Introduction
Cervical cancer is one of the most common malignant
tumors among women worldwide, with more than 85% of cervical
cancer-related deaths occurring in developing countries (1). Although the development of diagnosis
techniques has improved the detection of cervical cancer and the
use of vaccines can effectively prevent the disease, the overall
survival rate of cervical cancer patients at five years remains
only ~50% since most patients are diagnosed when the cancer is at
an advanced stage (2). It is,
therefore, crucially important to develop effective novel
therapeutic strategies to improve the survival rate of
patients.
Keratins, also known as cytokeratins, are the
intermediate filament (IF)-forming proteins of epithelial cells.
The primary role of keratins is to protect epithelial cells from
both mechanical and non-mechanical stressors (3). Keratins have also been reported to
have many roles in cancer, including as diagnostic markers
(4–7) and prognostic markers in epithelial
tumors (8–10), as well as roles in tumorigenesis
(11,12) and drug responsiveness (13–16).
Keratins 8, 18 and 19 (CK8, CK18 and CK19), which
are the most abundant keratins in simple epithelial cells, are
extensively used as the diagnostic markers (17,18).
CK18, also known as KRT18, and its caspase-cleaved fragment can be
released into the circulation and are indicative of epithelial cell
necrosis and apoptosis, respectively. Caspase-cleaved CK18 (M30)
and full length CK18 (M65) are assessed using tissue polypeptide
antigen (TPA) and tissue polypeptide specific antigen (TPS)
(19). The M30 and M65 assays may
provide important prognostic and predictive biomarkers in many
malignancies (14,18,20–23).
Notably, several studies have revealed that CK18 is
not only a biomarker but also a regulator in many diseases,
including cancer (24,25). CK18 knockdown can decrease
cell migration and increase chemosensitivity in non-small cell lung
cancer (15), decrease cell
migration in renal carcinoma (RCC5) cells (26), and increase cytokine-induced
apoptosis in HeLa cells (27),
suggesting that CK18 gene positively regulates tumorigenesis.
Conversely, CK18 has been reported to suppress tumor aggressiveness
and paclitaxel-resistance in paclitaxel-resistant prostate cancer
(DU145-TxR) cells (16), as well as
to induce cell adhesion and regression of malignancy in breast
cancer (28). However, it remains
unclear how CK18 exerts these biological functions.
Notably, CK18 has been reported to physically
interact with LRP16, also known as O-Acetyl-ADP-Ribose Deacetylase
MACROD1, thereby sequestering LRP16 in the cytoplasm and thus
inhibiting the proliferation of ERα-positive breast tumor cells
(29). It could be possible that
CK18 regulates gene expression via protein-protein interaction.
CK18 has also been revealed to be associated with mRNAs and could,
therefore, be an RNA binding protein (RBP) (30). RBPs are known to coordinate RNA
processing and post-transcriptional gene regulation (31). It is, however, unclear whether CK18
is able to affect post-transcriptional gene regulation in the
context of exerting biological functions.
In the present study, we firstly aimed to elucidate
the function of CK18 in HeLa cells, which are derived from cervical
cancer cells. It was revealed that knockdown of CK18 led to
significantly reduced apoptosis. RNA-seq analysis of the effect of
CK18 on the transcription and alternative splicing of the HeLa
transcriptome revealed that CK18 regulated the expression and
alternative splicing of a number of genes involved in apoptosis,
including FAS, FADD and CASP8. Additionally, CK18 was
revealed to regulate the transcription and alternative splicing of
many genes enriched in immunity and cancer-related pathways. Our
study revealed a novel apoptotic function of CK18, which was linked
to a feedforward regulation of apoptosis genes FAS and
CASP8, and to CK18-regulated transcriptomes favoring
apoptosis at both the transcription and alternative splicing
levels. These findings elucidated the dysregulated expression of
CK18 in cancers and supported an important role for CK18 in
tumorigenesis.
Materials and methods
Cell culture and transfections
HeLa (immortalized human cervical cancer) cells were
purchased from the Institute of Biochemistry and Cell Biology,
Chinese Academy of Sciences, HepG2 (immortalized human liver
cancer) cells were purchased from the American Type Culture
Collection (ATCC; Manassas, VA, USA). The cell lines were
identified by short terminal repeat (STR) genotyping, which
revealed a correspondence of >80% of the markers tested
(32). Cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine
serum (FBS).
To knockdown CK18, different shRNAs against
human CK18 (shRNA1: 5′-GATGACACCAATATCACACGA-3′; shRNA2:
5′-CTTCATGAAGAAGAACCACGA-3′; shRNA3: 5′-CCTGCTGAACATCAAGGTCAA-3′)
were designed, and scrambled shRNA (Scr-shRNA) that targeted a
non-specific sequence (5′-ACTGGACCAGGCAGCAGCGTCAGAAGACT-3′) was
used as the control. These shRNAs were transfected into HeLa cells
using Lipofectamine 2000 transfection reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. Transfected cells were harvested after 48
h for analysis.
Assesment of knockdown by shRNA
Total RNA was isolated from cells using TRIzol
reagent (Ambion; Thermo Fisher Scientific, Inc) and cDNA synthesis
was carried out using standard procedures. qPCR was performed on
the Bio-Rad S1000 Thermal Cycler, using Bestar SYBR-Green RT-PCR
Master Mix (DBI Bioscience, Shanghai, China). The PCR conditions
are consisted of denaturing at 95°C for 10 min, 40 cycles of
denaturing at 95°C for 15 sec, annealing and extension at 60°C for
1 min. The primers of CK18 used for quantitative real-time PCR
(qPCR) were: Forward, AAAGGCCTACAAGCCCAGAT and reverse,
CACTGTGGTGCTCTCCTCAA. Gene expression levels were calculated using
the 2−ΔΔCq method (33)
and CT values were normalized using glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) as an internal standard. The primers of GAPDH
were: Forward, GGTCGGAGTCAACGGATTTG and reverse,
GGAAGATGGTGATGGGATTTC.
MTT assay
An MTT assay was used to assess cell proliferation.
Briefly, HeLa cells were cultured in 96-well plates and transfected
with the vector using Lipofectamine 2000, according to the
manufacturer's protocol. The cells were then incubated at 37°C for
48 h. Subsequently, MTT solution (5 mg/ml, 0.025 ml) was added to
each well, and the cells were incubated for another 4 h. After
centrifugation at 4,000 rpm for 15 min, the supernatant was removed
from each well. The colored formazan crystals produced from MTT in
each well were dissolved in DMSO (0.15 ml) and the optical density
(OD) values were measured at 490 nm.
Flow cytometric analysis of cell
apoptosis
HeLa cells (5×104) were seeded in 24-well
culture plates. Once the cells reached 70% confluence, the cells
were were transfected with the vector using Lipofectamine 2000,
according to the manufacturer's protocol. The cells were then
incubated at 37°C for 48 h and viable cells were harvested and
washed twice with phosphate-buffered saline (PBS). Viable cells
were double-stained with fluorescein isothiocyanate
(FITC)-conjugated Annexin V and 7-amino actinomycin D (7-AAD) (4A
Biotech Co., Ltd. Beijing, China).
The percentage of cell apoptosis was defined as the
sum of the right lower quadrant and upper quadrant.
Library preparation and
sequencing
The RNA was treated with RQ1 DNase (Promega Corp.,
Madison, WI, USA) to remove DNA. The quality and quantity of the
purified RNA were assessed by measuring the absorbance at 260
nm/280 nm (A260/A280) using SmartSpec Plus spectrophotometer
(BioRad Laboratories, Inc., Hercules, CA, USA). The integrity of
each RNA sample was further verified by 1.5% agarose gel
electrophoresis.
For each sample, a total of 1 µg RNA was used for
RNA-seq library preparation. PolyA mRNAs were purified and
concentrated with oligo (dT)-conjugated magnetic beads (Invitrogen;
Thermo Fisher Scientific, Inc.) before directional RNA-seq library
preparation. The fragmented RNAs were end repaired and an adaptor
sequence was ligated at the 5′end. Reverse transcription was then
performed using an RT primer harboring a 3′adaptor sequence and
randomized hexamer. The cDNAs were purified, amplified, and stored
at −80°C until they were used for sequencing.
For high-throughput sequencing, the libraries were
prepared following the manufacturer's instructions. An Illumina
HiSeq 4000 sequencing system (ABLife, Inc., Wuhan, China) was used
to collect data from 151-bp pair-end sequencing.
Clean and alignment of RNA-Seq raw
data
Raw reads were filtered to remove the adaptors,
PolyN reads, and low-quality bases using FASTX-Toolkit (version
0.0.13). Short reads, less than 16 nt, were also discarded. Clean
reads were then aligned to the GRCh38 genome using TopHat2 software
(34), with four mismatches.
Uniquely mapped reads were used to calculate the reads number and
FPKM value (fragments per kilobase of transcript per million
fragments mapped) for each gene.
Analysis of differentially expressed
genes (DEGs)
EdgeR software (35)
was used to measure FPKM values and to analyze the differential
expression of genes using RNA-Seq data, in order to identify DEGs.
To determine whether a gene was differentially expressed, the
results were analyzed based on the fold change (fold change ≥2 or
≤0.5) and a false discovery rate (FDR<0.05).
We also analyzed the two replicates separately
(named as simple pair). In detail, the DEGs were identified between
shCK18-1st vs. Ctrl-1st and shCK18-2nd vs. Ctrl-2nd, respectively.
We then obtained the overlapping genes from the DEGs of each
replicate. The upregulated and downregulated DEGs shared by these
two simple pairs were then overlapped with those identified by
edgeR as biological replicates.
Gene Ontology (GO) and enriched KEGG pathway
analyses were carried out using the KOBAS 2.0 server (36) to predict the gene function and
calculate the frequency distribution of functional categories. The
hypergeometric test and Benjamini-Hochberg FDR controlling
procedure were used to define the enrichment of each pathway
(corrected P-value <0.05).
Alternative splicing analysis
The alternative splicing events (ASEs) and regulated
alternative splicing events (RASEs) between the samples were
defined and quantified by using the ABLas pipeline, as previously
described (37). In brief, eight
types of ASEs were identified, based on the splice junction reads.
The eight possible types of ASE included Cassette exon
(CassetteExon), Exon skipping (ES), Mutual exclusive exon skipping
(MXE), A5SS, A3SS, the MXE combined with alternative 5′promoter
(5pMXE) combined with alternative polyadenylation site (3pMXE) and
intron retention.
Having identified the ASEs in each RNA-seq sample,
Fisher's exact test was selected to determine statistical
significance, using the alternative reads and model reads of the
samples as input data. The altered ratio of alternatively spliced
reads and constitutively spliced reads between compared samples was
defined as the RASE ratio. The P-value <0.05 and RASE ratio
>0.2 were set as the thresholds for detection of RASEs.
Real time qPCR validation of DEGs and
ASEs
To determine the validity of the RNA-seq data, qPCR
was performed for selected DEGs and normalized with the
housekeeping gene GAPDH. Primers are designed in exon regions, and
sequences are presented in Table I.
The same RNA samples for RNA-seq and RNA samples isolated from
CK18 knockdown in HepG2 (using the same shRNAs) were used
for qPCR. The PCR conditions consisted of denaturing at 95°C for 10
min, 40 cycles of denaturing at 95°C for 15 sec, and annealing and
extension at 60°C for 1 min. PCR amplifications were performed in
triplicate for each sample.
 | Table I.Primer sequences used in q-PCR
experiments. |
Table I.
Primer sequences used in q-PCR
experiments.
| DEGsa | Forward | Reverse |
|---|
| FAS |
AAGCGGTTTACGAGTGACTTGG |
AGCATGGTTGTTGAGCAATCCT |
| FADD |
TCTACCTCCGAAGCGTCCTGAT |
AGGTGGTCTGTGGCTCACTCA |
| CD79B |
GGGCTGGAGACAAATGGCAG |
TGAAGTGGTCTGTAGGTGAGCA |
| CXCL2 |
CTTGGATTCCTCAGCCTCTAT |
GGTTTGCAGATATTCTCTAGTC |
|
|
RASGsb | Model
forward | AS
forward | Model/AS
reverse |
|
| CTNNB1 |
CATCCTTTAGCTGTATTGTC |
TTTATACAGCCTGTATTGTC |
AACAAGCAAGGCTAGGGTTTGA |
| FAS |
AAAGAGGAAGGATCCAGATC |
AAAGAGGAAGTGAAGAGAAA |
AGTTGGAGATTCATGAGAACCT |
Concurrently, a qPCR assay was used to analyze ASEs.
The primers for detecting the pre-mRNA splicing are presented in
Table I. To detect one of the
alternative isoforms, one primer was designed in the alternative
exon and an opposing primer was designed in a constitutive exon.
Other alternative isoforms were detected using a boundary-spanning
primer for the sequence encompassing the exon-exon junction, with
the opposing primer in a constitutive exon.
Western blot analysis
CK18 knockdown and control Hela cells were lysed in
RIPA buffer containing 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1.0%
deoxycholate, 1% Triton X-100, 1 mM EDTA and 0.1% SDS. Following
centrifugation of the homogenate (20,000 × g, 15 min) the
supernatants were used for western blotting. Protein concentrations
were measured using the BCA protein assay (Beyotime Institute of
Biotechnology, Nanjing, China) with bovine serum albumin (BSA) as a
standard. Equal amounts (20 µg/lane) of protein samples were loaded
on 12% SDS-PAGE gel for separation and then transferred to a
polyvinylidene difluoride (PVDF) membrane. The membrane was
incubated with blocking solution (1X TBS; 0.05% Tween-20; 5%
non-fat milk) at room temperature for 1 h and incubated overnight
with primary antibodies raised against CK18 (dilution 1:1,000; cat.
no. A0389), CASP8 (dilution 1:1,000; cat. no. A0215) and GAPDH
(dilution 1:1,000; cat. no. AC027) (all from ABclonal, Wuhan,
China). Immunoreactive proteins were detected using an ECL
chemiluminescence system (Clinx Science Instruments Co., Ltd.,
Shanghai, China) with default settings and GAPDH as the normalized
control.
Accession number
RNA-seq data presented in this study have been
deposited in the Gene Expression Omnibus of NCBI and are accessible
through GEO series accession no. GSE119255.
Statistical analysis
Experimental data are presented as the mean ±
standard deviation of at least three experiments. Statistical
analyses were performed with SPSS software (version 17.0; SPSS,
Inc., Chicago, IL, USA). Significance of differences was evaluated
with Student's t-test when only two groups were compared. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effect of CK18 knockdown on
proliferation and apoptosis of HeLa cells
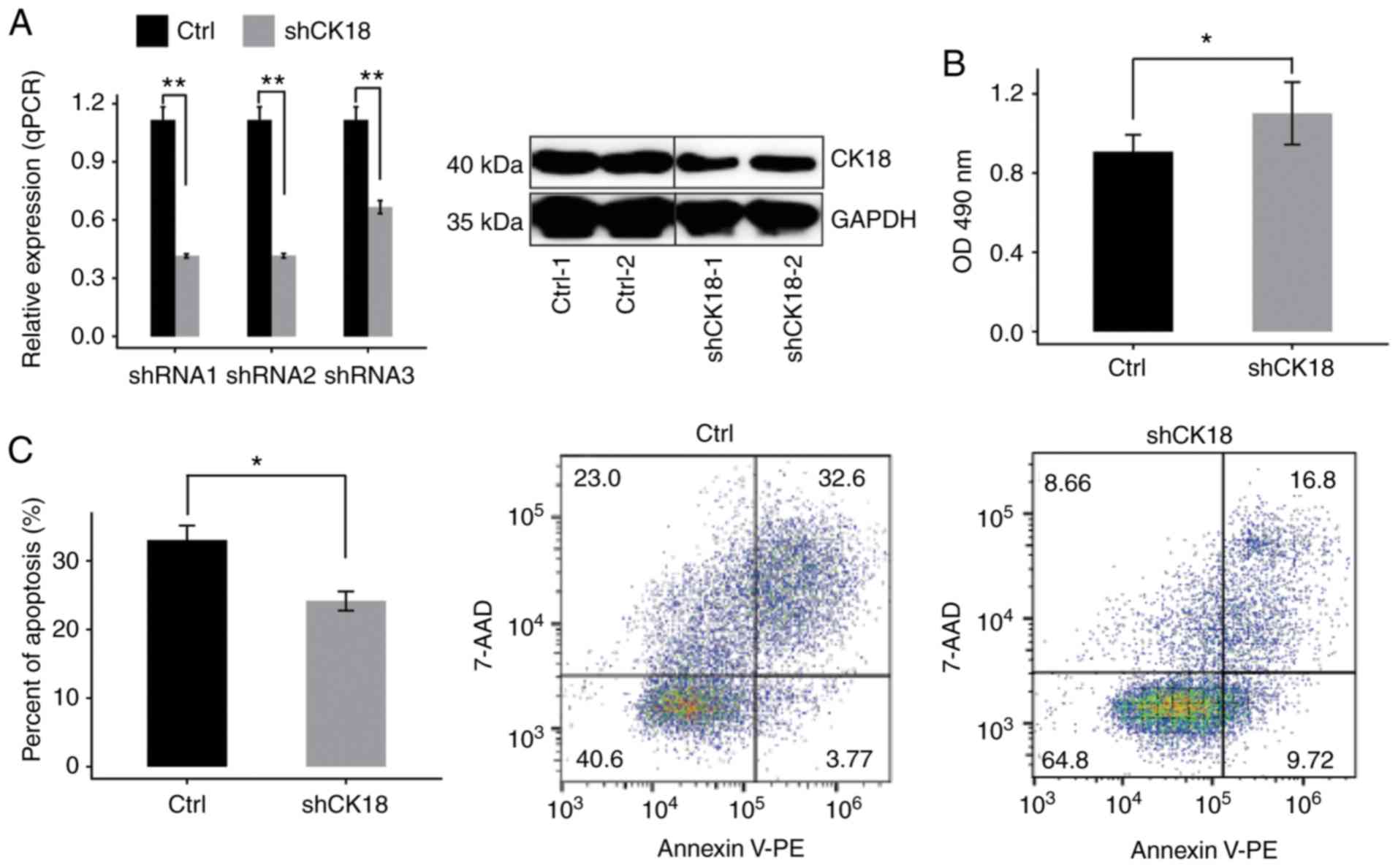
The expression of CK18 was examined in HeLa
cells transduced with different shRNAs (shRNA1, shRNA2 and shRNA3)
or an empty vector by RT-qPCR. Compared with cells treated with the
empty vector mRNA expression was effectively reduced in cells
treated with CK18-shRNAs. The effective shRNA (shRNA1) also
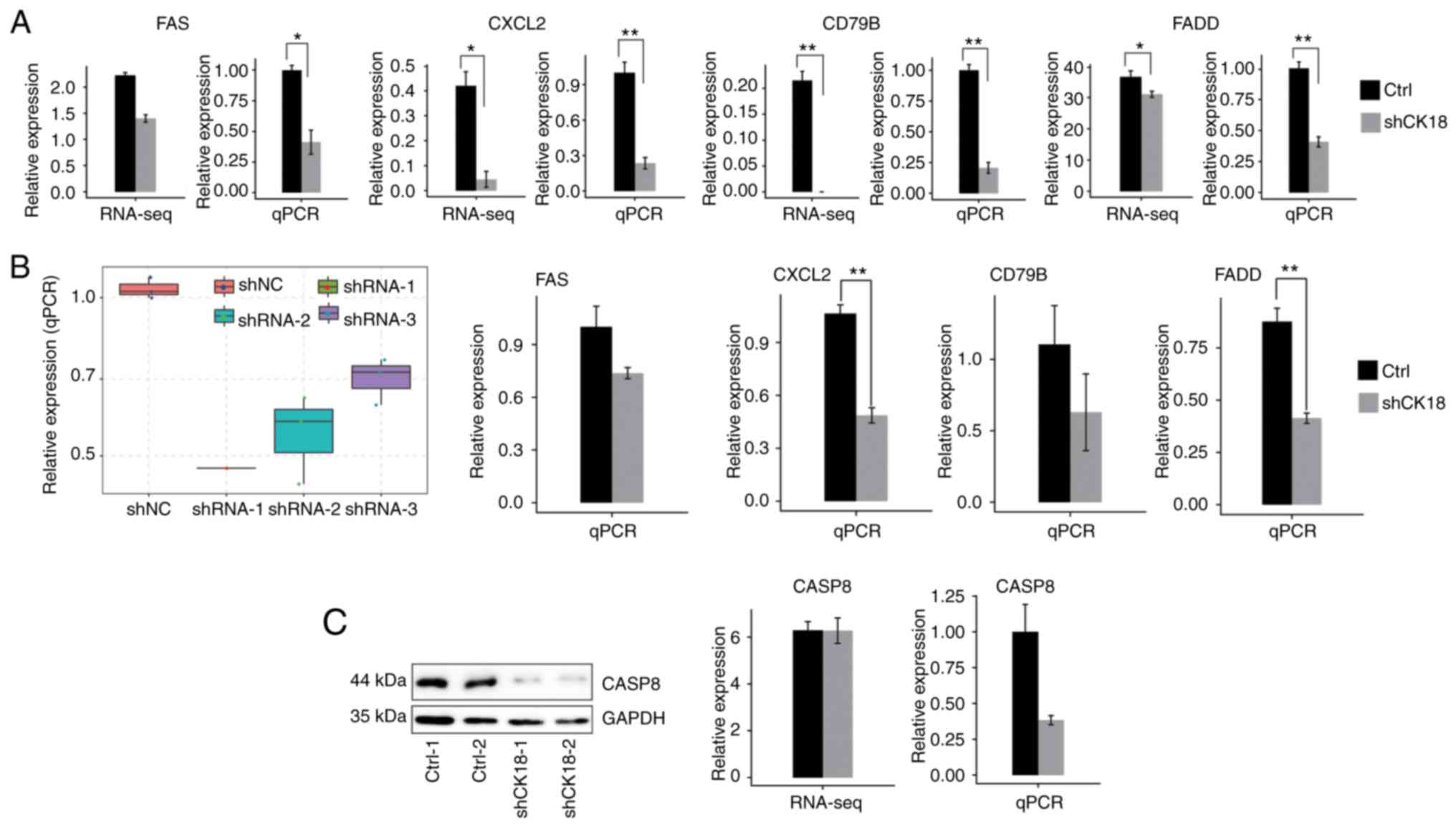
led to a significant reduction in the protein level (Fig. 1A). Therefore, CK18-shRNA1 was
used in subsequent experiments in order to knockdown CK18
expression in HeLa cells. Knockdown of CK18 (CK18 KD)
led to a significant increase in cell proliferation and a
significant decrease in cell apoptosis (Fig. 1B and C).
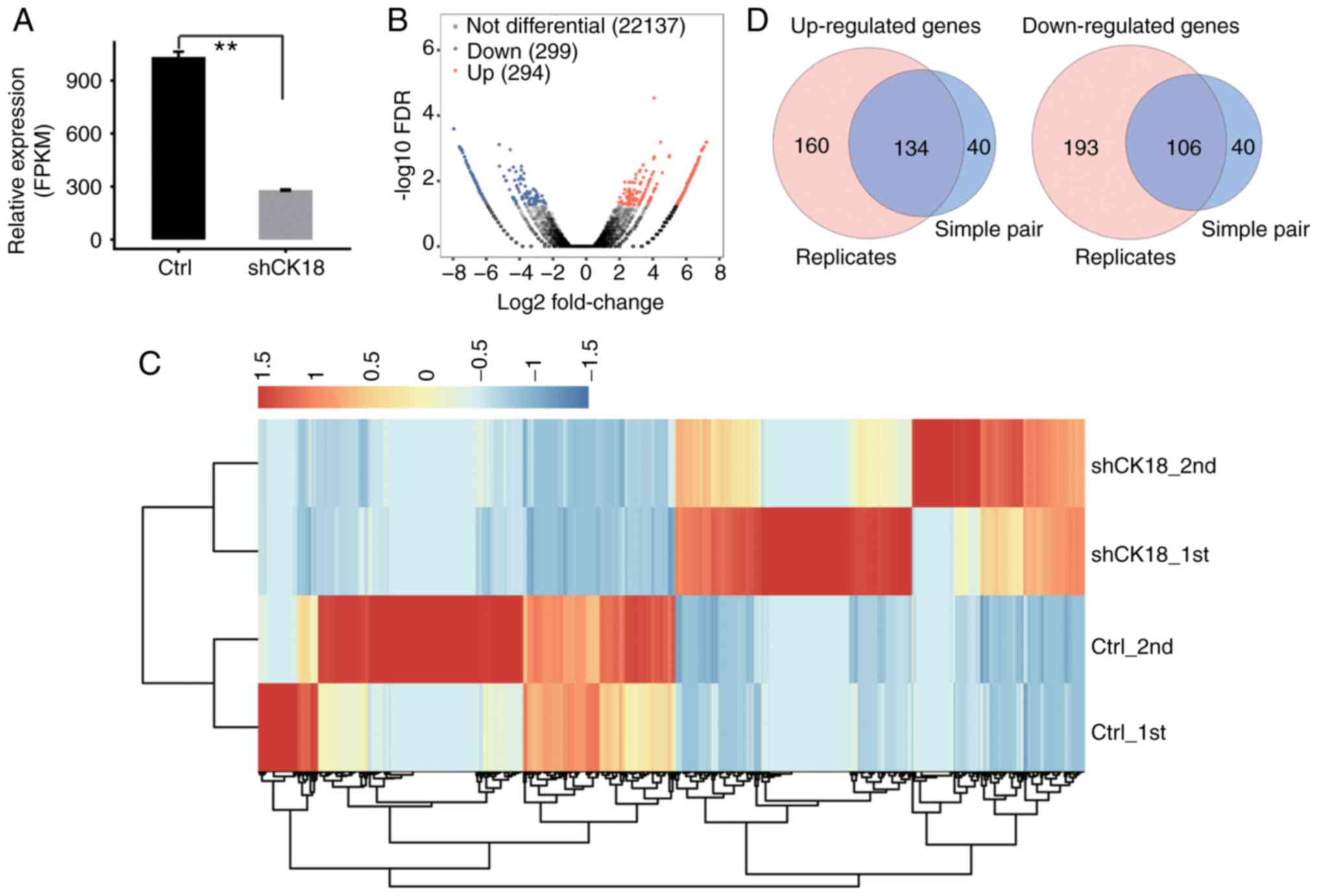
RNA-Seq and DEG analysis
CK18 KD and control cells were used to
construct cDNA libraries for sequencing on an Illumina HiSeq 4000
platform. Two biological replicates were used and a total of
76.7±4.9M 150 nucleotide paired-end raw reads per sample were
obtained. The raw reads were filtered by removing low-quality reads
and reads containing N and adaptor sequences, leaving 73.5±4.7M
clean reads for downstream bioinformatics analysis. The clean reads
were then mapped onto the human GRCh38 genome using TopHat2:
77.99-81.98% were aligned and ~86.77-95.05% were uniquely mapped
(Table II). Quantification of
genes and transcripts was reassessed using Cufflinks (38), to compare gene expression patterns
across individuals. FPKM values were then calculated. There were
22,677 genes expressed in the RNA-seq data (available upon
request). Effective KD of CK18 was further confirmed by in
parallel RNA-seq analysis (Fig.
2A).
| Table II.Summary of RNA-seq reads used in the
analysis. |
Table II.
Summary of RNA-seq reads used in the
analysis.
| Sample | shCK18_1st | ShCK18_2nd | shCtrl_1st | shCtrl_2nd |
|
|---|
| Raw reads | 75207886 | 83976994 | 73556828 | 73880306 |
76655503±4932995a |
| Clean reads | 72139395 | 80470749 | 70542901 | 70836199 |
73497311±4700455 |
| Paired-end
reads | 68071008 | 68521042 | 69757064 | 77558436 |
70976888±4445225 |
| Total mapped | 55639546 | 56175922 | 54402225 | 62167878 |
57096393±3461607 |
|
|
(81.74%b) | (81.98%) | (77.99%) | (80.16%) |
|
| Total uniquely
mapped | 52472346 | 53397850 | 51511527 | 53943874 |
52831399±1069182 |
|
|
(94.31%c) | (95.05%) | (94.69%) | (86.77%) |
|
| Splice reads | 28437401 | 30330300 | 29524140 | 29318683 |
|
|
| (54.2%d) | (56.8%) | (57.32%) | (54.35%) |
|
Using criteria of an absolute fold change ≥2 and FDR
<0.05 with the edge R package (35), 294 upregulated and 299 downregulated
genes related with CK18 KD (data not shown) were identified.
A volcano plot was constructed to display the significantly
expressed genes that were associated with CK18 KD (Fig. 2B). The heatmap demonstrated
distinguishable transcription profiles between CK18 KD and
the control groups (Fig. 2C).
The two replicate RNA-seq datasets were divided into
two groups (case 1 with control 1; case 2 with control 2). Using
the criteria of an absolute fold change ≥2 with the EdgeR software
package, 174 co-upregulated and 146 co-downregulated genes
overlapping in two groups associated with CK18 KD were
identified. A Venn-diagram demonstrated the upregulated and
downregulated genes between the two different analytical strategies
(Fig. 2D).
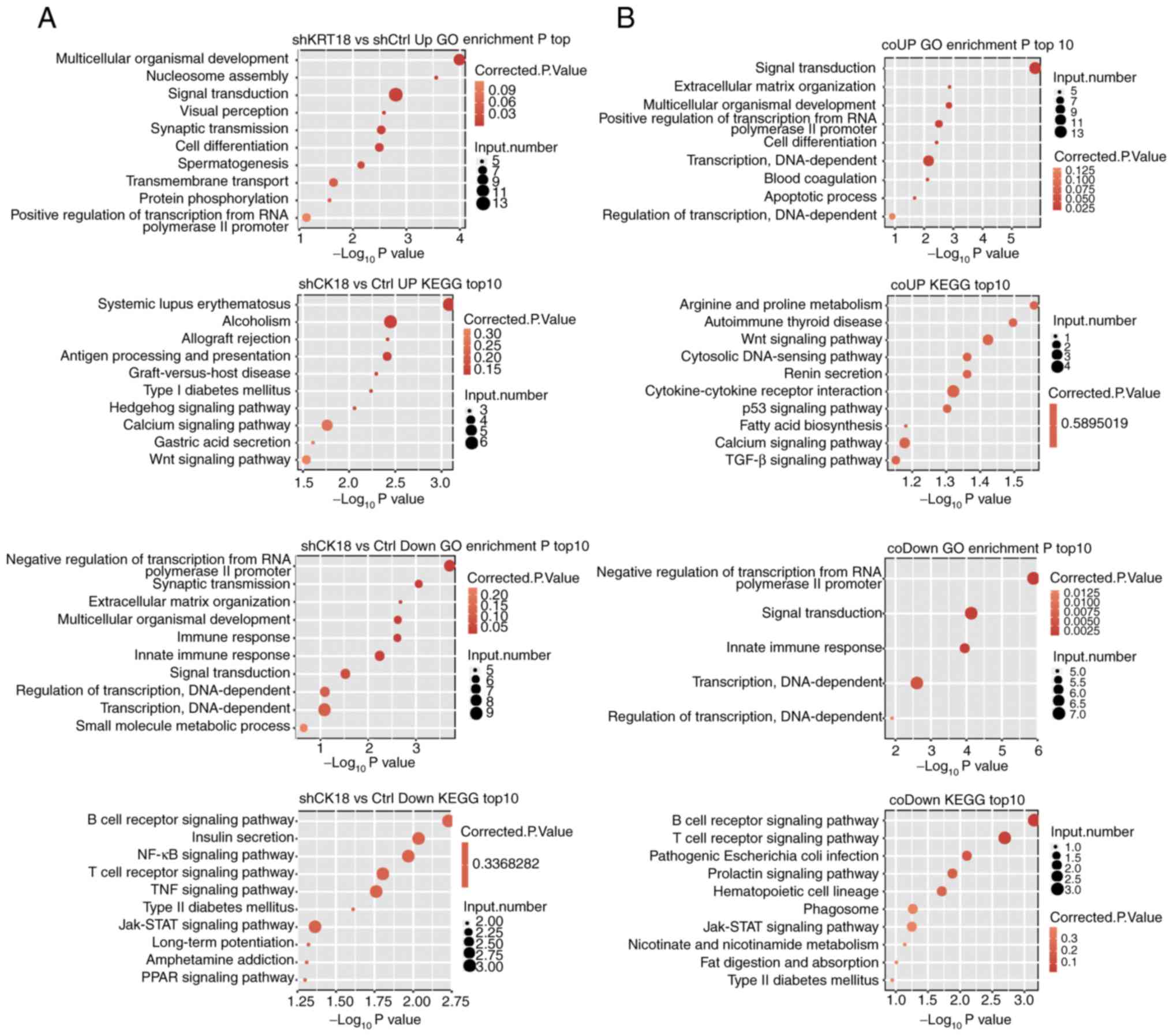
Functional analysis of CK18-regulated
genes
GO enrichment analysis was performed to further
explore the biological function of DEGs with two different
analytical strategies. All three ontologies of GO analysis,
molecular function, cellular component and biological process, were
obtained. The top biological process terms of the GO analysis that
involved upregulation or downregulation of genes following
CK18 KD are presented in Fig.
3A (repetition) and B (simple pair). Most of the terms
overlapped; the upregulated genes were mainly associated with
processes involved in multicellular organismal development, signal
transduction, cell differentiation and the apoptosis process. The
downregulated genes, conversely, were mainly associated with
negative regulation of transcription from RNA polymerase II
promoter, innate immune response, signal transduction, and
regulation of transcription, DNA-dependent. Based on the KEGG
analysis, the top ten pathways involved in up- and downregulated
DEGs were also presented in Fig. 3.
Coinciding with the GO analyses, many pathways were associated with
the immune response and apoptosis. The B-cell receptor signaling
pathway, the T-cell receptor signaling pathway and the Jak-STAT
signaling pathway were all significantly enriched in downregulated
gene sets.
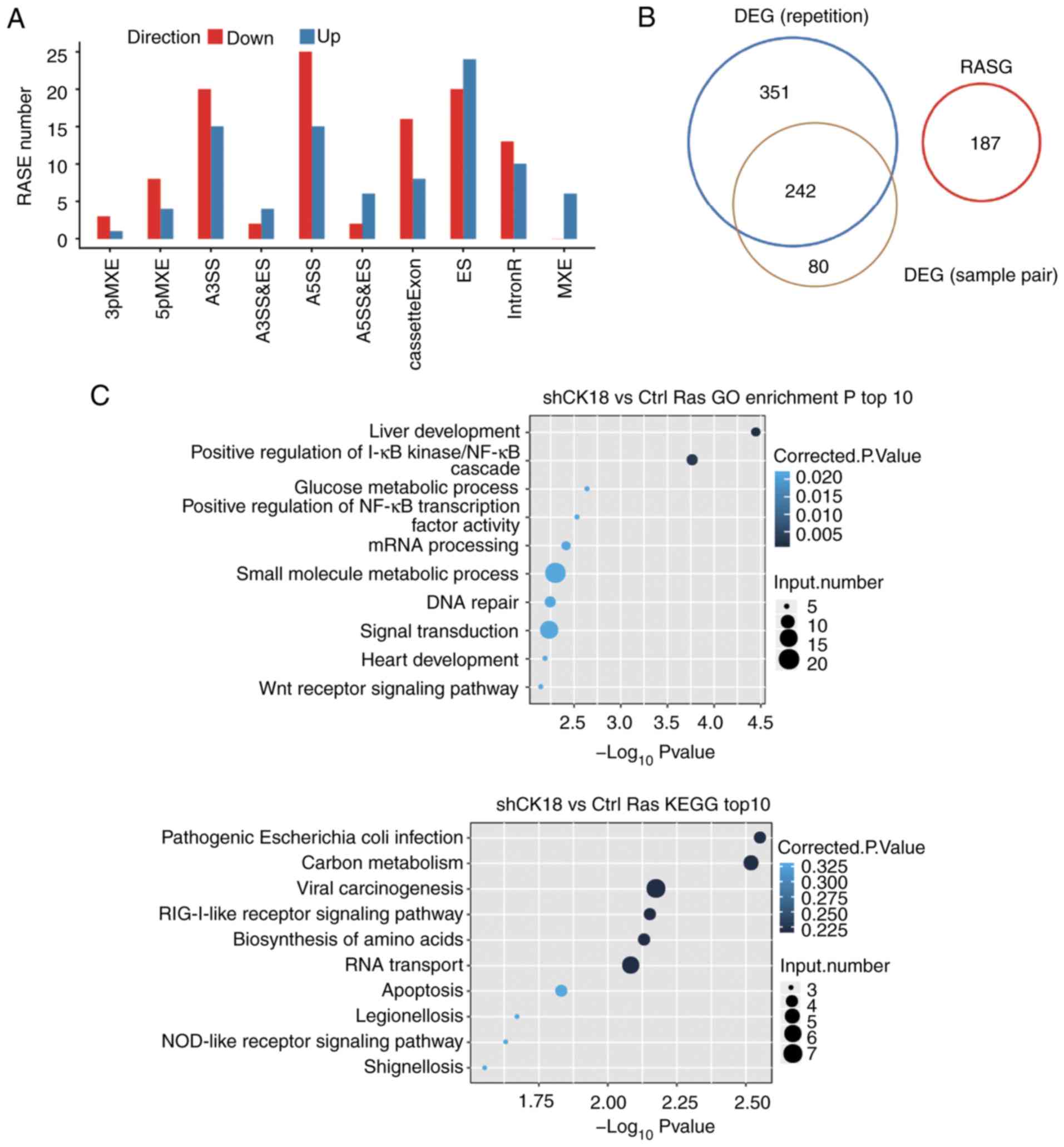
Analysis of CK18-regulated ASEs
Regulation of alternative splicing (AS) of CK18 in
the transcriptome sequencing data was also explored. Between 54.2
and 57.32% of the uniquely mapped reads from CK18 KD and
control samples were junction reads (Table II). A total of 58.9% of annotated
exons (216,356 out of 367,321) were detected. For splicing
junctions, 147,540 annotated and 86,961 novel junctions were
detected using TopHat2.
ASEs were analyzed using ABLas software to
investigate global changes in the occurrence of alternative
splicing. We detected 15,786 known ASEs in the model gene that we
designated in the reference genome, along with 40,946 novel ASEs,
excluding intron retention (Table
III).
| Table III.Known or novel splicing events
detected from ABLas. |
Table III.
Known or novel splicing events
detected from ABLas.
| Known splicing
events |
|---|
|
|---|
| Sample | 3pMXE | 5pMXE | A3SS | A3SS&ES | A5SS | A5SS&ES | ES | IntronR | MXE | Cassette exon | Totalb | Detected
junctionc |
|---|
| shCtrl_1st | 274 | 562 | 2614 | 296 | 3177 | 385 | 2058 | 1013 | 233 | 1463 | 12075 | 134568 |
| shCtrl_2nd | 181 | 383 | 1729 | 197 | 2103 | 277 | 1359 | 953 | 139 | 923 | 8244 | 114474 |
| shCK18_1st | 189 | 399 | 1838 | 202 | 2222 | 307 | 1460 | 959 | 135 | 1000 | 8711 | 119159 |
| shCK18_2nd | 224 | 451 | 2319 | 255 | 2725 | 371 | 1759 | 1009 | 194 | 1199 | 10506 | 128578 |
| All
samplesa | 362 | 733 | 3455 | 384 | 4118 | 515 | 2573 | 1443 | 322 | 1881 | 15786 | 147540 |
|
| Novel splicing
events |
|
| Sample | 3pMXE | 5pMXE | A3SS |
A3SS&ES | A5SS |
A5SS&ES | ES | IntronR | MXE | Cassette
exon |
Totalb | Detected
junctiond |
|
| shCtrl_1st | 612 | 1654 | 3795 | 485 | 5032 | 574 | 1872 | 3782 | 205 | 660 | 18671 | 46290 |
| shCtrl_2nd | 341 | 972 | 2644 | 321 | 3395 | 390 | 1382 | 3072 | 120 | 401 | 13038 | 22297 |
| shCK18_1st | 381 | 1061 | 2737 | 360 | 3494 | 407 | 1485 | 3466 | 104 | 396 | 13891 | 24036 |
| shCK18_2nd | 503 | 1420 | 3305 | 391 | 4101 | 490 | 1642 | 3647 | 139 | 508 | 16146 | 34627 |
| All
samplesa | 1313 | 3282 | 8950 | 1201 | 10843 | 1360 | 4380 | 7750 | 416 | 1451 | 40946 | 86961 |
We identified 263 high confidence RASEs by using a
stringent cutoff of P≤0.05, along with changed AS ratio ≥0.2.
CK18-regulated ASEs, which included 35 alternative 3'splice sites
(A3SS), 40 alternative 5'splice sites (A5SS), 44 examples of exon
skipping (ES) and 24 cassette exons, 12 mutually exclusive 5′UTRs
(5pMXE), four mutually exclusive 3′UTRs (3pMXE), six mutually
exclusive exons (MXE), 84 examples of intron retention (IR), eight
examples of alternative 5'splice site & exon skipping (A5SS
& ES) and six examples of alternative 3'splice site & exon
skipping (A3SS & ES), are summarized in Fig. 4A. These results indicated that
CK18 KD had a broad influence on splicing. Coupled to the
transcription data, there were no significant expression changes in
alternative splicing genes (Fig.
4B). The alternative spliced genes that were identified by GO
analyzed were, however, enriched in liver development, positive
regulation of I-κB kinase/NF-κB cascade, glucose metabolic process,
positive regulation of NF-κB transcription factor activity, mRNA
processing and small molecule metabolic process. These results were
similar to those obtained by the transcriptional GO analysis,
further confirming the important regulatory role of CK18 in these
biological functions. The detailed results of the GO and KEGG
pathway analyses are presented in Fig.
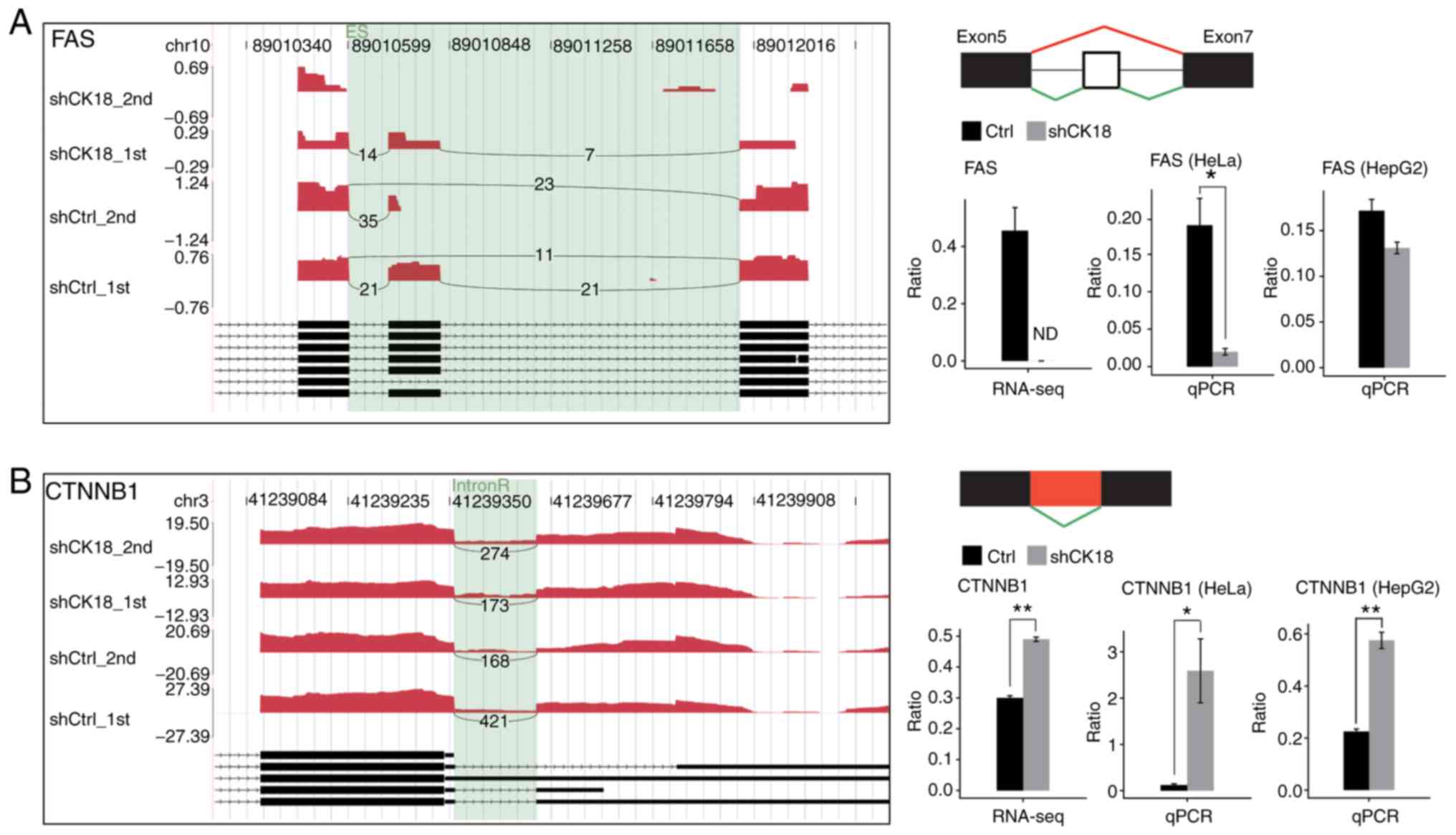
4C. It was observed that alternative splicing of CASP8 and FAS,
the known upstream genes regulating the production of CK18, was
under the regulation of CK18.
To confirm the important regulatory function of CK18
on both gene expression levels of alternative splicing in HeLa and
HepG2 cell lines, we validated certain DEGs and ASEs that were
important in immunity and apoptosis by RT-qPCR. The ASEs detected
by PCR primer pairs (Table I) were
designed to amplify both long splicing isoforms and the short
splicing isoforms in the same reaction.
Most of the DEGs associated with immune function and
apoptosis that we validated were in agreement with RNA-seq results
(Fig. 5A and B). For CASP8
gene expression, however, there was no significant change in
RNA-seq data, whereas the qPCR and western blotting results
revealed an evident decrease in CK18 KD cells (Fig. 5C). Two important ASEs were located
in FAS and CTNNB1, which have been well validated as
key genes in apoptotic pathways (Fig.
6).
Discussion
Knockdown of the expression level of CK18 by
a shRNA in HeLa cells led to a significant reduction of apoptosis,
indicating that CK18 is an apoptotic gene in the cancer
cells. Through genome-wide transcriptional and ASE analysis of
CK18 knockdown and control HeLa cells using RNA-seq, it was
revealed that the FAS-mediated apoptosis pathway was regulated by
CK18. Additionally, CASP8, which is downstream of the FAS-induced
apoptosis pathway, was revealed to be feedback-regulated by its
cleavage substrate, CK18. These findings indicated that CK18 had
more diverse regulatory functions than have been generally
recognized.
We have unbiasedly analyzed DEGs whose expression
was regulated by CK18. These DEGs were enriched in multiple GO
functional clusters and KEGG pathways. The finding that CK18
regulates the expression of genes enriched in multicellular
organismal development and signal transduction was consistent with
the reported function of CK18 in maintaining integrity of
epithelial cells and organismal development (39,40).
CK18-regulated genes were also enriched in immune response-related
pathways, which includes the B-cell receptor signaling pathway, the
T-cell receptor signaling pathway, the TNF signaling pathway and
the Jak-STAT signaling pathway.
In addition to transcriptional regulation,
CK18 also regulates alternative splicing of a large number
of genes, which are enriched in NF-κB regulatory processes. NF-κB
transcription factors are central coordinators of innate and
adaptive immune responses. Activated NF-κB regulates the expression
of over 300 different genes (41).
Activated NF-κB generally inhibits apoptosis by activation of
anti-apoptotic genes, although in some cases NF-κB promotes
apoptosis (42,43). Fortier et al have reported
that PI3K/Akt/NF-κB is activated when both CK8 and CK18 are knocked
down in HepG2 and KLE cells, thus increasing cell migration and
invasion (44).
CK18 has caspase recognition sites, which can be
cleaved during apoptosis caused by tumor pathogenesis or
chemotherapy. Fas- and TNF-mediated apoptosis are associated with
caspase-mediated CK18 degradation. A previous study revealed a
higher incidence of FasL-induced apoptosis in CK18− HeLa
cells compared with CK18+ HeLa cells (27). Another study revealed that
siRNA-mediated knockdown of K8/18 filament expression enhanced the
expression of the apoptotic gene FAS (>70% of cells) in
granulosa cell tumor cell line, KGN (45). CK18 mutation has, however, been
determined to disrupt FAS-mediated apoptosis in the livers of
transgenic mice (46).
In the present study, it was revealed that mRNA
levels of FAS and FADD in HeLa cells were positively
regulated by CK18, indicating that CK18-modulated apoptosis was
regulated by FAS but not TNF. A decline in the ratio of FAS
exon 6 skipping was also revealed. Skipping of FAS exon 6
resulted in an mRNA encoding a soluble form of FAS receptor that
acted as a decoy to prevent cell apoptosis (47–50).
It appears, therefore, that CK18 had dual regulatory effects on
FAS-mediated apoptosis, i.e., CK18 positively regulated FAS
transcription and also regulated production of the decoy isoform of
FAS mRNA.
FAS and FADD have been revealed to recruit and
activate CASP8, which plays an essential role in cleavage of the
actin cytoskeleton, including cytokeratin (51). In the present study, it was revealed
that CK18 regulated the alternative splicing of CASP8. In
addition, western blot analysis revealed that cleaved (activated)
CASP8 was significantly reduced upon CK18 knockdown. It is,
therefore, possible that CK18 exerts feedforward regulation of its
own cleavage by positively regulating the expression CASP8.
Full-length CK18 (M65) and the caspase-cleaved
fragment (M30) are recognized as useful markers in clinical
diagnosis and prognostic evaluation (18) and recently, the important regulatory
role of CK18 has also attracted attention. A recent study revealed
that CK18 could associate with histone H3, leading to aberrant
expression of histone deacetylase in hepatocellular carcinoma
(52). The finding that CK18
physically interacts with LRP16, sequestering LRP16 in the
cytoplasm and thus inhibiting the proliferation of ERα-positive
breast cancer cells (29), suggests
that CK18 may affect gene expression via protein-protein
interaction. Additionally, CK18 was revealed to be a potential RNA
binding protein (30). RNA binding
proteins play important roles in post-transcriptional gene
regulation. We therefore propose that CK18 may regulate gene
expression and alternative splicing through its protein and RNA
binding functions. Further studies are required to explore this
possibility.
CK18 dysregulation occurs widely in various tumors,
although the regulatory mechanism in tumors is largely unclear. The
knowledge that CK18 regulates the transcription and alternative
splicing of NF-κB genes and the transcription of genes related to
cancer cell migration and metastasis should expand our
understanding of the biological and regulatory functions of CK18.
It should also contribute to an improved understanding of the
prognostic value and clinicopathological significance of CK18.
Acknowledgements
We are especially grateful to Hong Wu for the
language polishing.
Funding
No funding was received.
Availability of data and materials
RNA-seq data mentioned in this article are publicly
available at the NCBI Gene Expression Omnibus (GEO) under the
accession no. GSE119255.
Author's contributions
YC performed experiments, contributed to analysis
and interpretation of the data, and was a major contributor in
writing the manuscript. KQ and NH performed experiments, and
contributed in drafting and revising the manuscript. ZZ contributed
in the data analysis and revision of the manuscript, HX and JZ
performed the cell experiments, and contributed in the revision of
the manuscript. YZ contributed to the design of the study and the
writing and revision of the manuscript. SY contributed to the
conception and design of the study, the data analysis and
interpretation, and the writing and revision of the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Koh WJ, Greer BE, Abu-Rustum NR, Apte SM,
Campos SM, Cho KR, Chu C, Cohn D, Crispens MA, Dorigo O, et al:
Cervical Cancer, Version 2.2015. J Natl Compr Canc Netw.
13:395–404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng R, Zeng H, Zhang S, Chen T and Chen
W: National estimates of cancer prevalence in China, 2011. Cancer
Lett. 370:33–38. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Coulombe PA and Omary MB: ‘Hard’ and
‘soft’ principles defining the structure, function and regulation
of keratin intermediate filaments. Curr Opin Cell Biol. 14:110–122.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Greystoke A, Dean E, Saunders MP, Cummings
J, Hughes A, Ranson M, Dive C and Renehan AG: Multi-level evidence
that circulating CK18 is a biomarker of tumour burden in colorectal
cancer. Br J Cancer. 107:1518–1524. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schneider J: Tumor markers in detection of
lung cancer. Adv Clin Chem. 42:1–41. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hernandez BY, Frierson HF, Moskaluk CA, Li
YJ, Clegg L, Cote TR, McCusker ME, Hankey BF, Edwards BK and
Goodman MT: CK20 and CK7 protein expression in colorectal cancer:
Demonstration of the utility of a population-based tissue
microarray. Hum Pathol. 36:275–281. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moll R, Divo M and Langbein L: The human
keratins: Biology and pathology. Histochem Cell Biol. 129:705–733.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ahn SK, Moon HG, Ko E, Kim HS, Shin HC,
Kim J, You JM, Han W and Noh DY: Preoperative serum tissue
polypeptide-specific antigen is a valuable prognostic marker in
breast cancer. Int J Cancer. 132:875–881. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Escobar-Hoyos LF, Shah R, Roa-Peña L,
Vanner EA, Najafian N, Banach A, Nielsen E, Al-Khalil R, Akalin A,
Talmage D and Shroyer KR: Keratin-17 promotes p27KIP1 nuclear
export and degradation and offers potential prognostic utility.
Cancer Res. 75:3650–3662. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan HS, Jiang WH, He Y, Wang DS, Wu ZJ, Wu
DS, Gao L, Bao Y, Shi JZ, Liu B, et al: KRT8 upregulation promotes
tumor metastasis and is predictive of a poor prognosis in clear
cell renal cell carcinoma. Oncotarget. 8:76189–76203. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheung KJ, Padmanaban V, Silvestri V,
Schipper K, Cohen JD, Fairchild AN, Gorin MA, Verdone JE, Pienta
KJ, Bader JS and Ewald AJ: Polyclonal breast cancer metastases
arise from collective dissemination of keratin 14-expressing tumor
cell clusters. Proc Natl Acad Sci USA. 113:E854–E863. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hsu J: Nuclear Keratin 17 and its role in
the DNA Damage Response. Doctoral dissertation. Johns Hopkins
University. 2017.
|
|
13
|
Saha SK, Choi HY, Kim BW, Dayem AA, Yang
GM, Kim KS, Yin YF and Cho SG: KRT19 directly interacts with
β-catenin/RAC1 complex to regulate NUMB-dependent NOTCH signaling
pathway and breast cancer properties. Oncogene. 36:332–349. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bilici A: Cytokeratin 18 for chemotherapy
efficacy in gastric cancer. Transl Gastrointest Cancer. 4:200–206.
2015.
|
|
15
|
Zhang B, Wang J, Liu W, Yin Y, Qian D,
Zhang H, Shi B, Li C, Zhu J, Zhang L, et al: Cytokeratin 18
knockdown decreases cell migration and increases chemosensitivity
in non-small cell lung cancer. J Cancer Res Clin Oncol.
142:2479–2487. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin B, Zhang M, Zeng Y, Li Y, Zhang C,
Getzenberg RH and Song Y: Downregulation of cytokeratin 18 is
associated with paclitaxel-resistance and tumor aggressiveness in
prostate cancer. Int J Oncol. 48:1730–1736. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang YL, Chen J, Yan W, Zang D, Qin Q and
Deng AM: Diagnostic accuracy of cytokeratin-19 fragment (CYFRA
21-1) for bladder cancer: A systematic review and meta-analysis.
Tumour Biol. 36:3137–3145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nagel M, Schulz J, Maderer A, Goepfert K,
Gehrke N, Thomaidis T, Thuss-Patience PC, Al-Batran SE,
Hegewisch-Becker S, Grimminger P, et al: Cytokeratin-18 fragments
predict treatment response and overall survival in gastric cancer
in a randomized controlled trial. Tumor Biol.
40:1010428318764002018. View Article : Google Scholar
|
|
19
|
Sjöström J, Alfthan H, Joensuu H, Stenman
UH, Lundin J and Blomqvist C: Serum tumour markers CA 15-3, TPA,
TPS, hCGbeta and TATI in the monitoring of chemotherapy response in
metastatic breast cancer. Scand J Clin Lab Invest. 61:431–441.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yaman E, Coskun U, Sancak B, Buyukberber
S, Ozturk B and Benekli M: Serum M30 levels are associated with
survival in advanced gastric carcinoma patients. Int
Immunopharmacol. 10:719–722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Demiray M, Ulukaya EE, Arslan M, Gokgoz S,
Saraydaroglu O, Ercan I, Evrensel T and Manavoglu O: Response to
neoadjuvant chemotherapy in breast cancer could be predictable by
measuring a novel serum apoptosis product, caspase-cleaved
cytokeratin 18: A prospective pilot study. Cancer Invest.
24:669–676. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ulukaya E, Yilmaztepe A, Akgoz S, Linder S
and Karadag M: The levels of caspase-cleaved cytokeratin 18 are
elevated in serum from patients with lung cancer and helpful to
predict the survival. Lung Cancer. 56:399–404. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ozturk B, Coskun U, Sancak B, Yaman E,
Buyukberber S and Benekli M: Elevated serum levels of M30 and M65
in patients with locally advanced head and neck tumors. Int
Immunopharmacol. 9:645–648. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Homberg M and Magin TM: Beyond
expectations: Novel insights into epidermal keratin function and
regulation. Int Rev Cell Mol Biol. 311:265–306. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Weng YR, Cui Y and Fang JY: Biological
functions of cytokeratin 18 in cancer. Mol Cancer Res. 10:485–493.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Messai Y, Noman MZ, Derouiche A, Kourda N,
Akalay I, Hasmim M, Stasik I, Ben Jilani S, Chebil M, Caignard A,
et al: Cytokeratin 18 expression pattern correlates with renal cell
carcinoma progression: Relationship with Snail. Int J Oncol.
36:1145–1154. 2010.PubMed/NCBI
|
|
27
|
Sullivan BT, Cherry JA, Sakamoto H, Henkes
LE, Townson DH and Rueda BR: Cytokeratin 18 expression inhibits
cytokine-induced death of cervical cancer cells. Int J Gynecol
Cancer. 20:1474–1481. 2010.PubMed/NCBI
|
|
28
|
Bühler H and Schaller G: Transfection of
keratin 18 gene in human breast cancer cells causes induction of
adhesion proteins and dramatic regression of malignancy in vitro
and in vivo. Mol Cancer Res. 3:365–371. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meng Y, Wu Z, Yin X, Zhao Y, Chen M, Si Y,
Yang J, Fu X and Han W: Keratin 18 attenuates estrogen receptor
alpha-mediated signaling by sequestering LRP16 in cytoplasm. BMC
Cell Biol. 10:962009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Castello A, Fischer B, Eichelbaum K, Horos
R, Beckmann BM, Strein C, Davey NE, Humphreys DT, Preiss T,
Steinmetz LM, et al: Insights into RNA biology from an atlas of
mammalian mRNA-binding proteins. Cell. 149:1393–1406. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gerstberger S, Hafner M and Tuschl T: A
census of human RNA-binding proteins. Nat Rev Genet. 15:829–845.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Landry JJ, Pyl PT, Rausch T, Zichner T,
Tekkedil MM, Stütz AM, Jauch A, Aiyar RS, Pau G, Delhomme N, et al:
The genomic and transcriptomic landscape of a HeLa cell line. G3
(Bethesda). 3:1213–1224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim D, Pertea G, Trapnell C, Pimentel H,
Kelley R and Salzberg SL: TopHat2: Accurate alignment of
transcriptomes in the presence of insertions, deletions and gene
fusions. Genome Biol. 14:R362013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Robinson MD, McCarthy D and Smyth GK:
edgeR: A Bioconductor package for differential expression analysis
of digital gene expression data. Bioinformatics. 26:139–140. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie C, Mao X, Huang J, Ding Y, Wu J, Dong
S, Kong L, Gao G, Li CY and Wei L: KOBAS 2.0: A web server for
annotation and identification of enriched pathways and diseases.
Nucleic Acids Res 39 (Web Server Issue). W316–W322. 2011.
View Article : Google Scholar
|
|
37
|
Xia H, Chen D, Wu Q, Wu G, Zhou Y, Zhang Y
and Zhang L: CELF1 preferentially binds to exon-intron boundary and
regulates alternative splicing in HeLa cells. Biochim Biophys Acta
Gene Regul Mech. 1860:911–921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Trapnell C, Williams BA, Pertea G,
Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ and Pachter
L: Transcript assembly and quantification by RNA-Seq reveals
unannotated transcripts and isoform switching during cell
differentiation. Nat Biotechnol. 28:511–515. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Karantza V: Keratins in health and cancer:
More than mere epithelial cell markers. Oncogene. 30:127–138. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Toivola DM, Boor P, Alam C and Strnad P:
Keratins in health and disease. Curr Opin Cell Biol. 32:73–81.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Serasanambati MR and Chilakapati SR:
Function of nuclear factor Kappa B (NF-κB) in human diseases-a
review. South Indian J Biol Sci. 2:368–387. 2016. View Article : Google Scholar
|
|
42
|
Magné N, Toillon RA, Bottero V, Didelot C,
Houtte PV, Gérard JP and Peyron JF: NF-kappaB modulation and
ionizing radiation: Mechanisms and future directions for cancer
treatment. Cancer Lett. 231:158–168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen X, Kandasamy K and Srivastava RK:
Differential roles of RelA (p65) and c-Rel subunits of nuclear
factor kappa B in tumor necrosis factor-related apoptosis-inducing
ligand signaling. Cancer Res. 63:1059–1066. 2003.PubMed/NCBI
|
|
44
|
Fortier AM, Asselin E and Cadrin M:
Keratin 8 and 18 loss in epithelial cancer cells increases
collective cell migration and cisplatin sensitivity through
claudin1 up-regulation. J Biol Chem. 288:11555–11571. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Trisdale SK, Schwab NM, Hou X, Davis JS
and Townson DH: Molecular manipulation of keratin 8/18 intermediate
filaments: modulators of FAS-mediated death signaling in human
ovarian granulosa tumor cells. J Ovarian Res. 9:82016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ku NO, Soetikno RM and Omary MB: Keratin
mutation in transgenic mice predisposes to Fas but not TNF-induced
apoptosis and massive liver injury. Hepatology. 37:1006–1014. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cheng J, Zhou T, Liu C, Shapiro JP, Brauer
MJ, Kiefer MC, Barr PJ and Mountz JD: Protection from Fas-mediated
apoptosis by a soluble form of the Fas molecule. Science.
263:1759–1762. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu C, Cheng J and Mountz JD: Differential
expression of human Fas mRNA species upon peripheral blood
mononuclear cell activation. Biochem J. 310:957–963. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Papoff G, Cascino I, Eramo A, Starace G,
Lynch DH and Ruberti G: An N-terminal domain shared by Fas/Apo-1
(CD95) soluble variants prevents cell death in vitro. J Immunol.
156:4622–4630. 1996.PubMed/NCBI
|
|
50
|
Tejedor JR, Papasaikas P and Valcárcel J:
Genome-wide identification of Fas/CD95 alternative splicing
regulators reveals links with iron homeostasis. Mol Cell. 57:23–38.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stegh AH, Herrmann H, Lampel S,
Weisenberger D, Andrä K, Seper M, Wiche G, Krammer PH and Peter ME:
Identification of the cytolinker plectin as a major early in vivo
substrate for caspase 8 during CD95- and tumor necrosis factor
receptor-mediated apoptosis. Mol Cell Biol. 20:5665–5679. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lai YC, Cheng CC, Lai YS and Liu YH:
Cytokeratin 18-associated histone 3 modulation in hepatocellular
carcinoma: A mini review. Cancer Genomics Proteomics. 14:219–223.
2017. View Article : Google Scholar : PubMed/NCBI
|