Introduction
Programmed cell death 4 (PDCD4) is an
apoptosis-related gene. It is currently identified as a
tumor-suppressor gene that inhibits neoplastic transformation
(1,2),
tumor progression (3) and protein
translation (4). PDCD4 is expressed
ubiquitously in different normal tissues, especially the liver
(5). However, the expression of PDCD4
in various types of cancers, such as liver cancer (6), lung cancer (7), breast cancer (8), is lost or decreased. We also found that
the expression of PDCD4 is downregulated in ovarian cancer tissues
compared with control ovarian epithelial tissues (9). It has been reported that multiple
factors participate in the regulation of PDCD4 mRNA and
protein. PDCD4 was first identified as an upregulated gene
during apoptosis (5). Different
pro-apoptotic substances, including ionomycin,
phorbol-12-myristate-13-acetate (PMA) or dexamethasone, were found
to induce increased levels of PDCD4 (10), but other inducers of apoptosis such as
arabinosyl cytosine (ara-C), UV irradiation or topoisomerase
inhibitors had no effect on the expression of PDCD4 (11,12).
Similarly, the levels of PDCD4 were found to be induced by
interleukin (IL)-12, but decreased by IL-2 and IL-15 treatment
(13). These results suggest that the
expression of PDCD4 may vary depending on different stimuli
(11).
The endometrium is regulated by changing
concentrations of ovarian hormones, such as estrogen and
progesterone, and shows periodical changes, including proliferative
phase, secretory phase and menstrual phase. Estrogen is increased
in the proliferative phase of the endometrium; after ovulation
progesterone is produced and drives the endometrium into a
secretory phase. When the levels of estrogen and progesterone are
decreased, the endometrium enters into a menstrual phase.
Therefore, the levels of ovarian hormones during the menstrual
cycle are changed. It has been reported that ovarian hormones
affect the expression of many genes, such as catalase (14) and IL-10R (15), and further induce gene expression
variation during the cycle changes of the endometrium. Ovarian
hormones not only regulate gene transcription by directly or
indirectly binding to DNA, but also regulate gene expression by
binding to corresponding membrane receptors and further initiate
various signaling pathways, including mitogen-activated protein
kinase/extracellular regulated protein kinase (MAPK/ERK) and
phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) pathways
(16). In a previous study, we found
that the expression of PDCD4 in the proliferative phase of the
endometrium was higher than that in the secretory phase of the
endometrium (17). This suggests that
the expression of PDCD4 may be regulated by ovarian hormones.
However, the effect and mechanism of ovarian hormones on the
expression of PDCD4 remain unclear.
In the present study, we aimed to investigate
whether PDCD4 expression is regulated by estrogen and progesterone
via the MAPK/ERK or PI3K/AKT pathways, and it was confirmed that
progesterone could downregulate the expression of PDCD4 protein via
PI3K/AKT pathway.
Materials and methods
Primary antibodies
Rabbit polyclonal or monoclonal antibodies specific
for PDCD4 (cat. no. cst-9535S); MAPK signaling pathway-related
molecules: Stress-activated protein kinase/Jun-amino-terminal
kinase (SAPK/JNK) (cat. no. cst-9252), p44/42MAPK (ERK1/2) (cat.
no. cst-4695), p38MAPK (cat. no. cst-8690); PI3K/AKT signaling
pathway-related molecules: AKT (cat. no. cst-4691), mammalian
target of rapamycin (mTOR) (cat. no. cst-2983), as well as
phospho-SAPK/JNK (phosphorylation site: Thr183/Tyr185) (cat. no.
cst-4668), phospho-p44/42 MAPK (ERK1/2) (phosphorylation site:
Thr202/Tyr204) (cat. no. cst-4370), phospho-p38MAPK
(phosphorylation site: Thr180/Tyr182) (cat. no. cst-4511),
phospho-AKT (phosphorylation site: Ser473) (cat. no. cst-4060),
phospho-mTOR (phosphorylation site: Ser2448) (cat. no. cst-5536)
were purchased from Cell Signaling Technology. Mouse monoclonal
antibody specific for β-actin (TA-09; ZSGB-Bio, Beijing, China) was
used as a loading control.
Cell culture
Well-differentiated human endometrial cancer
Ishikawa cells were kindly provided by Qilu Hospital. The cells
were cultured in Dulbecco's modified Eagle's medium (DMEM)-High
Glucose medium (Hyclone) containing 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.), penicillin and streptomycin (100X,
MACGENE). Moderately differentiated endometrial cancer HEC-1-A
cells were obtained from the China Center for Type Culture
Collection (Wuhan, China). The cells were cultured in McCoy's 5A
medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum, penicillin and streptomycin. All the cells
were incubated in a humidified atmosphere with 5% CO2 at
37°C.
Hormone treatments
The cells were respectively treated with 0, 0.1, 1,
10, 100 and 1,000 nM of 17β-estradiol (E2) (Sigma-Aldrich; Merck
KGaA) or 0, 0.01, 0.1, 1, 10 and 20 µM of progesterone (P4)
(Sigma-Aldrich; Merck KGaA). All treated cells were collected to
detect the expression of PDCD4 mRNA or protein by
quantitative real-time PCR (qPCR) or western blot analysis. The
cells were treated with 10 µM of progesterone for 0, 6, 12, 24, 36
and 48 h to determine the time when progesterone begins to reduce
the expression of PDCD4.
In order to investigate the signal transduction
pathway which was involved in PDCD4 down-regulation induced by
progesterone, the cells were firstly treated with 10 µM of
progesterone for 0, 1, 2 and 4 h, and the expression of signaling
molecules was detected by western blot analysis. In addition, the
cells were respectively pretreated with dimethyl sulfoxide (DMSO)
and 5 µM (18) of PI3K inhibitor
LY294002 (Selleckchem) for 1 h or 100 nM (19) of AKT inhibitor MK2206 (MedChem
Express) for 2 h, and then the cells were treated with 10 µM of
progesterone for 4 h. All treated cells were collected to detect
the expression of PDCD4 protein and signaling molecules.
RNA isolation and qPCR
Total RNA was extracted using RNAfast 200 (Fastagen)
according to the manufacturer's protocol, and then reversely
transcribed into complementary DNA using Reverse Transcription
System (Takara). The expression levels of genes were assessed by
qPCR using a 2X UltraSYBR Mixture and specific primers (CWBIO)
according to the manufacturer's instructions. qPCR was carried out
using the Applied Biosystems STEP One Plus Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The primers
for PDCD4 were as follows: forward, CCT GGA AGA TGG TGA TGG
GAT and reverse, AAC GGA TTT GGT CGT ATT GGG.
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used
for an internal control to analyze PDCD4 expression. The
primers for GAPDH were as follows: Forward: ACA GGT GTA TGA
TGT GGA GGA and reverse, TTC TCA AAT GCC CTT TCA TCC AA. The qPCR
Protocol was as follows: Pre-denaturation at 95°C for 10 min,
followed by 40 cycles with denaturation at 95°C for 15 sec and
annealing/elongation at 60°C for 1 min. The expression of
PDCD4 was analyzed according to the comparative quantitative
method (2−ΔΔCq) and the samples were examined in
triplicate.
Total protein extraction and western
blot analysis
The cells were lysed using radio-immunoprecipitation
assay (RIPA) lysis buffer (Beyotime) with protease inhibitor and
phosphatase inhibitor (Bimake). After homogenization, the cells
were centrifuged at 13,700 × g at 4°C for 30 min. The supernatant
was quantified using the bicinchoninic acid (BCA) assay kit (Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Equal amounts of proteins were separated by sodium
dodecyl sulfate (SDS) polyacrylamide gel, and then transferred onto
polyvinylidene fluoride (PVDF) membranes (Millipore). After being
blocked with 5% bovine serum albumin (Sigma-Aldrich; Merck KGaA) in
Tris-buffered saline Tween-20 (TBST) for 2~3 h. The membranes were
respectively incubated overnight at 4°C with a 1:1,000 dilution of
primary antibodies. Next day, the membranes were incubated with a
1:2,000 dilution of horseradish peroxidase-conjugated secondary
antibodies (Jackson Immuno Research) at room temperature for 1 h.
The signal was detected using the enhanced chemiluminescence kit
(Millipore). β-actin was used as an internal reference for
normalization of PDCD4. Total protein of mTOR, AKT, ERK1/2, P38,
JNK1/2 were used as loading control for normalization of their
phosphorylated protein levels.
Statistical analysis
Statistical analysis was performed with GraphPad
Prism 7.0 software (GraphPad Software, Inc.). All data are
presented as means ± SD. A Dunnett's post hoc test after one way
ANOVA was used to compared different test groups with one control
group. The Student's t-test was performed to evaluate the
statistical significance between two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of estrogen on the expression
of PDCD4 in human endometrial cancer cell lines
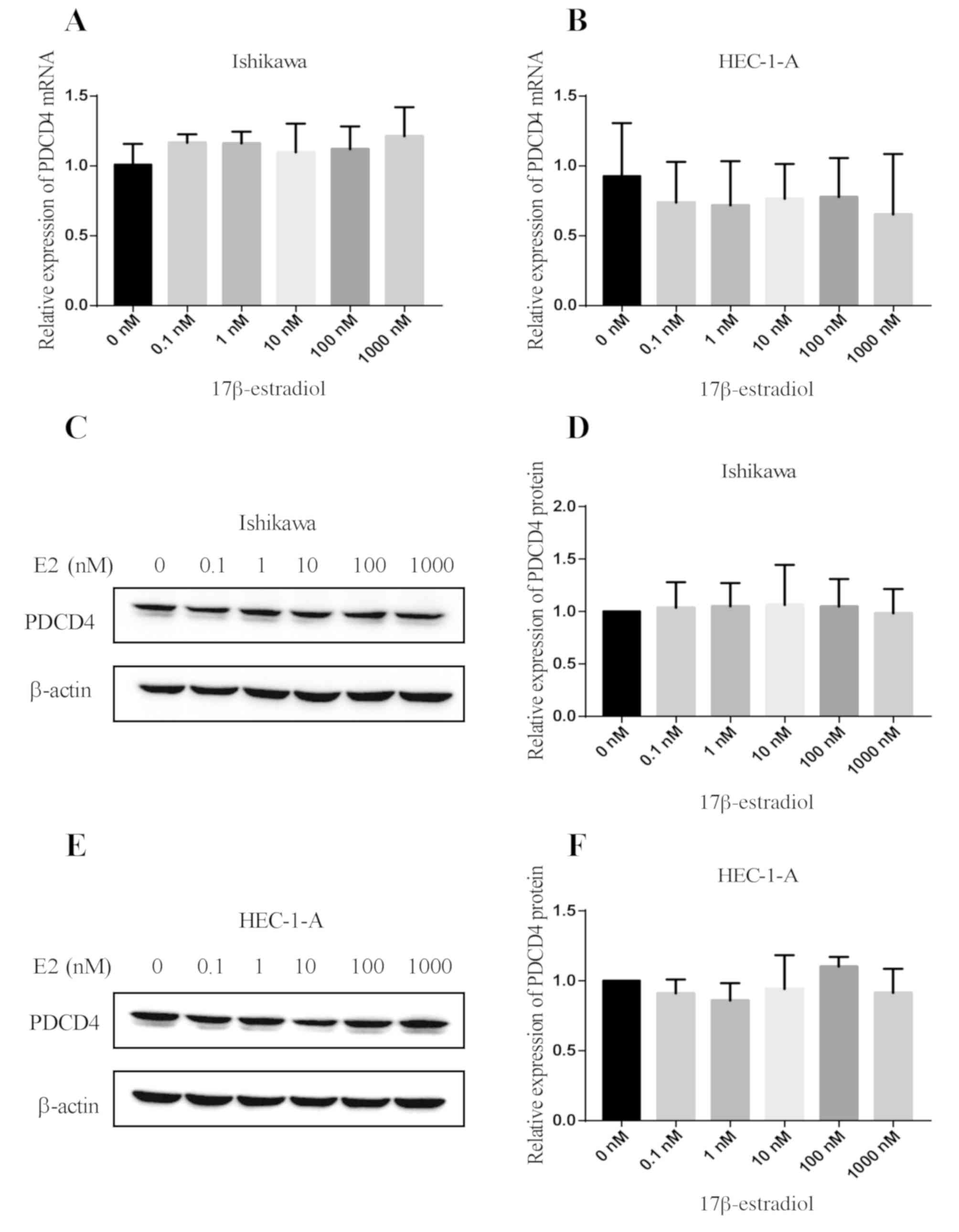
After Ishikawa and HEC-1-A cells were treated with
different concentrations of 17β-estradiol (E2) for 24 h, qPCR was
used to detect the expression of PDCD4 mRNA (Fig. 1A and B). After the above two cell
lines were treated with different concentrations of 17β-estradiol
(E2) for 48 h, western blot analysis was performed to detect PDCD4
protein expression (Fig. 1C-F). The
results showed that for any concentration of 17β-estradiol no
effect on the expression of PDCD4 mRNA and protein in the
two types of cell lines was observed (P>0.05) (Fig. 1).
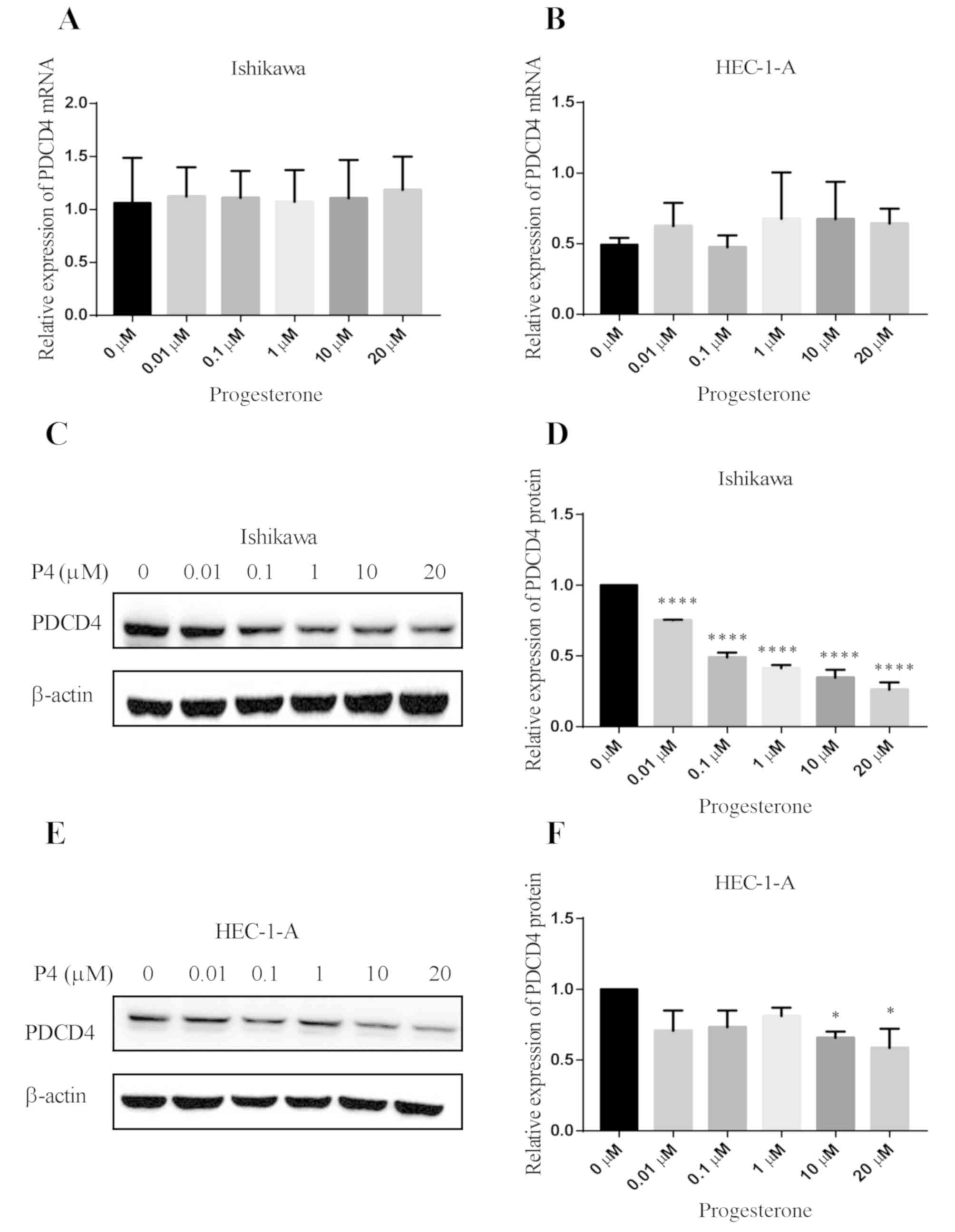
Effect of progesterone on the
expression of PDCD4 in human endometrial cancer cell lines
The effect of different concentrations of
progesterone on PDCD4 mRNA expression was determined in
Ishikawa and HEC-1-A cells. The results showed that at any
concentration of progesterone no significant effect on the
expression of PDCD4 mRNA was observed in the Ishikawa and
HEC-1-A cells (Fig. 2A and B).
Furthermore, an experiment of the time kinetic investigation on the
PDCD4 mRNA level in Ishikawa and HEC-1-A cells was
conducted. The results confirmed that progesterone failed to
regulate the expression of PDCD4 at the mRNA level (data not
shown). Then, the effect of different concentrations of
progesterone on PDCD4 protein expression was investigated in
Ishikawa and HEC-1-A cells. The results showed that different
concentrations (0.01, 0.1, 1, 10 and 20 µM) of progesterone
obviously downregulated the expression of PDCD4 protein in Ishikawa
cells (P<0.0001 vs. 0 µM) (Fig. 2C and
D); however, only 10 and 20 µM of progesterone effectively
reduced the expression of PDCD4 protein in HEC-1-A cells (P<0.05
vs. 0 µM) (Fig. 2E and F). Next, we
detected the effect of progesterone (10 µM) on the PDCD4 protein
expression at different time points in Ishikawa and HEC-1-A cells.
It was found that PDCD4 protein began to decrease after
progesterone treatment for 6 h in Ishikawa cells (P<0.01 vs. 0
h) (Fig. 3A and B). In HEC-1-A cells,
the expression of PDCD4 protein started to decrease after
progesterone treatment for 12 h (P<0.05 vs. 0 h) (Fig. 3C and D).
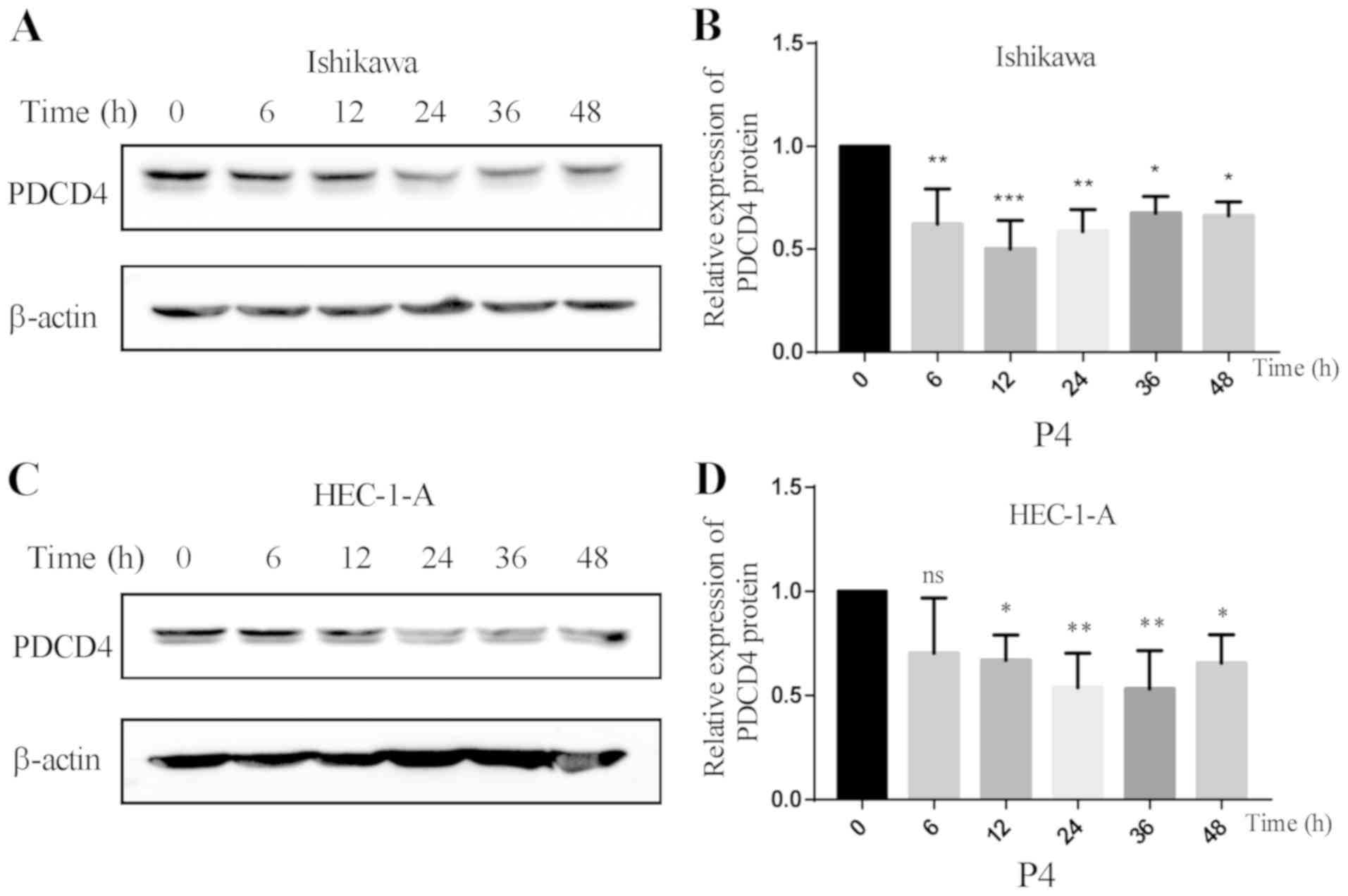 | Figure 3.Effect of progesterone on the
expression of PDCD4 protein at different time points. (A) The
expression of PDCD4 protein was detected by western blot analysis
in Ishikawa cells after treatment with 10 µM of progesterone at
different time points (0, 6, 12, 24, 36 and 48 h). (B) The
expression of PDCD4 protein began to decrease after progesterone
treatment for 6 h in Ishikawa cells (*P<0.05 vs. 0 h;
**P<0.01 vs. 0 h; ***P<0.01 vs. 0 h). (C) The expression of
PDCD4 protein was detected by western blot analysis in HEC-1-A
cells after treatment with 10 µM of progesterone at different time
points (0, 6, 12, 24, 36 and 48 h). (D) The expression of PDCD4
protein started to decrease after progesterone treatment for 12 h
in HEC-1-A cells (*P<0.05 vs. 0 h; **P<0.01 vs. 0 h; ns, not
significant). PDCD4, programmed cell death 4. |
Progesterone activates the
PI3K/AKT/mTOR pathway but not the MAPK pathway
We further determined whether the PI3K/AKT/mTOR
pathway participates in the effect of progesterone on PDCD4 protein
expression using 10 µM of progesterone to treat Ishikawa and
HEC-1-A cells for 0, 1, 2 and 4 h. It was found that the expression
of p-mTOR (P<0.05 vs. 0 h) and p-AKT (P<0.01 vs. 0 h) was
significantly higher after treatment with progesterone (P4) for 4 h
(Fig. 4A-C). In order to investigate
whether the MAPK signaling pathway participates in the
downregulation of PDCD4 protein expression induced by progesterone
(P4), the expression of p-ERK1/2, p-P38 and p-JNK was detected. It
was found that the levels of p-ERK1/2, p-P38 and p-JNK had no
significant changes following progesterone (P4) treatment
(P>0.05) (Fig. 4D-F).
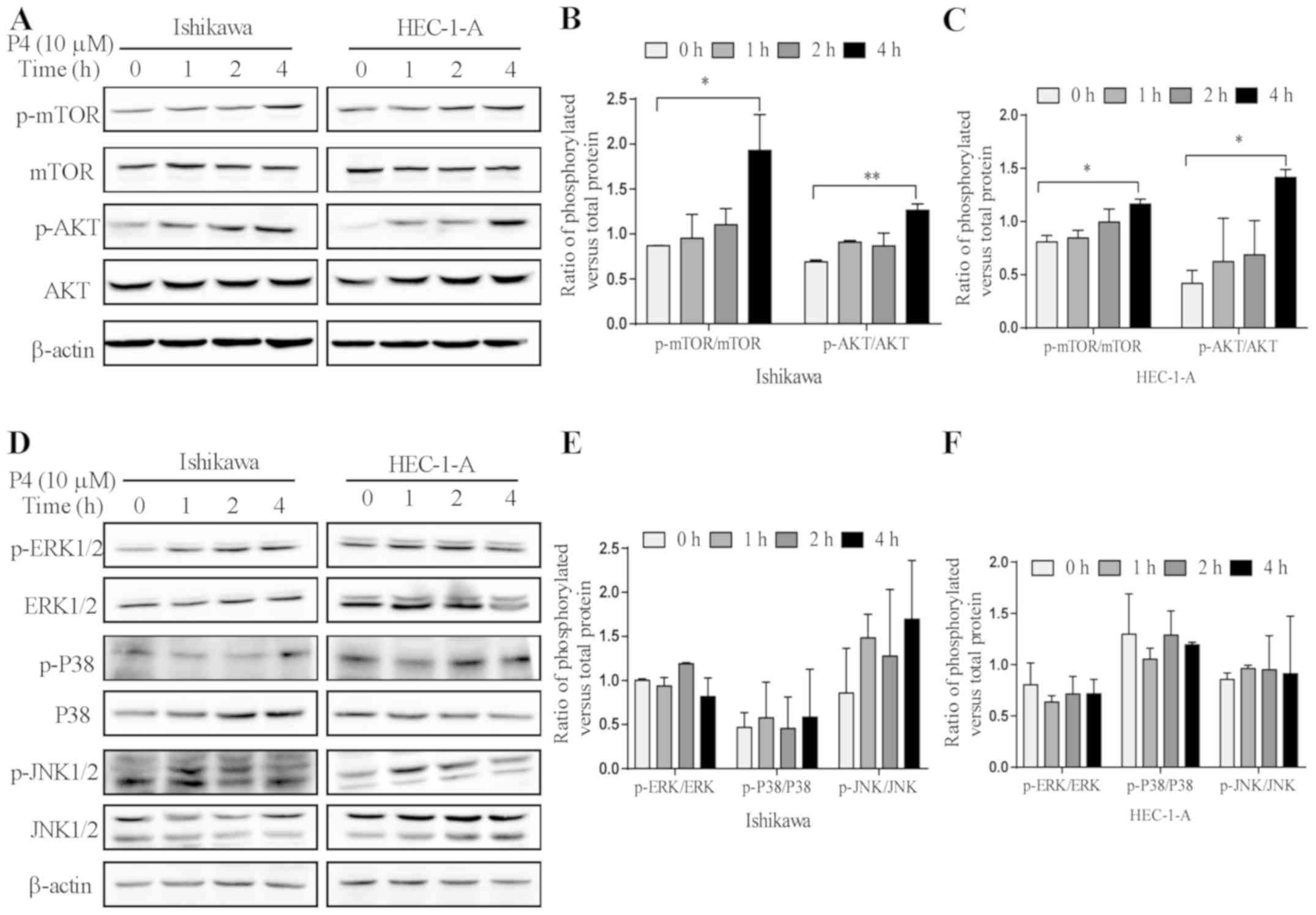 | Figure 4.Effect of progesterone on the
expression of signaling molecules in the PI3K/AKT/mTOR and MAPK
pathways. (A) The expression of phosphorylated (p)-mTOR, mTOR,
p-AKT, AKT and β-actin was detected by western blot analysis after
treatment with 10 µM of progesterone (P4) at different time points
(0, 1, 2 and 4 h). Quantitative analysis of the ratios of
p-mTOR/mTOR and p-AKT/AKT by measuring the relative band density in
(B) Ishikawa and (C) HEC-1-A cells. (D) The expression of p-ERK,
ERK, p-P38, P38, p-JNK, JNK and β-actin was detected by western
blot analysis after treatment with 10 µM of progesterone (P4) at
different time points (0, 1, 2 and 4 h). (E) Quantitative analysis
of the ratios of p-ERK/ERK, p-P38/P38 and p-JNK/JNK by measuring
the relative band density in (E) Ishikawa and (F) HEC-1-A cells.
*P<0.05 vs. 0 h; **P<0.01 vs. 0 h. |
The PI3K/AKT pathway participates in
the reduction of PDCD4 protein expression induced by
progesterone
In order to confirm that progesterone (P4)
downregulates the expression of PDCD4 protein via the PI3K/AKT
pathway in Ishikawa and HEC-1-A cells, we used a PI3K inhibitor
(LY294002) to pretreat Ishikawa and HEC-1-A cells for 1 h (Fig. 5A-C) or AKT inhibitor (MK2206) to
pretreat Ishikawa and HEC-1-A cells for 2 h (Fig. 5D-F), respectively, and then
progesterone (P4) was administered. The results showed that the
expression of p-AKT was significantly lower (P<0.05) and the
expression of PDCD4 was significantly increased (P<0.05) in both
cell lines when compared with P4 treatment alone, while the
expression of p-mTOR exhibited no obvious changes (P>0.05) in
Ishikawa and HEC-1-A cells (Fig.
5).
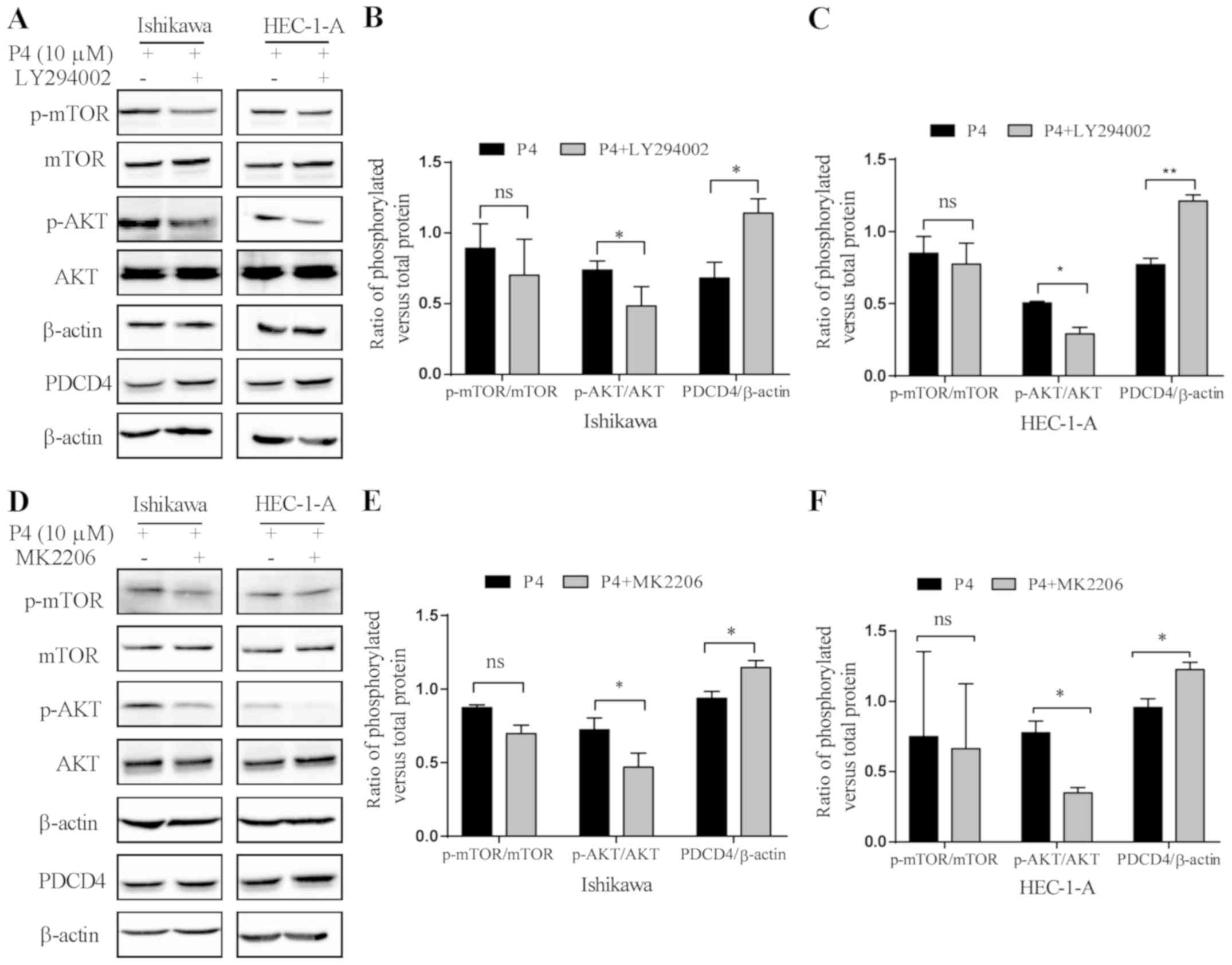 | Figure 5.The PI3K/AKT pathway participates in
the downregulation of PDCD4 protein expression induced by
progesterone. (A) The effect of LY294002 (PI3K inhibitor) on the
expression of phosphorylated (p)-mTOR, mTOR, p-AKT, AKT, PDCD4 and
β-actin in Ishikawa and HEC-1-A cells. Cells were pre-treated with
5 µM of LY294002 for 2 h, and then treated with 10 µM of
progesterone (P4) for 4 h, and the expression of p-mTOR, mTOR,
p-AKT, AKT, PDCD4 and β-actin was detected by western blot
analysis. (B) Quantitative analysis of the ratios of p-mTOR/mTOR,
p-AKT/AKT and PDCD4/β-actin by measuring the relative band density
in (B) Ishikawa and (C) HEC-1-A cells. (D) The effect of MK2206
(AKT inhibitor) on the expression of p-mTOR, mTOR, p-AKT, AKT,
PDCD4 and β-actin in Ishikawa and HEC-1-A cells. Cells were
pre-treated with 100 nM of MK2206 for 2 h, and then treated with 10
µM of progesterone (P4) for 4 h, and the expression of p-mTOR,
mTOR, p-AKT, AKT, PDCD4 and β-actin was detected by western blot
analysis. (E) Quantitative analysis of the ratios of p-mTOR/mTOR,
p-AKT/AKT and PDCD4/β-actin by measuring the relative band density
in (E) Ishikawa and (F) HEC-1-A cells. *P<0.05 vs. the control
group (P4 only); **P<0.01 vs. the control group (P4 only); ns,
not significant; P4, progesterone. |
Discussion
PDCD4, as a tumor suppressor, regulates the
expression of various genes related to tumor development and
progression at the transcriptional and translational levels. PDCD4
was found to be decreased or lost in different types of cancer,
including liver and lung cancer. The expression of PDCD4 is
regulated by various factors, such as apoptosis inducers and
cytokines. We previously found that PDCD4 expression was
significantly decreased in the progesterone-predominated secretory
phase of the normal endometrium compared with that in
estrogen-predominated proliferative phase (17), which suggests that PDCD4 expression is
hormonally regulated in the human endometrium.
The human endometrium is composed of epithelial and
stromal cells, and it is cyclically regenerated. The reproductive
cycle is induced by neuroendocrine signaling (20). The endometrium is driven into the
proliferative phase of the cycle by increased estrogen, and then
progesterone is produced by the ovary. The progesterone inhibits
proliferation of the endometrium and drives the endometrium into a
secretory phase in anticipation of fertilization (20). However, if there is no fertilization,
the endometrium is shed and hormone levels are decreased, and then
the cycle is reactivated (21,22). As
well known, prolonged exposure to estrogen is a major endocrine
risk factor for the establishment and progression of various
diseases, such as endometriosis (23)
and endometrial carcinoma (24).
Progesterone counteracts estrogen-mediated action and exhibits
anti-proliferative and anti-inflammatory roles (25–27).
Therefore, the antagonistic nature of progesterone to estrogen in
the endometrium empowers progesterone as the first-line of hormonal
therapy for the clinical treatment of endometriosis (28–31) and
endometrial carcinoma. However, the therapeutic efficacy and
beneficial effect of progesterone on the pathogenesis of
endometrial carcinoma remain debatable (32). In the present study, it was
demonstrated that progesterone downregulates PDCD4 protein
expression, which suggests that PDCD4 is a progesterone target
gene. Considering that PDCD4 is a type of tumor suppressor, which
inhibits tumor development and progression, downregulation of PDCD4
by progesterone may be one of the reasons that progesterone has
limited therapeutic efficacy.
Classically, the actions of progesterone are
attributed to the binding of progesterone and nuclear progesterone
receptor (nPR) and subsequent activation of its downstream target
genes (33). In addition, cell
membrane hormonal receptors, such as the membrane progesterone
receptor (mPR) family, have been identified and demonstrated to be
functional (34–37). mPR is able to activate the MAPK/ERK
and PI3K/AKT pathways, which also leads to regulation of gene
expression (38). Mammalian target of
rapamycin (mTOR) is a target gene of PI3K/AKT. It has been reported
that miR-21 inhibits PDCD4 expression and activates the
PI3K/AKT/mTOR signaling pathway (39). Dorrello et al (40) revealed a new signaling branch of the
mTOR pathway that controls the degradation of PDCD4. In the present
study, it was demonstrated that p-AKT and p-mTOR were increased and
PDCD4 was decreased after treatment of progesterone. Inhibition of
PI3K/AKT by LY294002/MK2206 reduced the expression of p-AKT and
upregulated the expression of PDCD4 protein, while expression of
p-mTOR exhibited no obvious changes. These results suggest that the
PI3K/AKT pathway is involved in progesterone-induced PDCD4 protein
downregulation.
In conclusion, it was demonstrated for the first
time that progesterone effectively decreases the expression of
PDCD4 protein, and the PI3K/AKT pathway may be involved in the
downregulation of PDCD4 protein. These results suggest that the
downregulation of PDCD4 induced by progesterone could affect the
therapeutic efficacy of progesterone in human endometrial cancer or
endometriosis, which may have important implications for
progesterone treatment in the clinic.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (81471437, 81771554),
Natural Science Foundation of Shandong (ZR2018MH013), Science and
Technology Development Plan provided by the Health and Family
Planning Committee in Shandong (2014–25).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZW designed the research project and conducted the
experimental study. XiaW participated in designing the experiments,
writing and reviewing the manuscript. XisW performed the
experiments and wrote the manuscript. YueL was involved in
performing various experiments. LW and YaL were involved in
statistical analysis. YSu and YuqL participated in preparation of
the manuscript. YSh and LZ were involved in revising it critically
for important intellectual content. HZ and JW assisted in data
analysis and reviewing the manuscript. All authors read and
approved the final manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
None of the authors has any potential financial
conflicts of interests related to this manuscript.
References
|
1
|
Cmarik JL, Min H, Hegamyer G, Zhan S,
Kulesz-Martin M, Yoshinaga H, Matsuhashi S and Colburn N:
Differentially expressed protein Pdcd4 inhibits tumor
promoter-induced neoplastic transformation. Proc Natl Acad Sci USA.
96:14037–14042. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang HS, Jansen AP, Nair R, Shibahara K,
Verma AK, Cmarik JL and Colburn NH: A novel transformation
suppressor, Pdcd4, inhibits AP-1 transactivation but not NF-kappaB
or ODC transactivation. Oncogene. 20:669–676. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ding L, Zhang X, Zhao M, Qu Z, Huang S,
Dong M and Gao F: An essential role of PDCD4 in progression and
malignant proliferation of gastrointestinal stromal tumors. Med
Oncology. 29:1758–1764. 2012. View Article : Google Scholar
|
|
4
|
LaRonde-LeBlanc N, Santhanam AN, Baker AR,
Wlodawer A and Colburn NH: Structural basis for inhibition of
translation by the tumor suppressor Pdcd4. Mol Cell Biol.
27:147–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lankat-Buttgereit B and Goke R: Programmed
cell death protein 4 (pdcd4): A novel target for antineoplastic
therapy? Biol Cell. 95:515–519. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang H, Ozaki I, Mizuta T, Hamajima H,
Yasutake T, Eguchi Y, Ideguchi H, Yamamoto K and Matsuhashi S:
Involvement of programmed cell death 4 in transforming growth
factor-beta1-induced apoptosis in human hepatocellular carcinoma.
Oncogene. 25:6101–6112. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen Y, Knosel T, Kristiansen G, Pietas A,
Garber ME, Matsuhashi S, Ozaki I and Petersen I: Loss of PDCD4
expression in human lung cancer correlates with tumour progression
and prognosis. J Pathol. 200:640–646. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wen YH, Shi X, Chiriboga L, Matsahashi S,
Yee H and Afonja O: Alterations in the expression of PDCD4 in
ductal carcinoma of the breast. Oncol Rep. 18:1387–1393.
2007.PubMed/NCBI
|
|
9
|
Wei NA, Liu SS, Leung TH, Tam KF, Liao XY,
Cheung AN, Chan KK and Ngan HY: Loss of programmed cell death 4
(Pdcd4) associates with the progression of ovarian cancer. Mol
Cancer. 8:702009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shibahara K, Asano M, Ishida Y, Aoki T,
Koike T and Honjo T: Isolation of a novel mouse gene MA-3 that is
induced upon programmed cell death. Gene. 166:297–301. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lankat-Buttgereit B and Goke R: The tumour
suppressor Pdcd4: Recent advances in the elucidation of function
and regulation. Biol Cell. 101:309–317. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Onishi Y and Kizaki H: Molecular cloning
of the genes suppressed in RVC lymphoma cells by topoisomerase
inhibitors. Biochem Biophysical Res Commun. 228:7–13. 1996.
View Article : Google Scholar
|
|
13
|
Azzoni L, Zatsepina O, Abebe B, Bennett
IM, Kanakaraj P and Perussia B: Differential transcriptional
regulation of CD161 and a novel gene, 197/15a, by IL-2, IL-15, and
IL-12 in NK and T cells. J Immunol. 161:3493–3500. 1998.PubMed/NCBI
|
|
14
|
Mundim TC, Ramos AF, Sartori R, Dode MA,
Melo EO, Gomes LF, Rumpf R and Franco MM: Changes in gene
expression profiles of bovine embryos produced in vitro, by natural
ovulation, or hormonal superstimulation. Genet Mol Res.
8:1398–1407. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qin X, Zhang H, Wang F, Xue J and Wen Z:
Expression and possible role of interleukin-10 receptors in
patients with adenomyosis. Eur j Obstet Gynecol Rep Biol.
161:194–198. 2012. View Article : Google Scholar
|
|
16
|
Lin SL, Yan LY, Zhang XT, Yuan J, Li M,
Qiao J, Wang ZY and Sun QY: ER-alpha36, a variant of ER-alpha,
promotes tamoxifen agonist action in endometrial cancer cells via
the MAPK/ERK and PI3K/Akt pathways. PLoS One. 5:e90132010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Y, Tan X, Wang Z, Li Y, Gao M, Li Y,
Fang Z, Sun Y, Zhang L, Wang X and Wei Z: Down-regulation of tumor
suppressor PDCD4 expression in endometrium of adenomyosis patients.
Curr Res Transl Med. 64:123–128. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li N, Wang J, Zhang N, Zhuang M, Zong Z,
Zou J, Li G, Wang X, Zhou H, Zhang L and Shi Y: Cross-talk between
TNF-α and IFN-γ signaling in induction of B7-H1 expression in
hepatocellular carcinoma cells. Cancer Immunol Immunother. 271–283.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pant A, Lee II, Lu Z, Rueda BR, Schink J
and Kim JJ: Inhibition of AKT with the orally active allosteric AKT
inhibitor, MK-2206, sensitizes endometrial cancer cells to
progestin. PLoS One. 7:e415932012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nabilsi NH, Broaddus RR, McCampbell AS, Lu
KH, Lynch HT, Chen LM and Loose DS: Sex hormone regulation of
survivin gene expression. J Endocrinol. 207:237–243. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bischof P, Krahenbuhl C and Desaulles PA:
Elucidation of the mechanism responsible for the luteolytic effect
of oestradiol during pseudogestation in the rat. Experientia.
30:1101–1102. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Knobil E: Hormonal control of the
menstrual cycle and ovulation in the rhesus monkey. Acta Endocrinol
Suppl (Copenh). 166:137–144. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Y, Gong P, Chen Y, Nwachukwu JC,
Srinivasan S, Ko C, Bagchi MK, Taylor RN, Korach KS, Nettles KW, et
al: Dual suppression of estrogenic and inflammatory activities for
targeting of endometriosis. Sci Transl Med. 7:271ra92015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stadel BV: Estrogen therapy and
endometrial carcinoma. Am J Obstet Gynecol. 125:571–573. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fang Z, Yang S, Lydon JP, DeMayo F, Tamura
M, Gurates B and Bulun SE: Intact progesterone receptors are
essential to counteract the proliferative effect of estradiol in a
genetically engineered mouse model of endometriosis. Ferti Steril.
82:673–678. 2004. View Article : Google Scholar
|
|
26
|
Cooke PS, Buchanan DL, Young P,
Calhaz-Jorge C, D'Hooghe T, De Bie B, Heikinheimo O, Horne AW,
Kiesel L and Nap A: Stromal estrogen receptors mediate mitogenic
effects of estradiol on uterine epithelium. Proc Natl Acad Sci USA.
94:6535–6540. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Adur MK, Kannan A, Davila J, Zhao Y,
Nowak RA, Bagchi MK, Bagchi IC and Li Q: Progesterone alleviates
endometriosis via inhibition of uterine cell proliferation,
inflammation and angiogenesis in an immunocompetent mouse model.
PLoS One. 11:e01653472016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dunselman GA, Vermeulen N, Becker C,
Calhaz-Jorge C, D'Hooghe T, De Bie B, Heikinheimo O, Horne AW,
Kiesel L, Nap A, et al: ESHRE guideline: Management of women with
endometriosis. Hum Reprod. 29:400–412. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mounsey AL, Wilgus A and Slawson DC:
Diagnosis and management of endometriosis. Am Fam Physician.
74:594–600. 2006.PubMed/NCBI
|
|
30
|
Vercellini P, Vigano P, Somigliana E and
Fedele L: Endometriosis: Pathogenesis and treatment. Nat Rev
Endocrinol. 10:261–275. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schweppe KW: Current place of progestins
in the treatment of endometriosis-related complaints. Gynecol
Endocrinol. 6:22–28. 2001. View Article : Google Scholar
|
|
32
|
Ruiz MP, Huang Y, Hou JY, Tergas AI, Burke
WM, Ananth CV, Neugut AI, Hershman DL and Wright JD: All-cause
mortality in young women with endometrial cancer receiving
progesterone therapy. Am J Obstet Gynecol. 217:669.e1–669.e13.
2017. View Article : Google Scholar
|
|
33
|
Xie M, Zhu X, Liu Z, Shrubsole M, Varma V,
Mayer IA, Dai Q, Chen Q and You S: Membrane progesterone receptor
alpha as a potential prognostic biomarker for breast cancer
survival: A retrospective study. PLoS One. 7:e351982012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thomas P: Characteristics of membrane
progestin receptor alpha (mPRalpha) and progesterone membrane
receptor component 1 (PGMRC1) and their roles in mediating rapid
progestin actions. Front Neuroendocrinol. 29:292–312. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dressing GE and Thomas P: Identification
of membrane progestin receptors in human breast cancer cell lines
and biopsies and their potential involvement in breast cancer.
Steroids. 72:111–116. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sleiter N, Pang Y, Park C, Horton TH, Dong
J, Thomas P and Levine JE: Progesterone receptor A (PRA) and
PRB-independent effects of progesterone on gonadotropin-releasing
hormone release. Endocrinology. 150:3833–3844. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu Y, Rice CD, Pang Y, Pace M and Thomas
P: Cloning, expression, and characterization of a membrane
progestin receptor and evidence it is an intermediary in meiotic
maturation of fish oocytes. Proc Natl Acad Sci USA. 100:2231–2236.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Piasecka D, Skladanowski AC, Kordek R,
Romanska HM and Sadej R: Aspects of progesterone receptor (PR)
activity regulation-impact on breast cancer progression. Postepy
Biochem. 61:198–206. 2015.PubMed/NCBI
|
|
39
|
Jiang LP, He CY and Zhu ZT: Role of
microRNA-21 in radiosensitivity in non-small cell lung cancer cells
by targeting PDCD4 gene. Oncotarget. 8:23675–23689. 2017.PubMed/NCBI
|
|
40
|
Dorrello NV, Peschiaroli A, Guardavaccaro
D, Colburn NH, Sherman NE and Pagano M: S6K1- and betaTRCP-mediated
degradation of PDCD4 promotes protein translation and cell growth.
Science. 314:467–471. 2006. View Article : Google Scholar : PubMed/NCBI
|