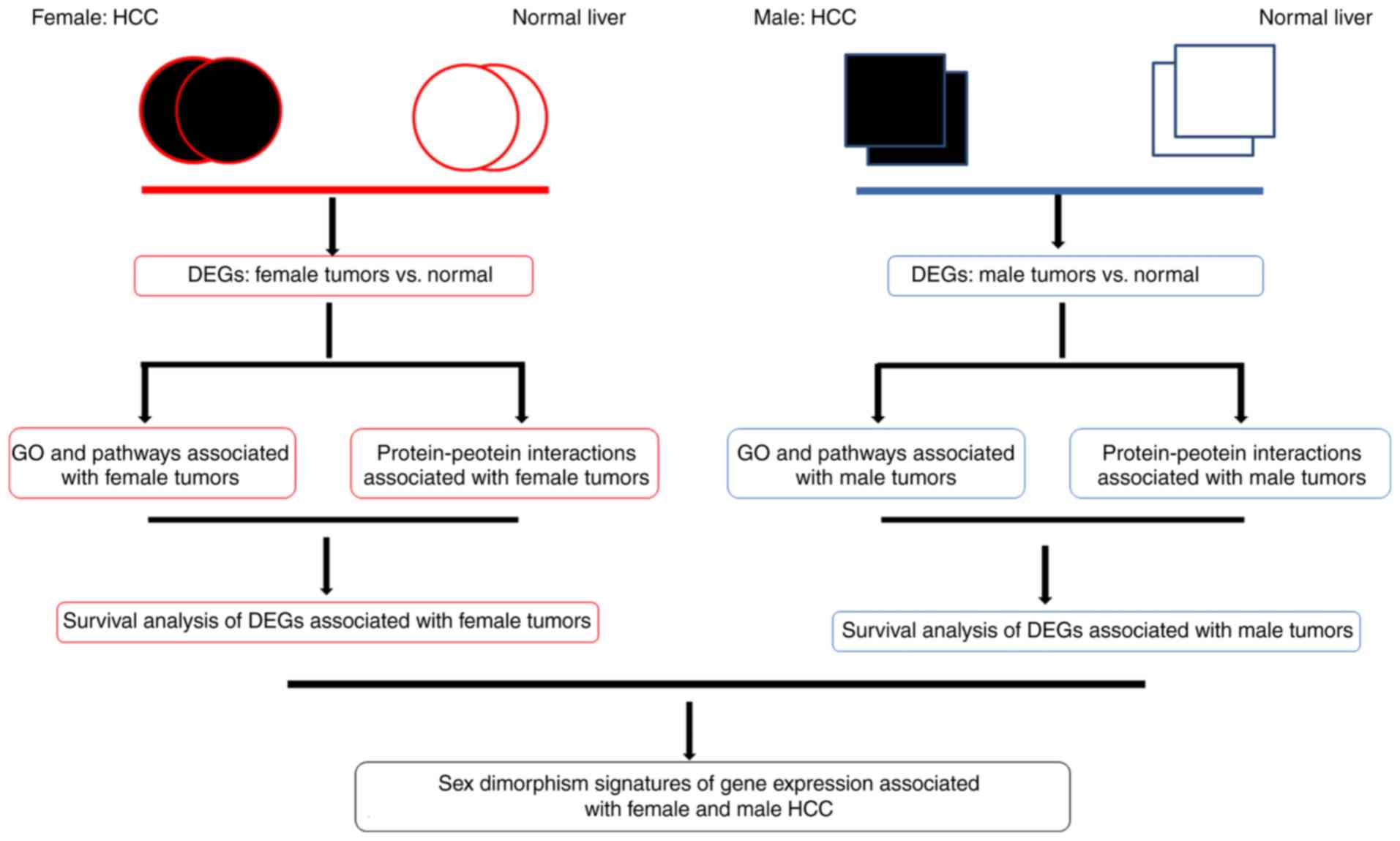
Introduction
Sexual disparity has long been recognized as an
independent key factor that can affect susceptibility to, and the
incidence, development and management of various types of diseases,
including vascular and heart disease, brain disorders and cancer
(1–3).
The susceptibility to cancer and the risk of malignancy are
generally much higher in male compared with in female subjects
(4–6).
Therefore, sex is considered a prognostic indicator for various
cancers (7); however, the endogenous
molecular signatures of sex-based differences in different types of
cancer require further investigation.
Liver cancer is the 5th most common type of cancer
and the second leading cause of mortality in males globally,
whereas it is the 7th most common cancer and the 6th leading cause
of mortality in women (3,8). Hepatocellular carcinoma (HCC) accounts
for 75–90% of primary liver cancers. The 5-year survival of
patients with HCC is only 7%, and the majority of the cases result
from risk factors, such as hepatitis B or hepatitis C virus
infection (9). A previous study
reported that the incidence of HCC is affected by sex (10) and predominant in males, with the
male:female ratio within populations usually ranging between 2:1
and 7:1 (11–13).
In human and rodent females, differences in the
incidence of HCC have been attributed to the expression of sexual
hormones and associated receptors, such as estrogen and the
estrogen receptor (ER) (14–16). For example, estrogen exerted
protective effects against chemically-induced HCC tumorigenesis by
inhibiting IL-6 production in macrophage-Kupffer cells, which in
turn reduced tumor proliferation (16). However, the expression levels of IL-6
did not necessarily correlate with tumor load in female and male
mice when forkhead box protein A (Foxa)1 and 2 is ablated in
hepatocytes (6). The steroid hormone
receptors ER and androgen receptor co-regulate Foxa factors, Foxa1
and Foxa2, which are transcription factors involved in
carcinogen-induced HCC development or proliferation (6,17);
however, no mutations in Foxa1/2 have been reported in
humans. Comprehensive signatures of significantly sex-affected
pathways, including immune response, cell cycle,
metabolism-related, and DNA repair and p53 signaling pathways have
been identified in numerous cancer types (10); however, sex-dependent gene expression
signatures in liver cancer remain unclear (18).
The goal of the present study was to identify
differences in the transcription of specific genes identified in
the tumors of male and female subjects using bioinformatic tools
(Fig. 1) (19). Microarray technology and bioinformatic
analysis have been increasing regarded as a useful approach to
detect differentially expressed genes (DEGs) and underlying
molecular functional pathways involved in tumor carcinogenesis and
progression (20). At present, no
detailed bioinformatic analysis has been performed on number of
microarrays of liver cancer and non-cancerous tissues which have
been uploaded to the Gene Expression Omnibus (GEO) (20,21).
Therefore, in the present study, Gene Ontology (GO) analysis, Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment, and
protein-protein interaction (PPI) network and survival analyses
were performed systematically to characterize gene expression
profiles downloaded from GEO, identify genes associated with the
effects of sex on carcinogenesis and progression, and emphasize the
importance of sex differences with respect to the available animal
or clinical trial data for HCC. Additionally, expression data were
obtained from The Cancer Genome Atlas (TCGA) to validate the
sex-biased gene expression profiles.
Materials and methods
Microarray data preparation
Gene expression profiles that provided information
concerning the sex features of subjects, including GSE19665
(22), GSE23343 (23) and GSE9843 (24) were downloaded from the GEO (http://www.ncbi.nlm.nih.gov/geo), a public
functional genomics dataset with high-throughput gene expression
data chips and microarrays (21). The
selection was based on a GeneChip Human Genome U133 Plus 2.0 Array
platform (Affymetrix; Thermo Fisher Scientific, Inc.). The datasets
comprised 108 tissue samples, including normal liver samples from 4
female subjects and 13 male subjects, in addition to 28 and 63 HCC
samples from female and male subjects, respectively.
Furthermore, raw tissue mRNA expression data and
corresponding clinical information from patients with HCC were
downloaded from The Cancer Genome Atlas (TCGA; http://cancergenome.nih.gov). The selection included
22 and 28 paired samples of normal liver tissues and corresponding
HCC samples from 117 male and 245 female subjects with HCC,
respectively.
Data process in analysis and
identification of sex-biased DEGs
The raw data were preprocessed and analyzed by
utilizing the affy (version 1.54.0; http://www.bioconductor.org/packages/release/bioc/html/affy.html)
package in R (version 3.5.1). Initially, a robust multi-array
average algorithm was used to preprocess data, including background
correction, quantile normalization and calculation of gene
expression values, following conversion of raw data into an
expression matrix. Subsequently, the t-test method in the Linear
Models for Microarray Data (limma; version 3.32.10) package in R
was used to calculate the P-values and identify the DEGs between
the HCC and normal liver tissue samples (25–27).
Quantified gene expression data obtained from TCGA datasets were
normalized and analyzed via the same process using the limma
package.
The Benjamini-Hochberg method (28) was applied to calculate adjusted
P-values and the false discovery rate (FDR). Probe sets without
corresponding gene symbols or genes with >1 probe set were
removed or averaged, respectively. Only the genes with |log2 fold
change| ≥1.5 and adjusted P-value ≤0.05 were selected as DEGs.
Hierarchical clustering analysis and heatmap analysis of DEGs were
performed using the heatmap (version 1.0.0) package in R (29). VennDiagram (version 1.6.20, http://CRAN.R-project.org/package=VennDiagram), a
package to generate high-resolution Venn and Euler plots in R, was
used to construct visual analyses of the common and exclusive DEGs
in the subjects of the two different sexes.
GO and pathway enrichment analysis of
DEGs
The Database for Annotation, Visualization and
Integrated Discovery (DAVID, http://david.abcc.ncifcrf.gov/) is a public biological
resource regularly utilized for functional annotation and pathway
analysis (21). In order to fully
understand the biological themes of DEGs, GO (30,31) and
KEGG pathway enrichment analyses (32–34) were
performed using DAVID in order to examine the functions of these
DEGs. A P<0.05 and a count >2 were considered as the
threshold for significant differences. The top 5 genes derived from
functional enrichment of each GO subset and KEGG pathway in female
and male groups were depicted in a bubble diagram by the ggplot2
(version 3.1.1) package in R (35).
PPI network construction of DEGs
The STRING database (version 10.0) is an online
biological resource used to detect PPIs and identify precise
functions of proteins (36). Protein
interactions were detected using a cut-off combination threshold of
0.4 using STRING. The data were further downloaded to construct a
PPI network using Cytoscape (version 3.5.0) (37). Significant modules with stronger PPIs
were selected in the plugin Molecular Complex Detection (MCODE)
(38) using the following default
parameters: Degree cut-off ≥2, node score cut-off ≥2, K-core ≥2 and
maximum depth=100. P<0.05 was considered to indicate a
statistically significant difference.
Survival analysis of sex-biased
genes
The Kaplan-Meier Plotter database (http://kmplot.com/analysis) is handled by a PostgreSQL
server, which integrates gene expression and clinical data
simultaneously, consisting of 364 patients from GEO, European
Genome-Phenome Archive (EGA; http://ega-archive.org/) and TCGA datasets (39). This database was used to perform
survival analysis for identified DEGs based on the hazard ratio
index, the 95% confidence interval and a log-rank P<0.05.
Results
Identification of DEGs
A total of 54,675 genes were obtained from the
integrated gene chips from the GEO datasets. A total of 856 and 496
DEGs were identified in tumors from female and male subjects,
respectively. The DEGs identified in the female patients included
616 upregulated and 240 downregulated genes, whereas the DEGs
identified in the male subjects comprised 173 upregulated and 323
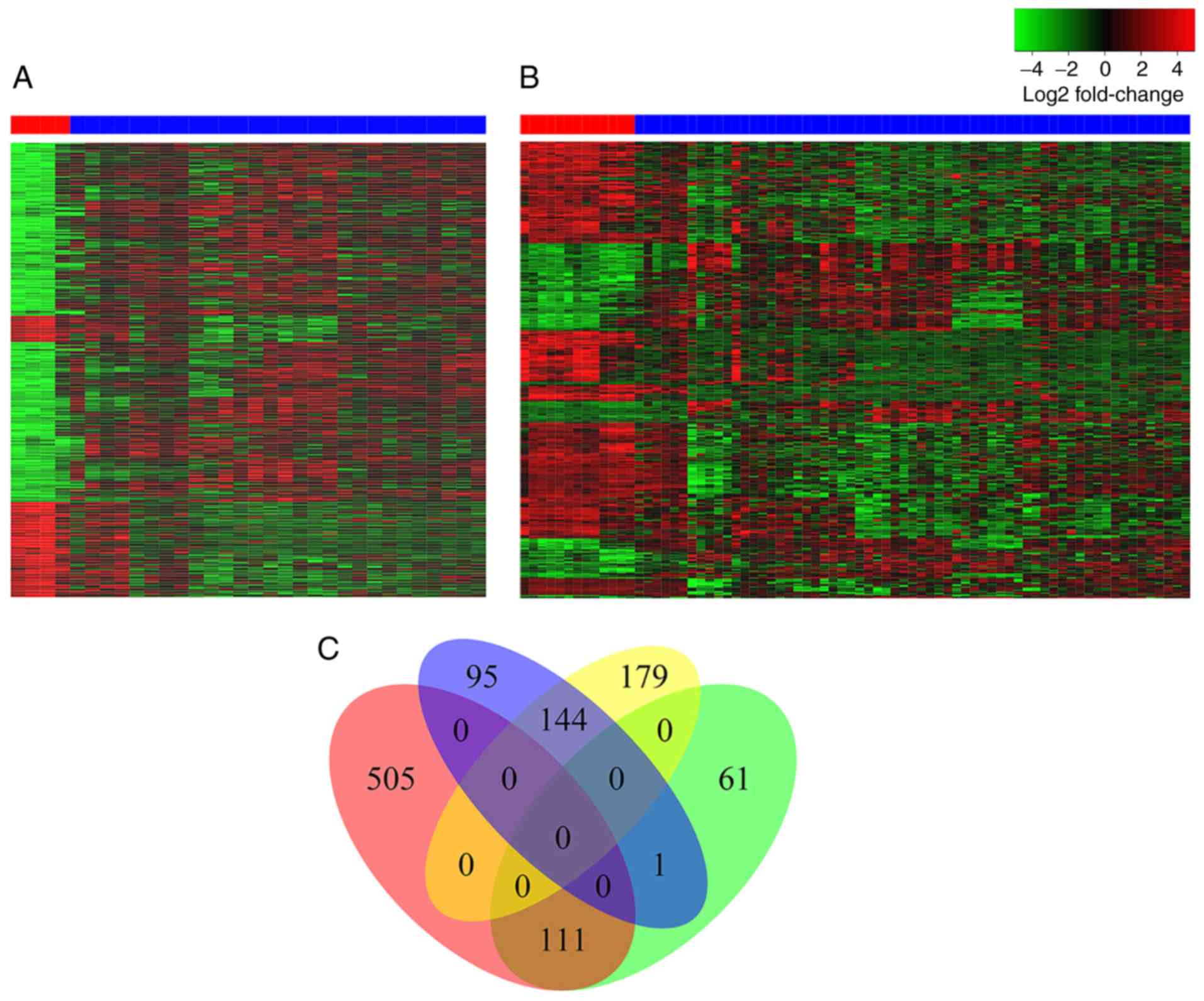
downregulated genes. The heatmaps of the bidirectional clustering
suggested significant differences between the two sexes (Fig. 2A and B). Furthermore, Venn diagram
analysis of DEG characterization between female and male subjects
(Fig. 2C) revealed that 600 DEGs were
identified only in female subjects, and 240 DEGs were found only in
male subjects. A total of 505 and 95 genes were upregulated and
downregulated, respectively, in female subjects, whereas 179 and 61
genes were downregulated and upregulated, respectively, in male
subjects. A total of 255 genes, containing 111 upregulated genes
and 144 downregulated genes, were common between female and male
patients with HCC. In addition, DEGs obtained from GEO and TCGA
samples intersected with each other according to sex, validating
the aforementioned results (Fig.
S1). The intersection contained almost all of the DEGs
identified in males, and ~40% of the DEGs identified in females
from GEO analysis, respectively.
Functional and pathway enrichment
analysis of DEGs
All the identified DEGs from the female and male
groups in the GEO datasets were uploaded separately into DAVID to
identify the functional enrichment of the representative DEGs. A
total of 220 GO terms for the female DEGs and 188 GO terms for the
male DEGs were obtained. The top 10 terms in the three GO
categories, including ‘biological process’ (BP), ‘cellular
component’ (CC) and ‘molecular function’ (MF), were presented in
Fig. 3.
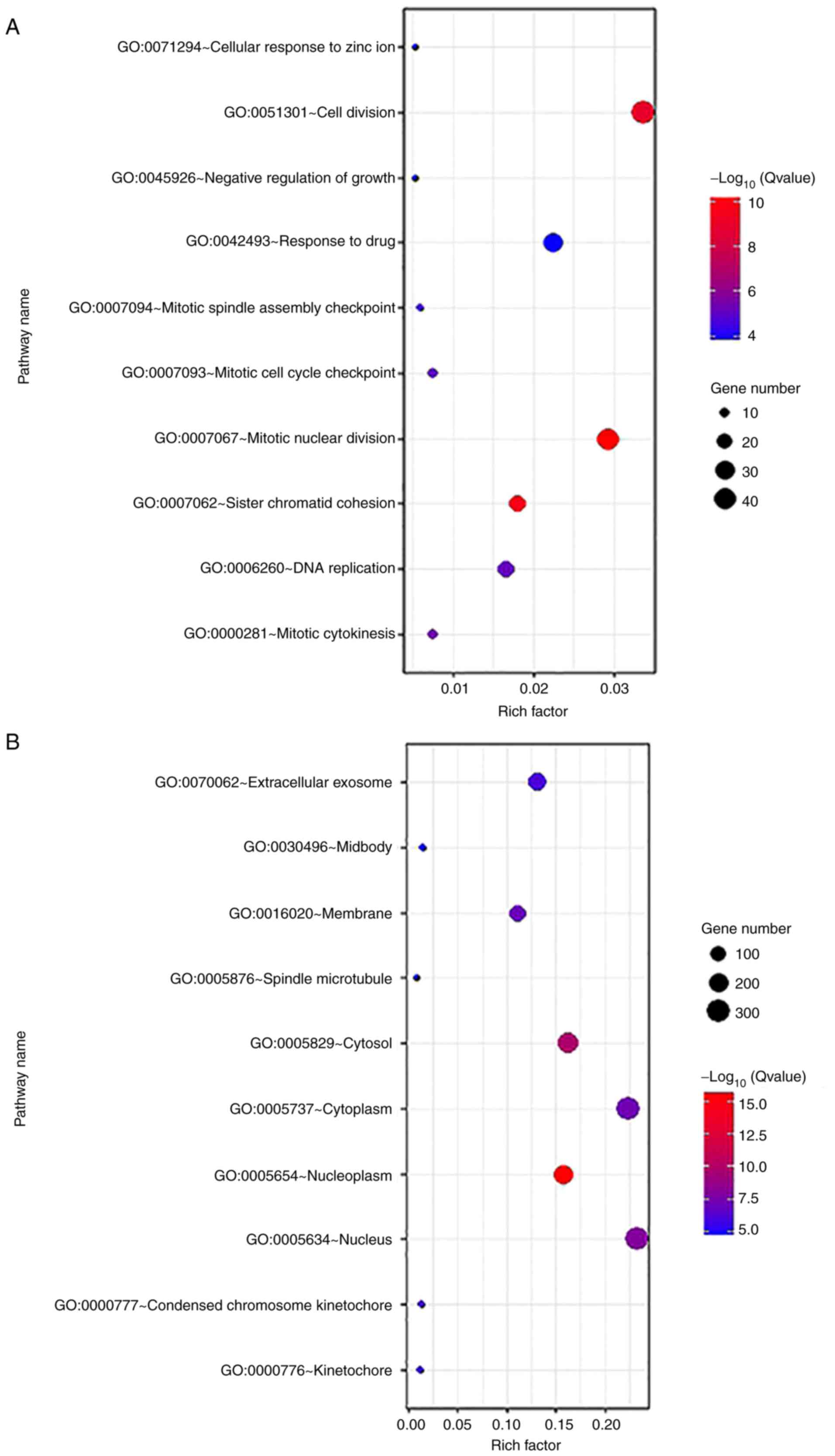 | Figure 3.Gene Ontology terms and KEGG pathway
enrichment of differentially expressed genes separated by sex. (A
and D) Biological processes, (B and E) cellular components and (C
and F) molecular functions, and (G and H) KEGG pathway analysis in
female and male cohorts, respectively. KEGG, Kyoto Encyclopedia of
Genes and Genomes; Rich factor, enrichment factor. Gene Ontology
terms and KEGG pathway enrichment of differentially expressed genes
separated by sex. (A and D) Biological processes, (B and E)
cellular components and (C and F) molecular functions, and (G and
H) KEGG pathway analysis in female and male cohorts, respectively.
KEGG, Kyoto Encyclopedia of Genes and Genomes; Rich factor,
enrichment factor. Gene Ontology terms and KEGG pathway enrichment
of differentially expressed genes separated by sex. (A and D)
Biological processes, (B and E) cellular components and (C and F)
molecular functions, and (G and H) KEGG pathway analysis in female
and male cohorts, respectively. KEGG, Kyoto Encyclopedia of Genes
and Genomes; Rich factor, enrichment factor. Gene Ontology terms
and KEGG pathway enrichment of differentially expressed genes
separated by sex. (A and D) Biological processes, (B and E)
cellular components and (C and F) molecular functions, and (G and
H) KEGG pathway analysis in female and male cohorts, respectively.
KEGG, Kyoto Encyclopedia of Genes and Genomes; Rich factor,
enrichment factor. |
The BPs of the DEGs from the female subjects were
primarily enriched in mitotic nuclear division, sister chromatid
cohesion, cell division, mitotic cytokinesis, DNA replication and
mitotic cell cycle checkpoint. The BPs of the DEGs from the male
subjects were mainly enriched in the epoxygenase P450 pathway, the
exogenous drug catabolic process, the oxidation-reduction process,
the drug metabolism process and the negative regulation of cell
growth (Fig. 3A and D).
The CC results revealed that DEGs in the female
cohorts were mainly enriched in nucleoplasm, cytosol, nucleus,
cytoplasm and membrane components. The DEGs in the tumor samples
from the male cohorts were mainly enriched in the following
modules: Extracellular region, extracellular exosome, extracellular
space, organelle membrane and blood microparticle (Fig. 3B and E).
The MF results indicated that DEGs identified in the
female cohorts were primarily enriched in protein binding, ATP
binding, poly(A) RNA binding, chromatin binding, protein
homodimerization activity and microtubule binding. The results
indicated that the DEGs in the samples from males were mainly
enriched in oxidoreductase activity, oxygen binding, heme binding,
iron ion binding and monooxygenase activity (Fig. 3C and F).
KEGG signaling pathway analysis was used to identify
the top five enriched pathways in female subjects, which included
‘Pathways in cancer’, ‘Cell cycle’, ‘Viral carcinogenesis’,
‘Biosynthesis of antibiotics’ and ‘Oocyte meiosis’, whereas in the
male subjects, the top enriched pathways included ‘Metabolic
pathways’, ‘PI3K-Akt signaling pathway’, ‘Cell cycle’, ‘Retinol
metabolism’, ‘Transcriptional dysregulation in cancer’ and ‘Drug
metabolism-cytochrome P450’ (Fig. 3G and
H).
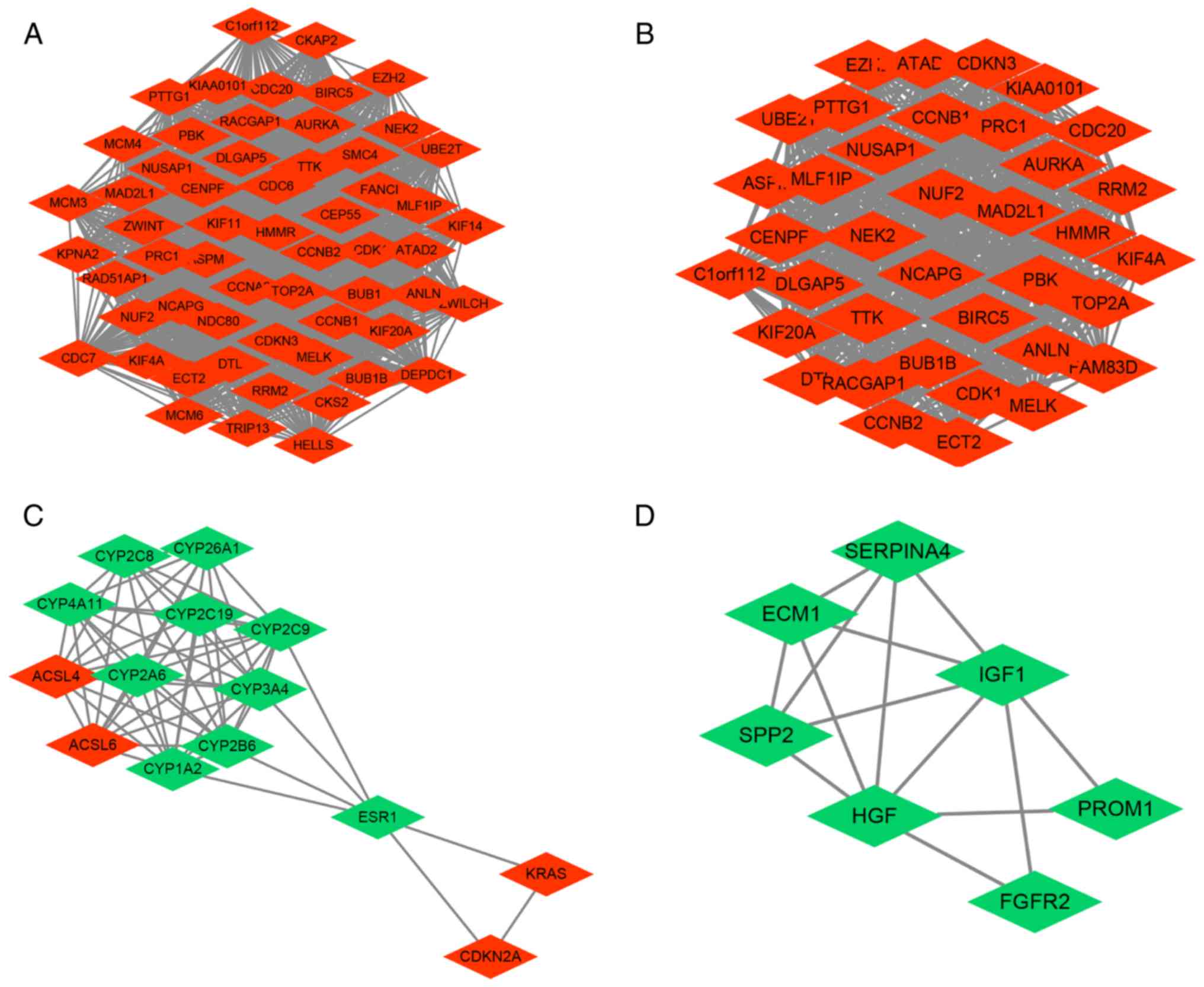
PPI network analysis of
DEGs-significant modules and core genes in network
DEGs were imported into STRING to analyze the PPIs
between the various gene targets. PPI networks were constructed in
Cytoscape (Fig. 4). Common hub genes
between the two sexes were selected with a cut-off degree >10,
and the ranked top 15 female DEGs included TOP2A, CDK1, GAPDH,
CCNB1, ACLY, CCNB2, BIRC5, NDC80, CCNA2, CDC20, CDKN3, MAD2L1,
BUB1, AURKA and KIF11. The top ranked male DEGs
comprised TOP2A, CDK1, CCNB1, CDKN3, BIRC5, AURKA, CCNB2,
MAD2L1, HMMR, TTK, EZH2, CDC20, PTTG1, RACGAP1 and NCAPG
(Table I). The hub genes in each
group were further analyzed for pathway enrichment, which exhibited
a certain degree of similarity with the top five pathways; of note,
the pathways ‘cell cycle’, ‘p53 signaling pathway’ and ‘oocyte
meiosis’ were included in the two sex groups (Table II).
 | Table I.Top hub genes in the protein-protein
interaction network with regard to their node degree between female
and male. |
Table I.
Top hub genes in the protein-protein
interaction network with regard to their node degree between female
and male.
| A, Female |
|---|
|
|---|
| Gene name | Node degree | Clustering
coefficient |
|---|
| TOP2A | 156 | 0.20239 |
| CDK1 | 114 | 0.36625 |
| GAPDH1 | 111 | 0.11433 |
| CCNB1 | 107 | 0.41474 |
| ACLY | 96 | 0.08794 |
| CCNB2 | 91 | 0.52454 |
| BIRC5 | 89 | 0.49413 |
| NDC80 | 88 | 0.54075 |
| CCNA2 | 87 | 0.53034 |
| CDC20 | 86 | 0.53844 |
| CDKN3 | 86 | 0.54118 |
| MAD2L1 | 85 | 0.57843 |
| BUB1 | 84 | 0.60614 |
| AURKA | 82 | 0.56218 |
| KIF11 | 79 | 0.63746 |
|
| B, Male |
|
| Gene
name | Node
degree | Clustering
coefficient |
|
| TOP2A | 78 | 0.26607 |
| CDK1 | 54 | 0.53948 |
| CCNB1 | 52 | 0.57919 |
| CDKN3 | 47 | 0.66605 |
| BIRC5 | 46 | 0.68213 |
| AURKA | 46 | 0.69952 |
| CCNB2 | 44 | 0.75159 |
| MAD2L1 | 44 | 0.75370 |
| HMMR | 44 | 0.72093 |
| TTK | 43 | 0.78295 |
| EZH2 | 42 | 0.60511 |
| CDC20 | 41 | 0.80610 |
| PTTG1 | 41 | 0.80488 |
| RACGAP1 | 41 | 0.83171 |
| NCAPG | 41 | 0.85854 |
| Table II.Top five enriched pathways of the hub
genes in female and male subjects. |
Table II.
Top five enriched pathways of the hub
genes in female and male subjects.
| A, Female |
|---|
|
|---|
| Term | P-value | Genes |
|---|
| cfa04110:Cell
cycle |
6.24×10−9 | CCNB1, CDK1, CCNB2,
MAD2L1, BUB1, CDC20, CCNA2 |
|
cfa04914:Progesterone-mediated oocyte
maturation |
7.00×10−8 | CCNB1, CDK1, CCNB2,
MAD2L1, BUB1, CCNA2 |
| cfa04114:Oocyte
meiosis |
1.14×10−5 | CDK1, MAD2L1, BUB1,
CDC20, AURKA |
| cfa04115:p53
signaling pathway |
3.87×10−3 | CCNB1, CDK1,
CCNB2 |
| cfa05203:Viral
carcinogenesis |
3.09×10−2 | CDK1, CDC20,
CCNA2 |
|
| B, Male |
|
| Term | P-value | Genes |
|
| bta04110:Cell
cycle |
8.73×10−11 | CCNB1, CDK1,
MAD2L1, CCNB2, TTK, CDC20, PTTG1 |
| bta04114:Oocyte
meiosis |
2.77×10−7 | CDK1, MAD2L1,
AURKA, CDC20, PTTG1 |
|
bta04914:Progesterone-mediated oocyte
maturation |
2.33×10−4 | CCNB1, CDK1,
MAD2L1, CCNB2 |
| bta04115:p53
signaling pathway |
4.50×10−3 | CCNB1, CDK1,
CCNB2 |
| bta05166:HTLV–I
infection |
5.89×10−3 | MAD2L1, CDC20,
PTTG1 |
GO and KEGG enrichment analysis of
DEGs from subgroup analysis of the PPI network
According to the cut-off criteria, several
significant modules were extracted from the male and female PPI
network using the MCODE plugin for Cytoscape (Fig. 4). Module A from the female PPI network
was constructed with 58 nodes and 1,564 edges, and contained a high
number of hub genes (Fig. 4A). Module
B in the male PPI network was constructed with 37 nodes and 651
edges (Fig. 4B). Module C in the male
PPI network was constructed with 14 nodes and 59 edges (Fig. 4C). Module D in the male PPI network
was constructed with 7 nodes and 14 edges (Fig. 4D). Notably, a large number of DEGs
that were upregulated in the female module A were also found in the
male module B. Additionally, the genes CYP3A4 and
SERPINA4 were included in the male downregulated gene
modules C and D (Fig. 4C and D).
To further explore the detailed functions of the
genes in each module, the DEGs in the PPI networks were analyzed
using GO, KEGG and STRING. Cell cycle, p53 signaling and oocyte
meiosis pathways were common in the two sexes. Additional metabolic
processes that were primarily enriched in the male sex, including
retinol metabolism, drug metabolism and linoleic acid metabolism,
were identified (Table III).
| Table III.GO and KEGG enrichment analyses of
DEGs as determined by survival analysis. |
Table III.
GO and KEGG enrichment analyses of
DEGs as determined by survival analysis.
| A, Module A |
|---|
|
|---|
| Pathway
description | P-value | Matching
proteins |
|---|
| Cell division |
2.38×10−41 | ANLN, ASPM, AURKA,
BIRC5, BUB1, BUB1B, CCNA2, CCNB1, CCNB2, CDC20, CDC6, CDC7, CDK1,
CENPF, CEP55, CKAP2, CKS2, ECT2, HELLS, KIF11 |
| Mitotic nuclear
division |
4.14×10−30 | ANLN, ASPM, BIRC5,
BUB1, CCNA2, CCNB1, CCNB2, CDC20, CDC6, CDK1, CENPF, CEP55, DLGAP5,
HELLS, KIF14, KIF4A, MAD2L1 |
| Oocyte meiosis |
2.67×10−6 | AURKA, BUB1, CCNB1,
CCNB2, CDC20, MAD2L1, PTTG1, CDK1, AURKA |
| p53 signaling
pathway |
4.17×10−2 | CCNB1, CDK1, CCNB2,
RRM2 |
|
| B, Module
B |
|
| Pathway
description | P-value | Matching
proteins |
|
| Cell cycle |
5.42×10−7 | CCNB1, CDK1, BUB1B,
CCNB2, CDC20, MAD2L1, PTTG1, TTK |
| Oocyte meiosis |
4.64×10−6 | CDK1, AURKA, CCNB1,
CCNB2, CDC20, MAD2L1, PTTG1 |
|
Progesterone-mediated oocyte
maturation |
2.25×10−2 | CCNB1, CDK1,
MAD2L1, CCNB2 |
| p53 signaling
pathway |
1.65×10−2 | CCNB1, CDK1, CCNB2,
RRM2 |
| HTLV–I
infection |
7.93×10−3 | BUB1B, CCNB2,
CDC20, MAD2L1, PTTG1 |
|
| C, Module
C |
|
| Pathway
description | P-value | Matching
proteins |
|
| Retinol
metabolism |
2.15×10−15 | CYP1A2, CYP26A1,
CYP2A6, CYP2B6, CYP2C8, CYP2C9, CYP3A4, CYP4A11 |
| Drug metabolism -
cytochrome P450 |
9.02×10−13 | CYP1A2, CYP2A6,
CYP2B6, CYP2C19, CYP2C8, CYP2C9, CYP3A4 |
| Chemical
carcinogenesis |
3.96×10−10 | CYP1A2, CYP2A6,
CYP2C19, CYP2C8, CYP2C9, CYP3A4 |
| Linoleic acid
metabolism |
4.69×10−10 | CYP1A2, CYP2C19,
CYP2C8, CYP2C9, CYP3A4 |
| Metabolic
pathways |
1.56×10−8 | ACSL4, ACSL6,
CYP1A2, CYP2A6, CYP2B6, CYP2C19, CYP2C8, CYP2C9, CYP3A4,
CYP4A11 |
|
| D, Module
D |
|
| Pathway
description | P-value | Matching
proteins |
|
| Negative regulation
of peptidase activity |
3.92×10−3 | ECM1, HGF,
SERPINA4, SPP2 |
| Negative regulation
of endopeptidase activity |
2.06×10−2 | ECM1, HGF, IGF1,
SERPINA4, SPP2 |
| Negative regulation
of macromolecule metabolic process |
4.80×10−3 | ECM1, FGFR2, HGF,
IGF1, SERPINA4, SPP2 |
| Negative regulation
of cellular metabolic process |
8.95×10−3 | ECM1, FGFR2, HGF,
IGF1, SERPINA4, SPP2 |
| Regulation of
molecular function |
4.80×10−3 | ECM1, FGFR2, HGF,
IGF1, SERPINA4, SPP2 |
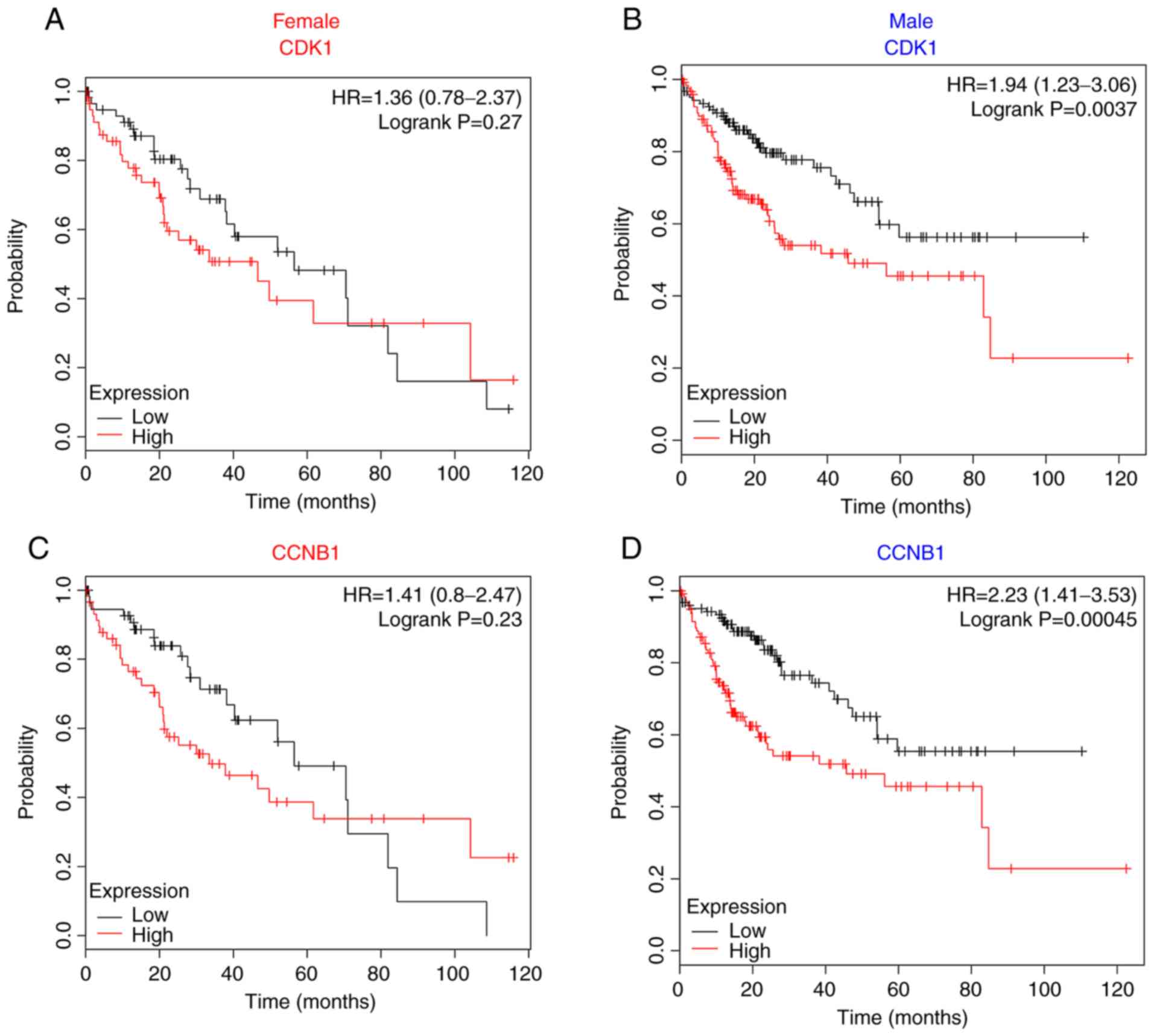
Survival analysis of candidate genes
in PPI modules
To explore the effects of each module on patient
survival, the prognostic value of DEGs for each sex was analyzed
using the Kaplan-Meier Plotter and data derived from the
combination of the GEO, EGA and TCGA databases. First, common hub
genes in modules A and B that were shared in specific common
pathways for the two sexes, such as CDK1 and CCNB1,
were analyzed. The expression of these two genes and other shared
genes were not significantly associated with the survival of female
patients (Fig. 5A and C; other genes
not shown). Conversely, significantly decreased survival was
observed for male patients with overexpression of these hub genes
(Fig. 5B and D; other genes not
shown).
Furthermore, genes identified in the subgroup PPI
analysis that were specific to the male modules C and D were
analyzed; it was revealed that CYP3A4 and SERPINA4,
which were downregulated in male patients, were significantly
associated with poor patient survival (Fig. 5F and H); however, no statistical
significance was observed for the association between the
expression of these genes and survival in female patients (Fig. 5E and G).
Discussion
HCC is a high-mortality malignancy with inadequate
current medical treatment. Males usually exhibit an increased risk
of developing HCC and poorer prognosis than females (3). The sex differences in physiological
liver gene expression have been widely studied in rodents and
humans (40–42); however, knowledge of sex-dependent
gene expression in human HCC remains scarce, and may be of
substantial biological and medical importance. In the present
study, several mRNA profile datasets from GEO and TCGA were
integrated using bioinformatic analysis methods to further identify
sex-associated characteristics of gene expression between HCC
tissues from female and male patients.
In the present study, liver samples from public
datasets were obtained, and the following sex-based gene expression
signatures were identified: A total of 858 DEGs from the female
patient group, of which 111 were upregulated, and 144 were
downregulated genes shared with the male cohort. A high number of
DEGs were identified in only one sex. The study highlighted common
molecular features for both sexes that were present in HCC tissue.
Subsequent analysis into differences in the gene function, GO
ontology and pathway analysis of DEGs between the two sexes were
performed. DEGs in the female cohort were mainly enriched in the
intracellular components, cell cycle, oocyte, mitotic nuclear
division and cell division. DEGs in the male cohort were primarily
enriched in extracellular components, metabolic pathways,
epoxygenase P450 pathway, exogenous drug catabolic processes,
oxidation-reduction processes and drug metabolism processes.
A majority of functional pathways were similar
between the two sexual groups, as demonstrated by analysis of the
common hub genes in the significant PPI networks and modules. It is
well known that the cell cycle serves an important role in
controlling cell proliferation, and that the dysregulation of the
mitotic cycle accelerates carcinogenesis and tumor progression
(43–45). Similar findings were noted in the
present study; the cell cycle module was enriched in both sexes.
Highly expressed hub genes, such as CDK1 and CCNB1,
were shared between both sexes. These common genes were enriched in
pathways such as the cell cycle pathway. Previous studies have
reported that the cell cycle is a sex-affected process (10,46,47). The
present study indicated that upregulated expression of CDK1
and CCNB1 was associated with poorer patient survival in
males, whereas no significant differences were observed in the
expression levels of these genes in female patients with HCC,
suggesting that sex may affect the functional activity of genes
involved in the cell cycle in HCC. Cyclin-dependent kinase 1 (CDK1)
is required for regulating cell cycle progression, DNA replication
and segregation, cell maturation and proliferation (48,49). The
aberrant activation of CDK1 leads to a significant
contribution to tumorigenesis by promoting cell proliferation
(49). Cyclin B1 (CCNB1) is a member
of the highly conserved cyclin family of proteins that is required
for appropriate cell cycle progression (50). CCNB1 binds to CDK and forms an active
complex known as maturation-promoting factor, which facilitates
cell entry to mitosis (48,51). Overexpression of either CDK1 or CCNB1
is associated with advanced stage, poor differentiation, and poor
prognosis in HCC (52,53). A previous study suggested that
knockdown of CDK1 is more effective in males with HCC (19), consistent with the results of the
present study. Previous studies demonstrated that 17 beta-estradiol
exerts inhibitory and apoptotic effects on SW-13 human adrenal
carcinoma cells in vitro by decreasing CCNB1 expression and
blocking G2/M entry, highlighting that the expression of CCNB1 can
be significantly decreased by estrogen (54). Therefore, high concentrations of
estradiol may aid the protection of female patients against the
development or progression of HCC by inhibiting activation of the
cell cycle and subsequently carcinogenesis. Similarly upregulated
expression levels of the cell cycle genes were detected between the
two sexes; different levels of estrogen or ER expression may affect
the activity of target genes and subsequently the prognosis of
HCC.
The liver is an important organ in metabolizing and
detoxifying dietary compounds and drugs. Sex-associated differences
in the metabolism of drugs may account for liver toxicity; it has
been shown that females exhibit increased susceptibility to drugs
and associated adverse effects (55,56). In
the present study, the findings indicated that DEGs in drug and
metabolic pathways were enriched in the tumors of the male
subjects. The majority of these genes were downregulated,
suggesting that metabolic pathways, the epoxygenase P450 pathway,
the exogenous drug catabolic process, the oxidation-reduction
process, the oxidoreductase activity and drug metabolism processes
were primarily downregulated in the tumors of male patients with
HCC. Previous studies reported that oxidoreductase activity is an
important component of antioxidant defenses, and that it acts as a
tumor suppressor that is frequently altered in tumors (57,58). The
relationship between the expression of candidate genes in
significant modules and patient survival was investigated.
Downregulation of the CYP3A4 gene was a significant
predictor of male patient survival. Cytochrome P450 3A4 (CYP3A4) is
mainly expressed in the liver and intestine, and is involved in the
metabolic activation and metabolism of several pro-carcinogens
(59,60). Sex-associated differences were
observed in CYP3A4 enzyme activity towards different substrates
with respect to their metabolic clearance (55). This enzyme exhibits higher activity in
women than in men, and may be regulated by steroid hormones
(55). A previous study reported that
the downregulation of CYP3A4 may be associated with lower metabolic
clearance of various procarcinogens, leading to higher accumulation
of these compounds in the body (55);
as a result, carcinogenesis is facilitated in patients with low
levels of the CYP3A4 gene (55,59).
Serpin family A member 4 (SERPINA4), also known as kallistatin, is
a novel antiangiogenesis protein, which exerts various effects on
inflammation, angiogenesis and tumor growth (61,62).
Previous studies reported that SERPINA4 suppressed tumor growth by
directly inhibiting cancer cell proliferation, migration and
invasion, and by adjusting cancer cell signaling in various
malignant tumors, including lung and breast cancers, and HCC
(61–63). Kallistatin levels were positively
correlated with free androgen index in women with polycystic ovary
syndrome (64). In the present study,
SERPINA4 was included in the analysis of the enrichment pathways
and was associated with negative regulation in the metabolic
process component. These findings may explain how low
SERPINA4 gene expression in male patients with HCC may lead
to poor survival.
Comparison of the gene expression levels between the
two sexes raises questions regarding the mechanisms that confer
additional protection in one sex compared with the other. The
differences in physiology between the two sexes result primarily
from the combination of the cell-autonomous effects of sex
chromosomes, and from the activation effects of male and female sex
hormones on their receptors (65).
Previous studies revealed that the cell cycle and metabolic
pathways are partially affected by steroid hormones and their
receptors (55,66). The candidate genes or pathways
identified in the present study require further investigate to
assess their potential associations with estrogen or androgen and
their receptors in an animal model of liver cancer.
Human sex-biased differences in hepatic genes at the
genetic, regulatory and functional level can be associated with the
hepatic disease state. Zhang et al (67) evaluated microarray data from liver
samples from Western Europeans following the removal of primary
liver tumors. They reported that significant sex-biased genes could
affect diverse physiological functions, such as zinc finger
clusters and lysine demethylase for epigenetic modification, and
CYP3A for hepatic drug metabolism. Yuan et al (10) performed a comprehensive analysis of
molecular differences with respect to sex across 13 cancer types
from TCGA. Numerous sex-biased genes were enriched in sex
chromosomes and accompanied with sex-biased DNA methylation. These
genes were associated with the immune response, cell cycle,
metabolism-pathways, DNA repair and p53 pathways. Ma et al
(19) analyzed DEGs between
neoplastic and normal tissue for each sex in HCC samples from TCGA.
They reported that pathways associated with lipid metabolism were
only significantly dysregulated in male subjects. The present study
contained not only TCGA samples, but also GEO expression profiles
in order to analyze different gene expression between male and
female patients with respect to tissue (neoplastic vs. normal). The
analyzed data included expression profiles from Asians and
Europeans. Sex-biased genes that were, consistent with previous
findings, associated with drug and metabolic pathways, as well as
shared genes enriched in cell cycle and p53 pathways, were
associated with patient prognosis in a sex-dependent manner.
The present study exhibited several limitations.
First, the lack of clinical information for patients downloaded
from GEO prevents adjustments for confounding factors, such as age,
disease state, tumor purity or survival time, and may reduce the
observation power. It should be noted that the bias of tumor and
patient characteristics between male and female patients for which
samples were downloaded from TCGA in the present study were not
significantly different to those observed in a previous study
(10). Second, the normal liver
tissues utilized in the present study were primarily obtained from
either non-tumor-bearing donors or liver samples adjacent to
tumors. The tissues may consist of distinct cell types to
corresponding tumor tissues, potentially confound comparison
between normal and tumor samples. Additional patients were enrolled
from TCGA to confirm the results from the GEO analysis; however,
the majority of the tissues were obtained from patients with
primary HCC tumors removed. Furthermore, the small number of normal
tissues used in the study further sensitizes the study to potential
confounds, limiting the detection power. Therefore, future studies
into this topic will require rigorous analyses of large patient
cohorts with greater control of clinical confounding variables.
Third, the presented results were entirely based on microarray
data. It remains to be established to what extent the observed sex
differences in mRNA expression would contribute to sex differences
at the protein level and the levels of biological activity in
vivo. Finally, our study did not explore the sex differences in
gene expression signatures and function derived from non-RNA-based
mechanisms, such as translational regulation, protein function and
post-translation modifications. Associations of sex-determined
factors, such as sex chromosomes, sex hormones and their activation
receptors, with sex-biased gene expression characteristics require
further elucidation. In addition, the impact of sex-biased somatic
alterations on the etiology, progression, treatment and
personalized therapy of cancer should be considered. In summary,
the present findings in sex-dependent DEGs and enriched functional
pathways exhibited a certain degree of consistency with previous
studies; however, the aforementioned limitations prevented a full
characterization of sex-biased gene expression.
In conclusion, the present study screened sex
dimorphism signatures of gene expression, gene functions and
pathways in HCC using computational bioinformatic methods. Genes
associated with the cell cycle and/or with the metabolic pathways
were identified as dysregulated in both sexes; however, these genes
exhibited distinct prognostic values for male and female patients
with HCC. The differences in physiology noted between the two
different sexes may account for the differences in the expression
of genes, including those involved in oncogenesis and in liver
cancer progression. Further studies into sex-biased DEGs in liver
cancer at the genetic, regulatory and functional level should be
conducted in animal models of HCC, or in larger independent cohorts
with increased controlling for potential confounding factors, in
order to further increase understanding of the role of these genes
in hepatic disease states and therapeutic strategies.
Supplementary Material
Supporting Data
Acknowledgements
We thank Professor Tianyan Chen and Dr Xiude Fan
(Department of Infectious Diseases and Institution of Hepatology,
First Affiliated Hospital, School of Medicine, Xi'an Jiaotong
University) for technical and writing assistance.
Funding
The present study was financially supported by the
National 13th Five-Year Special Grand Project for Infectious
Disease of China (grant no. 2017ZX10202203-007-009).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors were involved in the conception and
design of the study. YCW and NJY collected all data. YCW and YLF
analyzed and interpreted the data. YCW and ZT drafted the
manuscript. All authors assisted in revision of the manuscript of
the paper. All authors have approved the final draft of the
manuscript.
Ethical approval and consent to
participate
The present article does not include studies with
human participants or animals performed by any of the authors.
Patient consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing
interests.
References
|
1
|
Lagranha CJ, Silva TLA, Silva SCA, Braz
GRF, da Silva AI, Fernandes MP and Sellitti DF: Protective effects
of estrogen against cardiovascular disease mediated via oxidative
stress in the brain. Life Sci. 192:190–198. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li R, Cui J and Shen Y: Brain sex matters:
Estrogen in cognition and Alzheimer's disease. Mol Cell Endocrinol.
389:13–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dorak MT and Karpuzoglu E: Gender
differences in cancer susceptibility: An inadequately addressed
issue. Front Genet. 3:2682012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Institute of Medicine (US) Committee on
Understanding the Biology of Sex and Gender Differences, .
Exploring the Biological Contributions to Human Health: Does Sex
Matter? Wizemann TM and Pardue ML: National Academies Press (US);
Washington, DC: 2001,
|
|
6
|
Li Z, Tuteja G, Schug J and Kaestner KH:
Foxa1 and Foxa2 are essential for sexual dimorphism in liver
cancer. Cell. 148:72–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Molife R, Lorigan P and MacNeil S: Gender
and survival in malignant tumours. Cancer Treat Rev. 27:201–209.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273.e1. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ruggieri A, Barbati C and Malorni W:
Cellular and molecular mechanisms involved in hepatocellular
carcinoma gender disparity. Int J Cancer. 127:499–504. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan Y, Liu L, Chen H, Wang Y, Xu Y, Mao
H, Li J, Mills GB, Shu Y, Li L and Liang H: Comprehensive
characterization of molecular differences in cancer between male
and female patients. Cancer Cell. 29:711–722. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Villa E, Baldini GM, Pasquinelli C,
Melegari M, Cariani E, Di Chirico G and Manenti F: Risk factors for
hepatocellular carcinoma in Italy. Male sex, hepatitis B virus,
non-A non-B infection, and alcohol. Cancer. 62:611–615. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee CM, Lu SN, Changchien CS, Yeh CT, Hsu
TT, Tang JH, Wang JH, Lin DY, Chen CL and Chen WJ: Age, gender, and
local geographic variations of viral etiology of hepatocellular
carcinoma in a hyperendemic area for hepatitis B virus infection.
Cancer. 86:1143–1150. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
El-Serag HB: Hepatocellular carcinoma and
hepatitis C in the United States. Hepatology 36 (5 Suppl 1).
S74–S83. 2002.
|
|
14
|
Iyer JK, Kalra M, Kaul A, Payton ME and
Kaul R: Estrogen receptor expression in chronic hepatitis C and
hepatocellular carcinoma pathogenesis. World J Gastroenterol.
23:6802–6816. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hassan MM, Botrus G, Abdel-Wahab R, Wolff
RA, Li D, Tweardy D, Phan AT, Hawk E, Javle M, Lee JS, et al:
Estrogen replacement reduces risk and increases survival times of
women with hepatocellular carcinoma. Clin Gastroenterol Hepatol.
15:1791–1799. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Naugler WE, Sakurai T, Kim S, Maeda S, Kim
K, Elsharkawy AM and Karin M: Gender disparity in liver cancer due
to sex differences in MyD88-dependent IL-6 production. Science.
317:121–124. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao Y and Li Z: Interplay of estrogen
receptors and FOXA factors in the liver cancer. Mol Cell
Endocrinol. 418:334–339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng B, Zhu YJ, Wang HY and Chen L:
Gender disparity in hepatocellular carcinoma (HCC): Multiple
underlying mechanisms. Sci China Life Sci. 60:575–584. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma J, Malladi S and Beck AH: Systematic
analysis of sex-linked molecular alterations and therapies in
cancer. Sci Rep. 6:191192016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Angarica VE and Del Sol A: Bioinformatics
tools for genome-wide epigenetic research. Adv Exp Med Biol.
978:489–512. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Edgar R, Domrachev M and Lash AE: Gene
expression omnibus: NCBI gene expression and hybridization array
data repository. Nucleic Acids Res. 30:207–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Deng YB, Nagae G, Midorikawa Y, Yagi K,
Tsutsumi S, Yamamoto S, Hasegawa K, Kokudo N, Aburatani H and
Kaneda A: Identification of genes preferentially methylated in
hepatitis C virus-related hepatocellular carcinoma. Cancer Sci.
101:1501–1510. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Misu H, Takamura T, Takayama H, Hayashi H,
Matsuzawa-Nagata N, Kurita S, Ishikura K, Ando H, Takeshita Y, Ota
T, et al: A liver-derived secretory protein, selenoprotein P,
causes insulin resistance. Cell Metab. 12:483–495. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chiang DY, Villanueva A, Hoshida Y, Peix
J, Newell P, Minguez B, LeBlanc AC, Donovan DJ, Thung SN, Solé M,
et al: Focal gains of VEGFA and molecular classification of
hepatocellular carcinoma. Cancer Res. 68:6779–6788. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Diboun I, Wernisch L, Orengo CA and
Koltzenburg M: Microarray analysis after RNA amplification can
detect pronounced differences in gene expression using limma. BMC
Genomics. 7:2522006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc Ser B (Methodological). 57:289–300.
1995.
|
|
29
|
Perry M: Heatmaps: Flexible heatmaps for
functional genomics and sequence features. R package version 1.0.0.
https://rdrr.io/bioc/heatmaps/
|
|
30
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
The Gene Ontology Consortium, . The Gene
Ontology Resource: 20 years and still GOing strong. Nucleic Acids
Res. 47:D330–d338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kanehisa M, Sato Y, Furumichi M, Morishima
K and Tanabe M: New approach for understanding genome variations in
KEGG. Nucleic Acids Res. 47:D590–D595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wickham H.: ggplot2: Elegant graphics for
data analysis. Springer-Verlag; New York, NY: 2016
|
|
36
|
Snel B, Lehmann G, Bork P and Huynen MA:
STRING: A web-server to retrieve and display the repeatedly
occurring neighbourhood of a gene. Nucleic Acids Res. 28:3442–3444.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Menyhart O, Nagy A and Gyorffy B:
Determining consistent prognostic biomarkers of overall survival
and vascular invasion in hepatocellular carcinoma. R Soc Open Sci.
5:1810062018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Delongchamp RR, Velasco C, Dial S and
Harris AJ: Genome-wide estimation of gender differences in the gene
expression of human livers: Statistical design and analysis. BMC
Bioinformatics. 6 (Suppl 2):S132005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mayne BT, Bianco-Miotto T, Buckberry S,
Breen J, Clifton V, Shoubridge C and Roberts CT: Large scale gene
expression meta-analysis reveals tissue-specific, sex-biased gene
expression in humans. Front Genet. 7:1832016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Weng Y, DiRusso CC, Reilly AA, Black PN
and Ding X: Hepatic gene expression changes in mouse models with
liver-specific deletion or global suppression of the
NADPH-cytochrome P450 reductase gene. Mechanistic implications for
the regulation of microsomal cytochrome P450 and the fatty liver
phenotype. J Biol Chem. 280:31686–31698. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cai Z and Liu Q: Cell cycle regulation in
treatment of breast cancer. Adv Exp Med Biol. 1026:251–270. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Otto T and Sicinski P: Cell cycle proteins
as promising targets in cancer therapy. Nat Rev Cancer. 17:93–115.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Brandmaier A, Hou SQ and Shen WH: Cell
cycle control by PTEN. J Mol Biol. 429:2265–2277. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Berger C, Qian Y and Chen X: The
p53-estrogen receptor loop in cancer. Curr Mol Med. 13:1229–1240.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Feng H, Cheng AS, Tsang DP, Li MS, Go MY,
Cheung YS, Zhao GJ, Ng SS, Lin MC, Yu J, et al: Cell cycle-related
kinase is a direct androgen receptor-regulated gene that drives
β-catenin/T cell factor-dependent hepatocarcinogenesis. J Clin
Invest. 121:3159–3175. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Brown NR, Korolchuk S, Martin MP, Stanley
WA, Moukhametzianov R, Noble MEM and Endicott JA: CDK1 structures
reveal conserved and unique features of the essential cell cycle
CDK. Nat Commun. 6:67692015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liu P, Kao TP and Huang H: CDK1 promotes
cell proliferation and survival via phosphorylation and inhibition
of FOXO1 transcription factor. Oncogene. 27:4733–4744. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ding K, Li W, Zou Z, Zou X and Wang C:
CCNB1 is a prognostic biomarker for ER+ breast cancer. Med
Hypotheses. 83:359–364. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bednarek K, Kiwerska K, Szaumkessel M,
Bodnar M, Kostrzewska-Poczekaj M, Marszalek A, Janiszewska J,
Bartochowska A, Jackowska J, Wierzbicka M, et al: Recurrent CDK1
overexpression in laryngeal squamous cell carcinoma. Tumour Biol.
37:11115–11126. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Spaziani A, Alisi A, Sanna D and Balsano
C: Role of p38 MAPK and RNA-dependent protein kinase (PKR) in
hepatitis C virus core-dependent nuclear delocalization of cyclin
B1. J Biol Chem. 281:10983–10989. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Park TJ, Kim JY, Oh SP, Kang SY, Kim BW,
Wang HJ, Song KY, Kim HC and Lim IK: TIS21 negatively regulates
hepatocarcinogenesis by disruption of cyclin B1-Forkhead box M1
regulation loop. Hepatology. 47:1533–1543. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Brown JW, Prieto LM, Perez-Stable C,
Montoya M, Cappell S and Fishman LM: Estrogen and progesterone
lower cyclin B1 AND D1 expression, block cell cycle in G2/M, and
trigger apoptosis in human adrenal carcinoma cell cultures. Horm
Metab Res. 40:306–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tanaka E: Gender-related differences in
pharmacokinetics and their clinical significance. J Clin Pharm
Ther. 24:339–346. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ueno K and Sato H: Sex-related differences
in pharmacokinetics and pharmacodynamics of anti-hypertensive
drugs. Hypertens Res. 35:245–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Abu-Remaileh M and Aqeilan RI: The tumor
suppressor WW domain-containing oxidoreductase modulates cell
metabolism. Exp Biol Med (Maywood). 240:345–350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Roszczenko P, Radomska KA, Wywial E,
Collet JF and Jagusztyn-Krynicka EK: A novel insight into the
oxidoreductase activity of Helicobacter pylori HP0231 protein. PLoS
One. 7:e465632012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ashida R, Okamura Y, Ohshima K, Kakuda Y,
Uesaka K, Sugiura T, Ito T, Yamamoto Y, Sugino T, Urakami K, et al:
CYP3A4 gene is a novel biomarker for predicting a poor prognosis in
hepatocellular carcinoma. Cancer Genomics Proteomics. 14:445–453.
2017.PubMed/NCBI
|
|
60
|
Rodriguez-Antona C and Ingelman-Sundberg
M: Cytochrome P450 pharmacogenetics and cancer. Oncogene.
25:1679–1691. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sun HM, Mi YS, Yu FD, Han Y, Liu XS, Lu S,
Zhang Y, Zhao SL, Ye L, Liu TT, et al: SERPINA4 is a novel
independent prognostic indicator and a potential therapeutic target
for colorectal cancer. Am J Cancer Res. 6:1636–1649.
2016.PubMed/NCBI
|
|
62
|
Frühbeck G, Gómez-Ambrosi J, Rodríguez A,
Ramírez B, Valentí V, Moncada R, Becerril S, Unamuno X, Silva C,
Salvador J and Catalán V: Novel protective role of kallistatin in
obesity by limiting adipose tissue low grade inflammation and
oxidative stress. Metabolism. 87:123–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Nallagangula KS, Shashidhar KN, Lakshmaiah
V and Muninarayana C: Cirrhosis of liver: Interference of serpins
in quantification of SERPINA4-a preliminary study. Pract Lab Med.
9:53–57. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Calan M, Guler A, Unal Kocabas G, Alarslan
P, Bicer M, Imamoglu C, Yuksel A, Bozkaya G and Bilgir O:
Association of kallistatin with carotid intima-media thickness in
women with polycystic ovary syndrome. Minerva Endocrinol.
43:236–245. 2018.PubMed/NCBI
|
|
65
|
Mauvais-Jarvis F, Arnold AP and Reue K: A
guide for the design of pre-clinical studies on sex differences in
metabolism. Cell Metab. 25:1216–1230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kim HI, Lim H and Moon A: Sex differences
in cancer: Epidemiology, genetics and therapy. Biomol Ther.
26:335–342. 2018. View Article : Google Scholar
|
|
67
|
Zhang Y, Klein K, Sugathan A, Nassery N,
Dombkowski A, Zanger UM and Waxman DJ: Transcriptional profiling of
human liver identifies sex-biased genes associated with polygenic
dyslipidemia and coronary artery disease. PLoS One. 6:e235062011.
View Article : Google Scholar : PubMed/NCBI
|