Introduction
Cervical cancer (CC) is one of the most common
malignancies worldwide. Although screening for CC is widespread,
over 500,000 new cases of CC were estimated to occur worldwide in
2018, thus ranking it as the second most common malignant tumor in
females (1). With advances in
surgery, radiotherapy, and chemotherapy, the five-year survival
rate of patients receiving standardized treatment can reach 80%.
However, once patients develop metastases, the five-year survival
rate drops below 50% (2). Thus, a
deeper understanding of the mechanisms of CC development and
identification of its critical signaling markers as potential
targets for improved treatment strategies is crucial.
Bradykinin (BK) is an active peptide that is
generated by the kallikrein-kinin system (KKS) (3). It has been reported that BK is involved
in the regulation of various cellular processes in cancer cells,
including cell proliferation and angiogenesis (4,5). BK also
facilitates cancer migration and invasion by stimulating the
activity of membrane matrix metalloproteases (MMPs) and integrins
(6,7).
The effect of BK is mediated via two G protein-coupled receptors,
B1 and B2, which have been pharmacologically characterized and
defined by molecular cloning (8).
Furthermore, recent studies have indicated that activation of the
bradykinin B2 receptor (B2R) by BK is involved in most BK-mediated
biological actions (9–11). Studies performed by our group
demonstrated that BK/B2R promoted angiogenesis of cervical cancer
and facilitated tumorigenesis (12).
In the present study, other underlying molecular mechanisms by
which BK promotes tumor progression in CC were explored.
Signal transducer and activator of transcription 3
(STAT3) is persistently activated in many cancers and animal cancer
models (13). Studies have also shown
that STAT3 is constitutively activated in CC and cervical
high-grade lesions (14,15). STAT3 Tyrosine705 phosphorylation
actives this canonical pathway, and results in malignant
transformation by promoting cell proliferation, angiogenesis,
invasion, and metastasis (16–18). The
breakdown of the basement membrane and remodeling of the
extracellular matrix (ECM) are markers for the invasion and
metastasis of malignant tumors. The degradation of ECM proteins is
mainly accomplished by a variety of MMPs (19,20).
Activated STAT3 upregulates MMP expression and activity (15,21) and
downregulates expression of pro-apoptotic proteins (22), thus promoting metastasis and
proliferation in many cancers. Therefore, the expression of BK and
B2R was detected, and their relationship with STAT3 was explored in
CC. The effect of BK treatment on CC cells was examined and the
function of B2R was further characterized in BK-mediated effects.
It was also demonstrated that BK promoted the proliferation,
migration, and invasion of CC cells by activating STAT3 in
vitro. These results highlighted the important role of BK/B2R
in CC.
Materials and methods
Cell lines and cultures
Human cervical cancer cell lines (SiHa and HeLa)
were purchased from the American Type Culture Collection. The cells
were maintained in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% fetal bovine serum (FBS). All the cells were
cultured at 37°C in a humidified atmosphere containing 5%
CO2.
Transduction with lentiviruses
B2R-overexpressed and -knockdown cell lines were
constructed as previously described (12). Target cells were selected with
puromycin (2 µg/ml) for 48 h. B2R expression was detected using
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blotting. All lentiviruses were obtained from
Shanghai GeneChem Co., Ltd. Cell experiments were finished within
three months of cell construction.
RNA isolation and RT-qPCR
Total RNA was extracted from cells using a Total RNA
Kit (Omega Bio-tek) and reverse transcribed into cDNA using M-MLV
reverse transcriptase (Takara Bio, Inc.). Real-time quantitative
PCR was performed using a Bio-Rad CFX96 (Bio-Rad Laboratories,
Inc.) system with SYBR Green (Bio-Rad Laboratories, Inc.) in
triplicate. Each sample was normalized to the control gene GAPDH.
The primer sequences for PCR are presented as follows: human BDKRB2
forward, 5′-CCGAAAGAAGTCTTGGGAGGT-3′ and reverse,
5′-CTGGCGTTCCACGGAGATG-3′; human GAPDH forward,
5′-GACAGTCAGCCGCATCTTCT-3′ and reverse, 5′-TTAAAAGCAGCCCTGGTGAC-3′.
The cycling conditions were as follows: 95°C for 3 min, then 45
cycles of 95°C for 15 sec and 60°C for 30 sec.
Western blot analysis
Cells were lysed in radioimmunoprecipitation assay
(RIPA) lysis buffer (Beyotime Institute of Biotechnology)
supplemented with a protease inhibitor cocktail (Roche
Diagnostics). Concentrations of the proteins extracted from cell
lines were determined using the Coomassie brilliant blue G-250
(BioFroxx; neoFroxx GmbH) staining method. Equal amounts of cell
lysate (30 µg) were resolved by 10% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a
polyvinylidene difluoride membrane. The blots were blocked with 5%
bovine serum albumen (BSA) at room temperature for 1 h and then
incubated with primary antibodies overnight at 4°C. After washing
three times, appropriate secondary antibodies were added, followed
by incubation for 1 h at room temperature. Primary antibodies were
as follows: p-STAT3 (dilution 1:2,000; product no. 9145T) and STAT3
(dilution 1:2,000; product no. 4904T; both from Cell Signaling
Technology, Inc.), MMP2 (dilution 1:1,000; product code ab92536;
Abcam), MMP9 (dilution 1:600; cat. no. 10375-2-AP; ProteinTech
Group, Inc.), caspase-9 (dilution 1:1,000; product no. 9504T) and
cleaved caspase-9 (dilution 1:1,000; product no. 9509T; both from
Cell Signaling Technology, Inc.), BDKRB2 (dilution 1:500; product
code ab134118; Abcam), GAPDH (dilution 1:2,000, cat. no. AC001;
ABclonal Technology), and α-tubulin (dilution 1:2,000; cat. no.
A41641; Antgene). The proteins were detected using an enhanced
chemiluminescence system (Pierce; Thermo Fisher Scientific, Inc.).
The statistical data on protein levels in western blots were
analyzed using ImageJ 1.8.0 software (National Institutes of
Health).
Plate colony formation assay
Cells were digested and diluted with DMEM. For each
group, 800 cells in 2 ml DMEM were seeded in six-well culture
plates and then maintained in a 5% CO2 incubator at
37°C. Various concentrations of BK (1, 2.5 and 5 µM) and the
bradykinin B2 receptor antagonist HOE140 (Tocris Bioscience) (1, 5
and 10 µM) were added. Constructed cells were treated with 2.5 µM
BK. The culture plates were collected 12 days later, and the clones
were stained with 0.1% crystal violet for 10 min at room
temperature for visualization. Experiments were performed in
triplicate.
Migration assays
Using a 24-well Transwell plate containing PET
membranes with 8-µm pores (Corning Inc.), CC cells were added into
the upper chambers with non-coated membranes. Approximately
1×104 cells in 100 µl of serum-free DMEM were placed in
the upper chamber, and 500 µl of the same medium containing BK (5
µM; Tocris Bioscience) was placed in the lower chamber. For cells
transduced with lentiviruses, 500 µl medium supplemented with 20%
FBS and 2.5 µM BK was placed into the lower chambers. After 24 h at
37°C, the cells that migrated into the underside of the membrane
were fixed in 4% paraformaldehyde for 15 min, stained with 0.5%
crystal violet for 20 min, and washed with PBS. At least six random
fields were assessed under an Olympus IX73 microscope (Olympus
Corp.) (×10, magnification). Experiments were performed in
triplicate.
Invasion assay
The procedure for the invasion assay was similar to
the migration assay described above, except that the upper chambers
were coated with Matrigel (10 mg/ml; Corning Incorporated) diluted
five times in serum-free DMEM. Briefly, the cells were placed in
the upper chamber, and medium containing BK (5 µM) or 20% FBS with
2.5 µM BK was placed into the lower chambers. After 36 h at 37°C,
the cells were fixed, stained, washed, and counted. Experiments
were performed in triplicate.
Scratch assay
Cells were added to six-well plates and then
cultured in a 5% CO2 incubator at 37°C. Once the cell
density reached 80%, a vertical wound was produced in the monolayer
and BK (5 µM) or HOE140 (10 µM) were added. Constructed cells were
treated with 2.5 µM BK. The closure of the wound was monitored at
0, 24, 36, 48, and 60 h and measured using ImageJ 1.8.0 software
(National Institutes of Health). For each experiment, at least
three scratched fields were recorded, and all scratch assays were
performed in triplicate (×10, magnification).
Statistical analysis
The data are reported as the means ± standard error.
Statistical evaluation between groups was conducted with the
Student's t-test. Comparisons of more than two groups were
performed using one-way analysis of variance (ANOVA) followed by
Tukey's post hoc test. Differences were considered statistically
significant at P<0.05.
Results
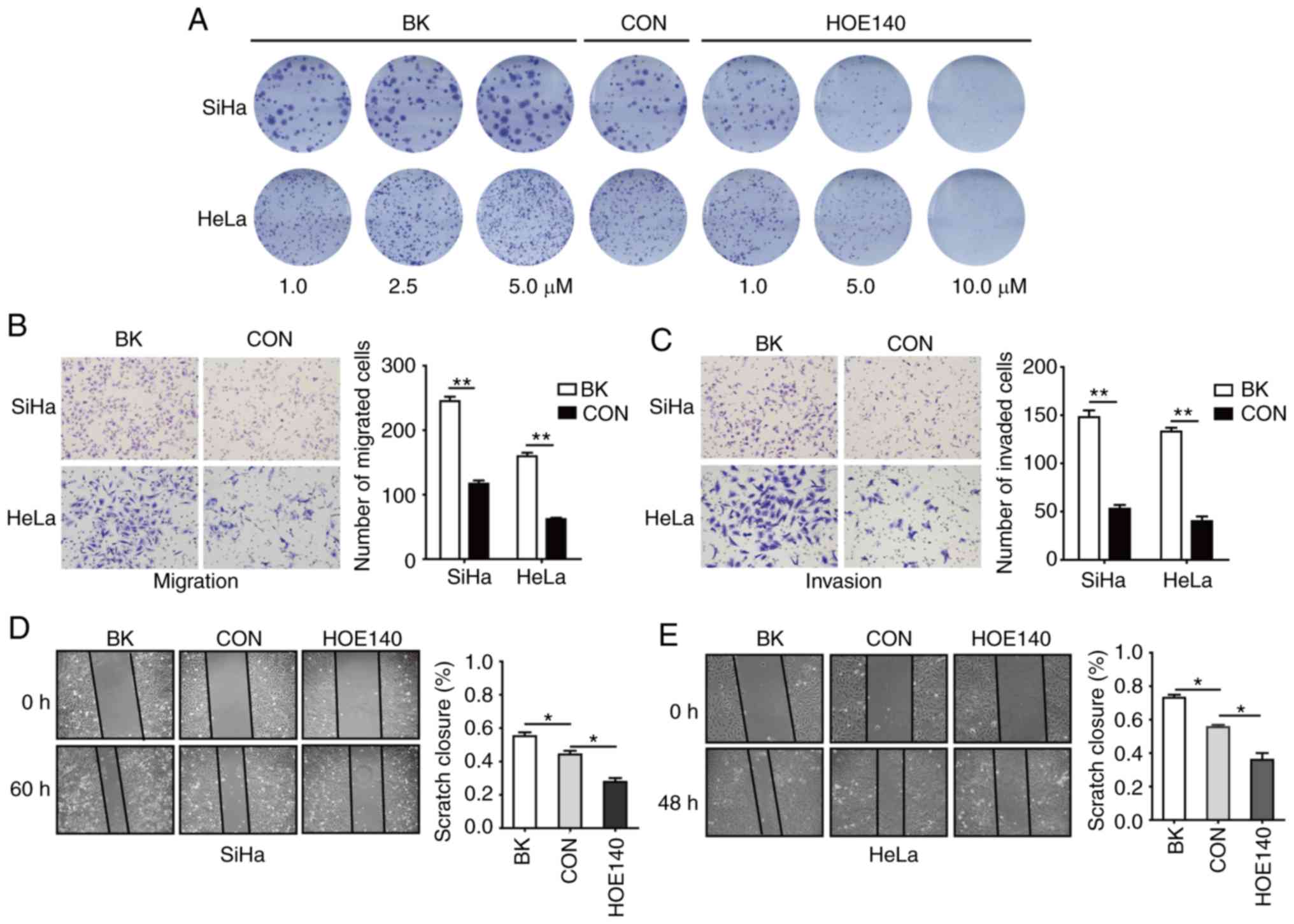
BK enhances the proliferation,
migration, and invasion of CC cells
To assess the potential role of BK in CC, colony
formation, Transwell, and cell wound-healing assays were conducted.
Plate colony formation assays were used to assess the proliferative
activity of cells (14). In this
assay, SiHa and HeLa cells were treated with 1, 2.5 or 5 µM BK or
1, 5, or 10 µM HOE140, respectively. As revealed in Fig. 1A, BK treatment resulted in a
dose-dependent increase in cell proliferation, whereas HOE140 had
the opposite effect, inhibiting cell proliferation in a
concentration-dependent manner as compared with the control cells.
Transwell assays revealed that BK enhanced the migration and
invasion of human CC cells (Fig. 1B and
C). In addition, cell wound-healing assays also revealed that
BK promoted CC cell migration and that HOE140 blocked CC cell
migration (Fig. 1D and E). These data
indicated that BK promoted cell proliferation, migration, and
invasion in CC cells.
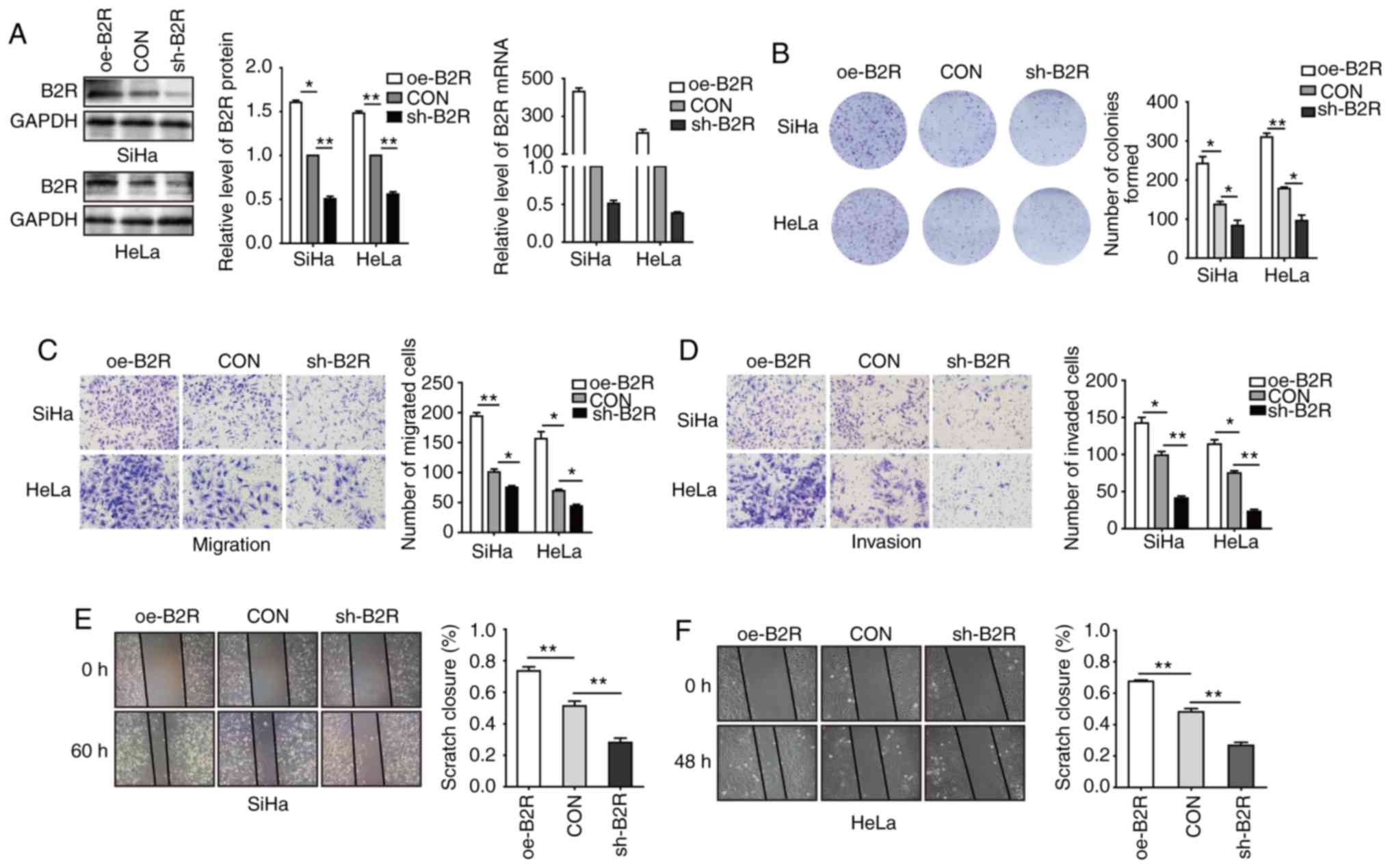
Involvement of B2R in BK-mediated
proliferation, migration, and invasion of CC cells
To further determine whether B2R is involved in the
BK-mediated biological function of CC cell lines, B2R-overexpressed
and -knockdown cell lines were constructed. After confirming mRNA
and protein levels (Fig. 2A), plate
cell colony formation assays were conducted. Fig. 2B revealed that B2R overexpression
significantly increased cell proliferation and that B2R knockdown
markedly reduced cell proliferation. Transwell assays revealed that
B2R-overexpressed CC cells exhibited greater migration and invasion
abilities compared to those of the control cells, while in
B2R-knockdown CC cells, the effect was reversed (Fig. 2C and D). The results of the scratch
assays were similar to those of the Transwell assays (Fig. 2E and F). During the incubation period,
2.5 µM BK was added to all cell supernatants. Collectively, these
data indicated that BK promoted CC cell proliferation, migration,
and invasion via B2R.
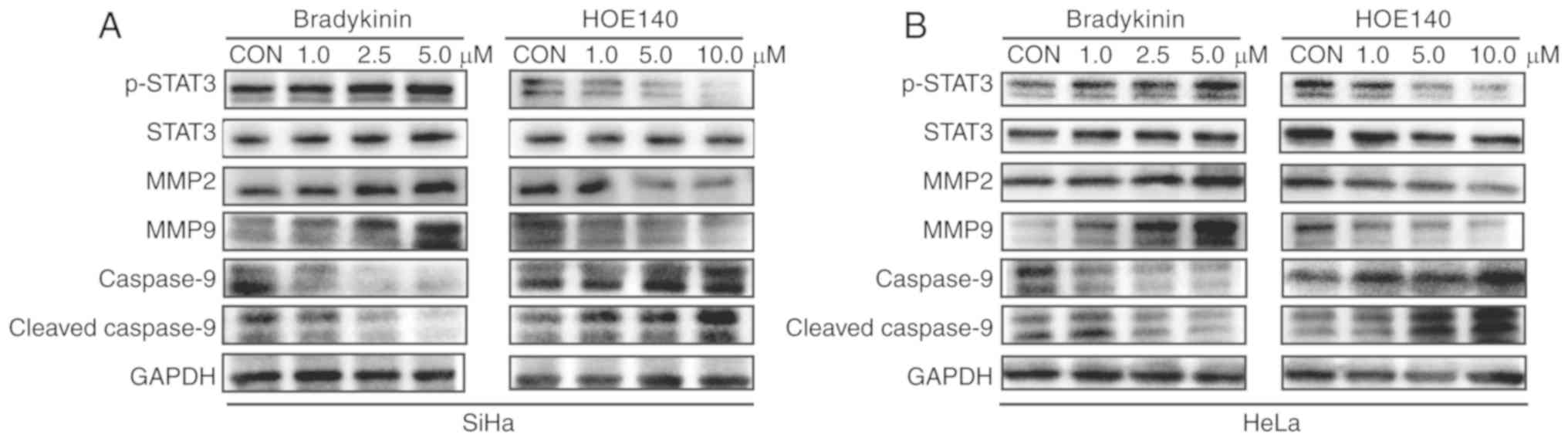
BK upregulates MMP expression in CC
cells
Whether MMPs are involved in BK-induced invasion and
migration of CC cells was also investigated. Notably, treatment
with BK resulted in a concentration-dependent upregulation of MMP2
and MMP9 in SiHa and HeLa cells (Fig. 3A
and B). However, MMP2 and MMP9 expression was inhibited by
HOE140 in a dose-dependent manner (Fig.
3A and 3B). Moreover, there was a
positive association between B2R expression and MMP2 and MMP9
expression (Fig. 4A and B). Thus,
BK/B2R mediated migration and invasion of CC cells by increasing
the expression of MMP2 and MMP9.
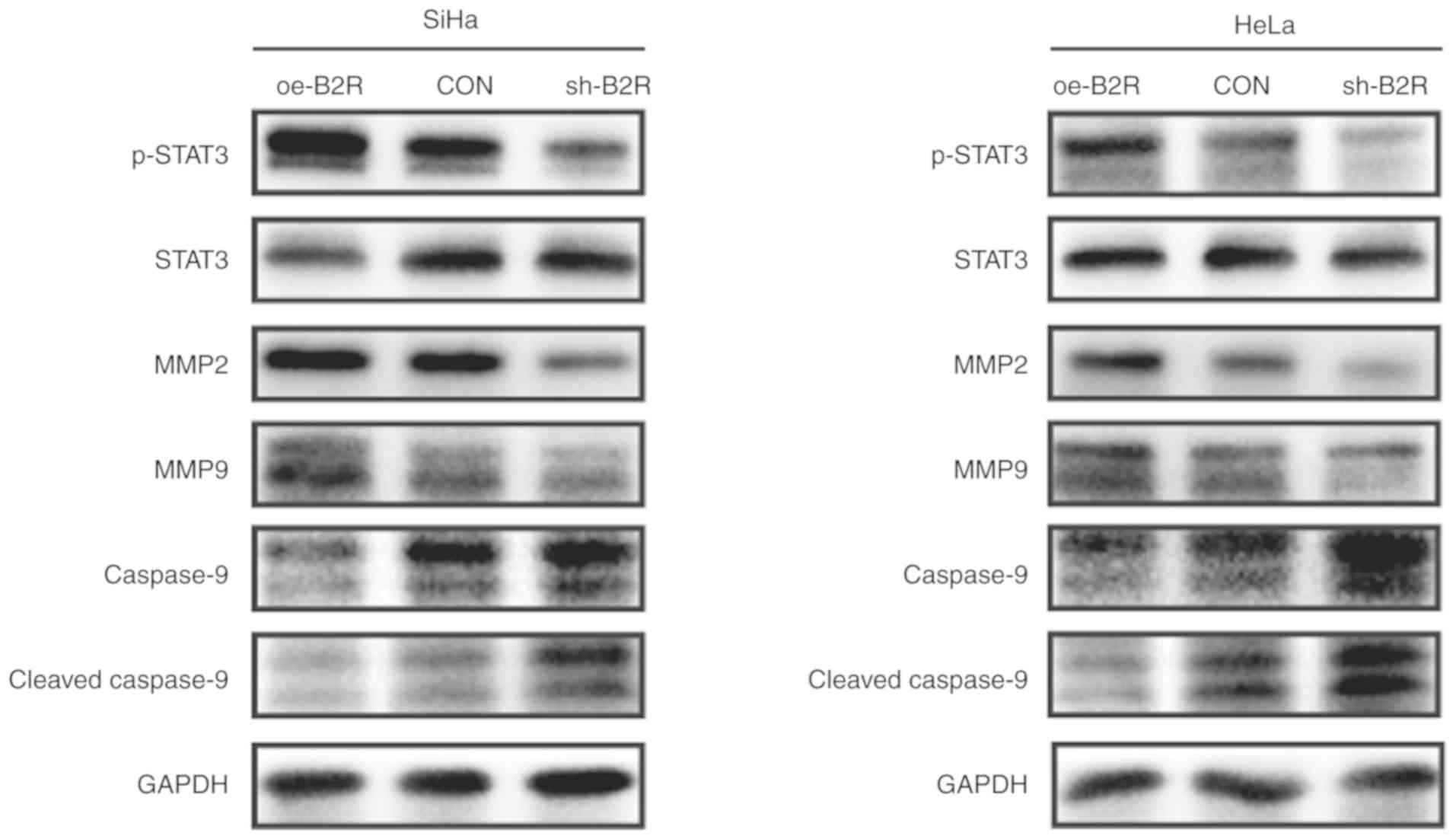 | Figure 4.Representative blots revealing the
protein levels of p-STAT3, STAT3, MMP2, MMP9, cleaved caspase-9,
and caspase-9 in (A) SiHa and (B) HeLa cells transfected with
oe-B2R, sh-B2R, or an empty lentiviral vector. GAPDH was loaded as
a control. The results were similar for three independent
experiments. p-STAT3, phosphorylated-STAT3; B2R, bradykinin B2
receptor; MMP, matrix metalloprotease. |
BK enhances the activation of STAT3
signaling pathways to promote CC cell proliferation, migration, and
invasion
Given that abnormal activation of STAT3 signaling
plays an important role in CC cell proliferation, migration, and
invasion (23,24), whether STAT3 signaling pathways are
involved in the BK-mediated biological function of CC cells was
investigated. Western blotting indicated that phosphorylated
(p)-STAT3 protein levels were increased after treatment with BK
while HOE140 decreased p-STAT3 protein levels in a dose-dependent
manner. For the STAT3-regulated pro-apoptotic protein cleaved
caspase-9, the effect was reversed (Fig.
3A and B). Similarly, the protein level of p-STAT3 in
B2R-overexpressed cells was markedly increased compared with that
in the control cells, while B2R-knockdown CC cells exhibit the
opposite effect. The expression of B2R also exhibited a positive
association with those of MMP2 and MMP9 and a negative association
with that of the apoptosis pathway-related protein cleaved
caspase-9 (Fig. 4A and B). In
summary, the present findings indicated that the activation of B2R
and STAT3 played important roles during BK-induced proliferation,
migration, and invasion in CC cells.
Discussion
BK, the most prominent member of the kinin group,
plays an important role in many cellular processes (25). An increasing number of studies have
revealed that BK treatment promotes proliferation, angiogenesis,
and metastasis in many cancers (11,26). The
present findings have revealed for the first time, to the best of
our knowledge, that BK treatment promoted proliferation, migration,
and invasion of CC cells, whereas the B2R inhibitor HOE140
exhibited the opposite effect. Previous research performed by our
group determined that BK promoted CC angiogenesis via B2R but not
B1R (12). Furthermore, in the
present study, overexpression of B2R facilitated BK-treated CC cell
proliferation, migration, and invasion, whereas downregulatoin of
B2R blocked them. Collectively, the results of the present study
indicated that BK plays a biological role via B2R in CC cells.
Several studies have demonstrated that BK promotes
tumorigenesis through the MAPK, PKC, NF-κB, and ERK signaling
pathways (7,9,11,27). Constitutive STAT3 activation was
revealed to be associated with various human cancers and indicated
poor prognosis (14,17,28,29). The
present study revealed that treatment of CC cells with BK induced
an increase in the expression of p-STAT3 Tyrosine705 (Y705) in a
concentration-dependent manner. Moreover, the B2R antagonist HOE140
has the opposite effect. Activated STAT3 has been revealed to
upregulate MMP expression and reduce the expression of
pro-apoptotic proteins (15,22). To the best of our knowledge, the
present findings are the first to demonstrate that treatment of CC
cells with BK/B2R upregulated p-STAT3 expression while it
downregulated the expression of the pro-apoptotic protein cleaved
caspase-9, consequently facilitating CC cell migration and invasion
by stimulating the activity of MMP2 and MMP9, whereas silencing of
B2R blocked these effects. These data confirmed that BK/B2R plays
an important role in the malignant progression of CC by activating
STAT3.
STAT3 plays an important role in tumorigenesis, and
many studies have demonstrated that STAT3 inhibitors are promising
agents for antitumor therapy (13).
However, although small-molecule STAT3 inhibitors exhibit antitumor
effects, they also cause serious side effects, some of which are
fatal (30,31). The present study revealed that the B2R
inhibitor HOE140 hindered the activation of STAT3, thus, it was
speculated whether B2R inhibitors can also achieve the same
antitumor effect as STAT3 inhibitors, but with reduced side
effects. This is a potentially fruitful area of future research and
may bring new hope to many cancer patients.
Notably, based on the biological function of BK in
tumors, there are several studies that suggest that BK antagonists
can be used as novel antitumor drugs (32). In the present study, the B2R
antagonist HOE140 was revealed to inhibit the proliferation,
invasion, and metastasis of CC cells. Moreover, Avdieiev et
al reported that the combination of a BK receptor antagonist
and thiazolidinone derivatives had a significant synergistic effect
on fatality to rat and human glioma cells and also reduced drug
resistance (33). BK antagonists have
also been used in human trials and no harmful side effects have
been observed. Thus, BK antagonists can be used for anticancer
therapy (34). Accordingly, BK
antagonists may serve as very promising agents for the treatment of
CC. For patients with recurrent CC and/or that are drug resistant,
the combination of cisplatin/paclitaxel with Bevacizumab, an
antibody directed against VEGF-A, provides a new treatment strategy
which can increase the toxicity of drugs to tumor cells (35). For such patients, B2R antagonist
treatment may also provide an option worthy of investigation.
In summary, it was demonstrated for the first time
that BK promoted CC cell proliferation, migration, and invasion
through activation of the B2 receptor and STAT3 signaling pathways.
It was also revealed that BK/B2R facilitated CC cell migration and
invasion by upregulating the expression of the STAT3-regulated
products MMP2 and MMP9, and that BK/B2R downregulated the
expression of the pro-apoptotic protein cleaved caspase-9. These
findings will enhance our understanding of the mechanisms of CC
tumorigenesis and metastasis, and they may provide an alternative
to current CC therapies.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Natural Science
Foundation Committee of China (NSFC81472444, NSFC81472783,
NSFC81630060) and The Graduates' Innovation Fund of Huazhong
University of Science and Technology (no. 2019ygscxcy060).
Availability of data and materials
All data used or analyzed during the current study
are included in this published article.
Authors' contributions
WW conceived and designed the study, participated in
every experiment, and drafted and critically revised the
manuscript. YZ participated in experimental design and revised the
manuscript. RW, GJ, FL, XC, XW analyzed the data. DM provided
experimental technical support. LX provided final approval of the
version to be published and was involved in the conception of the
study. All authors revised, read and approved the final version of
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petignat P and Roy M: Diagnosis and
management of cervical cancer. BMJ. 335:765–768. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moreau ME, Garbacki N, Molinaro G, Brown
NJ, Marceau F and Adam A: The kallikrein-kinin system: Current and
future pharmacological targets. J Pharmacol Sci. 99:6–38. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Maeda H, Wu J, Okamoto T, Maruo K and
Akaike T: Kallikrein-kinin in infection and cancer.
Immunopharmacology. 43:115–128. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Searovic P, Alonso M, Oses C,
Pereira-Flores K, Velarde V and Saez CG: Effect of tamoxifen and
retinoic acid on bradykinin induced proliferation in MCF-7 cells. J
Cell Biochem. 106:473–481. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu J, Akaike T, Hayashida K, Okamoto T,
Okuyama A and Maeda H: Enhanced vascular permeability in solid
tumor involving peroxynitrite and matrix metalloproteinases. Jpn J
Cancer Res. 92:439–451. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang WH, Chang JT, Hsu SF, Li TM, Cho DY,
Huang CY, Fong YC and Tang CH: Bradykinin enhances cell migration
in human chondrosarcoma cells through BK receptor signaling
pathways. J Cell Biochem. 109:82–92. 2010.PubMed/NCBI
|
|
8
|
Menke JG, Borkowski JA, Bierilo KK,
MacNeil T, Derrick AW, Schneck KA, Ransom RW, Strader CD, Linemeyer
DL and Hess JF: Expression cloning of a human B1 bradykinin
receptor. J Biol Chem. 269:21583–21586. 1994.PubMed/NCBI
|
|
9
|
Wang G, Ye Y, Zhang X and Song J:
Bradykinin stimulates IL-6 production and cell invasion in
colorectal cancer cells. Oncol Rep. 32:1709–1714. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vassou D, Notas G, Hatzoglou A, Castanas E
and Kampa M: Opioids increase bladder cancer cell migration via
bradykinin B2 receptors. Int J Oncol. 39:697–707. 2011.PubMed/NCBI
|
|
11
|
Yu HS, Wang SW, Chang AC, Tai HC, Yeh HI,
Lin YM and Tang CH: Bradykinin promotes vascular endothelial growth
factor expression and increases angiogenesis in human prostate
cancer cells. Biochem Pharmacol. 87:243–253. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Y, Wang W, Wei R, Jiang G, Li F, Chen
X, Wang X, Long S, Ma D and Xi L: Serum bradykinin levels as a
diagnostic marker in cervical cancer with a potential mechanism to
promote VEGF expression via BDKRB2. Int J Oncol. 55:131–141.
2019.PubMed/NCBI
|
|
13
|
Wang X, Crowe PJ, Goldstein D and Yang JL:
STAT3 inhibition, a novel approach to enhancing targeted therapy in
human cancers (Review). Int J Oncol. 41:1181–1191. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yao S, Xu J, Zhao K, Song P, Yan Q, Fan W,
Li W and Lu C: Down-regulation of HPGD by miR-146b-3p promotes
cervical cancer cell proliferation, migration and
anchorage-independent growth through activation of STAT3 and AKT
pathways. Cell Death Dis. 9:10552018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saini U, Naidu S, ElNaggar AC, Bid HK,
Wallbillich JJ, Bixel K, Bolyard C, Suarez AA, Kaur B, Kuppusamy,
et al: Elevated STAT3 expression in ovarian cancer ascites promotes
invasion and metastasis: A potential therapeutic target. Oncogene.
36:168–181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Alvarez JV, Greulich H, Sellers WR,
Meyerson M and Frank DA: Signal transducer and activator of
transcription 3 is required for the oncogenic effects of
non-small-cell lung cancer-associated mutations of the epidermal
growth factor receptor. Cancer Res. 66:3162–3168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang S, Yang C, Yu F, Ding W, Hu Y, Cheng
F, Zhang F, Guan B, Wang X, Lu L and Rao J: Endoplasmic reticulum
resident oxidase ERO1-Lalpha promotes hepatocellular carcinoma
metastasis and angiogenesis through the S1PR1/STAT3/VEGF-A pathway.
Cell Death Dis. 9:11052018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pradeep S, Huang J, Mora EM, Nick AM, Cho
MS, Wu SY, Noh K, Pecot CV, Rupaimoole R, Stein MA, et al:
Erythropoietin stimulates tumor growth via EphB4. Cancer Cell.
28:610–622. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Smith HA and Kang Y: The
metastasis-promoting roles of tumor-associated immune cells. J Mol
Med (Berl). 91:411–429. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Z, Tian Z, Cao K, Zhang B, Wen Q, Zhou
X, Yang W, Wang T, Shi H and Wang R: TSG101 promotes the
proliferation, migration and invasion of hepatocellular carcinoma
cells by regulating the PEG10. J Cell Mol Med. 23:70–82. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shishodia G, Shukla S, Srivastava Y,
Masaldan S, Mehta S, Bhambhani S, Sharma S, Mehrotra R, Das BC and
Bharti AC: Alterations in microRNAs miR-21 and let-7a correlate
with aberrant STAT3 signaling and downstream effects during
cervical carcinogenesis. Mol Cancer. 14:1162015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guha P, Gardell J, Darpolor J, Cunetta M,
Lima M, Miller G, Espat NJ, Junghans RP and Katz SC: STAT3
inhibition induces Bax-dependent apoptosis in liver tumor
myeloid-derived suppressor cells. Oncogene. 38:533–548. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J, Jia J, Zhao L, Li X, Xie Q, Chen
X, Wang J and Lu F: Down-regulation of microRNA-9 leads to
activation of IL-6/Jak/STAT3 pathway through directly targeting
IL-6 in HeLa cell. Mol Carcinog. 55:732–742. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao H, Wang W, Zhao Q, Hu G, Deng K and
Liu Y: BCL3 exerts an oncogenic function by regulating STAT3 in
human cervical cancer. Onco Targets Ther. 9:6619–6629. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
da Costa PL, Sirois P, Tannock IF and
Chammas R: The role of kinin receptors in cancer and therapeutic
opportunities. Cancer Lett. 345:27–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang G, Sun J, Liu G, Fu Y and Zhang X:
Bradykinin promotes cell proliferation, migration, invasion, and
tumor growth of gastric cancer through ERK signaling pathway. J
Cell Biochem. 118:4444–4453. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sabatini F, Luppi F, Petecchia L, Stefano
AD, Longo AM, Eva A, Vanni C, Hiemstra PS, Sterk PJ, Sorbello V, et
al: Bradykinin-induced asthmatic fibroblast/myofibroblast
activities via bradykinin B2 receptor and different MAPK pathways.
Eur J Pharmacol. 710:100–109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Morikawa T, Baba Y, Yamauchi M, Kuchiba A,
Nosho K, Shima K, Tanaka N, Huttenhower C, Frank DA, Fuchs CS and
Ogino S: STAT3 expression, molecular features, inflammation
patterns, and prognosis in a database of 724 colorectal cancers.
Clin Cancer Res. 17:1452–1462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yin W, Cheepala S, Roberts JN, Syson-Chan
K, DiGiovanni J and Clifford JL: Active Stat3 is required for
survival of human squamous cell carcinoma cells in serum-free
conditions. Mol Cancer. 5:152006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wong AL, Soo RA, Tan DS, Lee SC, Lim JS,
Marban PC, Kong LR, Lee YJ, Wang LZ, Thuya WL, et al: Phase I and
biomarker study of OPB-51602, a novel signal transducer and
activator of transcription (STAT) 3 inhibitor, in patients with
refractory solid malignancies. Ann Oncol. 26:998–1005. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Murakami T, Takigawa N, Ninomiya T, Ochi
N, Yasugi M, Honda Y, Kubo T, Ichihara E, Hotta K, Tanimoto M and
Kiura K: Effect of AZD1480 in an epidermal growth factor
receptor-driven lung cancer model. Lung Cancer. 83:30–36. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stewart JM: Bradykinin antagonists as
anti-cancer agents. Curr Pharm Des. 9:2036–2042. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Avdieiev S, Gera L, Havrylyuk D, Hodges
RS, Lesyk R, Ribrag V, Vassetzky Y and Kavsan V: Bradykinin
antagonists and thiazolidinone derivatives as new potential
anti-cancer compounds. Bioorg Med Chem. 22:3815–3823. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Stewart JM, Gera L, Chan DC, Bunn PJ Jr,
York EJ, Simkeviciene V and Helfrich B: Bradykinin-related
compounds as new drugs for cancer and inflammation. Can J Physiol
Pharmacol. 80:275–280. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Monk BJ, Sill MW, Burger RA, Gray HJ,
Buekers TE and Roman LD: Phase II trial of bevacizumab in the
treatment of persistent or recurrent squamous cell carcinoma of the
cervix: A gynecologic oncology group study. J Clin Oncol.
27:1069–1074. 2009. View Article : Google Scholar : PubMed/NCBI
|