancer cells presenting different MDR mechanisms
(18). Our results (Fig. 5) showed that PA is quite effective,
reducing the viability of the studied cell lines to levels below
that obtained by treatment with DTX. The similarity of PA effects
on PC3 and PC3R indicates that the triterpene is able to bypass not
only the resistance mechanisms induced by DTX but also the
intrinsic resistance mechanism present in the lineages. Indeed, PA
did not interfere with P-gp/ABCB1 activity (Fig. 7B) present on PC3R cells, confirming
previous findings in leukemic cell lines (17,18)
that this triterpene is not a substrate for this transporter.
Moreover, in agreement with previous observations of our group
(36), PA negatively modulated the
activity of MRP1/ABCC1 (Fig. 8A and
B), suggesting that this effect contributes to PA cytotoxicity.
However in addition to the activity of MDR proteins, resistance
mechanisms present in CRPC may also be mediated by alterations of
antiapoptotic proteins of the Bcl2 family (37) and innate activation of pathways
involved in the EMT process, such as NF-κB and PI3K/AKT/mTOR among
others (24). Literature data show
that the cytotoxic effect of PA includes modulation of several of
these pathways. Thus, PA is capable of killing Bcl-2 overexpressing
cells (14), a protein whose
inhibition restores the response to DTX (37). PA also inhibits the activation of
NF-κB (38) a factor involved in PC
progression (39), drug resistance
(40) and activation and
maintenance of the AR (41).
Moreover, PA inhibits the activation of PI3K/AKT/mTOR (38) a pathway involved in disease
progression and whose inhibition sensitizes PC cells to the
apoptotic effect induced by chemotherapy (42).
Resistance of CRPC to DTX has also been related to
endogenous cellular mechanisms or mechanisms dependent on the tumor
microenvironment such as the EMT process. Previous studies have
demonstrated that resistant prostate lines, including those
resistant to docetaxel, exhibit an EMT phenotype (43,44).
Analysis of the expression of EMT markers in the cell lines by
fluorescence microscopy revealed that PC3R exhibited an increase in
N-cadherin and vimentin expression and a slight decrease in
E-cadherin when compared to PC3 indicating that PC3R underwent the
EMT process (Fig. 9). PA partially
reverted the increase in the expression of N-cadherin and vimentin
induced by DTX treatment although its effect on E-cadherin was
negligible. These results corroborate literature data showing that
the triterpene ursolic acid blocks EMT by increasing E-cadherin
expression and decreasing vimentin expression in different strains
(45,46).
The results obtained in the present study
demonstrated that PA may be a possible candidate for the
development of novel antitumor agents for future PC chemotherapy.
Indeed, in addition to being a powerful cytotoxic agent, this
triterpene was found to interfere with important mechanisms of
resistance in the treatment of PC such as that mediated by
transporter proteins (P-gp/ABCB1 and MRP1/ABCC1) and its effects on
EMT may be useful in preventing disease progression. Moreover,
literature data have shown that PA also acts on several pathways
involved in DTX resistance and the EMT process. Together, these
data suggest a possible advantage of using PA as a co-adjuvant to
prevent progression of PC and to the treatment of CRCP.
Acknowledgements
Not applicable.
Funding
The present study was supported by funds of the
Conselho de Desnvolvimento Científico e Tecnológico (CNPq) Grants
485193/2012-4 (CMT) and 309056/2014-4 (CRG), Fundação de Amparo a
Pesquisa do Estado do Rio de Janeiro (FAPERJ) Grant
E-26-102.283/2013 (CMT) and Fundação do Câncer 2016. The authors
wish to thank the Conselho de Aperfeiçoamento do Pessoal de Nível
Superior (CAPES) for the graduate fellowship awarded to Carollina
de Araújo Martins.
Availability of data and materials
The datasets generated in the present study are
available from the corresponding author on reasonable request.
Authors' contributions
CRG and CMT conceived and designed the study. CAM
performed the experiments with the collaboration of GGR. CRG, CAM,
GGR and CMT reviewed and edited the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests. The authors alone are responsible for the content and
writing of this article.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yap TA, Smith AD, Ferraldeschi R,
Al-Lazikani B, Workman P and de Bono JS: Drug discovery in advanced
prostate cancer: Translating biology into therapy. Nat Rev Drug
Discov. 15:699–718. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Keyes M, Crook J, Morton G, Vigneault E,
Usmani N and Morris WJ: Treatment options for localized prostate
cancer. Can Fam Physician. 59:1269–1274. 2013.PubMed/NCBI
|
|
4
|
Dai C, Heemers H and Sharifi N: Androgen
signaling in prostate cancer. Cold Spring Harb Perspect Med.
7(pii): a0304522017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sumanasuriya S and de Bono J: Treatment of
advanced prostate cancer-A review of current therapies and future
promise. Cold Spring Harb Perspect Med. 8(pii): a0306352018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al: Docetaxel plus prednisone or mitoxantrone plus prednisone for
advanced prostate cancer. N Engl J Med. 351:1502–2512. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
De Bono JS, Oudard S, Ozguroglu M, Hansen
S, Machiels JP, Kocak I, Gravis G, Bodrogi I, Mackenzie MJ, Shen L,
et al: Prednisone plus cabazitaxel or mitoxantrone for metastatic
castration-resistant prostate cancer progressing after docetaxel
treatment: A randomised open-label trial. Lancet. 376:1147–1154.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seruga B, Ocana A and Tannock IF: Drug
resistance in metastatic castration-resistant prostate cancer. Nat
Rev Clin Oncol. 8:12–23. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakazawa M, Paller C and Kyprianou N:
Mechanisms of therapeutic resistance in prostate cancer. Curr Oncol
Rep. 19:132017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sridhar SS, Freedland SJ, Gleave ME,
Higano C, Mulders P, Parker C, Sartor O and Saad F:
Castration-resistant prostate cancer: From new pathophysiology to
new treatment. Eur Urol. 65:289–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Arsov C, Winter C, Rabenalt R and Albers
P: Current second-line treatment options for patients with
castration resistant prostate cancer (CRPC) resistant to docetaxel.
Urol Oncol. 30:762–771. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chandrasekar T, Yang JC, Gao AC and Evans
CP: Mechanisms of resistance in castration-resistant prostate
cancer (CRPC). Transl Androl Urol. 4:365–380. 2015.PubMed/NCBI
|
|
13
|
Gill BS, Kumar S and Navgeet: Triterpenes
in cancer: Significance and their influence. Mol Biol Rep.
43:881–896. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fernandes J, Weinlich R, Castilho RO,
Amarante-Mendes GP and Gattass CR: Pomolic acid may overcome
multidrug resistance mediated by overexpression of anti-apoptotic
Bcl-2 proteins. Cancer Lett. 245:315–320. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lúcio KA, Rocha Gda G, Monção-Ribeiro LC,
Fernandes J, Takiya CM and Gattass CR: Oleanolic acid initiates
apoptosis in non-small cell lung cancer cell lines and reduces
metastasis of a B16F10 melanoma model in vivo. PLoS One.
6:e285962011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yan XJ, Gong LH, Zheng FY, Cheng KJ, Chen
ZS and Shi Z: Triterpenoids as reversal agents for anticancer drug
resistance treatment. Drug Discov Today. 19:482–488. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fernandes J, Castilho RO, da Costa MR,
Wagner-Souza K, Coelho Kaplan MA and Gattass CR: Pentacyclic
triterpenes from Chrysobalanaceae species: Cytotoxicity on
multidrug resistant and sensitive leukemia cell lines. Cancer Lett.
190:165–169. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vasconcelos FC, Gattass CR, Rumjanek VM
and Maia RC: Pomolic acid-induced apoptosis in cells from patients
with chronic myeloid leukemia exhibiting different drug resistance
profile. Invest New Drugs. 25:525–533. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Song Y, Zhang P, Zhu H, Chen L, Xiao
Y and Xing Y: Oleanolic acid inhibits cell survival and
proliferation of prostatecancer cells in vitro and in vivo through
the PI3K/Akt pathway. Tumor Biol. 37:7599–7613. 2016. View Article : Google Scholar
|
|
20
|
Shin J, Lee HJ, Jung DB, Jung JH, Lee HJ,
Lee EO, Lee SG, Shim BS, Choi SH, Ko SG, et al: Suppression of
STAT3 and HIF-1 alpha mediates anti-angiogenic activity of
betulinic acid in hypoxic PC-3 prostate cancer cells. PLoS One.
6:e214922011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shanmugam MK, Manu KA, Ong TH,
Ramachandran L, Surana R, Bist P, Lim LH, Kumar AP, Hui KM and
Sethi G: Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid
leads to suppression of metastasis in transgenic adenocarcinoma of
mouse prostate model. Int J Cancer. 129:1552–1563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
O'Neill AJ, Prencipe M, Dowling C, Fan Y,
Mulrane L, Gallagher WM, O'Connor D, O'Connor R, Devery A, Corcoran
C, et al: Characterisation and manipulation of docetaxel resistant
prostate cancer cell lines. Mol Cancer. 10:1262011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakazawa M, Paller C and Kyprianou N:
Mechanisms of therapeutic resistance in prostate cancer. Curr Oncol
Rep. 19:132017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kojima K, Fujita Y, Nozawa Y, Deguchi T
and Ito M: MiR-34a attenuates paclitaxel-resistance of
hormone-refractory prostate cancer PC3 cells through direct and
indirect mechanisms. Prostate. 70:1501–1512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee BY, Hochgräfe F, Lin HM, Castillo L,
Wu J, Raftery MJ, Martin Shreeve S, Horvath LG and Daly RJ:
Phosphoproteomic profiling identifies focal adhesion kinase as a
mediator of docetaxel resistance in castrate-resistant prostate
cancer. Mol Cancer Ther. 13:190–201. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Karatas OF, Guzel E, Duz MB, Ittmann M and
Ozen M: The role of ATP-binding cassette transporter genes in the
progression of prostate cancer. Prostate. 76:434–444. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu Y, Liu C, Armstrong C, Lou W, Sandher
A and Gao AC: Anti-androgens inhibit ABCB1 efflux and ATPase
activity and reverse docetaxel resistance in advanced prostate
cancer. Clin Cancer Res. 21:4133–4142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lombard AP, Liu C, Armstrong CM, Cucchiara
V, Gu X, Lou W, Evans CP and Gao AC: ABCB1 mediates
cabazitaxel-docetaxel cross-resistance in advanced prostate cancer.
Mol Cancer Ther. 16:2257–2266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu Y, Liu C, Nadiminty N, Lou W, Tummala
R, Evans CP and Gao AC: Inhibition of ABCB1 expression overcomes
acquired docetaxel resistance in prostate cancer. Mol Cancer Ther.
12:1829–1836. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sánchez C, Mercado A, Contreras HR,
Mendoza P, Cabezas J, Acevedo C, Huidobro C and Castellón EA:
Chemotherapy sensitivity recovery of prostate cancer cells by
functional inhibition and knock down of multidrug resistance
proteins. Prostate. 71:1810–1817. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zalcberg J, Hu XF, Slater A, Parisot J,
El-Osta S, Kantharidis P, Chou ST and Parkin JD: MRP1 not MDR1 gene
expression is the predominant mechanism of acquired multidrug
resistance in two prostate carcinoma cell lines. Prostate Cancer
Prostatic Dis. 3:66–75. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rocha Gda G, Simões M, Oliveira RR, Kaplan
MA and Gattass CR: 3β-acetyl tormentic acid induces apoptosis of
resistant leukemia cells independently of P-gp/ABCB1 activity or
expression. Invest New Drugs. 30:105–113. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meng Y, Lin ZM, Ge N, Zhang DL, Huang J
and Kong F: Ursolic acid induces apoptosis of prostate cancer cells
via the PI3K/Akt/mTOR Pathway. Am J Chin Med. 43:1471–1486. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yoo KH, Park JH, Lee DK, Fu YY, Baek NI
and Chung IS: Pomolic acid induces apoptosis in SK-OV-3 human
ovarian adenocarcinoma cells through the mitochondrial-mediated
intrinsic and death receptor-induced extrinsic pathways. Oncol
Lett. 5:386–390. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guimarães LPTP, Rocha GDG, Queiroz RM,
Martins CA, Takiya CM and Gattass CR: Pomolic acid induces
apoptosis and inhibits multidrug resistance protein MRP1 and
migration in glioblastoma cells. Oncol Rep. 38:2525–2534. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tamaki H, Harashima N, Hiraki M, Arichi N,
Nishimura N, Shiina H, Naora K and Harada M: Bcl-2 family
inhibition sensitizes human prostate cancer cells to docetaxel and
promotes unexpected apoptosis under caspase-9 inhibition.
Oncotarget. 5:11399–11412. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Park JH, Yoon J and Park B: Pomolic acid
suppresses HIF1α/VEGF-mediated angiogenesis bytargeting p38-MAPK
and mTOR signaling cascades. Phytomedicine. 23:1716–1726. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
McCall P, Bennett L, Ahmad I, Mackenzie
LM, Forbes IW, Leung HY, Sansom OJ, Orange C, Seywright M,
Underwood MA, et al: NFκB signalling is upregulated in a subset of
castrate-resistant prostate cancer patients and correlates with
disease progression. Br J Cancer. 107:1554–1563. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Codony-Servat J, Marín-Aguilera M, Visa L,
García-Albéniz X, Pineda E, Fernández PL, Filella X, Gascón P and
Mellado B: Nuclear factor-kappa B and interleukin-6 related
docetaxel resistance in castration-resistant prostate cancer.
Prostate. 73:512–521. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang L, Altuwaijri S, Deng F, Chen L, Lal
P, Bhanot UK, Korets R, Wenske S, Lilja HG, Chang C, et al:
NF-kappaB regulates androgen receptor expression and prostate
cancer growth. Am J Pathol. 175:489–499. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chang L, Graham PH, Hao J, Ni J, Bucci J,
Cozzi PJ, Kearsley JH and Li Y: PI3K/Akt/mTOR pathway inhibitors
enhance radiosensitivity in radioresistant prostate cancer cells
through inducing apoptosis, reducing autophagy, suppressing NHEJ
and HR repair pathways. Cell Death Dis. 5:e14372014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Marín-Aguilera M, Codony-Servat J, Reig Ò,
Lozano JJ, Fernández PL, Pereira MV, Jiménez N, Donovan M, Puig P,
Mengual L, et al: Epithelial-to-mesenchymal transition mediates
docetaxel resistance and high risk of relapse in prostate cancer.
Mol Cancer Ther. 13:1270–1284. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Puhr M, Hoefer J, Schäfer G, Erb HH, Oh
SJ, Klocker H, Heidegger I, Neuwirt H and Culig Z:
Epithelial-to-mesenchymal transition leads to docetaxel resistance
in prostate cancer and is mediated by reduced expression of
miR-200c and miR-205. Am J Pathol. 181:2188–2201. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu K, Guo L, Miao L, Bao W, Yang J, Li X,
Xi T and Zhao W: Ursolic acid inhibits epithelial-mesenchymal
transition by suppressing the expression of astrocyte-elevated
gene-1 in human nonsmall cell lung cancer A549 cells. Anticancer
Drugs. 24:494–503. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sohn EJ, Won G, Lee J, Yoon SW, Lee I, Kim
HJ and Kim SH: Blockage of epithelial to mesenchymal transition and
upregulation of let 7b are critically involved in ursolic acid
induced apoptosis in malignant mesothelioma cell. Int J Biol Sci.
12:1279–1288. 2016. View Article : Google Scholar : PubMed/NCBI
|