Introduction
Cervical cancer (CC) is the second leading cause of
cancer- associated mortalities among women aged 20–39 years old in
USA in 2015, and it should receive attention due to the requirement
for preventing the disease in young women (1). Chronic infection with human
papillomavirus (HPV) can result in cervical intraepithelial
neoplasia and deterioration of the cervix (2). Expression of the viral proteins E6 and
E7 is involved in CC, and the former can enhance the degradation of
the p53 tumor suppressor protein, whereas the latter serves key
roles in the HPV life cycle and carcinogenic transformation
(3,4).
The comprehensive mechanism underlying CC still remains to be fully
clarified.
Casiopeina II-gly (Cas-II-gly) is a novel
chemotherapeutic compound based on a copper core, and it is an
efficient antitumor agent for carcinoma cell lines in in
vitro experiments (5). A number
of studies indicated that Cas-II-gly could involve interaction with
DNA, via intercalation or direct adduct formation (6), or poisoning and oxidation of
mitochondrion (7). Additionally, a
previous report demonstrated that Cas-II-gly can significantly
suppress HeLa cell proliferation (8);
however, its mechanism remains unclear. The present study focuses
on the effects of Cas-II-gly on gene expression and cellular
functions in HeLa and CaSki CC cells.
Microarray analysis is an effective method to
examine specific genes and their properties among thousands of
samples (9–11). Almost all differentially-expressed
genes (DEGs) are divided into three major categories based on Gene
Ontology (GO) annotations: Biological processes; cellular
components; and molecular functions (12). A number of studies have been performed
using this approach. For example, using a microRNA (miRNA)
microarray, Xia et al (13)
determined that miR-3156-3p expression was significantly suppressed
in CC cells, compared with the control cells. In another study, Li
et al (14) derived data from
The Cancer Genome Atlas database and identified 11 upregulated and
28 downregulated DEGs in CC, as well as 5 significantly-enriched
pathways being validated in the ClueGO and DAVID databases,
including the systemic lupus erythematosus pathway (15). Liang et al (16) employed functional enrichment analysis
to demonstrate that the target genes of three miRNAs
(miRNA-145/200c/218-1) may participate in diverse pathways in CC,
including the mitogen-activated protein kinase (MAPK), adenosine
5′-monophosphate-activated protein kinase, focal adhesion,
cGMP-protein kinase G, Wnt and mechanistic target of rapamycin
kinase signaling pathways, which have significant impacts on CC
cells (16). In another study, Liang
et al (17) determined that
miR-101 (screened out using 3 bioinformatics web sites) suppressed
the G1-to-S phase transition in CC cells. All of these examples
demonstrate that microarray analysis is a powerful and effective
method that can help to identify the underlying genes and signaling
pathways responsible for diseases. However, the underlying
regulatory networks involve greater complexity that remains to be
comprehensively dissected. The aim of the present study was to
analyze the DEGs between untreated CC cells and CC cells treated
with Cas-II-gly.
Long non-coding (lnc)RNAs regulate the growth,
invasion and deterioration of tumors (18). The results of Fan et al
(19) demonstrated that upregulation
of lnc-interleukin 7 receptor could reduce tumor size and limit
lymph node metastasis in CC tissue. Another study indicated that
transfection of CC cells with a small interfering (si)RNA against
prostate cancer associated transcript-1 (PCAT-1) lncRNA could
significantly suppress PCAT-1 expression, while PCAT-1
overexpression could limit the proliferation, metastasis and
invasion of CC cells (20). miRNAs
are a class of 18–25 nucleotide-long RNA molecules that are
involved in maintaining normal cellular function by regulating mRNA
transcription (21). For instance,
miR-519d-3p overexpression inhibited cell proliferation and cell
cycle progression, while it enhanced apoptosis in HeLa cells by
targeting hypoxia inducible factor 1α (22). In another study, Cai et al
(23) determined that by targeting
transforming growth factor β receptor 2, miR-17-5p had a positive
correlation with CC proliferation and metastasis. Recently, the
lncRNA-miRNA-mRNA axis was assigned a broad role in the regulatory
mechanisms of disease-associated genes, including genes involved in
CC (12,24–27).
Furthermore, the lncRNA-miRNA-mRNA axis also serves an important
role in regulating genes involved in CC sensitivity to
chemotherapy, including cancer susceptibility candidate 2-miR-21
and phosphatase and tensin homolog (28). Therefore, the present study also aimed
to clarify the genetic basis of CC, which is essential for
investigating the underlying mechanism of Cas-II-gly in CC.
The Wnt signaling pathway is a complex regulatory
network that participates in cell differentiation, proliferation,
migration, polarity, processes that are crucial for embryonic
development, tissue regeneration, stem cell maintenance and
homeostasis (29). For example, it
has been reported that the Wnt/β-catenin/Notch and phosphoinositide
3-kinase (PI3K)/protein kinase B (Akt) pathways are deregulated in
HPV-induced cancer (30). The results
from Li et al (31) indicated
that by targeting the APC gene, miR-182 activated the
canonical Wnt signaling pathway and increased CC cell
proliferation. Those observations provided a molecular-level
perspective on CC cell regulation. However, this network, which
includes miRNAs, lncRNAs and the Wnt pathway, is complex and
further studies are required to investigate the regulatory
associations between the lncRNA-miRNA axis and the Wnt signaling
pathway.
Materials and methods
Dataset
The data were harvested from the Gene Expression
Omnibus database accession GSE41827 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE41827).
A total of 6 samples were divided into three control HeLa groups
and three Cas-II-gly treated HeLa groups (3 samples/group) in the
microarray data (7). In the dataset,
HeLa cells in the treatment group were exposed to Cas-II-gly (40
mM) in 96-well microplates for 6 h at 37°C and maintained at 37°C
in 5% CO2 under sterile conditions in Dulbecco's
modified Eagle's medium supplemented with 10% fetal bovine serum
(FBS), along with an untreated control group. The R (version 3.4.1)
Limma package (32) was used to
analyze the DEGs. |log2FC|>1 and P<0.05 served as
the standard to identify DEGs. The probes of the DEGs were then
replaced by the Gene Symbol listed in GPL570 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GPL570).
Functional enrichment analysis
The GO (33) analysis
consisted of the cellular composition, molecular function and
biological process. The ClusterProfiler (http://www.bioconductor.org/packages/release/bioc/html/clusterProfiler.html)
package was used to perform the GO biological process functional
enrichment analysis of the relevant DEGs, and the hypergeometric
distribution method (34) was used to
calculate the P-value. P<0.05 served as the threshold for
statistical significance. The results of ClusterProfiler enrich
analysis were visualized via the enrichplot (https://github.com/GuangchuangYu/enrichplot) and
cnetplot (34) package.
Pathway enrichment analysis
Based on the Kyoto Encyclopedia of Genes and Genomes
database, which includes systems information, genomic information
and chemical information (35), gene
set enrichment analysis was conducted via the ClusterProfiler
package to identify statistically significant signaling pathways
(P<0.05). The visualization was produced using the enrichplot
package.
Correlation network analysis among the
DEGs
To identify the correlation between mRNAs and
lncRNAs, the psych package (https://cran.rstudio.com/web/packages/psych/index.html)
was used to establish a correlation network, which was built
according to the normalized expression levels of the lncRNAs and
mRNAs. Pearson's coefficient was calculated to form the network
(36). Cutoff >0.7 and P<0.05
served as the threshold for statistical significance. The network
was visualized via the Cytoscape software (version 3.6.0;
http://www.cytoscape.org/).
miRNA target prediction
The TargetScan (http://www.targetscan.org/vert_72/) and starBase
databases (http://starbase.sysu.edu.cn/) were employed to predict
the targeting associations between miRNAs and lncRNAs/mRNAs. The
Venny tool (http://bioinfogp.cnb.csic.es/tools/venny/) was applied
to identify the common miRNAs among the predicted results.
Cell line cultivation
The CC cell lines [CaSki (cat. no. BNCC338223) and
HeLa (cat. no. BNCC337633; BeNa Culture Collection, Beijing,
China)] and the human cervical epithelial cell line HcerEpic (cat.
no. BNCC340374; BeNa Culture Collection) were maintained at 37°C in
5% CO2 under sterile conditions. The CaSki and HeLa cell
lines were cultured in CM2-1 solution, which consisted of RPMI-1640
(Sigma-Aldrich; Merck KGaA; Darmstadt, Germany) and FBS (9:1
ratio). The HcerEpic cell line was cultured in Eagle's Minimum
Essential medium (Minimum Essential medium and non-essential amino
acid; Sigma-Aldrich; Merck KGaA) with FBS (9:1 ratio).
Plasmid construction, oligonucleotides
and cell transfection
The MALAT1- and FZD2-encoding sequences were
amplified by polymerase chain reaction (PCR), according to the
subsequent protocol, and then cloned into a pcDNA3.1 vector (25 nM;
Shanghai GenePharma Co., Ltd., Shanghai, China) for overexpression.
MALAT1-siRNA (si-MALAT1; 50 nM) was used to target MALAT1 and
FZD2-siRNA (si-FZD2; 50 nM) was used to knockdown
FZD2, which were purchased from Shanghai GenePharma Co.,
Ltd. The sequences were as follows: si-MALAT1,
5′-CAGCCCGAGACTTCTGTAA-3′; and si-FZD2,
5′-CCCGACTTCACGGTCTACATGATCA-3′. The hsa-miR-17-5p mimic and
inhibitor were also purchased from Shanghai GenePharma Co., Ltd.
The oligonucleotides and constructs were transfected separately
into cells using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocols. The subsequent experimentation was
performed 24 h after transfection.
Synthesis of Cas-II-gly
The Cas-II-gly copper complex was synthesized.
Firstly, equimolar solutions of copper (II) nitrate and the
corresponding substituted diamine were mixed. Subsequently, a
previously deprotonated N-O donor was added (37). The product was consistent with the
characterization reported by Alemon-Medina et al (38) (Table
SI).
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was used to monitor the amplification of
target DNA molecules during the PCR in real time, which was used to
estimate the expression levels of the RNAs in the present study.
HeLa and CaSki cells were treated with different concentrations of
Cas-II-gly (0, 10, 20, 30 or 40 mM) for 6 h at 37°C. Subsequently,
total RNA was isolated using TRIzol® Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocols. miRNA-specific probes were designed using
the PrimerExpress® V3.0.1 software (Applied Biosystems;
Thermo Fisher Scientific, Inc.) and purchased from Integrated DNA
Technologies (Intergrated DNA Technologies, Inc., Coralville, IA,
USA). RT was performed using a Super-Script First-Strand Synthesis
kit (Invitrogen; Thermo Fisher Scientific, Inc.). qPCR was
performed using a standard TaqMan PCR kit protocol on an Applied
Biosystems 7900HT Sequence Detection System (cat. no. 4329002;
Applied Biosystems; Thermo Fisher Scientific, Inc.). The PCR
reaction mixture included the RT product, 2X TaqMan®
Universal PCR Master Mix (cat. no. 4324018), 0.2 µM
TaqMan® probe (cat. no. 4316033; both Applied
Biosystems; Thermo Fisher Scientific, Inc.), 1.5 µM forward primer
and 0.7 µM reverse primer. The thermocycling conditions for the PCR
were as follows: Preheating at 90°C for 10 min, followed by 30
cycles of 95°C for 30 sec, 55°C for 30 sec, 72°C for 60 sec, with a
final elongation step at 72°C for 10 min and 4°C on hold. Following
the completion of the reactions, the Cq values were determined
using the default threshold settings. The ratio of mRNA to lncRNA
was calculated by using the 2−∆∆Cq equation (39). The sequences are listed in Table I.
 | Table I.Primer sequences used for reverse
transcription- quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription- quantitative polymerase chain reaction.
| Genes | Primer sequences
(5′-3′) |
|---|
| MALAT1 | F:
AAAGCAAGGTCTCCCCACAAG |
|
| R:
GGTCTGTGCTAGATCAAAAGGC |
| FZD2 | F:
GCGAAGCCCTCATGAACAAG |
|
| R:
TCCGTCCTCGGAGTGGTTCT |
| miR-17-5p | F:
CAAAGTGCTTACAGTGCAGGTAG |
|
| R:
GAATACCTCGGACCCTGC |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
| GAPDH | F:
ACAACTTTGGTATCGTGGAAGG |
|
| R:
GCCATCACGCCACAGTTTC |
Western blot analysis
Western blot analysis was conducted to evaluate
protein expression levels. Following transfection in HeLa and CaSki
cells, according to the aforementioned protocol, proteins were
extracted in lysis buffer (Sigma-Aldrich; Merck KGaA), boiled for 5
min at 100°C and then cooled on ice for 5 min. Protein
concentrations were quantified by the bicinchoninic acid protein
assay. Subsequently, 30 µg total protein was resolved via 12%
SDS-PAGE and the transferred to a polyvinylidene difluoride
membrane (EMD Millipore, Billerica, MA, USA). The membranes were
blocked in 5% fat-free milk in TBST buffer (50 mM Tris-HCl, 150 mM
NaCl and 0.05% Tween-20; pH 7.6) at 4°C for 1 h. Subsequently, the
membranes were incubated with anti-FZD2 (1:2,000; cat. no.
ab94913), anti-disheveled segment polarity protein (Dvl; 1:2,000;
cat. no. ab233003), anti-glycogen synthase kinase-3β (GSK-3β;
1:5,000; cat. no. ab32391), anti-β-catenin (1:5,000; cat. no.
ab32572) and anti-GAPDH (1:2,500; cat. no. ab9485) primary
antibodies at 4°C overnight. Following three washes in TBST buffer,
the membranes were incubated with the goat anti-rabbit horseradish
peroxidase-conjugated immunoglobulin G secondary antibody (1:2,000;
cat. no. ab6721) at room temperature for 1 h and then washed three
times in TBST buffer. The blots were visualized by intense
chemiluminescence detection (GE Healthcare Life Sciences, Little
Chalfont, UK) and exposed to Kodak XAR-5-ray film (Sigma-Aldrich;
Merck KGaA). All antibodies were purchased from Abcam (Cambridge,
MA, USA).
Dual-luciferase reporter assays
For the luciferase reporter assays, HeLa cells
(3×104) without Cas-II-gly treatment were plated in a
24-well plate and then co-transfected with 400 ng pc3.1-miR-17-5p
(miR-17-5p mimics) or pcDNA3.1 empty vector (miR-NC) and 200 ng
wild-type (WT) or mutant (mut) luciferase constructs
(Sigma-Aldrich; Merck KGaA) using Lipofectamine 2000, according to
manufacturer's protocols. HeLa cells were co-transfected with:
miR-17-5p NC and MALAT1-WT; miR-17-5p mimics and MALAT1-WT;
miR-17-5p NC and MALAT1-mut; miR-17-5p mimics and MALAT1-mut;
miR-17-5p NC and FZD2-WT; miR-17-5p mimics and
FZD2-WT; miR-17-5p NC and FZD2-mut; or miR-17-5p
mimics and FZD2-mut. A Dual-Luciferase Reporter Gene Test
kit (Beyotime Institute of Biotechnology, Shanghai, China) was used
to detect the luciferase activities in the different groups. The
firefly and the Renilla luciferase reagents were added to
detect the luciferase activity in each group. Relative luciferase
activities were calculated as ratios of firefly to Renilla
luciferase activities at 36 h post-transfection.
Cell viability assays
Cell viability was assessed using an MTT assay
(Roche Diagnostics GmbH, Mannheim, Germany). Following treatment
with different Cas-II-gly concentrations or transfected with
plasmid, according to the aforementioned protocols, HeLa and CaSki
cells were plated into a 96-well plate (2×105
cells/well) and then incubated for 48 h at 37°C. Subsequently, 10
µl MTT (5 mg/ml) was added followed by incubation for 3 h at 37°C
in darkness. Following removal of the MTT solution, the
precipitated formazan was dissolved by the addition of 100 µl
dimethyl sulfoxide. After a 10-min incubation at 37°C with shaking,
the absorbance was quantified with a spectrophotometer at 560 nm
using with a microplate reader (Thermo Fisher Scientific,
Inc.).
EdU staining
A Click-iT Plus EdU Alexa Fluor 1594 Imaging kit
(Invitrogen; Thermo Fisher Scientific, Inc.) was used to measure
cell proliferation, according to the manufacturer's protocol.
Transfected HeLa and CaSki cells, according to the aforementioned
protocol, were fixed with 50 µl cold 4% formaldehyde for 30 min at
room temperature. DAPI (1:2,000) was used to stain the cell nuclei
for 30 min at room temperature, and the signal was detected using
an Olympus FLUOVIEW FV1000 confocal laser-scanning microscope
(Olympus Corporation, Tokyo, Japan) at a magnification of ×100.
Cell apoptosis assays
HeLa and CaSki cell apoptosis was measured using
fluorescein isothiocyanate (FITC)-conjugated Annexin V and
propidium iodide (PI) (BD Pharmingen; Becton-Dickinson and Company,
Franklin Lakes, NJ, USA). The cells were washed twice in cold PBS
and then resuspended in Annexin V-binding buffer (BD Pharmingen;
Becton-Dickinson and Company) at a concentration of
3×106 cells/ml. Additionally, 100 µl of the suspension
was incubated with 5 µl Annexin V-FITC and 5 µl PI in dark at 37°C
for 15 min. After gentle vortexing, the cells were incubated for 15
min at room temperature in the dark. Following the addition of 400
µl binding buffer (Sigma-Aldrich; Merck KGaA) to each tube, the
cells were analyzed using FACSAria™ III Cell Sorter flow cytometer
(BD Biosciences, San Jose, CA, USA) and analyzed using FlowJo V10
software (FlowJo LLC, Ashland, OR, USA).
Statistical analysis
All values were expressed as mean value ± standard
deviation. The data were analyzed using the GraphPad Prism v7.0
software (GraphPad Software, Inc., La Jolla, CA, USA). Unpaired
Student's t-test was used to analyze the significance between the
measurements of two independent groups and one-way analysis of
variance followed by the Tukey post hoc test was used to make
comparisons in data sets containing multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Differentially-expressed mRNAs and
lncRNAs following Cas-II-gly treatment
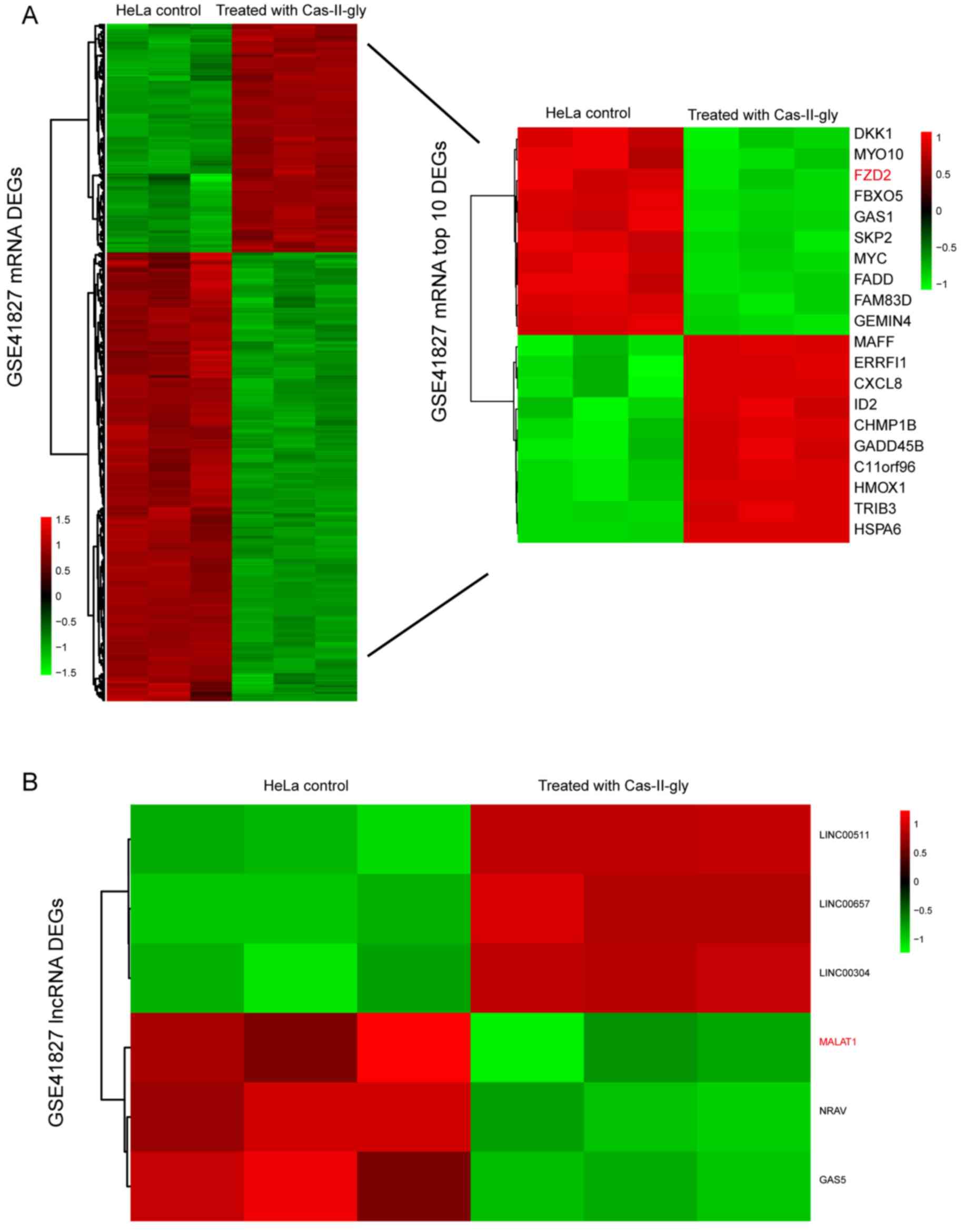
Fig. 1A demonstrates
that there were 723 differentially-expressed mRNAs, among which 244
were overexpressed and 479 were suppressed in the Cas-II-gly
treatment group. The top 10 upregulated and top 10 downregulated
mRNAs are depicted in the heatmap on the left. Compared with the
control group, FZD2 was significantly decreased in the
Cas-II-gly-treated HeLa cells (Fig.
1A). In reference to lncRNAs, Fig.
1B depicts that only 6 differentially-expressed lncRNAs were
identified, including MALAT1. MALAT1 was notably suppressed in
comparison with the control group (|log2FC|>1;
P<0.05).
Functional enrichment of DEGs
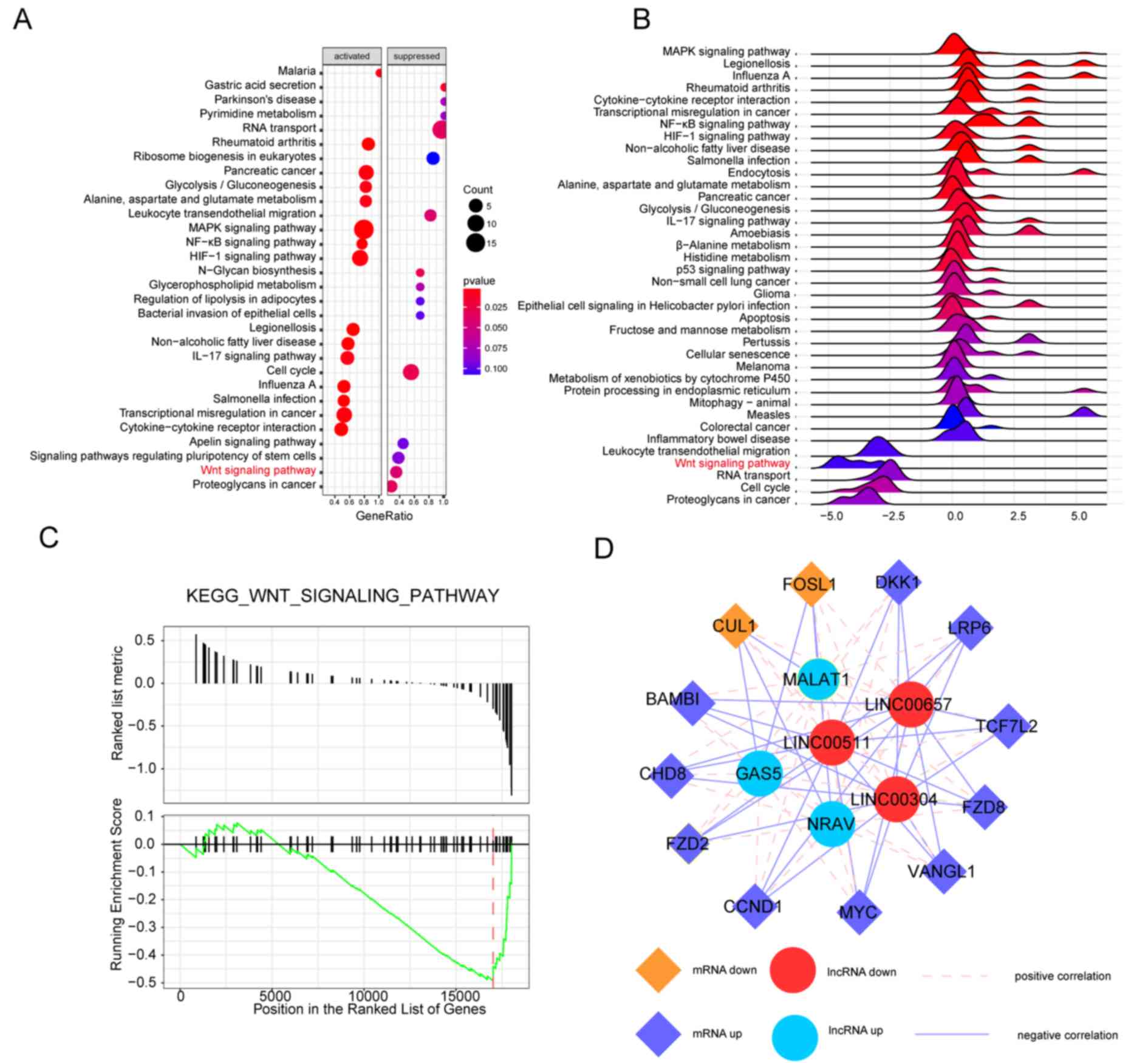
The dotplot in Fig. 2A
depicts the distribution of the 30 GO biological process terms (15
were activated and 15 were suppressed). The ‘cellular response to
heat’ term was activated in Cas-II-gly-treated HeLa cells,
indicating that Cas-II-gly may disrupt proper protein folding as a
result of nonspecific aggregation (39). The ‘cell cycle’ term, which involves
cell division and DNA replication, was suppressed in the treatment
group. The horizontal axis represents the gene ratio, which was the
result of the gene count divided by the set size. In the enrichment
map depicted in Fig. 2B, larger sets
of the enrichment results were combined into a network to gather
highly similar terms into clusters to highlight overall trends
(40). The ‘cellular response to
unfolded protein’, ‘response to unfolded protein’ and ‘cellular
response to topologically incorrect protein’ terms had intersecting
gene sets with each other. The ridgeplot in Fig. 2C depicts the exact gene set expression
of these biological process terms and the enrichment results of the
biological process terms. If the peak values are <0, then the
terms were suppressed, and the terms with peak values >0 were
activated. For instance, the ‘apoptotic mitochondrial changes’
term, which reflects the cell metabolism level, was activated in
the treatment group. In cnetplot in Fig.
2D depicts the association between the 5 biological
process-enriched terms and the DEGs, including the ‘cell division’,
‘DNA package’, ‘apoptotic mitochondrial changes’, ‘regulation of
transcription from RNA polymerase II promoter in response to
stress’ and ‘regulation of DNA-templated transcription in response
to stress’ terms.
Wnt signaling pathway is
suppressed
Fig. 3A depicts the
distribution of 15 activated and 15 suppressed signaling pathways.
The Wnt signaling pathway was notably suppressed in
Cas-II-gly-treated HeLa cells. Furthermore, the exact gene set
expression and enrichment results are depicted in the ridgeplot in
Fig. 3B. The Wnt signaling pathway
was significantly suppressed, as its peak was <0. The running
enrichment score of the Wnt signaling pathway is depicted in
Fig. 3C, which indicates the
suppression of the Wnt signaling pathway. Based on these combined
bioinformatics results, it was concluded that the Wnt signaling
pathway may participate in the Cas-II-gly dependent effects on HeLa
cells. The correlation analysis Fig.
3D depicts the association between the DEGs in the Wnt
signaling pathway and the differentially-expressed lncRNAs.
FZD2 was the upstream core gene in the Wnt signaling
pathway, which was downregulated in the treatment group. The MALAT1
lncRNA had positive correlation with FZD2.
Targeting association between
miR-17-5p and MALAT1/FZD2
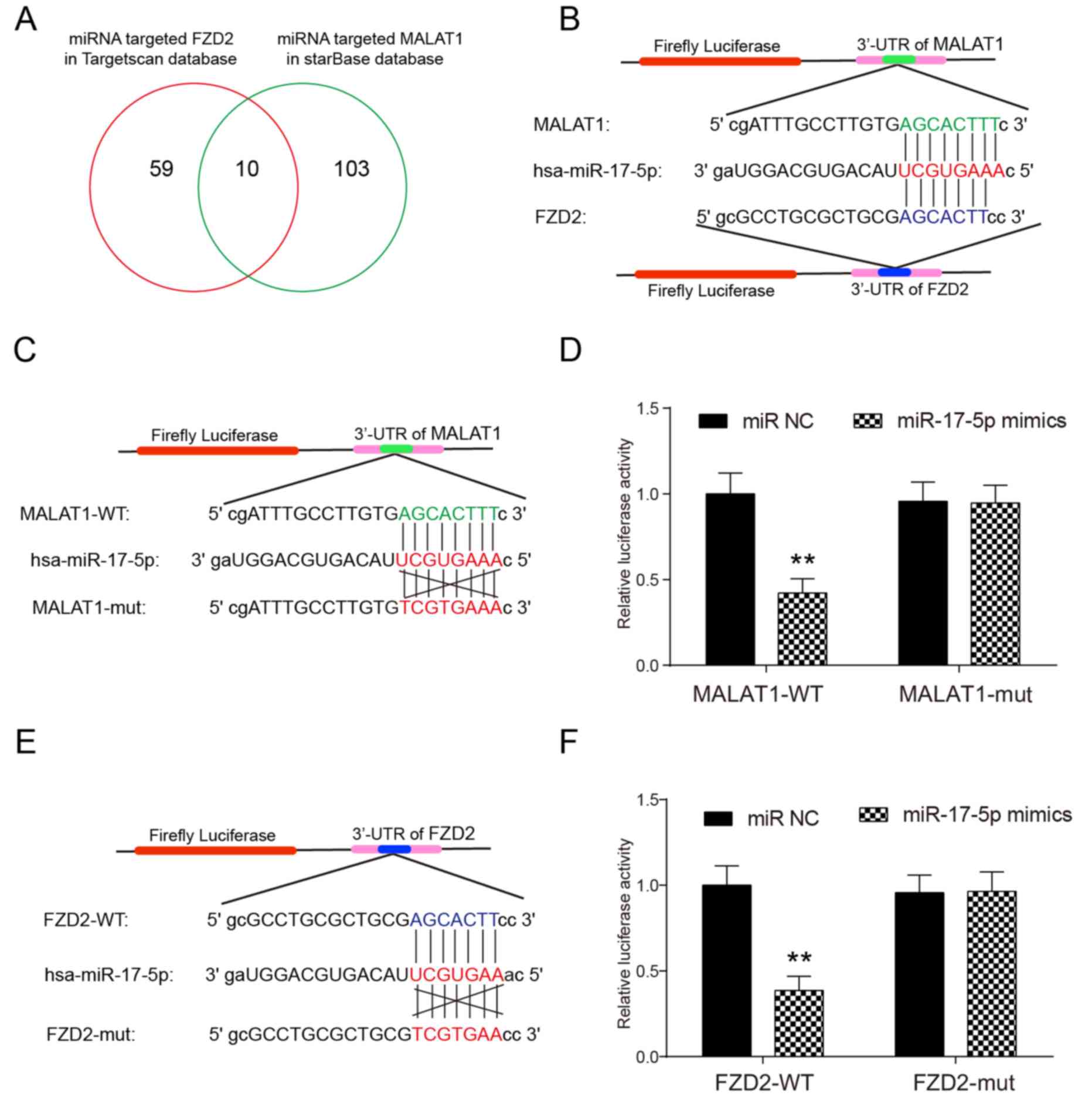
Fig. 4A depicts that
there were 69 miRNAs targeting FZD2 in the TargetScan
database and 113 miRNAs targeting the MALAT1 lncRNA in the starBase
database. A total of 10 miRNAs were common between these two
databases and hsa-miR-17-5p was selected as the target miRNA.
Fig. 4B depicts the target sites that
hsa-miR-17-5p could bind in the 3′-untranslated regions of MALAT1
and FZD2 determined in the starBase database. The target
sites were further verified using a luciferase reporter assay. As
depicted in Fig. 4C and D, miR-17-5p
overexpression resulted in a significant decrease in luciferase
activity in MALAT1-WT without changing the luciferase activity of
MALAT1-mut in HeLa cells, which confirmed that miR-17-5p binds to
MALAT1 via the target sites. A similar result was obtained in the
verification of the targeting association between miR-17-5p and
FZD2, as depicted in Fig. 4E and
F.
Cas-II-gly regulates the expression of
MALAT1, miR-17-5p and FZD2
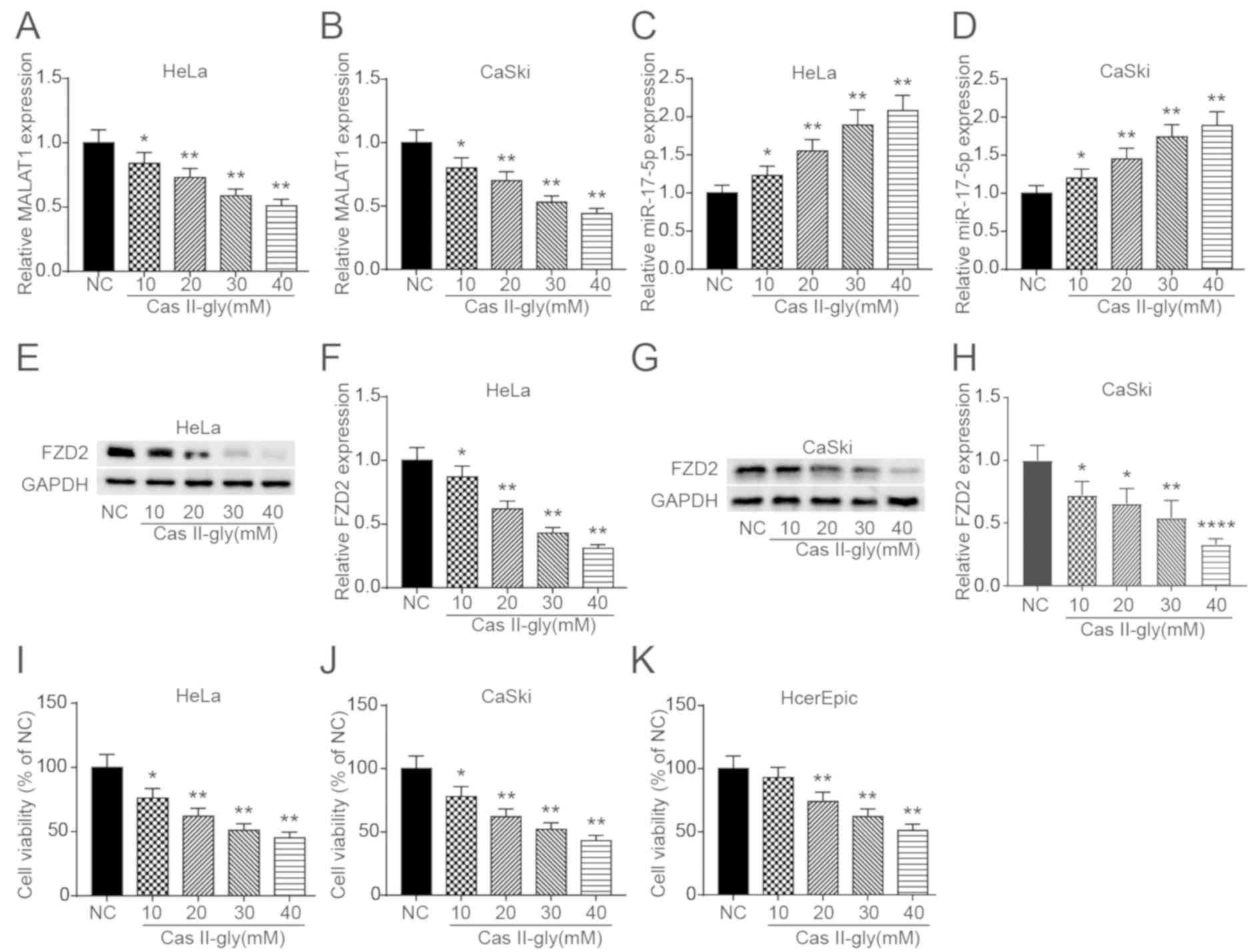
To further investigate whether Cas-II-gly can
regulate the MALAT1-miR-17-5p-FZD2 axis, the expression
profiles of MALAT1, miR-17-5p and FZD2 was measured in HeLa
and CaSki cells treated with increasing doses of Cas-II-gly.
Fig. 5A and B depicts that the
expression of MALAT1 was significantly suppressed with increasing
Cas-II-gly concentrations. The opposite result is depicted in
Fig. 5C and D, as the miR-17-5p
expression significantly increased with increasing Cas-II-gly
concentrations. The western blot results depicted in Fig. 5E-H indicate that FZD2 was also
significantly inhibited by Cas-II-gly. Subsequently, to determine
the optimum Cas-II-gly concentration, MTT assays were performed to
assess the influence of Cas-II-gly on the proliferation of HeLa,
CaSki and HcerEpic cells. The results in Fig. 5I-K demonstrate that 10 mM Cas-II-gly
could inhibit the proliferation of HeLa and CaSki cells, with no
significant killing effect on the HcerEpic cells; therefore, 10 mM
Cas-II-gly was selected for the subsequent experiments.
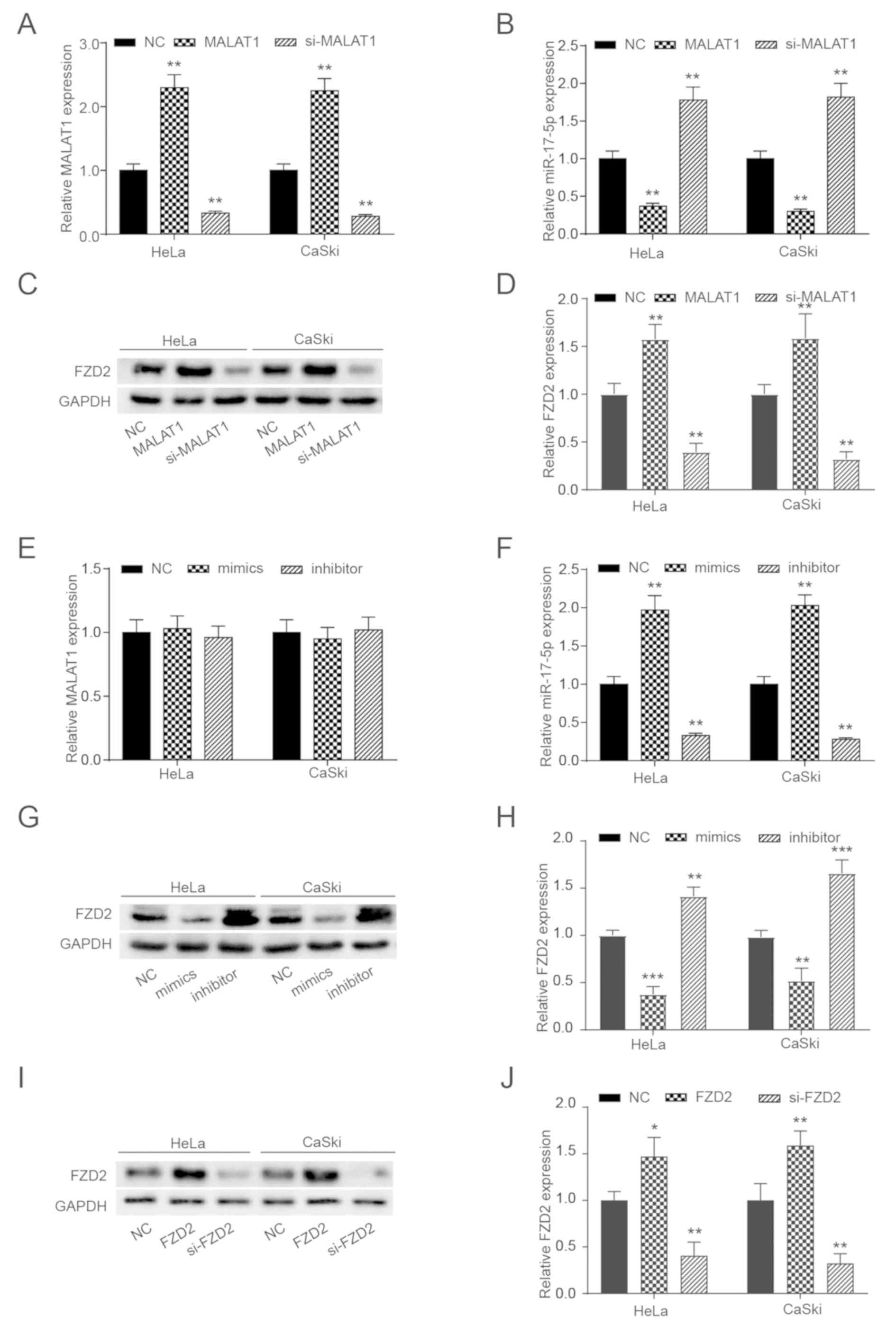
MALAT1 regulates FZD2 expression by
targeting miR-17-5p
To modulate the expression levels of MALAT1, pcDNA
3.1-MALAT1 and si-MALAT1 were designed to overexpress or knockdown
MALAT1 expression, respectively. As depicted in Fig. 6A-D, the expression levels of MALAT1
and FZD2 were significantly increased by pcDNA 3.1-MALAT1
and decreased by si-MALAT1, while the opposite effect was observed
for miR-17-5p expression. Similarly, miR-17-5p mimics and the
inhibitor were designed to modulate the expression levels of
miR-17-5p. The results in Fig. 6E-H
indicate that the miR-17-5p mimics could significantly increase
miR-17-5p expression and significantly suppress FZD2
expression, while miR-17-5p inhibitor had the opposite effects.
However, the miR-17-5p mimics and the inhibitor did not
significantly affect MALAT1 expression. The results of the western
blot analysis depicted in Fig. 6I and
J demonstrated that pcDNA 3.1-FZD2 could significantly
enhance FZD2 expression and si-FZD2 could
significantly reduce it.
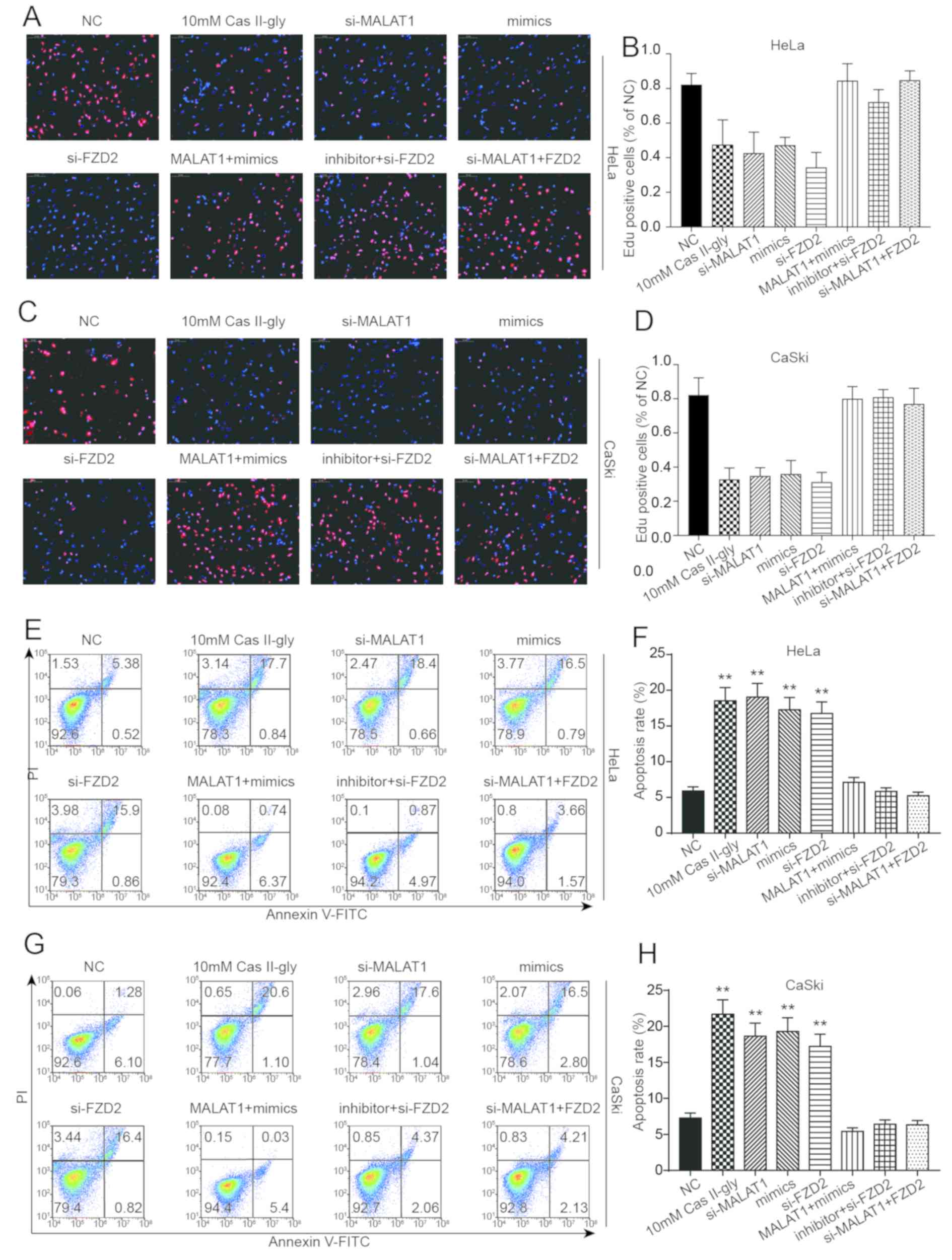
Cas-II-gly affects CC cells via the
MALAT1/miR-17-5p/FZD2 axis
To investigate the effects of these factors on the
proliferation and apoptosis of HeLa and CaSki cells, the
experiments were divided into 8 groups: NC; 10 mM Cas-II-gly;
si-MALAT1; mimics; si-FZD2; MALAT1 + mimics; inhibitor +
si-FZD2; and si-MALAT1 + FZD2. Fig. 7A-D depicts the EdU staining in HeLa
and CaSki cells, and the results indicated that 10 mM Cas-II-gly,
si-MALAT1, mimics and si-FZD2 could significantly reduce the
proliferation of HeLa and CaSki cells, while MALAT1 + mimics,
inhibitor + si-FZD2 and si-MALAT1 + FZD2 did not
cause any significant changes in cell proliferation. The effects on
apoptosis in HeLa and CaSki cells are depicted in Fig. 7E-H. Opposite results were from those
of the EdU staining were obtained, with 10 mM Cas-II-gly,
si-MALAT1, mimics and si-FZD2 significantly increasing the
apoptosis rate of HeLa and CaSki cells, while MALAT1 + mimics,
inhibitor + si-FZD2 and si-MALAT1 + FZD2 had no
significant effects. In conclusion, all these data indicate that
Cas-II-gly can affect the proliferation and apoptosis of CC cells
via the MALAT1/miR-17-5p/FZD2 axis.
Cas-II-gly regulates the Wnt signaling
pathway in CC cells via the MALAT1/miR-17-5p/FZD2 axis
At the conclusion of the present experiments,
western blot analysis was performed to assess the levels of Wnt
signaling pathway-associated proteins (Dvl, GSK-3β and β-catenin)
in HeLa and CaSki cells. As the results in Fig. 8A-D depict, 10 mM Cas-II-gly,
si-MALAT1, mimics and si-FZD2 could increase the expression
of Dvl, β-catenin and GSK-3β in HeLa and CaSki cells, and no
significant effects on these proteins were observed for MALAT1 +
mimics, inhibitor + si-FZD2 and si-MALAT1 + FZD2.
Additionally, all these data demonstrate that Cas-II-gly can
regulate the Wnt signaling pathway via the
MALAT1/miR-17-5p/FZD2 axis in HeLa and CaSki cells.
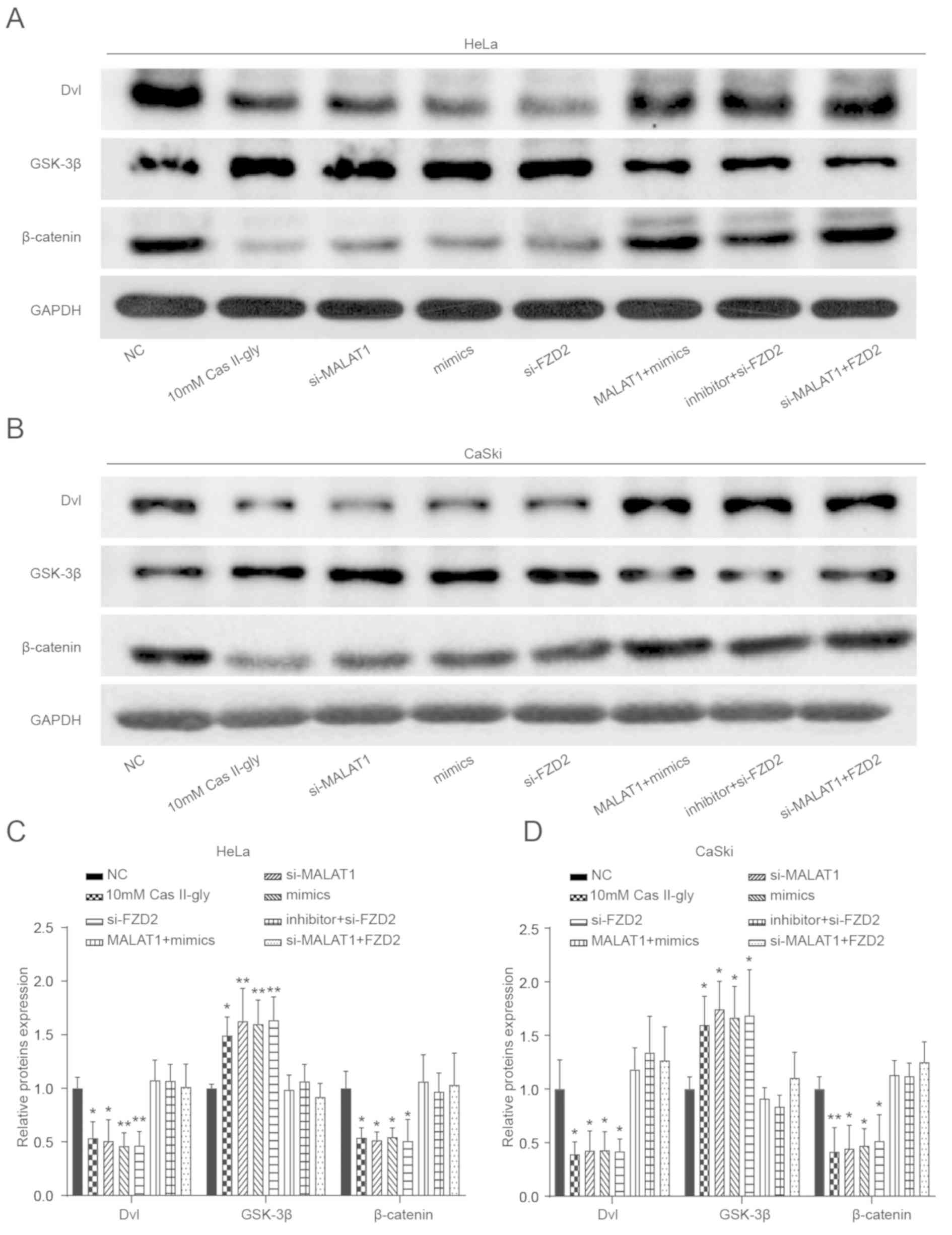 | Figure 8.The effects of Cas-II-gly on the Wnt
signaling pathway in cervical cancer cells. (A-D) Western blotting
was used to measure the levels of Wnt signaling pathway-associated
proteins (Dvl, GSK-3β and β-catenin) in the NC, 10 mM Cas-II-gly,
si-MALAT1, mimics, si-FZD2, MALAT1 + mimics, inhibitor +
si-FZD2 and si-MALAT1 + FZD2 groups. **P<0.01 vs.
the NC group. Cas-II-gly, Casiopeina II-gly; FZD2, frizzled class
receptor 2; MALAT1, metastasis associated lung adenocarcinoma
transcript 1; NC, negative control; si, small interfering; Dvl,
disheveled segment polarity protein; GSK-3β, glycogen synthase
kinase-3β. |
Discussion
In the present study, it was determined that
Cas-II-gly altered the expression levels of the lncRNA MALAT1, the
mRNA FZD2 and suppressed the Wnt signaling pathway in HeLa
and CaSki cells. In vitro experiments were conducted to
further investigate the effects of Cas-II-gly on gene expression
and cell functions, and the results indicated that Cas-II-gly
suppressed the progression of CC cells by suppressing the
MALAT1/miR-17-5p/FZD2/Wnt signaling pathway axis.
Cas-II-gly has been reported to be beneficial in the
treatment of cancer with limited side effects (8). In the present study, the underlying
mechanism, which is not yet fully understood, was investigated. A
number of studies have revealed outcomes associated with Cas-II-gly
treatment. In cervix-uterine and neuroblastoma tumors, Cas-II-gly
may result in mitochondrial apoptosis (7). More specifically, it was reported that
it inhibited the state 3 rates and uncoupled respiration in
mitochondria (41), and that it
induced cytotoxicity associated with oxidative damage (8). Correia et al (42) demonstrated that Cas-II-gly may be
transported by human serum albumin, as low Cas-II-gly
concentrations are expected to be found in blood, which may serve
as a selective tumor target. Another study revealed that Cas-II-gly
may activate the MAPK signaling pathway (7), which is involved in cellular programs,
including proliferation, differentiation, development,
transformation and apoptosis (43).
Cas-II-gly had the same effects on other tumor types. For instance,
in glioma C6 cells, Cas-II-gly inhibited cell proliferation,
increased reactive oxygen species levels and contributed to
apoptosis (44). In CH1 human ovarian
carcinoma cells, Cas-II-gly may induce apoptosis without exhibiting
DNA oligonucleosomal fragments against CC cells (45). A slight decrease in the proliferation
of normal cervical epithelial cells was observed following
treatment with 40 mM Cas-II-gly. Cas-II-gly is a copper-based drug
with anticancer activity (8). While
this compound has some toxicity (46), it may also have a slight side effect
on normal cervical epithelial cells, as indicated by the results of
the present experiments.
In the present study, MALAT1 was downregulated in CC
cells following Cas-II-gly treatment, indicating that MALAT1 may be
a cancer-promoting lncRNA that could contribute to tumor
development. Xia et al (47)
supported this result using metformin. During the process of CC
metastasis, miR-142-3p is significantly upregulated, whereas the
lncRNA MALAT1 and high mobility group AT-hook 2 were suppressed by
metformin (47). Furthermore, MALAT1
and polypyrimidine tract binding protein 2 were overexpressed, and
both are associated with invasion and metastasis in colorectal
cancer (48). Besides, they may also
be oncogenic through other processes. Some results indicated that
MALAT1 promotes proliferation and metastasis in osteosarcoma by
activating the PI3K/Akt pathway (49)
and by sponging miR-200s in clear cell renal carcinoma (50). Knockdown of MALAT1 expression
decreases renal cancer cell proliferation, migration, and invasion
(51). All of these data indicated
that Cas-II-gly may suppress CC cells by regulating MALAT1;
therefore, MALAT1 was selected for the present study.
FZD2, which acts as a Wnt receptor, had
reduced expression in HeLa and CaSki cells following Cas-II-gly
treatment, indicating its cancer-promoting function in CC.
FZD2 belongs to the Fz gene family, which are well
documented to be involved in carcinogenesis. For instance, Li et
al (52) determined that receptor
tyrosine kinase like orphan receptor 2 modulates canonical Wnt
signaling in lung epithelial cells through FZD2. They
demonstrated that FZD2 overexpression in HEC-1B and Ishikawa
cells promoted their migration and that it induced an
epithelial-mesenchymal transition phenotype in endometrial cancer
(53). Another study demonstrated
that overexpression of FZD2 promoted tongue squamous cell
migration and invasion, while RNA interference-mediated FZD2
knockdown had opposite effects on tongue squamous cell migration
and invasion (54). However,
FZD2 expression was reduced in metastatic salivary adenoid
cystic carcinoma tissue, compared with tissue without metastasis
(55). A possible reason for this
difference may be that the metastatic tumor has more complex
behaviors, compared with the original tumor.
Garzon et al (56) reported that miR-17-5p acts as a tumor
suppressor in prostate, cervical and breast cancer. In the present
study, it served as an inhibitor of FZD2 and was
demonstrated to be a target of MALAT1. Wei et al (57) demonstrated that miR-17-5p functions as
a tumor suppressor in CC cells by targeting TP53INP1. It was
reported that miR-17-5p expression was downregulated, while α-1,2
fucosyltransferase 2 expression was increased during osteoarthritis
progression (58). Another study
demonstrated that miR-17 downregulation promoted B cell
CLL/lymphoma 11B overexpression in T cells and promoted the
development of acute lymphoblastic leukemia (59). Furthermore, Li et al (60) revealed that miR-17-5p and miR-20a
alleviate the suppressive function of myeloid-derived suppressor
cells by modulating signal transducer and activator of
transcription 3 expression in acute myeloid leukemia. Furthermore,
miR-17-5p was demonstrated to sensitize MCF-7 cells (a breast
cancer cell line) to paclitaxel-induced apoptosis (15), and these data indicated that miR-17-5p
may be a potential target miRNA of MALAT1 and that it could be
involved in the antitumor mechanism of Cas-II-gly in CC; therefore,
miR-17-5p was selected for further study.
In the present study, the involvement of the
MALAT1-miR-17-5p-FZD2 axis during the treatment of CC cells
with Cas-II-gly was revealed. MALAT1 acted as a competing
endogenous RNA for miR-126-5p to modulate FZD2 expression,
resulting in the activation of the Wnt signaling pathway. Liu et
al (61) indicated that MALAT1
could promote high risk human papillomavirus (+) CC cell growth and
invasion, at least partially through the MALAT1-miR-124-RBG2 axis.
As previously described, MALAT1 may not only function as an
oncogene but could also serve as a drug target (47). Furthermore, MALAT1 and miR-17-5p were
demonstrated to be associated with chemoresistance (15,62,63).
Consistent with other studies, the present data demonstrated that
Cas-II-gly targets the MALAT1-miR-17-5p-FZD2 axis and that
it inactivates the Wnt signaling pathway, thus resulting in cell
apoptosis.
In the present study, it was determined that the Wnt
pathway was suppressed in the Cas-II-gly treatment group, resulting
in apoptosis. Additionally, other studies supported this result.
For example, Pećina-Slaus et al (64) reported that the Wnt pathway regulates
apoptosis. More specifically, Wnt-1 functions as an antiapoptotic
signal, and that it inhibits apoptosis by activating β-catenin/T
cell factor-mediated transcription (65). Similarly, You et al (66) determined that a β-catenin-independent
non-canonical pathway, including the Wnt/JNK pathway, may serve a
role in the Wnt-1 signaling-dependent inhibition of apoptosis. In
other cancer types, it was also demonstrated that the Wnt pathway
was involved in apoptosis. For instance, Wnt signaling regulates
the early and late stages of apoptosis during development and
cellular injury in neurons, endothelial cells, vascular smooth
muscle cells and cardiomyocytes (67). Additionally, in non-small cell lung
cancer, the Wnt pathway was activated by disheveled overexpression,
which contributed to tumor development (68).
In conclusion, it was demonstrated that Cas-II-gly
acts on MALAT1 by targeting miR-17-5p to inhibit FZD2
expression via inactivation of the Wnt signaling pathway, thus
inhibiting cell proliferation and promoting apoptosis in HeLa and
CaSki CC cells.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81771531,
81571395, 81671408 and 81701634).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
YX and RO conducted critical revision of the
manuscript. YX, QZ and FL provided substantial contribution to the
conception and design of the work, and manuscript drafting. LZ, FH
and LZ conducted acquisition, analysis and interpretation of the
data. YX and RO revised the manuscript critically and provided
final approval of the version to be published. All authors have
read and approved the final article.
Ethics approval and consent to
participate
The present study was authorized by ethics
committee of The First Affiliated Hospital of Wenzhou Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Cas-II-gly
|
Casiopeina II-gly
|
|
CC
|
cervical cancer
|
|
DEGs
|
differentially-expressed genes
|
|
FBS
|
fetal bovine serum
|
|
GO
|
Gene Ontology
|
|
GSEA
|
gene set enrichment analysis
|
|
lncRNAs
|
long non-coding RNAs
|
|
PI3K
|
phosphoinositide 3-kinase
|
|
RT
|
reverse transcription
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tewari KS and Monk BJ: New strategies in
advanced cervical cancer: From angiogenesis blockade to
immunotherapy. Clin Cancer Res. 20:5349–5358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McLaughlin-Drubin ME and Münger K: The
human papillomavirus E7 oncoprotein. Virology. 384:335–344. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scheffner M, Werness BA, Huibregtse JM,
Levine AJ and Howley PM: The E6 oncoprotein encoded by human
papillomavirus types 16 and 18 promotes the degradation of p53.
Cell. 63:1129–1136. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leal-Garcia M, Garcia-Ortuno L,
Ruiz-Azuara L, Gracia-Mora I, Luna-Delvillar J and Sumano H:
Assessment of acute respiratory and cardiovascular toxicity of
casiopeinas in anaesthetized dogs. Basic Clin Pharmacol Toxicol.
101:151–158. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chikira M, Tomizawa Y, Fukita D, Sugizaki
T, Sugawara N, Yamazaki T, Sasano A, Shindo H, Palaniandavar M and
Antholine WE: DNA-fiber EPR study of the orientation of Cu(II)
complexes of 1,10-phenanthroline and its derivatives bound to DNA:
Mono(phenanthroline)-copper(II) and its ternary complexes with
amino acids. J Inorg Biochem. 89:163–173. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Valencia-Cruz AI, Uribe-Figueroa LI,
Galindo-Murillo R, Baca-López K, Gutiérrez AG, Vázquez-Aguirre A,
Ruiz-Azuara L, Hernández-Lemus E and Mejía C: Whole genome gene
expression analysis reveals casiopeina-induced apoptosis pathways.
PLoS One. 8:e546642013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alemón-Medina R, Muñoz-Sánchez JL,
Ruiz-Azuara L and Gracia-Mora I: Casiopeina IIgly induced
cytotoxicity to HeLa cells depletes the levels of reduced
glutathione and is prevented by dimethyl sulfoxide. Toxicol In
Vitro. 22:710–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Winegarden N: Microarrays in cancer:
Moving from hype to clinical reality. Lancet. 362:14282003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brown H: The real value of microarray
technology. Lancet Oncol. 4:3262003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Butte A: The use and analysis of
microarray data. Nat Rev Drug Discov. 1:951–960. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin X, Chen X, Hu Y, Ying F, Zou R, Lin F,
Shi Z, Zhu X, Yan X, Li S and Zhu H: LncRNA-TCONS_00026907 is
involved in the progression and prognosis of cervical cancer
through inhibiting miR-143-5p. Cancer Med. 6:1409–1423. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xia YF, Pei GH, Wang N, Che YC, Yu FS, Yin
FF, Liu HX, Luo B and Wang YK: miR-3156-3p is downregulated in
HPV-positive cervical cancer and performs as a tumor-suppressive
miRNA. Virol J. 14:202017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li X, Tian R, Gao H, Yang Y, Williams BRG,
Gantier MP, McMillan NAJ, Xu D, Hu Y and Gao Y: Identification of a
histone family gene signature for predicting the prognosis of
cervical cancer patients. Sci Rep. 7:164952017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liao XH, Xiang Y, Yu CX, Li JP, Li H, Nie
Q, Hu P, Zhou J and Zhang TC: STAT3 is required for
MiR-17-5p-mediated sensitization to chemotherapy-induced apoptosis
in breast cancer cells. Oncotarget. 8:15763–15774. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang B, Li Y and Wang T: A three miRNAs
signature predicts survival in cervical cancer using bioinformatics
analysis. Sci Rep. 7:56242017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liang X, Liu Y, Zeng L, Yu C, Hu Z, Zhou Q
and Yang Z: miR-101 inhibits the G1-to-S phase transition of
cervical cancer cells by targeting Fos. Int J Gynecol Cancer.
24:1165–1172. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao Y, Huang J, Liu T, He S, Shang C, Guo
L, Du Q and Yao S: Overexpression of long non-coding RNA
RP11-396F22.1 correlates poor prognosis of patients with
early-stage cervical cancer. Am J Transl Res. 10:684–695.
2018.PubMed/NCBI
|
|
19
|
Fan Y, Nan Y, Huang J, Zhong H and Zhou W:
Up-regulation of inflammation-related LncRNA-IL7R predicts poor
clinical outcome in patients with cervical cancer. Biosci Rep.
38(pii): BSR201804832018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma TT, Zhou LQ, Xia JH, Shen Y, Yan Y and
Zhu RH: LncRNA PCAT-1 regulates the proliferation, metastasis and
invasion of cervical cancer cells. Eur Rev Med Pharmacol Sci.
22:1907–1913. 2018.PubMed/NCBI
|
|
21
|
Gao D, Zhang Y, Zhu M, Liu S and Wang X:
miRNA expression profiles of HPV-infected patients with cervical
cancer in the uyghur population in China. PLoS One.
11:e01647012016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang L, Shi S, Shi Q, Zhang H, Xia Y and
Zhong T: MicroRNA-519d-3p inhibits proliferation and promotes
apoptosis by targeting HIF-2α in cervical cancer under hypoxic
conditions. Oncol Res. 26:1055–1062. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cai N, Hu L, Xie Y, Gao JH, Zhai W, Wang
L, Jin QJ, Qin CY and Qiang R: MiR-17-5p promotes cervical cancer
cell proliferation and metastasis by targeting transforming growth
factor-β receptor 2. Eur Rev Med Pharmacol Sci. 22:1899–1906.
2018.PubMed/NCBI
|
|
24
|
Rui X, Xu Y, Jiang X, Ye W, Huang Y and
Jiang J: Long non-coding RNA C5orf66-AS1 promotes cell
proliferation in cervical cancer by targeting miR-637/RING1 axis.
Cell Death Dis. 9:11752018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang H, Zhang C, Guan H, Liu J and Cui Y:
LncRNA DANCR promotes cervical cancer progression by upregulating
ROCK1 via sponging miR-335-5p. J Cell Physiol. 234:7266–7278. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen X, Xiong D, Yang H, Ye L, Mei S, Wu
J, Chen S, Shang X, Wang K and Huang L: Long noncoding RNA
OPA-interacting protein 5 antisense transcript 1 upregulated SMAD3
expression to contribute to metastasis of cervical cancer by
sponging miR-143-3p. J Cell Physiol. 234:5264–5275. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng P, Yin Z, Wu Y, Xu Y, Luo Y and
Zhang TC: LncRNA HOTAIR promotes cell migration and invasion by
regulating MKL1 via inhibition miR206 expression in HeLa cells.
Cell Commun Signal. 16:52018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Feng Y, Zou W, Hu C, Li G, Zhou S, He Y,
Ma F, Deng C and Sun L: Modulation of CASC2/miR-21/PTEN pathway
sensitizes cervical cancer to cisplatin. Arch Biochem Biophys.
623-624:20–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang M, Wang M, Li X, Xie Y, Xia X, Tian
J, Zhang K and Tang A: Wnt signaling in cervical cancer? J Cancer.
9:1277–1286. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gupta S, Kumar P and Das BC: HPV:
Molecular pathways and targets. Curr Probl Cancer. 42:161–174.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li P, Hu J, Zhang Y, Li J, Dang Y, Zhang
R, Wei L and Shi M: miR-182 promotes cell proliferation of cervical
cancer cells by targeting adenomatous polyposis coli (APC) gene. Xi
Bao Yu Fen Zi Mian Yi Xue Za Zhi. 34:148–153. 2018.(In Chinese).
PubMed/NCBI
|
|
32
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:Article32004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hollander SL: An unusual case of chronic
lymphocytic leukemia. J Med. 22:289–297. 1991.PubMed/NCBI
|
|
34
|
Yu G, Wang LG, Han Y and He QY:
ClusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kanehisa M, Goto S, Sato Y, Furumichi M
and Tanabe M: KEGG for integration and interpretation of
large-scale molecular data sets. Nucleic Acids Res. 40:D109–D114.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Prieto C, Risueno A, Fontanillo C and De
las Rivas J: Human gene coexpression landscape: Confident network
derived from tissue transcriptomic profiles. PLoS One. 3:e39112008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bravo-Gómez ME, Garcia-Ramos JC,
Gracia-Mora I and Ruiz-Azuara L: Antiproliferative activity and
QSAR study of copper(II) mixed chelate
[Cu(N-N)(acetylacetonato)]NO3 and [Cu(N-N)(glycinato)]NO3
complexes, (Casiopeinas). J Inorg Biochem. 103:299–309. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Alemon-Medina R, Bravo-Gomez ME,
Gracia-Mora MI and Ruiz-Azuara L: Comparison between the
antiproliferative effect and intracellular glutathione depletion
induced by Casiopeina IIgly and cisplatin in murine melanoma B16
cells. Toxicol In Vitro. 25:868–873. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Richter K, Haslbeck M and Buchner J: The
heat shock response: Life on the verge of death. Mol Cell.
40:253–266. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Isserlin R, Merico D, Voisin V and Bader
GD: Enrichment Map-a Cytoscape app to visualize and explore OMICs
pathway enrichment results. F1000Res. 3:1412014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marin-Hernandez A, Gracia-Mora I,
Ruiz-Ramirez L and Moreno-Sanchez R: Toxic effects of copper-based
antineoplastic drugs (Casiopeinas) on mitochondrial functions.
Biochem Pharmacol. 65:1979–1989. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Correia I, Borovic S, Cavaco I, Matos CP,
Roy S, Santos HM, Fernandes L, Capelo JL, Ruiz-Azuara L and Pessoa
JC: Evaluation of the binding of four anti-tumor
Casiopeínas® to human serum albumin. J Inorg Biochem.
175:284–297. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang W and Liu HT: MAPK signal pathways
in the regulation of cell proliferation in mammalian cells. Cell
Res. 12:9–18. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Trejo-Solis C, Palencia G, Zuniga S,
Rodríguez-Ropon A, Osorio-Rico L, Luvia ST, Gracia-Mora I,
Marquez-Rosado L, Sánchez A, Moreno-García ME, et al: Cas IIgly
induces apoptosis in glioma C6 cells in vitro and in vivo through
caspase-dependent and caspase-independent mechanisms. Neoplasia.
7:563–574. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
De Vizcaya-Ruiz A, Rivero-Muller A,
Ruiz-Ramirez L, Kass GE, Kelland LR, Orr RM and Dobrota M:
Induction of apoptosis by a novel copper-based anticancer compound,
casiopeina II, in L1210 murine leukaemia and CH1 human ovarian
carcinoma cells. Toxicol In Vitro. 14:1–5. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Rivero-Muller A, De Vizcaya-Ruiz A, Plant
N, Ruiz L and Dobrota M: Mixed chelate copper complex, Casiopeina
IIgly, binds and degrades nucleic acids: A mechanism of
cytotoxicity. Chem Biol Interact. 165:189–199. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xia C, Liang S, He Z, Zhu X, Chen R and
Chen J: Metformin, a first-line drug for type 2 diabetes mellitus,
disrupts the MALAT1/miR-142-3p sponge to decrease invasion and
migration in cervical cancer cells. Eur J Pharmacol. 830:59–67.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ji Q, Zhang L, Liu X, Zhou L, Wang W, Han
Z, Sui H, Tang Y, Wang Y, Liu N, et al: Long non-coding RNA MALAT1
promotes tumour growth and metastasis in colorectal cancer through
binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2
complex. Br J Cancer. 111:736–748. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dong Y, Liang G, Yuan B, Yang C, Gao R and
Zhou X: MALAT1 promotes the proliferation and metastasis of
osteosarcoma cells by activating the PI3K/Akt pathway. Tumour Biol.
36:1477–1486. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xiao H, Tang K, Liu P, Chen K, Hu J, Zeng
J, Xiao W, Yu G, Yao W, Zhou H, et al: LncRNA MALAT1 functions as a
competing endogenous RNA to regulate ZEB2 expression by sponging
miR-200s in clear cell kidney carcinoma. Oncotarget. 6:38005–38015.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang HM, Yang FQ, Chen SJ, Che J and
Zheng JH: Upregulation of long non-coding RNA MALAT1 correlates
with tumor progression and poor prognosis in clear cell renal cell
carcinoma. Tumour Biol. 36:2947–2955. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li C, Chen H, Hu L, Xing Y, Sasaki T,
Villosis MF, Li J, Nishita M, Minami Y and Minoo P: Ror2 modulates
the canonical Wnt signaling in lung epithelial cells through
cooperation with Fzd2. BMC Mol Biol. 9:112008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bian Y, Chang X, Liao Y, Wang J, Li Y,
Wang K and Wan X: Promotion of epithelial-mesenchymal transition by
Frizzled2 is involved in the metastasis of endometrial cancer.
Oncol Rep. 36:803–810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang E, Li Z, Xu Z, Duan W, Sun C and Lu
L: Frizzled2 mediates the migration and invasion of human oral
squamous cell carcinoma cells through the regulation of the signal
transducer and activator of transcription-3 signaling pathway.
Oncol Rep. 34:3061–3067. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ding LC, Huang XY, Zheng FF, Xie J, She L,
Feng Y, Su BH, Zheng DL and Lu YG: FZD2 inhibits the cell growth
and migration of salivary adenoid cystic carcinomas. Oncol Rep.
35:1006–1012. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Garzon R and Croce CM: MicroRNAs in normal
and malignant hematopoiesis. Curr Opin Hematol. 15:352–358. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wei Q, Li YX, Liu M, Li X and Tang H:
MiR-17-5p targets TP53INP1 and regulates cell proliferation and
apoptosis of cervical cancer cells. IUBMB Life. 64:697–704. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hu J, Wang Z, Shan Y, Pan Y, Ma J and Jia
L: Long non-coding RNA HOTAIR promotes osteoarthritis progression
via miR-17-5p/FUT2/β-catenin axis. Cell Death Dis. 9:7112018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
He Z, Liao Z, Chen S, Li B, Yu Z, Luo G,
Yang L, Zeng C and Li Y: Downregulated miR-17, miR-29c, miR-92a and
miR-214 may be related to BCL11B overexpression in T cell acute
lymphoblastic leukemia. Asia Pac J Clin Oncol. 14:e259–e265. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li Z, Lu J, Sun M, Mi S, Zhang H, Luo RT,
Chen P, Wang Y, Yan M, Qian Z, et al: Distinct microRNA expression
profiles in acute myeloid leukemia with common translocations. Proc
Natl Acad Sci USA. 105:15535–15540. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Liu S, Song L, Zeng S and Zhang L:
MALAT1-miR-124-RBG2 axis is involved in growth and invasion of
HR-HPV-positive cervical cancer cells. Tumour Biol. 37:633–640.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen W, Zhao W, Zhang L, Wang L, Wang J,
Wan Z, Hong Y and Yu L: MALAT1-miR-101-SOX9 feedback loop modulates
the chemo-resistance of lung cancer cell to DDP via Wnt signaling
pathway. Oncotarget. 8:94317–94329. 2017.PubMed/NCBI
|
|
63
|
Chen W, Xu XK, Li JL, Kong KK, Li H, Chen
C, He J, Wang F, Li P, Ge XS and Li FC: MALAT1 is a prognostic
factor in glioblastoma multiforme and induces chemoresistance to
temozolomide through suppressing miR-203 and promoting thymidylate
synthase expression. Oncotarget. 8:22783–22799. 2017.PubMed/NCBI
|
|
64
|
Pećina-Slaus N: Wnt signal transduction
pathway and apoptosis: A review. Cancer Cell Int. 10:222010.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen S, Guttridge DC, You Z, Zhang Z,
Fribley A, Mayo MW, Kitajewski J and Wang CY: Wnt-1 signaling
inhibits apoptosis by activating beta-catenin/T cell
factor-mediated transcription. J Cell Biol. 152:87–96. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
You L, He B, Uematsu K, Xu Z, Mazieres J,
Lee A, McCormick F and Jablons DM: Inhibition of Wnt-1 signaling
induces apoptosis in beta-catenin-deficient mesothelioma cells.
Cancer Res. 64:3474–3478. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li F, Chong ZZ and Maiese K: Winding
through the WNT pathway during cellular development and demise.
Histol Histopathol. 21:103–124. 2006.PubMed/NCBI
|
|
68
|
Uematsu K, He B, You L, Xu Z, McCormick F
and Jablons DM: Activation of the Wnt pathway in non small cell
lung cancer: Evidence of dishevelled overexpression. Oncogene.
22:7218–7221. 2003. View Article : Google Scholar : PubMed/NCBI
|