Introduction
Prostate cancer (PCa) is one of the most common
types of cancer among men and accounts for 1/5 patients
newly-diagnosed with cancer in the USA alone (1). There were at least 180,000
newly-diagnosed cases and 26,000 estimated-deaths in 2016. Current
treatments include hormonal therapy, radical prostatectomy and
radiation therapy. However, almost all patients with PCa eventually
develop incurable metastatic castration-resistant PCa (mCRPC) and
generally succumb to this disease 2 years later (2,3).
Currently, the mechanism underlying PCa development is still far
from being understood, and thus investigating its molecular
mechanisms is important and will help improve therapeutic
efficacy.
p62 (also known as sequestosome-1, SQSTM-1 or A170)
contains 440 amino acid residues, is named based on its molecular
weight of 62 kDa, and plays a key role in the regulation of cell
proliferation and survival (4). p62
serves as a selective autophagy adaptor protein that links
ubiquitinated proteins to autophagy machinery for degradation. p62
harbors multiple functional domains and plays important roles in
multiple critical signaling pathways. An increasing amount of
evidence has demonstrated that high levels of p62 are associated
with multiple diseases including liver disorders, neurodegenerative
diseases and cancers (5).
The Kelch-like ECH-associated protein 1/nuclear
factor erytheroid-derived-2-like 2/antioxidant response element
(Keap1/Nrf2/ARE) system is currently recognized as one of the major
defense mechanisms against oxidative stress and xenobiotics
(6,7), thus, the dysregulation of the
Keap1/Nrf2/ARE system is involved in several human illnesses,
particularly cancers (8,9). Nrf2 is a critical leucine zipper
(bZIP) transcription factor that serves as a major regulator in
cellular defenses against reactive oxygen species (ROS) and
chemical detoxification to protect from oxidative damages. Under
normal conditions, Keap1 specifically recognizes Nrf2 (10) and constantly carries Nrf2 to
proteasomes for degradation (11,12).
Conversely, the activity of a Keap1-associated E3 ubiquitin ligase
is inhibited so that Nrf2 protein is activated and transfers from
the cytoplasm to the nucleus leading to nuclear accumulation under
oxidative stress. Nuclear Nrf2 increases the expression of
ARE-bearing genes including heme oxygenase-1 (HO-1), NAD(P)H
quinine oxidoreductase (NQO-1), glutamate-cysteine ligase (GCL),
glutathione (GSH) generation enzymes and GSH peroxidase (GPx), to
neutralize ROS and maintain a balance of cellular redox (13–15).
When autophagy is impaired, p62 forms a complex with
Keap1 and sequesters Keap1 into inclusion bodies, resulting in a
stabilization of Nrf2 protein and an induction of ARE-containing
genes. Therefore, p62 emerges as a linker molecule between
autophagy and oxidative stress (15).
The crosstalk between autophagy and the
Keap1/Nrf2/ARE axis was reported in 2010, and since then great
efforts have been made to investigate the underlying molecular
mechanisms, particularly in different types of cancers such as
hepatocellular carcinoma (16),
ovarian cancer (17) and lung
cancer (18). A previous study
demonstrated that the clinical drug verteporfin induced aggregation
of p62 and inhibited autophagy thus suppressing prostate
tumorigenesis in a xenograft model (19). However, the interplay between the
two signaling pathways remains largely unknown in PCa. Given that
p62 acts as a regulator of the Keap1/Nrf2 pathway, the previous
study hypothesized that p62 is involved in the Keap1/Nrf2/ARE
pathway and plays a critical role in the pathogenesis of mCRPC.
The present study demonstrated that p62 functioned
as a molecular regulator in the Keap1/Nrf2/ARE axis to promote the
transcription of Nrf2-mediated genes and decrease the levels of ROS
in PCa. Oxidative stresses generated under defective autophagy
trigger genome instability and tumorigenesis (20). Therefore, inhibiting p62 and Nrf2
activity may suppress carcinogenicity.
Materials and methods
Collection of animal and patient
tissue samples
In the present study, PTEN−/− mice and
transgenic adenocarcinoma of mouse prostate (TRAMP) mice were
obtained from Jackson Laboratory. All animal protocols were
approved by the Animal Care and Use Committee of the Texas A&M
Health Science Center at Houston (AUP IACUC 2015-0132-IBT). Normal
prostate tissues and prostate tumor tissues were collected from
three sets of 11-month-old wild-type, PTEN−/− and TRAMP
mice after mice were sacrificed by performing cervical dislocation
by appropriately trained personnel approved on the animal protocol.
A total of 150 patients including 112 patients with PCa and 38 with
BPH as controls were evaluated in the present study. As previously
described (21), these patients had
consented to donate prostate tissue samples from surgery or via
biopsy and underwent treatment including radiotherapy,
prostatectomy, transurethral resection of the prostate (TURP) and
hormone therapy between 1999 and 2003 at The Fifth Affiliated
Hospital of Guangzhou Medical University, Sun Yat-Sen Memorial
Hospital, and The First People's Hospital of Guangzhou and The
First Affiliated Hospital of Sun Yat-Sen University, Guangzhou,
China.
Immunohistochemistry analyses
All primary samples were fixed in 10% formalin,
embedded in paraffin, and stained with hematoxylin and eosin
(H&E). Paraffin-embedded, formalin-fixed prostate tissues of
patients and mice were immunostained for p62, Keap1 and Nrf2
protein using an immunohistochemical detection kit (GTVision)
according to the manufacturer's protocol. Immunostained slides were
independently evaluated under a Leica DM3000 microscope by two
pathologists in a double-blind manner.
Cell culture, transfection and siRNA. DU145
cells were obtained from the American Type Culture Collection
(ATCC); maintained in RPMI-1640 medium (HyClone Laboratories; GE
Healthcare Life Sciences) supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.), 100 units/ml penicillin
and 100 µg/ml streptomycin in a humidified incubator at 37°C with
5% CO2; and sub-cultured for a passage every 3–4 days.
Stable DU145 cell strains expressing different levels of p62 were
constructed as previously described: C1 carrying the control
plasmid vector, OE overexpressing p62, KO with p62 knocked out by
CRISPER technology and C2 negative control of KO (22). Validated siRNAs against Keap1 and
Nrf2 were purchased from Guangzhou Ribobio Co., Ltd. Cells were
treated with 10 mM bafilomycin A1 (BAF) or 10 mM MG-132 for 6 h
when necessary.
Immunoblotting analyses
Whole-cell extracts were prepared with RIPA buffer
containing the broad protease inhibitor cocktail (Roche
Diagnostics). Nucleoprotein extraction was prepared with NE-PER
Nuclear Cytoplasmic Extraction Reagent kit (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
protein concentration was determined using a BCA kit. Equal amounts
of total proteins (25 µg) were separated via 10% SDS-PAGE gel and
transferred to PVDF membranes. Then, 5% skimmed milk (Devondale
Milk) was used to incubate at room temperature for 2 h. Then the
membranes were blotted with primary antibodies against p62 (product
code ab56416, dilution 1:2,500), Keap1 (product code ab218815,
dilution 1:1,500) and Nrf2 (product code ab194986, dilution
1:1,000; all were from Abcam), Lamin B1 (cat. no. sc-374015,
dilution 1:1,000; Santa Cruz Biotechnology, Inc.) or GAPDH (cat.
no. AF0911, dilution 1:4,000; Affinity Biosciences), all the
primary antibodies were incubated at 4°C overnight. Protein bands
were visualized using their respective goat anti-rabbit IgG (H+L)
(dilution 1:1,000; incubated at room temperature for an 1 h) or
goat anti-mouse IgG (H+L) (dilution 1:1,000; incubated at room
temperature for an 1 h) (Wuhan Boster Biological Technology, Ltd.)
and ECL reagent (Thermo Fisher Scientific, Inc.).
ImageJ® software (v1.52p; National Institutes of Health)
was used for densitometric analysis.
Confocal fluorescent microscopy
Confocal fluorescent microscopy was performed as
previously described (22).
Briefly, cells were grown on glass coverslips in 6-well plates for
6 h, washed three times with PBS, fixed in 4% paraformaldehyde,
permeabilized in 0.1% Triton X-100, and incubated with 2% BSA for 1
h at room temperature. Treated cells were first blotted with an
antibody against Keap1 (product code ab139729) or Nrf2 (product
code EP1808Y; both from Abcam) for 16 h at 4°C, and subsequently
incubated with DYLight649-conjugated goat anti-rabbit IgG (H+L)
secondary antibody (cat. no. GAR6492; Multi Sciences) for 1 h at
room temperature. After washing with PBS, coverslips were mounted
with Fluoromount-G™ (Invitrogen; Thermo Fisher Scientific, Inc.)
and visualized under a confocal laser scanning microscope (Zeiss)
at an ×20 magnification.
Reverse transcription-quantitative PCR
(RT-qPCR) analyses
Total RNA was isolated using the UNlQ-10 Column
TRIzol Total RNA Isolation kit (Sangon Biotech Co., Ltd.) according
to the manufacturer's protocol, and reverse-transcribed to cDNA
using the PrimeScript™ RT reagent kit (Takara Bio, Inc.). The
conditions used were as follows: 30 sec 95°C, followed by 50 cycles
of 5 sec 95°C, 34 sec 60°C, then 15 sec 95°C, and 1 min 50°C. The
gene-specific primers presented in Table I were obtained from PrimerBank.
Differential expression was calculated using the 2−ΔΔCq
method as described (23). Data are
presented as the mean ± standard deviation of three independent
experiments.
 | Table I.The sequences of primers used for
qPCR experiments. |
Table I.
The sequences of primers used for
qPCR experiments.
| Primer | Sequence |
|---|
| hKeap1 | F:
GCTGATGAGGGTCACCAGTT |
|
| R:
CCAACTTCGCTGAGCAGATT |
| hNrf2 | F:
GCTCATACTCTTTCCGTCGC |
|
| R:
ATCATGATGGACTTGGAGCTG |
| hNQO-1 | F:
GCACTGATCGTACTGGCTCA |
|
| R:
GAACACTCGCTCAAACCAG |
| hGCLC | F:
GGATGATGCTAATGAGTCTGACC |
|
| R:
TCTACTCTCCATCCAATGTCTGAG |
| HO-1 | F:
CCAGGCAGAGAATGCTGAGTTC |
|
| R:
AAGACTGGGCTCTCCTTGTTGC |
| hGCLM | F:
TTGGGAACTCCATTCATTCA |
|
| R:
CGGGAACCTGCTCAACTG |
| GAPDH | F:
GAAGGTGAAGGTCGGAGTC |
|
| R:
GAAGATGGTGATGGGATTTC |
Apoptosis analyses
The Annexin V-APC/7AAD Apoptosis Detection system
was used to measure rates of apoptosis according to the
manufacturer's protocol. Briefly, cells were cultured in RPMI-1640
medium in 10-cm dishes to a confluence of 80–90%, digested with
0.25% trypsin without EDTA, re-suspended in 500 µl of binding
buffer supplied with 5 µl APC-Annexin V and 10 µl 7AAD, and
incubated at room temperature for 15 min after mixing. The
apoptosis rate was detected using BD Accuri C6 flow cytometer and
analyzed with the associated software from BD Biosciences.
Cell proliferation assays
The rates of cell proliferation were measured using
the Real Time Cellular Analysis (RTCA) system. Briefly, cells were
harvested at 48 h after infection with siRNA, diluted and seeded
into a 16-well strip at a density of 1×104 cells/well.
Cell proliferation was monitored in real time with sensor devices
placed in the incubator with 5% CO2.
Cell invasion assays
Cell invasion was continuously monitored every 15
min for 36 h by real-time monitoring on CIM-Plate 16 pre-coated
with Matrigel (356234; Corning Inc.) with an RTCA iCELLigence
Analyzer (ACEA Biosciences). Briefly, the infected cells were
starved for 6 h in serum-starved medium, diluted and seeded into a
16-well strip at a density of 3×105 cells/well in the
upper chamber. The lower chamber was filled with medium containing
10% serum.
ROS detection
The detection of ROS was performed using the Cell
ROX Orange reagent (Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The cell-permeable reagents are
non-fluorescent or weakly fluorescent in a reduced state, and emit
strong fluorescence signals upon oxidation. DU145 cells in a 6-well
plate were incubated in medium containing a final concentration of
5 µM CellROX Orange reagent at 37°C for 30 min and washed three
times with pre-warmed PBS after the medium was removed. Levels of
ROS of cells were quantified using ImageJ software (v1.52p;
National Institutes of Health) based on the fluorescence
intensities of cells in captured fields.
Statistical analysis
Experimental data were presented as the mean ±
standard error of at least three independent experiments.
Differences in results were analyzed by a two-tailed unpaired
t-test using the GraphPad Prism version 7 for Windows (GraphPad
Software, Inc.). P<0.05, P<0.01, and P<0.001 were
considered to indicate a statistically significant result
(*P<0.05, **P<0.01 and ***P<0.001 as indicated in the
figures).
Results
Expression of P62, Keap1 and Nrf2 in
prostate tissues
It has previously been reported that PCa tissues
have higher levels of cytoplasmic p62 than benign prostate
hyperplasia (BPH) tissues although p62 whose significance remains
unknown was also observed to be distributed in nuclei. In addition,
PCa tissues from patients with higher grades or metastasis have
significantly higher levels of p62 than those from patients with
lower grades or no metastasis (20). The present study further
demonstrated that p62 was primarily located in the cytoplasm of
epithelial cells of human PCa and BPH tissues (Fig. 1A-C). Keap1 was also primarily
located in the cytoplasm of epithelial cells of human PCa and BPH
tissues (Fig. 1D and E) while Nrf2
was detected in both the nucleus and cytoplasm (Fig. 1F and G). The levels of p62 and Nrf2
were higher in human PCa than those in BPH (Fig. 1A-C, F and G), while the levels of
Keap1 were higher in BPH than those in PCa (Fig. 1D and E). Similarly, higher levels of
p62 and Nrf2 and lower levels of Keap1 were detected in PCa tissues
collected from PTEN−/− and TRAMP mice compared with
normal prostate tissues collected from normal wild-type mice
(Fig. 1H-P). The results indicated
that p62 and Nrf2 may act as oncogenes of PCa.
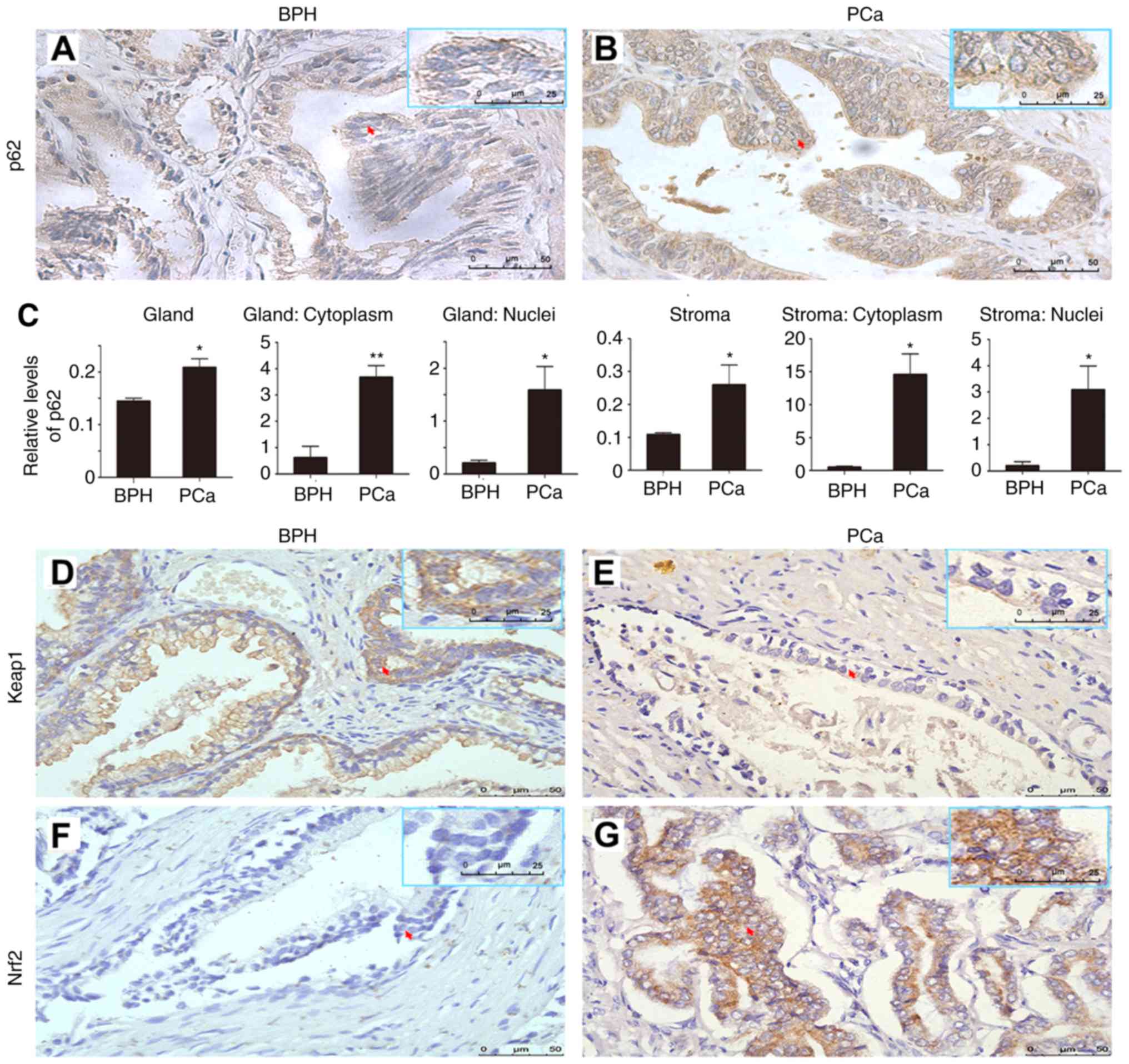 | Figure 1.The expression of p62, Keap1 and
Nrf2. (A and B) Representative images and (C) quantification
revealing the levels of p62 in (A) BPH and (B) PCa. A total of 150
patients including 112 patients with PCa and 38 with BPH were
evaluated. The relative levels of p62 in prostate glands and
stromas and in cytoplasm and nuclei were compared between BPH and
PCa. *P≤0.05 and **P≤0.01 Representative images showing (D) high
levels of Keap1 in BPH, or (E) low levels of Keap1 in PCa, and (F)
low levels of Nrf2 in BPH, or (G) high levels of Nrf2 in PCa.
Representative images showing (H) low or (I and J) high levels of
p62 expressed in prostate tissues from (H) normal, (I)
PTEN−/−, or (J) TRAMP mice; (K) high, or (L and M) low
levels of Keap1 expressed in (K) prostate tissues from normal, (L)
PTEN−/−, or (M) TRAMP mice; and (N) low, or (O and P)
high levels of Nrf2 expressed in (N) prostate tissues from normal,
(O) PTEN−/−or (P) TRAMP mice. Normal prostate tissues
and prostate tumors were collected from three sets of wild-type,
PTEN−/− and TRAMP mice. Scale bar, 50 µm. Keap1,
Kelch-like ECH-associated protein 1; Nrf2, nuclear factor
erytheroid-derived 2-like 2; BPH, benign prostate hyperplasia; PCa,
prostate cancer; TRAMP, transgenic adenocarcinoma of mouse
prostate. |
p62 suppresses the expression of Keap1
and enhances the activity of Nrf2
To examine the association between the levels of
p62, Keap1 and Nrf2 proteins, the present study prepared extracts
from four different DU145 cell strains expressing different levels
of p62. Increasing the expression levels of p62 significantly
decreased the levels of Keap1 and enhanced the levels of total Nrf2
and nuclear Nrf2 (Fig. 2A-H),
respectively. In contrast, decreasing the expression level of p62
significantly increased levels of Keap1 and reduced levels of total
Nrf2 and nuclear Nrf2 (Fig. 2I-P),
respectively. In addition, the effects of p62 on the levels of
Keap1 and total Nrf2 were further confirmed by immunofluorescence
microscopy analyses (Fig. 2Q-T).
Therefore, p62 suppressed the levels of Keap1 and enhanced the
levels of Nrf2.
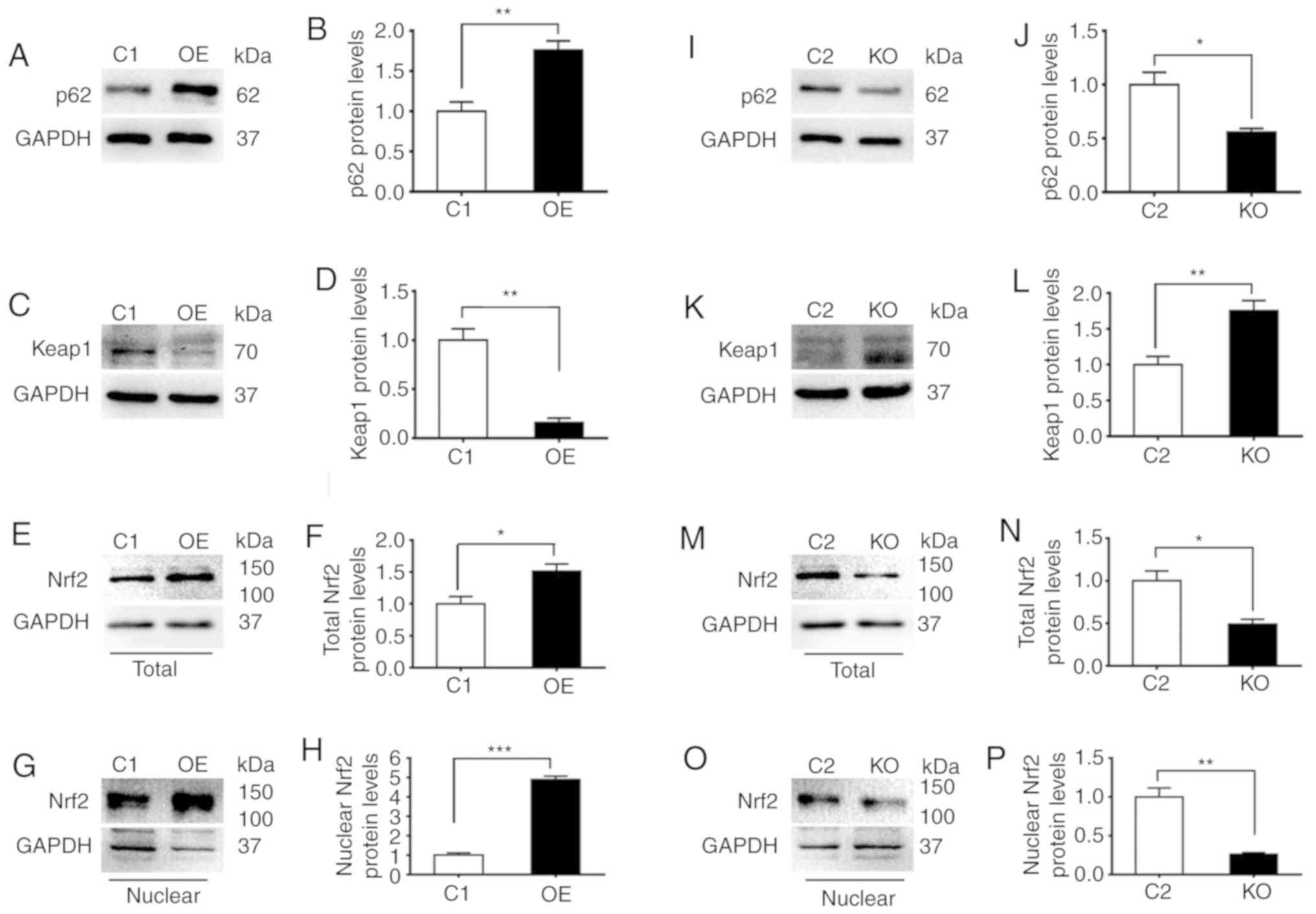 | Figure 2.p62 suppresses the expression of
Keap1 and enhances the activity of Nrf2. C1, DU145 cells carrying
the control plasmid vector; OE, DU145 cells overexpressing p62; KO,
OE cells with p62 knocked out by CRISPER technology; and C2, OE
cells carrying negative control for CRISPR knockout. (A-P)
Representative (A, C, E, G, I, K, M and O) immunoblots and (B, D,
F, H, J, L, N and P) quantification showing levels of (A, B, I and
J) p62, (C, D, K and L) Keap1, and (E, F, M and N) total and (G, H,
O and P) nuclear Nrf2 in (A-H) C1 and OE cells and (I-P) C2 and KO
cells. (Q-T) Representative images revealing the (Q and R)
immunostaining intensities and (S and T) quantification of (Q and
S) Keap1 and (R and T) Nrf2 in C1, OE, C2 and KO cells. The nuclei
were stained with DAPI (blue). All images were captured under an
identical setting. The results represent three independent
experiments. *P≤0.05, **P≤0.01, and ***P≤0.001. Keap1, Kelch-like
ECH-associated protein 1; Nrf2, nuclear factor erytheroid-derived
2-like 2. |
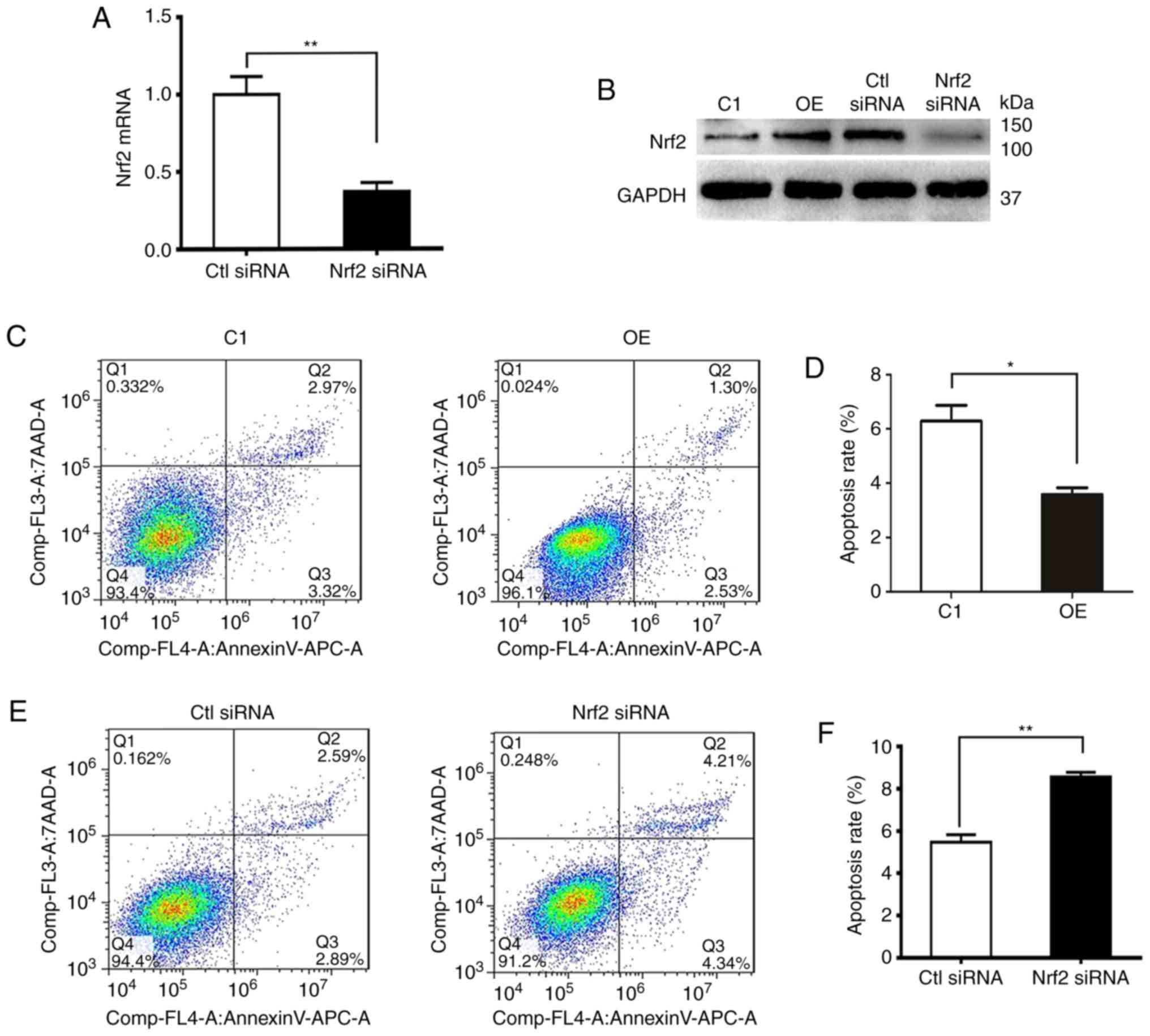
p62 decreases apoptosis and promotes
proliferation and invasion of DU145 cells through Nrf2
In order to determine whether aberrant Nrf2 activity
was associated with apoptosis, proliferation and invasion in
vitro, DU145 cells overexpressing p62 (OE) were transfected
with negative control siRNA (Ctl siRNA) and Nrf2-specific siRNA
(Nrf2 siRNA). A knockdown efficiency of 80% was confirmed by qPCR
and immunoblotting (Fig. 3A and B).
Flow cytometric analyses of cell apoptosis revealed that DU145
cells overexpressing p62 (OE) had lower rates of apoptosis than its
control C1 (Fig. 3C and D), while
silencing the expression of Nrf2 in OE cells induced prominently
higher rates of apoptosis (Fig. 3E and
F). In contrast, the overexpression of p62 led to increases in
cell proliferation and invasion, while further decreasing the
increased levels of Nrf2 expression with siRNA led to decreases in
cell proliferation and invasion (Fig.
3G and H). Notably, rates of cell proliferation and invasion
between OE and Ctl siRNA cells were different potentially due to
the impact of control siRNA (Fig. 3G
and H). Therefore, p62 decreased apoptosis and promoted
proliferation and invasion of DU145 cells through Nrf2.
p62 increases the expression levels of
Nrf2-targeted genes and suppresses ROS
In order to investigate the mechanism by which p62
enhances proliferation, invasion and anti-apoptosis, RT-qPCR was
used to assess the mRNA levels of Nrf2 target genes such as HO-1,
NQO-1, GCLC and glutamate-cysteine ligase modifier subunit (GCLM).
It was revealed that p62 increased the mRNA levels of Nrf2 target
genes (Fig. 4A-D). Overexpression
of p62 led to a decrease in ROS levels, while suppressing the
expression of p62 induced an increase in ROS levels (Fig. 4E and F). Thus, p62 increased the
expression of antioxidant-responding genes and suppressed ROS.
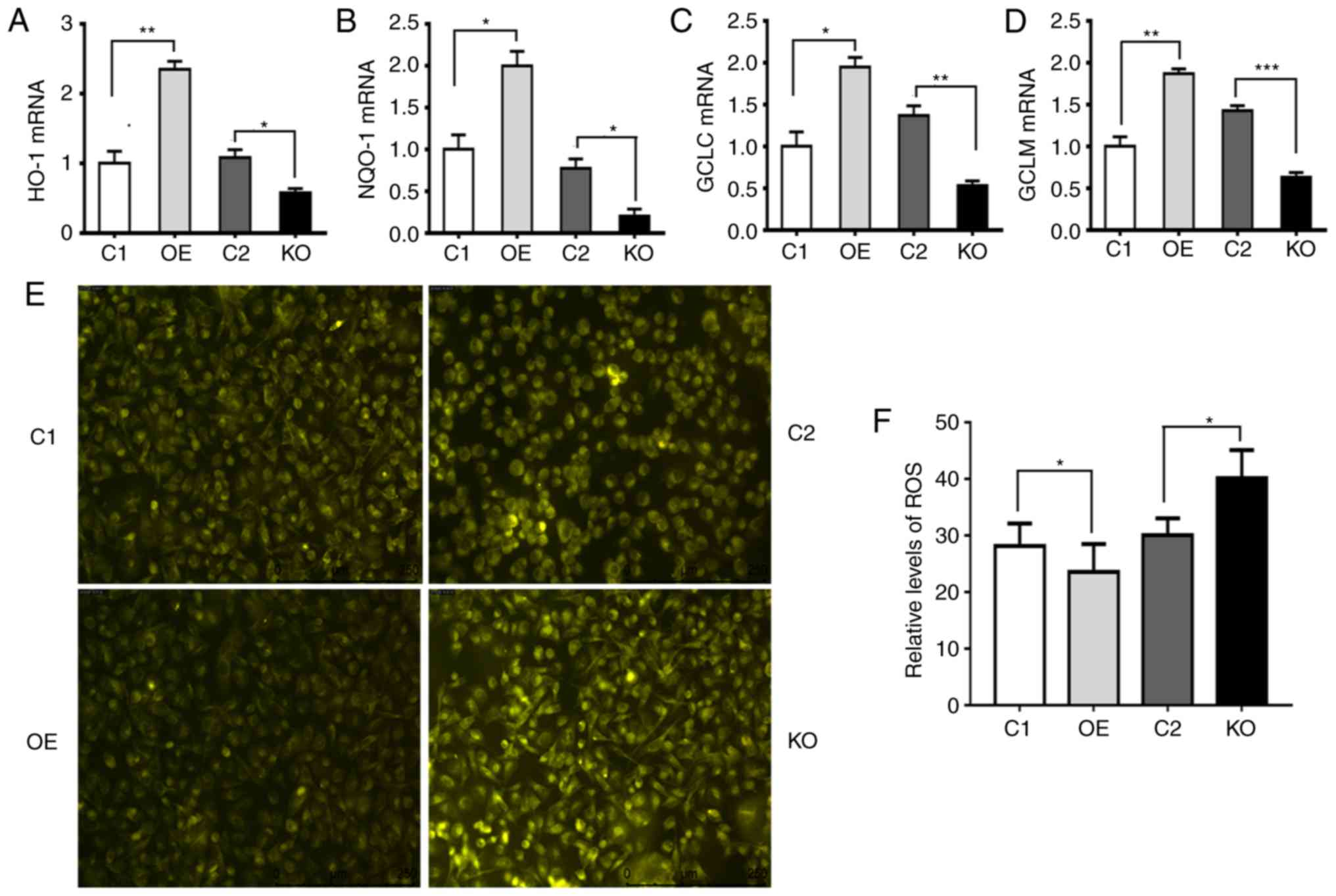 | Figure 4.p62 increases the expression of
Nrf2-targeted genes and decreases the levels of ROS. C1, DU145
cells carrying the control plasmid vector; OE, DU145 cells
overexpressing p62; KO, OE cells with p62 knocked out by CRISPER
technology; and C2, OE cells carrying negative control for CRISPR
knockout. (A-D) Plots revealing the mRNA levels of (A) HO-1, (B)
NQO-1, (C) GCLC and (D) GLCM in C1, OE, C2 and KO cells. Values
were normalized to GAPDH. (E and F) Representative (E) images and
(F) quantification revealing the levels of cellular ROS reflected
by the intensities of fluorescence in C1, OE, C2 and KO cells. Data
represent the mean ± standard error of the mean of readings from
>8 fields of views. *P<0.05, **P<0.01, ***P<0.001.
Nrf2, nuclear factor erytheroid-derived 2-like 2; HO-1, heme
oxygenase-1; NQO-1, NAD(P)H quinine oxidoreductase; GCLC,
glutamate-cysteine ligase catalytic subunit; GLCM,
glutamate-cysteine ligase modifier subunit; ROS, reactive oxygen
species. |
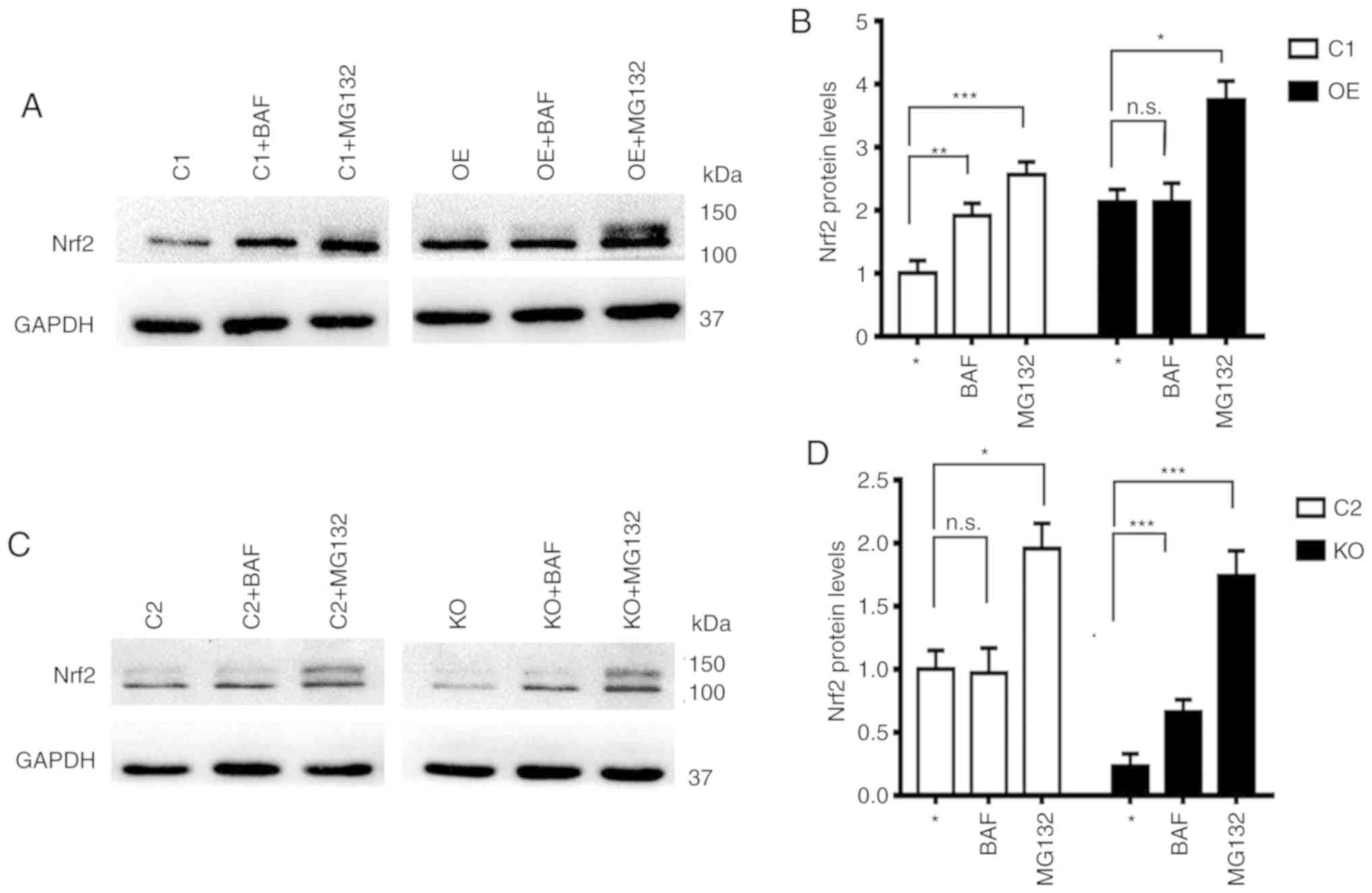
Nrf2 is degraded through the
proteasomal system independent of the p62 levels but the autophagy
system in the absence of p62
Numerous studies have demonstrated that Nrf2 protein
is degraded through the a proteasomal system (13,24,25).
It was revealed in the present study that the levels of Nrf2 in the
presence of proteasomal inhibitor MG132 were always increased
independent of the levels of p62, while levels of Nrf2 in the
presence of autophagy inhibitor BAF were only increased in cells
with low levels of p62 (C1 and KO), but remained constant in cells
with high levels of p62 (C2 and OE) (Fig. 5). These results indicated that some
portions of Nrf2 protein are always degraded through the
proteasomal system but some portions of the Nrf2 protein that are
modified differentially in the absence of p62 are degraded through
the autophagy system.
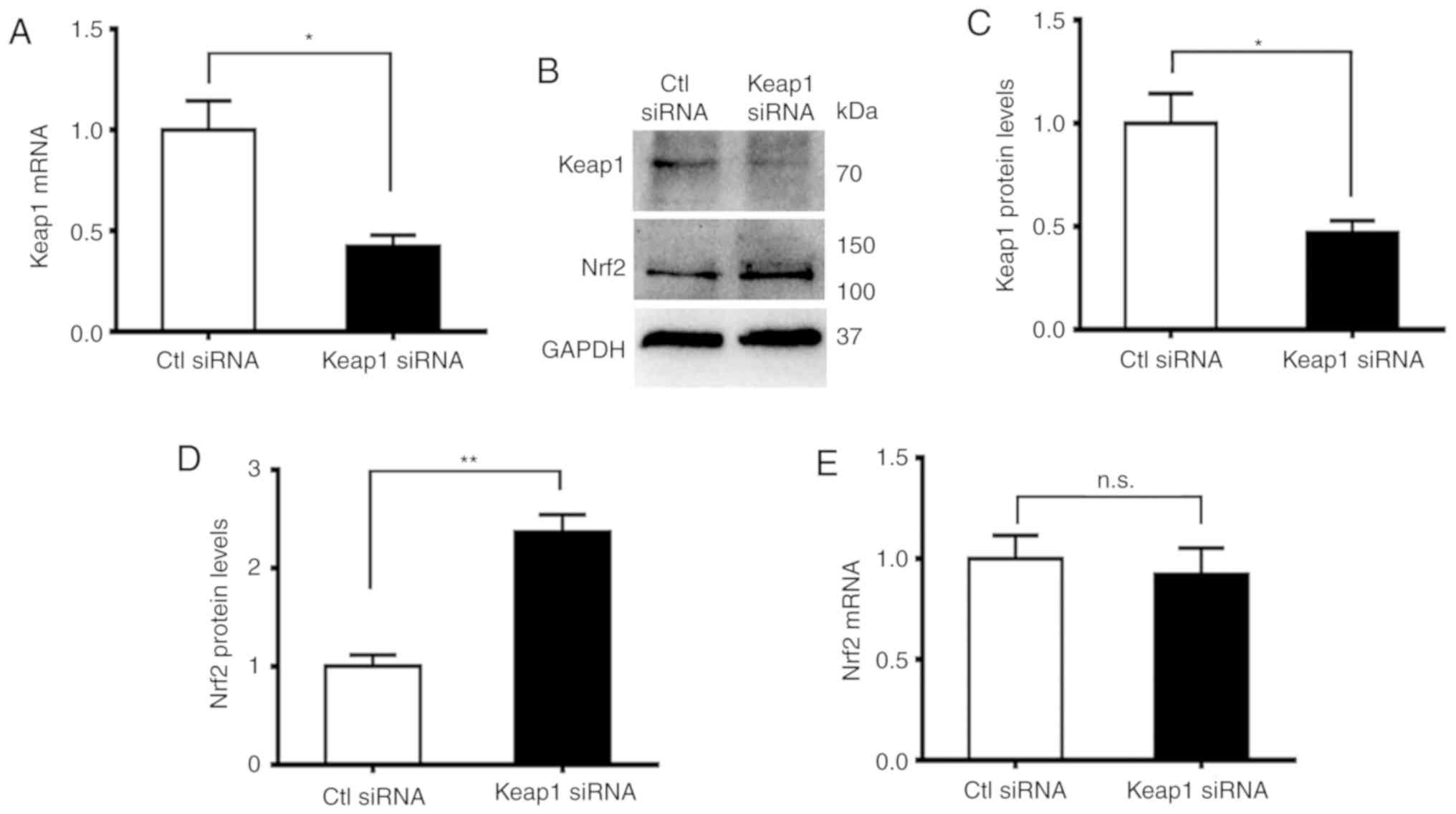
p62 regulates Nrf2 through Keap1
In order to investigate the association between
Keap1 and Nrf2, the expression of Keap1 in DU145 cells with p62
knocked down was suppressed. The levels of Nrf2 were increased
(Figs. 2G and H and 6A). The suppression of Keap1 was confirmed
in both mRNA (Fig. 6A) and protein
levels (Fig. 6B and C). Such
suppression caused a significant increase in the levels of Nrf2
protein (Fig. 6B and D) but had no
impact on the levels of Nrf2 mRNA (Fig.
6E). Therefore, p62 increased the level of Nrf2 through Keap1
in a post-transcriptional manner.
Discussion
It has previously been demonstrated that p62
functions as an oncogene in 73% of PCa cases, particularly at the
advanced stage of disease (26).
However, the mechanism underlying the carcinogenicity of P62
remains unclear in PCa. The present study revealed new evidence
that the p62/Keap1/Nrf2 pathway is a crucial regulator of PCa by
transcriptional inhibition of genes involved in regulating ROS. p62
was positively correlated with total Nrf2, particularly nuclear
Nrf2, and negatively correlated with Keap1. Notably, high levels of
p62 were crucial for Nrf2 activity in association with cell
proliferation, apoptosis and invasion. Inhibiting the
p62/Keap1/Nrf2 pathway significantly suppressed PCa development
both in vitro and in vivo.
There are multiple mechanisms that regulate the
Keap1/Nrf2/ARE pathway. For example, p62, a substrate and an
adaptor for selective autophagy (27) and a linker between autophagy and
oxidative stress (15), could be
involved in the regulation of this signaling pathway. Autophagy
deficiencies cause the accumulation of p62 and the formation of
aggregates (28). p62 blocks the
interaction between Keap1 and Nrf2 and promotes nuclear
translocation of Nrf2 to regulate a battery of genes encoding
antioxidant proteins (15,29). Increased levels of both p62 and Nrf2
were observed in PCa cells and PCa tissues from PTEN−/−
and TRAMP mice, while decreased levels of p62 led to the
accumulation of Nrf2. Collectively, these studies provide evidence
to suggest that the accumulation of p62 and prolonged activation of
Nrf2 activity play a critical role in the pathogenesis of numerous
human diseases including cancers.
The Keap1-Nrf2-ARE pathway is a key mechanism of
cellular defense that helps maintain the homeostasis of cellular
redox (30). The activation of the
Keap1/Nrf2/ARE pathway leads to the activation of diverse
antioxidant and phase II detoxifying enzymes, including NQO-1,
HO-1, GCLC and GCLM, to cope with oxidative stress. It was reported
that the suppression of p62 prevents UVA-induced production of ROS
potentially as p62 induction disrupts mitochondrial function
leading to increased production of ROS (31). Notably, it was revealed in the
present study that decreasing the levels of p62 in DU145 cells
resulted in an increase in basal levels of ROS, while high levels
of p62 caused a decrease in ROS levels. The role of ROS in cancers
has been under debate for decades. Recent studies have revealed
that low levels of ROS induced the activation of pathways
regulating proliferation and survival of cells (32), moderately increased levels of ROS
caused DNA damage and promoted mutagenesis in cells (33), but high levels of ROS could
ultimately cause cell senescence or death (34). It was speculated in the present
study that the decrease in the levels of ROS caused by p62
overexpression through the Keap1/Nrf2/ARE pathway was more
significant than the increase of ROS levels caused by p62
overexpression through the inhibition of the autophagy pathway and
the induction of mitochondrial dysfunction. Therefore, p62 may
induce low levels of ROS to promote the progression and survival
pathways of tumor cells.
It has been reported that overexpressed Keap1 is
degraded through autophagy (35)
and the degradation of Keap1 depends on a direct physical
interaction between Keap1 and p62 (13). In addition, it has been reported
that Nrf2 is degraded through the proteasomal system (35,36).
The present study confirmed that the degradation of Nrf2 protein is
always through the proteasomal system. However, Nrf2 is degraded
through the autophagy system in the absence of p62.
It was previously demonstrated that p62 enhances the
epithelial-mesenchymal transition (EMT) and promotes development of
PCa (22). The present study
further revealed that p62 increased the rates of proliferation and
invasion and decreased the rates of apoptosis of PCa cells through
the p62/Keap1/Nrf2 pathway. Once the activity of Nrf2 was
inhibited, the overexpression of p62 led to an enhancement of
apoptosis and a decrease in the rates of cell proliferation and
invasion. Therefore, Nrf2 was revealed to be a key mediator for p62
to exert its oncogenetic impact.
Epidemiological studies have attributed obesity as
an important risk factor of PCa progression and poor outcome of
those patients diagnosed, and one recent study demonstrated that
the underlying molecular mechanism was associated with p62 activity
in adipocytes (37,38). The deletion of p62 in adipocytes was
revealed to induce EMT and promote PCA aggressiveness independent
of the differences in locomotor activity and food intake in mice.
It was previously reported that p62 enhanced EMT of PCa, and the
present study further demonstrated that Nrf2 is a key regulator of
such a phenomenon. Nrf2 is reported to be involved in
obesity-induced oxidative stress and suggested to be targeted to
decrease obesity-triggered oxidative stress damage (39). However, obesity has been
characterized by significantly-activated oxidative stress and
increased levels of autophagy flux, which have been demonstrated to
be detrimental for the male reproductive system (40). p62 in combination with Nrf2 was
revealed to be beneficial for sustaining anti-oxidation in
adipocytes, while p62 deletion in adipocytes leads to an
upregulation of oxidative stress that may cause EMT in PCa. Since
Nrf2 is degraded through the autophagy system in the absence of
p62, the degradation of Nrf2 is accordingly enhanced in
p62-deficient adipocytes where autophagy is enhanced due to p62
inactivation. Collectively, these results contribute to the
progression of PCa, such as EMT. It has previously been reported
that high levels of p62 caused a promotion of EMT, in addition to
an impairment of autophagy flux in PCa. The present study has
further demonstrated that high levels of p62 suppressed ROS in PCa
through the anti-oxidative Keap1/Nrf2/ARE pathway. This is
consistent with a previous study which revealed that p62 and Nrf2
could cooperatively regulate antioxidants (41).
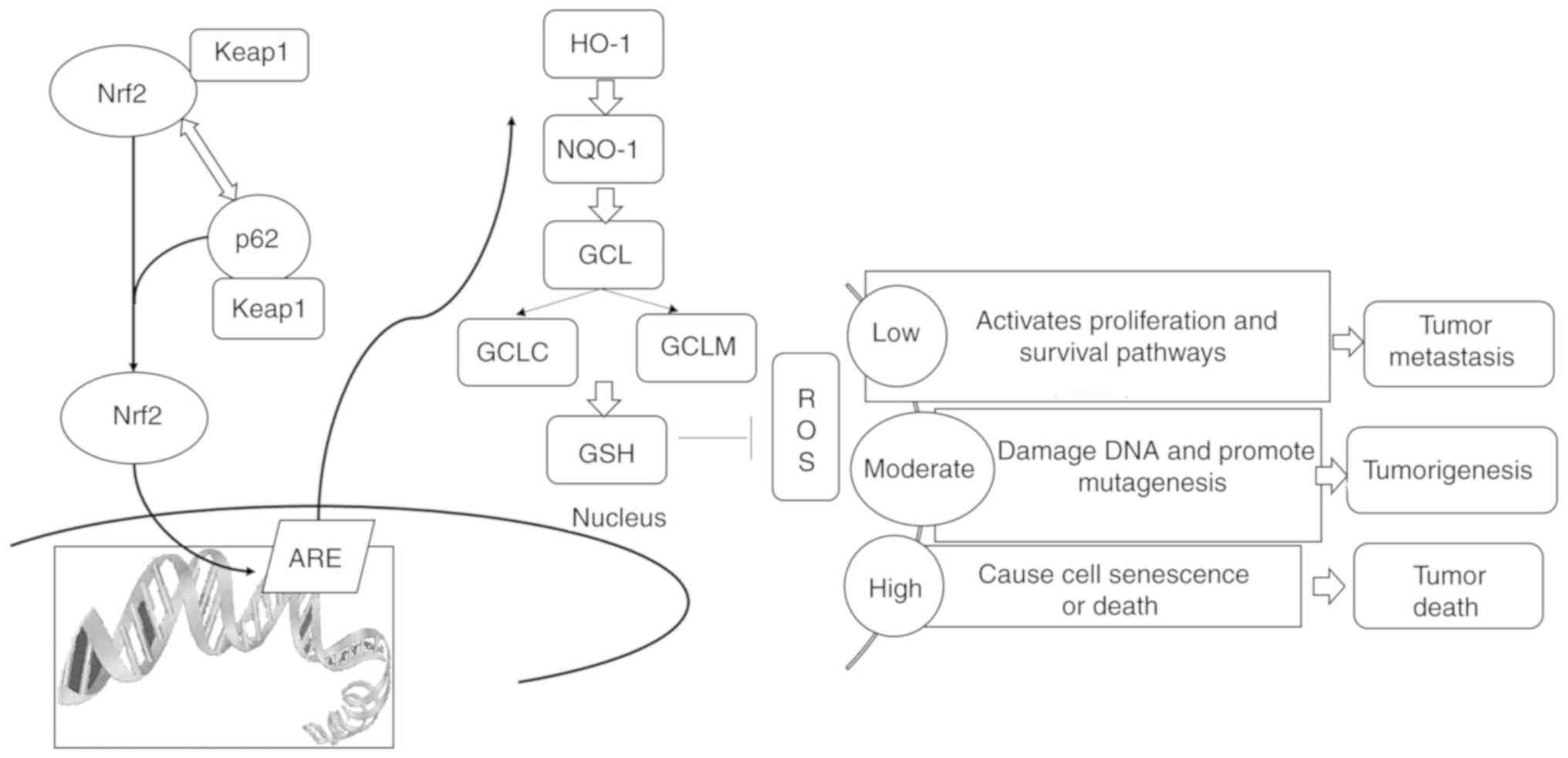
In conclusion, p62 interacted with Keap1 to prevent
the degradation of Nrf2 through the proteasomal system. In the
absence of p62, a portion of Nrf2 was delivered to the autophagy
machinery to be degraded. p62 suppressed the Keap1/Nrf2/ARE system
and induced low levels of ROS to promote PCa metastasis (Fig. 7). Therefore, p62 inhibitors or Nrf2
inhibitors may be utilized to precisely treat patients with PCa in
the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Guangzhou
Municipal Science and Technology Project (grant no. 1563000448),
the National Natural Science Foundation of China (grant no.
81772931) and the National Cancer Institute of National Institutes
of Health (grant no. CA142862).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
GJ, XL, YH, ZL, ZZ, ZS, ZF, YL, WY, TL, LH, FW and
HH performed the experiments. GJ, XL and YH wrote the original
draft of the manuscript. LL and XJ conceived, designed and directed
the project and reviewed the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The collection of human prostate samples was
approved by the Fifth Affiliated Hospital of Guangzhou Medical
University, Sun Yat-sen Memorial Hospital, the Guangzhou First
People's Hospital and the First Affiliated Hospital of Sun Yat-sen
University in Guangzhou, China. Animal protocols were approved by
the Animal Care and Use Committee of the Texas A&M Health
Science Center at Houston (AUP IACUC 2015-0132-IBT).
Patient consent for publication
Not applicable.
Competing interests
The authors have declared that they have no
competing interests.
References
|
1
|
DeSantis CE, Siegel RL, Sauer AG, Miller
KD, Fedewa SA, Alcaraz KI and Jemal A: Cancer statistics for
African Americans, 2016: Progress and opportunities in reducing
racial disparities. CA Cancer J Clin. 66:290–308. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Bono JS, Logothetis CJ, Molina A,
Fizazi K, North S, Chu L, Chi KN, Jones RJ, Goodman OB Jr, Saad F,
et al: Abiraterone and increased survival in metastatic prostate
cancer. New Engl J Med. 364:1995–2005. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Scher HI, Fizazi K, Saad F, Taplin ME,
Sternberg CN, Miller K, de Wit R, Mulders P, Chi KN, Shore ND, et
al: Increased survival with enzalutamide in prostate cancer after
chemotherapy. New Engl J Med. 367:1187–1197. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sanchez P, De Carcer G, Sandoval IV,
Moscat J and Diaz-Meco MT: Localization of atypical protein kinase
C isoforms into lysosome-targeted endosomes through interaction
with p62. Mol Cell Biol. 18:3069–3080. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moscat J, Karin M and Diaz-Meco MT: p62 in
Cancer: Signaling Adaptor Beyond Autophagy. Cell. 167:606–609.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jaramillo MC and Zhang DD: The emerging
role of the Nrf2-Keap1 signaling pathway in cancer. Gene Dev.
27:2179–2191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suzuki T and Yamamoto M: Molecular basis
of the Keap1-Nrf2 system. Free Rad Biol Med. 88:93–100. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kansanen E, Kuosmanen SM, Leinonen H and
Levonen AL: The Keap1-Nrf2 pathway: Mechanisms of activation and
dysregulation in cancer. Redox Biol. 1:45–49. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
O'Connell MA and Hayes JD: The Keap1/Nrf2
pathway in health and disease: From the bench to the clinic.
Biochem Soc Trans. 43:687–689. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hayes JD and Dinkova-Kostova AT:
Oncogene-stimulated congestion at the KEAP1 stress signaling hub
allows bypass of NRF2 and induction of NRF2-target genes that
promote tumor survival. Can Cell. 32:539–541. 2017. View Article : Google Scholar
|
|
11
|
Furukawa M and Xiong Y: BTB protein Keap1
targets antioxidant transcription factor Nrf2 for ubiquitination by
the Cullin 3-Roc1 ligase. Mol Cell Bol. 25:162–171. 2005.
View Article : Google Scholar
|
|
12
|
McMahon M, Itoh K, Yamamoto M and Hayes
JD: Keap1-dependent proteasomal degradation of transcription factor
Nrf2 contributes to the negative regulation of antioxidant response
element-driven gene expression. J Biol Chem. 278:21592–21600. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jain A, Lamark T, Sjottem E, Larsen KB,
Awuh JA, Overvatn A, McMahon M, Hayes JD and Johansen T: p62/SQSTM1
is a target gene for transcription factor NRF2 and creates a
positive feedback loop by inducing antioxidant response
element-driven gene transcription. J Biol Chem. 285:22576–22591.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Komatsu M, Kurokawa H, Waguri S, Taguchi
K, Kobayashi A, Ichimura Y, Sou YS, Ueno I, Sakamoto A, Tong KI, et
al: The selective autophagy substrate p62 activates the stress
responsive transcription factor Nrf2 through inactivation of Keap1.
Nat Cell Biol. 12:213–223. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lau A, Wang XJ, Zhao F, Villeneuve NF, Wu
T, Jiang T, Sun Z, White E and Zhang DD: A noncanonical mechanism
of Nrf2 activation by autophagy deficiency: Direct interaction
between Keap1 and p62. Mol Cell Biol. 30:3275–3285. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Inami Y, Waguri S, Sakamoto A, Kouno T,
Nakada K, Hino O, Watanabe S, Ando J, Iwadate M, Yamamoto M, et al:
Persistent activation of Nrf2 through p62 in hepatocellular
carcinoma cells. J Cell Biol. 193:275–284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xia M, Yu H, Gu S, Xu Y, Su J, Li H, Kang
J and Cui M: p62/SQSTM1 is involved in cisplatin resistance in
human ovarian cancer cells via the Keap1-Nrf2-ARE system. Intern J
Oncol. 45:2341–2348. 2014. View Article : Google Scholar
|
|
18
|
Wang Y, Zhang J, Huang ZH, Huang XH, Zheng
WB, Yin XF, Li YL, Li B and He QY: Isodeoxyelephantopin induces
protective autophagy in lung cancer cells via Nrf2-p62-keap1
feedback loop. Cell Death Dis. 8:e28762017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang L, Kim D, Wise JTF, Shi X, Zhang Z
and DiPaola RS: p62 as a therapeutic target for inhibition of
autophagy in prostate cancer. Prostate. 78:390–400. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang X, Zhong W, Huang H, He H, Jiang F,
Chen Y, Yue F, Zou J, Li X, He Y, et al: Autophagy defects
suggested by low levels of autophagy activator MAP1S and high
levels of autophagy inhibitor LRPPRC predict poor prognosis of
prostate cancer patients. Mol Carcinog. 54:1194–1204. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang X, Li X, Huang H, Jiang F, Lin Z, He
H, Chen Y, Yue F, Zou J, He Y, et al: Elevated levels of
mitochondrion-associated autophagy inhibitor LRPPRC are associated
with poor prognosis in patients with prostate cancer. Cancer.
120:1228–1236. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang X, Huang Y, Liang X, Jiang F, He Y,
Li T, Xu G, Zhao H, Yang W, Jiang G, et al: Metastatic prostate
cancer-associated P62 inhibits autophagy flux and promotes
epithelial to mesenchymal transition by sustaining the level of
HDAC6. Prostate. 78:426–434. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kobayashi A, Kang MI, Okawa H, Ohtsuji M,
Zenke Y, Chiba T, Igarashi K and Yamamoto M: Oxidative stress
sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to
regulate proteasomal degradation of Nrf2. Mol Cell Biol.
24:7130–7139. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Taguchi K, Motohashi H and Yamamoto M:
Molecular mechanisms of the Keap1-Nrf2 pathway in stress response
and cancer evolution. Gene Cell. 16:123–140. 2011. View Article : Google Scholar
|
|
26
|
Duran A, Linares JF, Galvez AS,
Wikenheiser K, Flores JM, Diaz-Meco MT and Moscat J: The signaling
adaptor p62 is an important NF-kappaB mediator in tumorigenesis.
Can Cell. 13:343–354. 2008. View Article : Google Scholar
|
|
27
|
Moscat J and Diaz-Meco MT: p62: A
versatile multitasker takes on cancer. Trend Biochem Sci.
37:230–236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang DD: Mechanistic studies of the
Nrf2-Keap1 signaling pathway. Drug Metab Rev. 38:769–789. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ichimura Y, Waguri S, Sou YS, Kageyama S,
Hasegawa J, Ishimura R, Saito T, Yang Y, Kouno T, Fukutomi T, et
al: Phosphorylation of p62 activates the Keap1-Nrf2 pathway during
selective autophagy. Mol Cell. 51:618–631. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lau A, Villeneuve NF, Sun Z, Wong PK and
Zhang DD: Dual roles of Nrf2 in cancer. Pharmacol Res. 58:262–270.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sample A, Zhao B, Wu C, Qian S, Shi X,
Aplin A and He YY: The autophagy receptor adaptor p62 is
Up-regulated by UVA radiation in melanocytes and in melanoma cells.
Photochem Photobiol. 94:432–437. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sena LA and Chandel NS: Physiological
roles of mitochondrial reactive oxygen species. Mol Cell.
48:158–167. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wiseman H and Halliwell B: Damage to DNA
by reactive oxygen and nitrogen species: Role in inflammatory
disease and progression to cancer. Biochem J. 313:17–29. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Trachootham D, Alexandre J and Huang P:
Targeting cancer cells by ROS-mediated mechanisms: A radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Taguchi K, Fujikawa N, Komatsu M, Ishii T,
Unno M, Akaike T, Motohashi H and Yamamoto M: Keap1 degradation by
autophagy for the maintenance of redox homeostasis. Proce Nat Acad
Sci USA. 109:13561–13566. 2012. View Article : Google Scholar
|
|
36
|
Kerr F, Sofola-Adesakin O, Ivanov DK,
Gatliff J, Gomez Perez-Nievas B, Bertrand HC, Martinez P, Callard
R, Snoeren I, Cocheme HM, et al: Direct Keap1-Nrf2 disruption as a
potential therapeutic target for Alzheimer's disease. PLoS Genet.
13:e10065932017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rodriguez A, Duran A, Selloum M, Champy
MF, Diez-Guerra FJ, Flores JM, Serrano M, Auwerx J, Diaz-Meco MT
and Moscat J: Mature-onset obesity and insulin resistance in mice
deficient in the signaling adapter p62. Cell Metabol. 3:211–222.
2006. View Article : Google Scholar
|
|
38
|
Huang J, Duran A, Reina-Campos M, Valencia
T, Castilla EA, Muller TD, Tschop MH, Moscat J and Diaz-Meco MT:
Adipocyte p62/SQSTM1 suppresses tumorigenesis through opposite
regulations of metabolism in adipose tissue and tumor. Can Cell.
33:770–784.e776. 2018. View Article : Google Scholar
|
|
39
|
Huo L, Su Y, Xu G, Zhai L and Zhao J:
Sulforaphane protects the male reproductive system of mice from
obesity-induced damage: Involvement of oxidative stress and
autophagy. Intern J Environ Res Public Health. 16(pii): E37592019.
View Article : Google Scholar
|
|
40
|
Ebato C, Uchida T, Arakawa M, Komatsu M,
Ueno T, Komiya K, Azuma K, Hirose T, Tanaka K, Kominami E, et al:
Autophagy is important in islet homeostasis and compensatory
increase of beta cell mass in response to high-fat diet. Cell
Metabol. 8:325–332. 2008. View Article : Google Scholar
|
|
41
|
Yamada M, Iwata M, Warabi E, Oishi H, Lira
VA and Okutsu M: p62/SQSTM1 and Nrf2 are essential for
exercise-mediated enhancement of antioxidant protein expression in
oxidative muscle. FASEB J. 33:8022–8032. 2019. View Article : Google Scholar : PubMed/NCBI
|