Introduction
Hepatocellular carcinoma (HCC) is one of the leading
causes of cancer-related deaths worldwide and more than 600,000
deaths are attributed to HCC every year (1). In the last decade, many novel
molecules targeting the HCC pathway have been discovered,
exhibiting promising results (2).
Although early detection and diagnosis play a critical role in
reducing the incidence and mortality of HCC, the 5-year overall
survival (OS) rate of HCC remains unsatisfactory (3). Previous research indicates that only
10–20% of HCC patients can undergo curative treatment at initial
diagnosis, while the remaining patients are inoperable due to a
poor prognosis (4,5). Therefore, it is urgently necessary to
explore new molecular targets for HCC treatment.
The family of WD-repeat proteins consists of more
than 30 regulatory proteins within the beta-propeller fold.
Originally, the WD-repeat has been identified as a core unit
composed of approximately 40 amino acids, which ends with the
residues, tryptophan and aspartic acid. The WD-repeat proteins
regulate the assembly of multiprotein complexes by presenting a
stable platform for simultaneous and reversible protein-protein
interactions (6). By this means,
WD-repeat proteins play important roles in many biological
processes, including signal transduction, cell cycle control,
apoptosis and chromatin assembly (7). The association between WD-repeat
proteins and tumor phenotypes has been further studied by
researchers (8). WD-repeat proteins
are associated with the cell death pathway since previous studies
have shown that mouse Apg16L acts as a scaffold in mammalian
autophagy (9–11). WD-repeat domain
phosphoinositide-interacting protein 2 (WIPI2) is reported to be
regulated by mTORC1 to control autophagic flux (12).
In our previous study, we found that WIPI2
expression in HCC tissues was markedly increased when compared with
that in adjacent normal cells, and WIPI was found to be associated
with patient survival rate. Then we showed that the proliferation
of cells was inhibited and the apoptosis rate was promoted after
depletion of WIPI2 in HCC cell lines. Furthermore, we explored the
molecular mechanisms underlying the WIPI2-mediated tumor cell
growth using WIPI2-depleted and control cells. The results showed
that the AMPK signaling pathway was significantly altered after
WIPI2 depletion. We also detected the downstream factors of the
AMPK signaling pathway and found that the variation tendency was
consistent with our hypothesis. Taken together, our findings
support that WIPI2 regulates the growth of HCC cells mainly through
the AMPK signaling pathway.
Materials and methods
Extraction and analysis of TCGA
datasets
Data for WIPI2 expression and clinical information
in the The Cancer Genome Atlas (TCGA) (https://www.cancer.gov/tcga) were extracted and
analyzed in the UALCAN platform (http://ualcan.path.uab.edu) (13).
Tissue array and immunohistochemistry
(IHC) assay
HCC tissue arrays were purchased from the National
Engineering Center for Biochips (Shanghai, China). The expression
of WIPI2 in the tissues was evaluated by IHC assay using the WIPI2
antibody (dilution 1:150; cat. no. ab105459; Abcam). The tissue
sections were collected, dewaxed in xylene, and rehydrated in
graded ethanol solutions. All the following steps were carried out
in a moist chamber. Then the tissue sections were blocked with 3%
peroxide-methanol at room temperature for endogenous peroxidase
ablation and incubated with blocking buffer (normal goat serum) at
room temperature. After that, the blocking buffer was discarded and
the sections were washed with PBS for 3 times. The sections were
then incubated with purified primary antibody overnight at 4°C, and
incubated with the corresponding secondary antibody (dilution
1:200; cat. no. KGAA35; Nanjing KeyGen Biotech. Co. Ltd.). Finally,
the sections were stained with DAB and hematoxylin. The staining
was scored according to the staining intensity (no color, 0; faint
yellow, 1; claybank, 2; brown, 3) and the proportion of positive
cells (0, <5; 1, <5 and <25%; 2, <25 and <50%, 3,
<50 and <75%; 4, <75% positive cells), and the final
staining scores were calculated as the score of the staining
intensity multiplied by the score of the proportion of positive
cells [score of 0 indicates negative staining (0+); 1–4 is weak
staining (1+); 5–8 is moderate staining (2+); 9–12 scores indicates
strong staining (3+)].
Cell culture
Huh7 and Hep3B cells were purchased from the
Shanghai Institutes for Biological Sciences (China). The cells were
maintained in DMEM (cat. no. 11965; Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS),
100 U/ml of penicillin and 100 µg/ml of streptomycin (complete
medium) at 37°C in a humidified atmosphere containing 5%
CO2.
Small interfering RNA and
transfection
The siRNAs targeting the WIPI2 sequence (KD,
TACGGAAGATGTGTGCATT) and the non-silencing sequence (NC,
TTCTCCGAACGTGTCACGT) were purchased from Guangzhou RiboBio Co.,
Ltd. Transfection with siRNAs was completed using riboFECT CP
transfection reagent (Guangzhou RiboBio Co., Ltd) according to the
manufacturer's instructions.
MTT assay
Human cell lines (1×105 cells in 0.2
ml/well) were seeded into 96-well plates and transfected with the
siRNA and the non-silencing sequence. The cells were cultured at
37°C for 1–5 days. Then 100 µl (5 mg/ml) of MTT solution was added
into each well, followed by incubation at 37°C for 4 h.
Subsequently, the supernatant was removed, and 150 µl of dimethyl
sulfoxide (DMSO) was added into each well. The plate was oscillated
at room temperature for 30 min, and then the absorbance at a
wavelength of 490 nm was determined by a multifunctional microplate
reader (Tecan). The values were calculated after background
subtraction. All the MTT experiments were repeated at least three
times.
EdU incorporation assay
The cell proliferation was determined by the
incorporation of 5-ethynyl-2′-deoxyuridine (EdU) into newly
synthesized DNA strands, using a Click-iT EdU microplate assay kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. The HCC cell lines were resuspended
and seeded into a 6-well plate and transfected with the siRNA and
negative control, respectively. Two hours before cell collection,
EdU was added to the cells at a final concentration of 10 µM. After
2 h, the incorporated EdU in DNA was coupled with Azide 488 dye,
and subsequently the nuclei were stained with Hoechst 33342. The
cells were observed by fluorescence microscope at ×100
magnification. The experiments were repeated at least three
times.
TUNEL assay
Analysis of cell apoptosis was performed by terminal
deoxynucleotidyl transferase-mediated dUTP nick-end labeling
(TUNEL) assay (Nanjing KeyGen Biotech Co., Ltd.) according to the
manufacturer's instructions. Following transfection for 24 h, the
cells were collected and fixed with 40 g/l paraformaldehyde for 30
min at room temperature. The TdT enzyme solution was prepared and
incubated with cells for 60 min at 37°C. Next, the cells were
washed with PBS and incubated with streptavidin-TRITC for 30 min at
37°C. The cell nuclei were then stained with DAPI (dilution
1:1,000) at room temperature for 10 min. Finally, the cells were
mounted in anti-fade mounting medium and were observed with a
fluorescence microscope at ×100 magnification. The experiments were
repeated at least three times.
Quantitative real-time PCR
(RT-qPCR)
Total RNA was extracted from HCC cell lines using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
following the manufacturer's instructions. RT-qPCR was performed as
previously reported (14,15). Briefly, 1 µg of total RNA was
reversely transcribed into cDNA using random primers and
Primescript reverse transcriptase (Takara). RT-qPCR for the
indicated genes was carried out using the SYBR Green qPCR kit
(Takara Dalian) on a fluorescent temperature cycler (Applied
Biosystems ABI™ Vii7 Real Time PCR System; Thermo Fisher
Scientific, Inc.). The following primers were used to detect the
expression of WIPI2 (forward, 5′-CCATCGTCAGCCTTAAAGCAC-3′ and
reverse, 5′-TCCAGGCATACTATCAGCCTC-3′) and GAPDH (forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′). Briefly, after an initial
denaturation step at 95°C for 5 min, amplifications were carried
out with 45 cycles at a melting temperature of 95°C for 15 sec and
an annealing temperature of 60°C for 1 min. GAPDH was
selected as an endogenous control, and the relative gene expression
was determined by the comparative Ct method. The experiment was
repeated at least three times.
Flow cytometry (FCM)
Cells transfected with the siRNA and control were
harvested, and washed twice with cold PBS. Then the cells were
stained with Annexin V-FITC and PI (eBioscience Inc.) and incubated
for 15 min at room temperature, and then determined using a BD
FACSCalibur Flow Cytometer (BD Biosciences) and analyzed using
FlowJo software (v10.0; TreeStar). The experiment was repeated at
least three times.
Western blot analysis
Cells transfected with siRNA were harvested and
total proteins were extracted from tumor cell lines using RIPA
buffer containing fresh protease and phosphatase inhibitors. The
protein concentration was determined using the BCA assay (Pierce;
Thermo Fisher Scientific, Inc.). Briefly, equal amounts of proteins
(50 µg) were subjected to 10% SDS-PAGE and transferred onto PVDF
membranes. The membranes were blocked with 3% BSA in 10 mM Tris-HCl
(pH 7.4) containing 0.05% Tween-20 and incubated with a primary
antibody at 4°C for 12 h. After washing with Tris-HCl buffer for
three times, the membranes were incubated with a corresponding
peroxidase-conjugated secondary antibody (Abcam). Immunoreactive
bands were visualized using Super-Signal West Pico Chemiluminescent
Substrate (Pierce; Thermo Fisher Scientific, Inc.). The
densitometry of the protein bands was quantified by ImageJ software
(1.8.0; National Institutes of Health). The experiment was repeated
at least three times. The details of the primary antibodies used in
the experiments are documented in Table
I.
 | Table I.Primary and secondary antibodies used
in the WB analysis. |
Table I.
Primary and secondary antibodies used
in the WB analysis.
| Antibody | Dilution | Catalog no. | Supplier |
|---|
| WIPI2 | 1:1,000 | ab105459 | Abcam |
| GAPDH | 1:1,000 | 5174 | Cell Signaling
Technology |
| β-actin | 1:1,000 | 3700 | Cell Signaling
Technology |
| AMPK | 1:1,000 | 5831 | Cell Signaling
Technology |
| p-AMPK | 1:1,000 | 50081 | Cell Signaling
Technology |
| AKT | 1:1,000 | 4691 | Cell Signaling
Technology |
| p-AKT | 1:1,000 | 5012 | Cell Signaling
Technology |
| Caspase 3 | 1:1,000 | 9662 | Abcam |
| Bcl2 | 1:1,000 | ab32124 | Abcam |
| Bax | 1:1,000 | ab32503 | Abcam |
| Cyclin D1 | 1:8,000 | ab134175 | Abcam |
| Goat anti-mouse IgG
H&L (HRP) | 1:10,000 | ab205719 | Abcam |
| Goat anti-rabbit
IgG H&L (HRP) | 1:10,000 | ab205718 | Abcam |
Microarray and analysis
Total RNA was isolated from Hep3B cells transfected
with si-WIPI2 (n=3) and Hep3B cells transfected with si-NC (n=3).
RNA samples were analyzed by microarray expression profiling using
the Affymetrix Human GeneChip PrimeView (Affymetrix) according to
the manufacturer's instructions. Briefly, cDNA target preparation
and in vitro transcription were conducted using the GeneChip
3′ IVT PLUS Kit (Affymetrix). Arrays were washed, stained and
processed using the GeneChip Hybridization Wash and Stain Kit
(Affymetrix), after which they were imaged using the Affymetrix
GeneChip Scanner 3000 (Affymetrix) for subsequent generation of raw
data. Genes significantly differentially expressed between the
Hep3B/KD and Hep3B/NC cells were selected based on a threshold
setting of fold change >1.3 and P<0.05. Functional pathway
analysis was conducted using Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway enrichment analysis (16) and Gene Ontology (GO) analysis
(17) according to the
manufacturer's instructions.
Statistical analysis
Data are presented as the mean ± standard deviation
(SD) of three independent experiments. All statistical analyses
were performed using SPSS 18.0 software (SPSS Inc.). P<0.05 was
considered statistically significant; P<0.01 was considered
statistically very significant. Differences among categorical
variables were analyzed using one-way ANOVA/SNK test or
independent-sample Student's t-test. The immunoreactive scores for
WIPI2 for tissue array were analyzed using non-parametric
Mann-Whitney U, Kruskal-Wallis H and Wilcoxon tests.
Results
Expression of WIPI2 is upregulated in
HCC tumor tissues and WIPI2 predicts a poor prognosis
In all 24 types of cancers included in the UACLAN
platform, WIPI2 expression was higher in tumor tissues compared
with normal tissues (Fig. 1A and
B). The survival rate showed that high WIPI2 expression
predicted a poor patient prognosis (Fig. 1C).
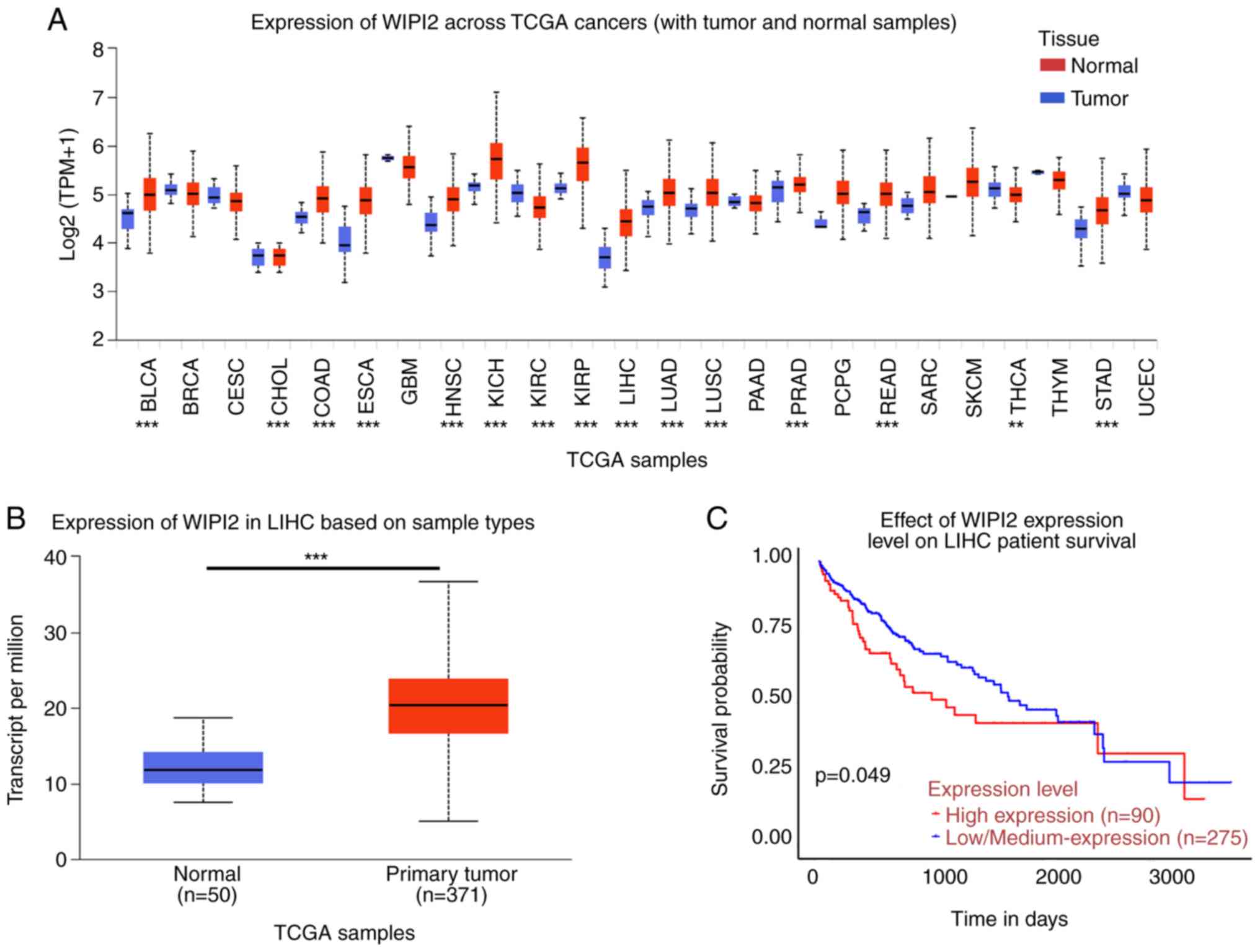 | Figure 1.Expression analysis of WIPI2 based on
the UALCAN platform. (A) Pan-cancer analysis of WIPI2 expression
across cancers from the UALCAN platform. BLCA, bladder urothelial
carcinoma; BRCA, breast invasive carcinoma; CESC, cervical squamous
cell carcinoma and endocervical adenocarcinoma; CHOL,
cholangiocarcinoma; COAD, colon adenocarcinoma; ESCA, esophageal
carcinoma; GBM, glioblastoma multiforme; HNSC, head and neck
squamous cell carcinoma; KICH, Kidney chromophobe; KIRC, kidney
renal clear cell carcinoma; KIRP, kidney renal papillary cell
carcinoma; LIHC, liver hepatocellular carcinoma; LUAD, lung
adenocarcinoma; LUSC, lung squamous cell carcinoma; PAAD,
pancreatic adenocarcinoma; PRAD, prostate adenocarcinoma; PCPG,
pheochromocytoma and paraganglioma; READ, rectum adenocarcinoma;
SARC, sarcoma; SKCM, skin cutaneous melanoma; THCA, thyroid
carcinoma; THYM, thymoma; STAD, stomach adenocarcinoma; UCEC,
uterine corpus endometrial carcinoma. **P<0.01, ***P<0.001.
(B) The WIPI2 level was higher in tumor tissues compared with that
in normal tissues. ***P<0.001. (C) Survival probability revealed
that high WIPI2 expression predicted poor prognosis, P<0.05.
WIPI2, WD repeat domain phosphoinositide-interacting protein 2. |
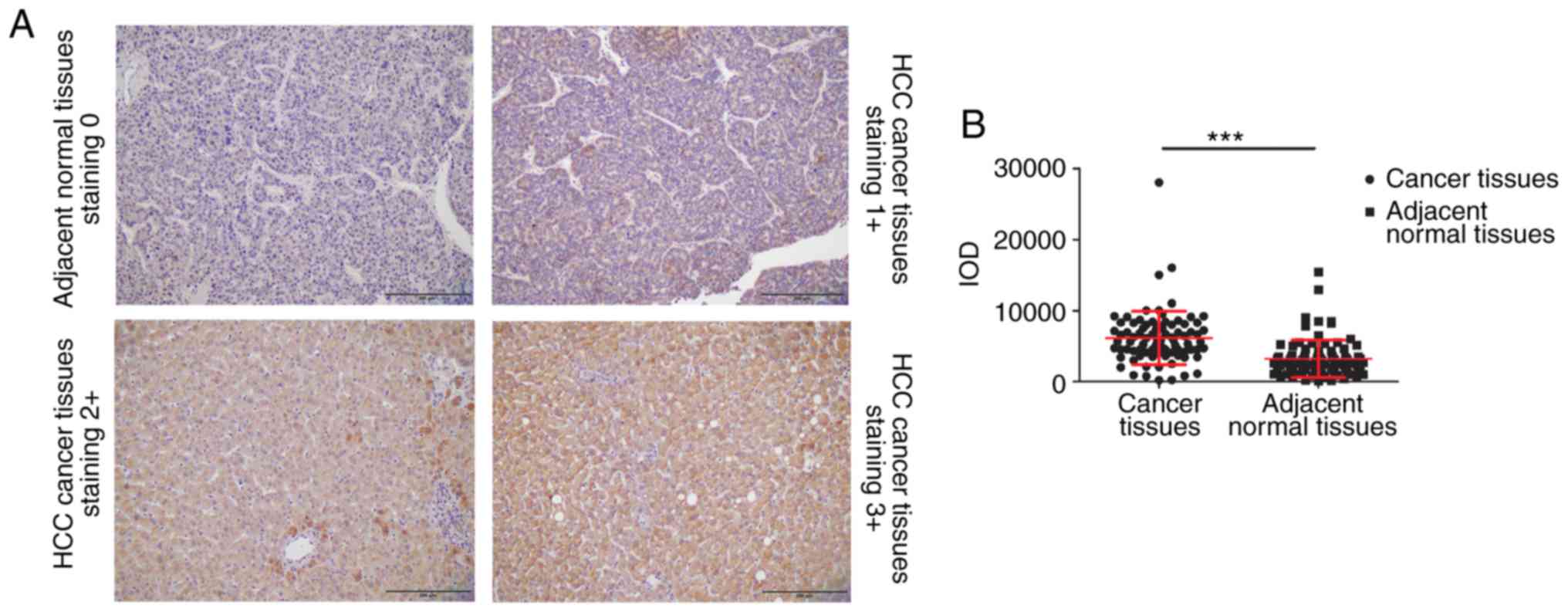
WIPI2 expression was then detected in HCC tissue
array by IHC assay. The array consisted of 86 HCC tissue pairs,
including tumor and corresponding adjacent normal tissues. Among
these samples, 65 tissue samples were obtained from male patients,
while 21 samples were collected from female patients, with a median
age of 67 years (range, 26–88 years). Table II and Fig. 2A show that WIPI2 was highly enriched
in HCC tumor tissue samples, evidenced by the fact that 66 tumor
tissue samples showed a score of ≥2. However, adjacent normal
tissue samples displayed significantly reduced WIPI2 expression
compared with tumor tissues (P<0.001). The results also
indicated that WIPI2 expression was markedly elevated in patients
older than 60 years (Table II). In
addition, WIPI2 expression was not associated with other factors.
However, the statistical analysis indicated that WIPI2 was
significantly higher in tumor tissues than that in the
corresponding adjacent normal tissues (P<0.001) (Fig. 2B).
| Table II.Correlation of WIPI2 expression and
clinicopathological features in the HCC tissue array. |
Table II.
Correlation of WIPI2 expression and
clinicopathological features in the HCC tissue array.
|
|
| WIPI2
expression |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
|
Characteristics | N | 0 | 1 | 2 | 3 | None | Mean rank | Z | P-value |
|---|
| Sex |
|
|
|
|
|
|
|
|
|
|
Male | 65 | 1 | 12 | 23 | 27 | 2 | 39.82 | −1.904 | 0.057 |
|
Female | 21 | 1 | 2 | 3 | 15 | 0 | 50.55 |
|
|
| Age (years) |
|
|
|
|
|
|
|
|
|
|
<60 | 28 | 2 | 12 | 7 | 6 | 1 | 32.02 | −2.989 | 0.003 |
|
≥60 | 58 | 3 | 15 | 1 | 38 | 1 | 47.46 |
|
|
| Tumor size
(cm) |
|
|
|
|
|
|
|
|
|
|
<5 | 43 | 2 | 9 | 3 | 27 | 2 | 42.39 | −0.048 | 0.962 |
| ≥5 | 43 | 1 | 10 | 4 | 28 | 0 | 42.60 |
|
|
|
Differentiation |
|
|
|
|
|
|
|
|
|
| 1 | 7 | 1 | 3 | 1 | 2 | 0 | 31.07 | −0.608 | 0.543 |
| 2 | 24 | 3 | 2 | 7 | 11 | 1 | 25.24 |
|
|
| 3 | 52 | 1 | 21 | 0 | 29 | 1 | 51.74 |
|
|
| 4 | 3 | 0 | 0 | 2 | 1 | 0 | 44.50 |
|
|
| TNM stage |
|
|
|
|
|
|
|
|
|
| I | 17 | 0 | 2 | 1 | 13 | 1 | 14.25 | −0.581 | 0.561 |
| II | 28 | 2 | 13 | 1 | 12 | 0 | 30.32 |
|
|
|
III | 40 | 1 | 4 | 14 | 20 | 1 | 63.00 |
|
|
| IV | 1 | 0 | 0 | 1 | 0 | 0 | 36.00 |
|
|
| Location |
|
|
|
|
|
|
|
|
|
| Tumor
tissue | 86 | 1 | 17 | 10 | 56 | 2 | 109.79 | −6.800 | 0.000 |
|
Adjacent tissue | 86 | 44 | 13 | 6 | 23 | 0 | 61.77 |
|
|
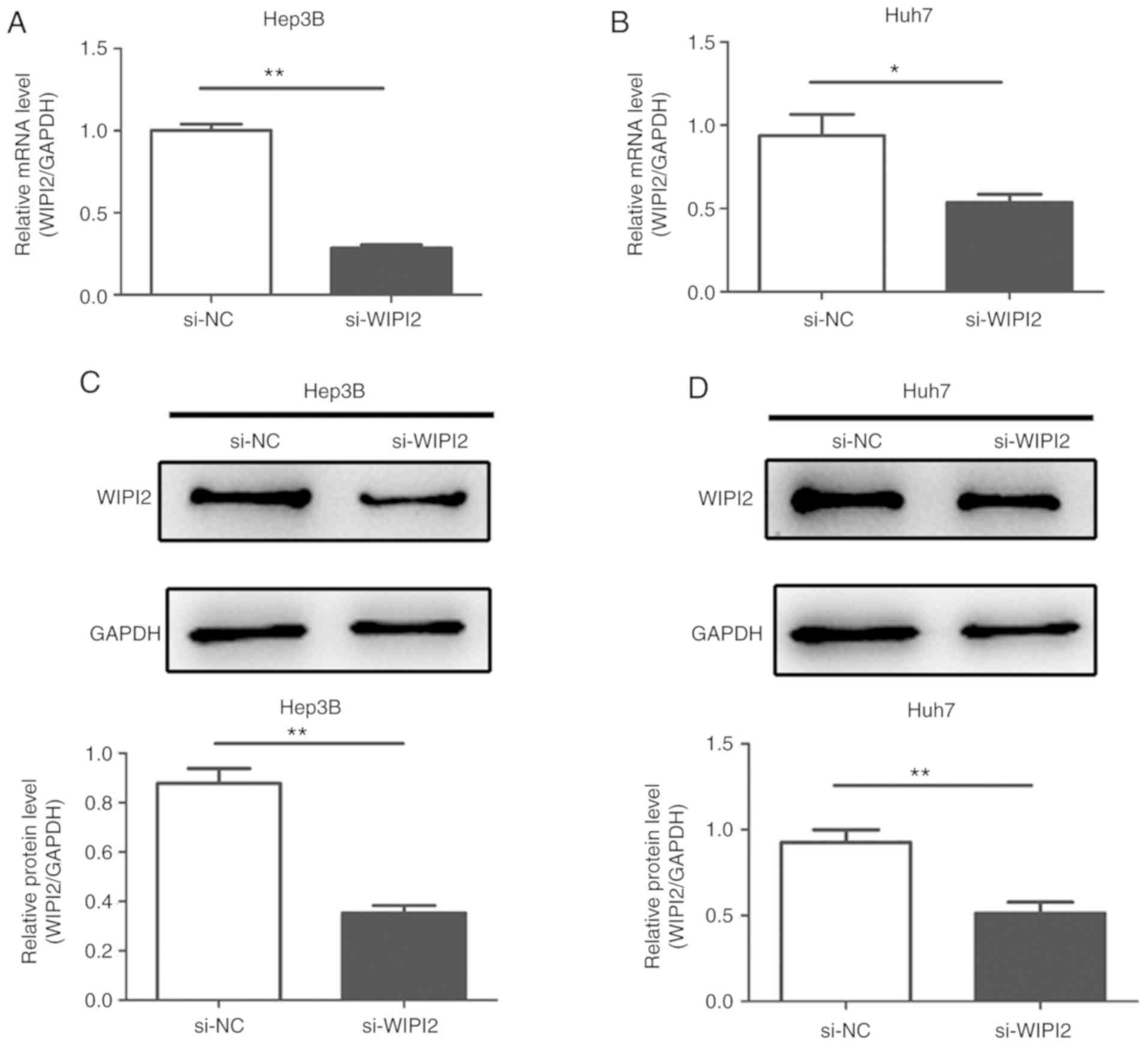
Downregulation of WIPI2 suppresses the
proliferation and increases the cytotoxicity of HCC cells
In the present study, siRNA targeting WIPI2 was
employed to suppress WIPI2 expression in the HCC cell lines in
order to verify its roles in the tumorigenesis of HCC in
vitro. After transfection with siRNA, the expression of WIPI2
at the mRNA and protein levels were both decreased in the HCC Huh7
and Hep3B cell lines (Fig.
3A-D).
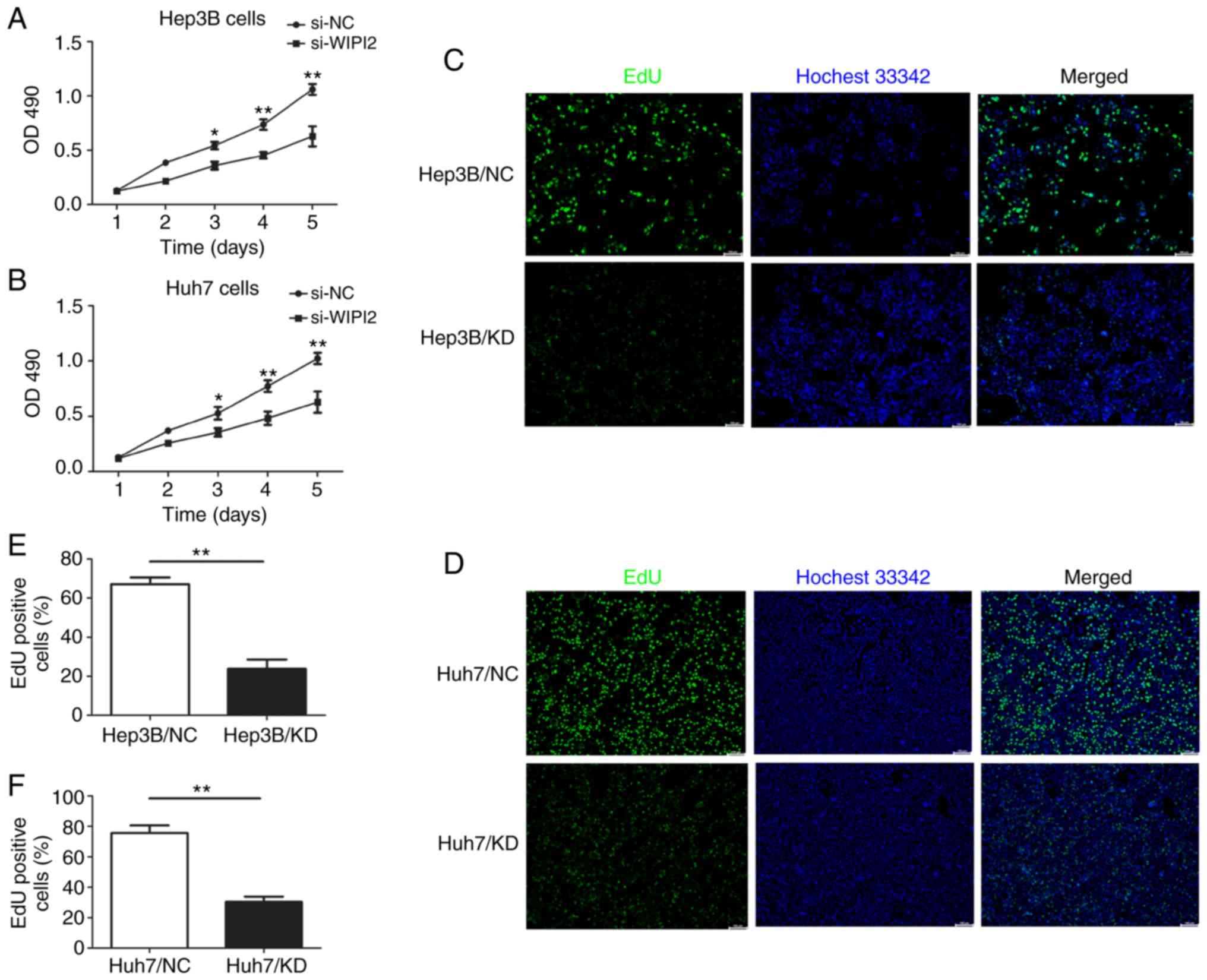
In order to determine the effect of WIPI2 on the
cell proliferation of HCC cells, MTT and EdU assays were used after
WIPI2 downregulation. MTT results showed that the proliferation of
HCC cells was increased following WIPI2 depletion (si-WIPI2)
(Fig. 4A and B). In addition, the
EdU assay indicated that cell proliferation was significantly
decreased after WIPI2 ablation. The number of EdU-positive cells
was significantly reduced upon WIPI2 knockdown (KD) in contrast to
the negative control (NC) cells (Fig.
4C-F).
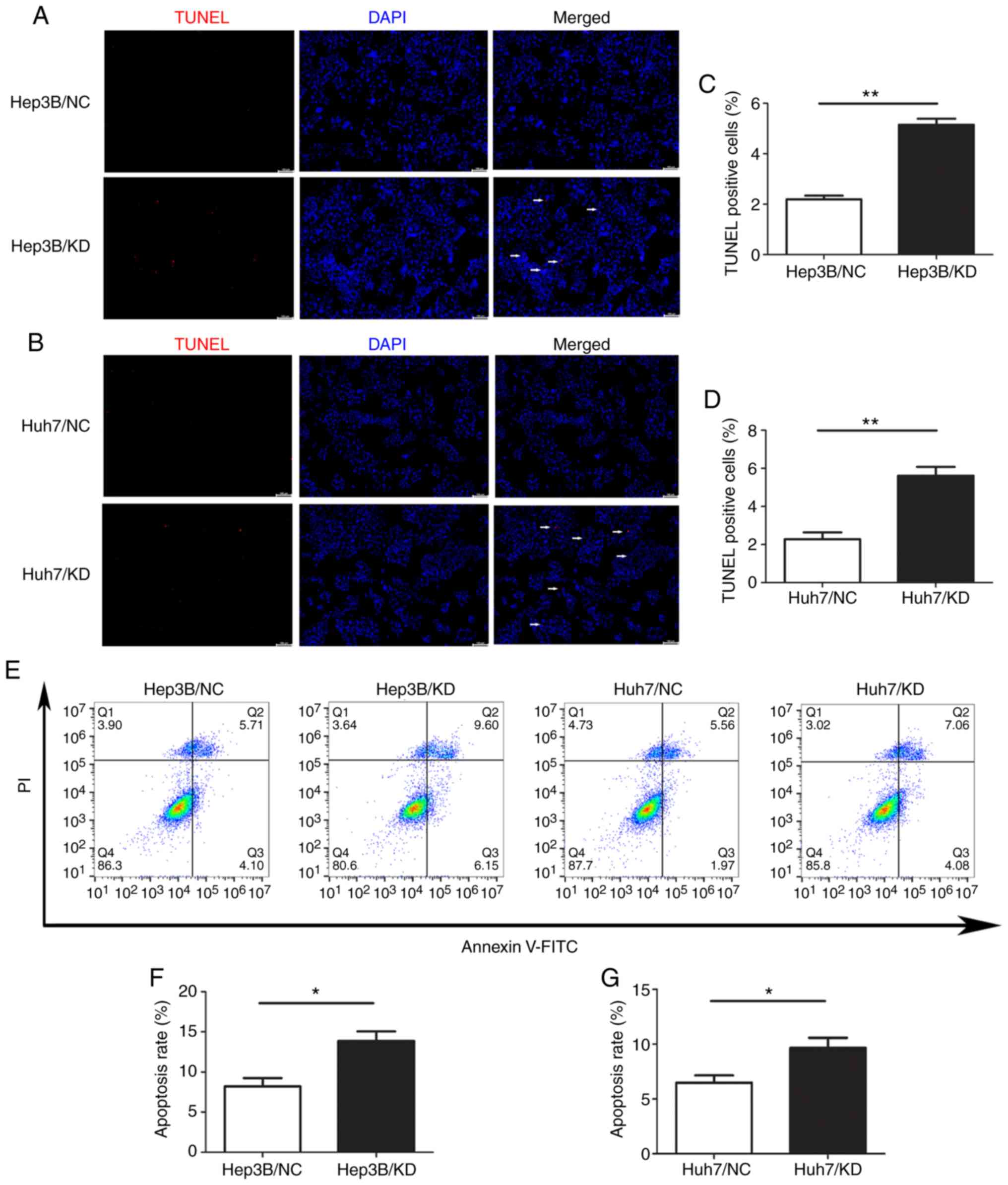
Knockdown of WIPI2 increases the
apoptosis of HCC cells
To explore the role of WIPI2 in regulating the
growth of HCC cells, we assessed the changes in apoptosis after
WIPI2 depletion by TUNEL assay and FCM. The results of TUNEL
staining revealed that the percentage of TUNEL-positive cells was
significantly increased after WIPI2 knockdown (KD) compared with
the negative control (NC) cells (Fig.
5A-D). Consistently, the FCM results indicated that the
apoptotic rate of HCC cells was significantly increased after WIPI2
knockdown both in the Huh7 and Hep3B cell lines (Fig. 5E and F). These results showed that
WIPI2 ablation could promote the apoptosis of HCC cells.
Depletion of WIPI2 inhibits cell
proliferation through the AMPK signaling pathway
To uncover the mechanism underlying the
WIPI2-induced proliferation and apoptosis, we conducted a
microarray analysis to identify differentially expressed genes upon
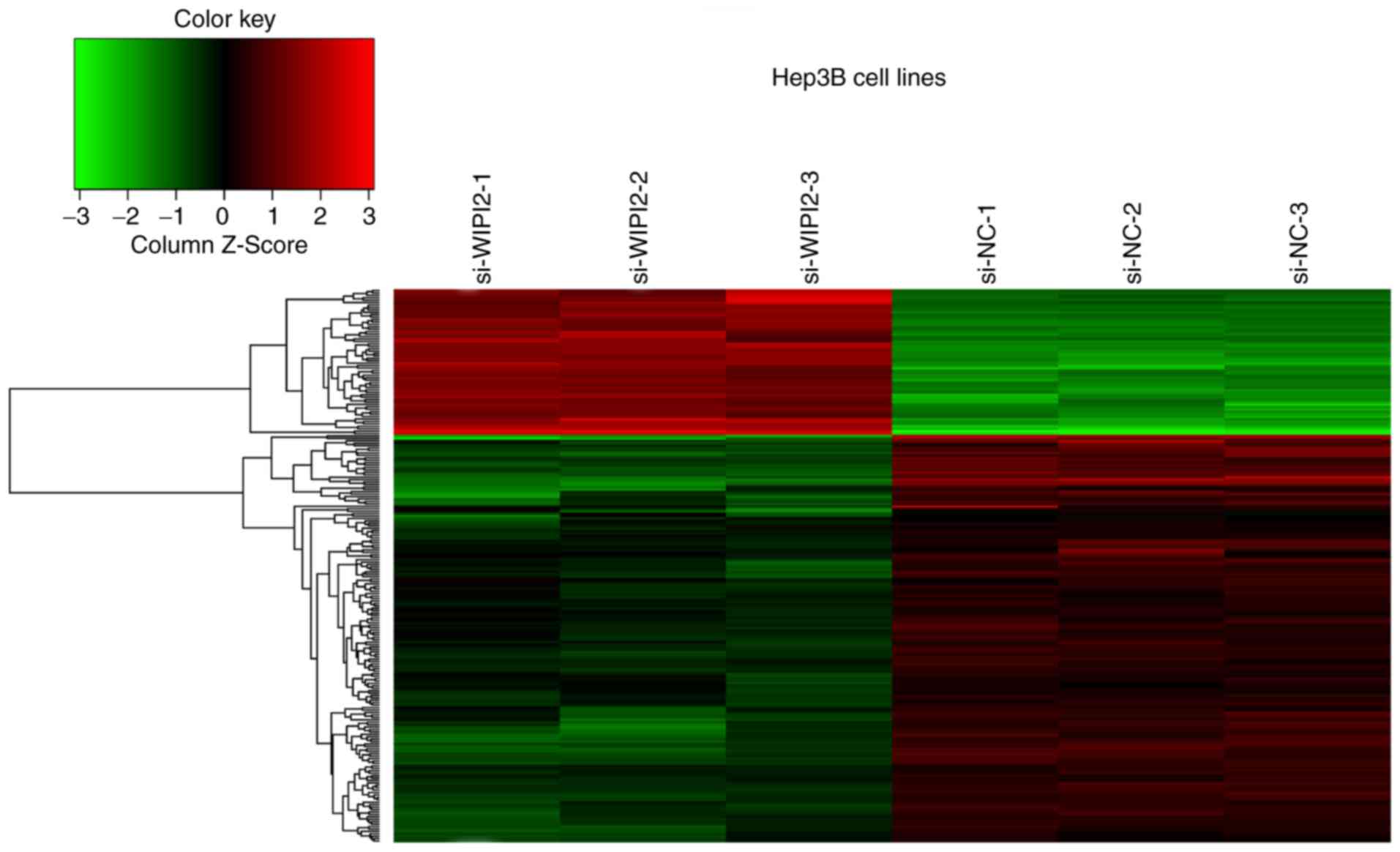
WIPI2 depletion using Hep3B/KD and Hep3B/NC cell lines (Fig. 6). The results showed that a total of
63 genes were upregulated, and 199 genes were downregulated after
WIPI2 depletion (P<0.05 and fold change >1.3), and all the
details of the 216 gene symbols and the fold changes are shown in
Table SI. Supervised analysis was
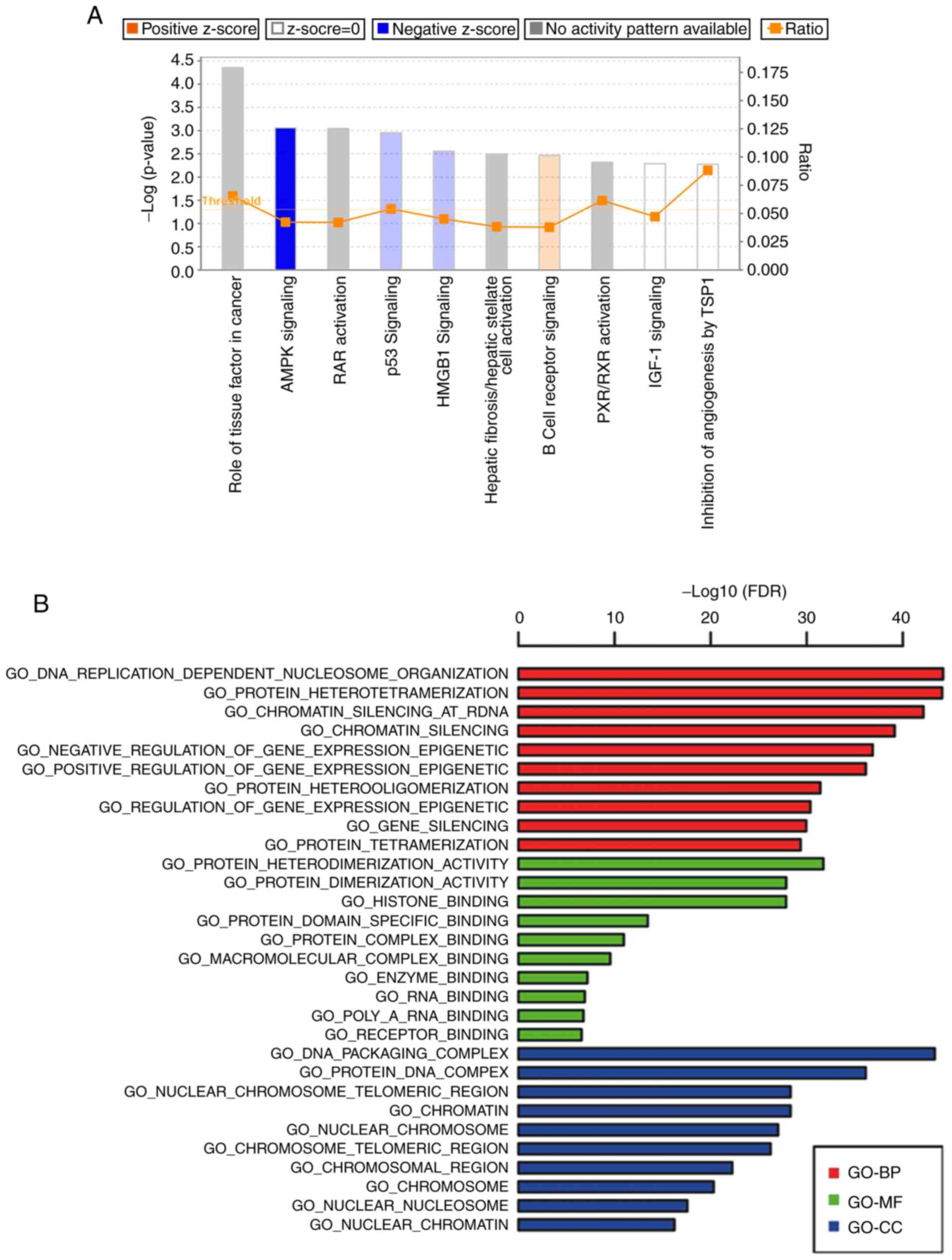
conducted following the KEGG and GO technical route (Fig. 7A and B). We found that the AMPK
signaling pathway was markedly modified after WIPI2 depletion.
In regards to the downstream factors of the AMPK
pathway, AKT is associated with cell proliferation and apoptosis.
Thus, we hypothesized that AKT may be involved in the inhibition of
HCC cells. Further verification was conducted by western blot
analysis. Phosphorylation of AKT (p-AKT) was significantly
decreased, while total AKT remained unchanged after WIPI depletion
(Fig. 8A and C) in the Hep3B cells.
The results also revealed that the ratios of p-AMPK/AMPK and
p-AKT/AKT were reduced after WIPI2 knockdown (Fig. 8E). Cyclin D1 is associated with cell
proliferation; thus, we detected the expression of cyclin D1 and
found that it was significantly decreased after WIPI2 depletion
(Fig 8A and C) in the Hep3B cells.
Since TUNEL assay indicated that the apoptosis was increased, we
also determined the levels of Bcl2, Bax, caspase-3 and
cleaved-caspase 3, key biomarkers of apoptosis (Fig. 8B and D). The results indicated that
the Bcl2/Bax ratio was decreased (Fig.
8F) and cleaved-caspase 3 was also evaluated and caspase-3 was
not altered (Fig. 8B and D) after
WIPI2 knockdown. Similar results were observed in the Huh7 cells
(Fig. 8G-L). Taken together, our
data revealed that WIPI2 regulated the growth of HCC cells mainly
through the AMPK pathway.
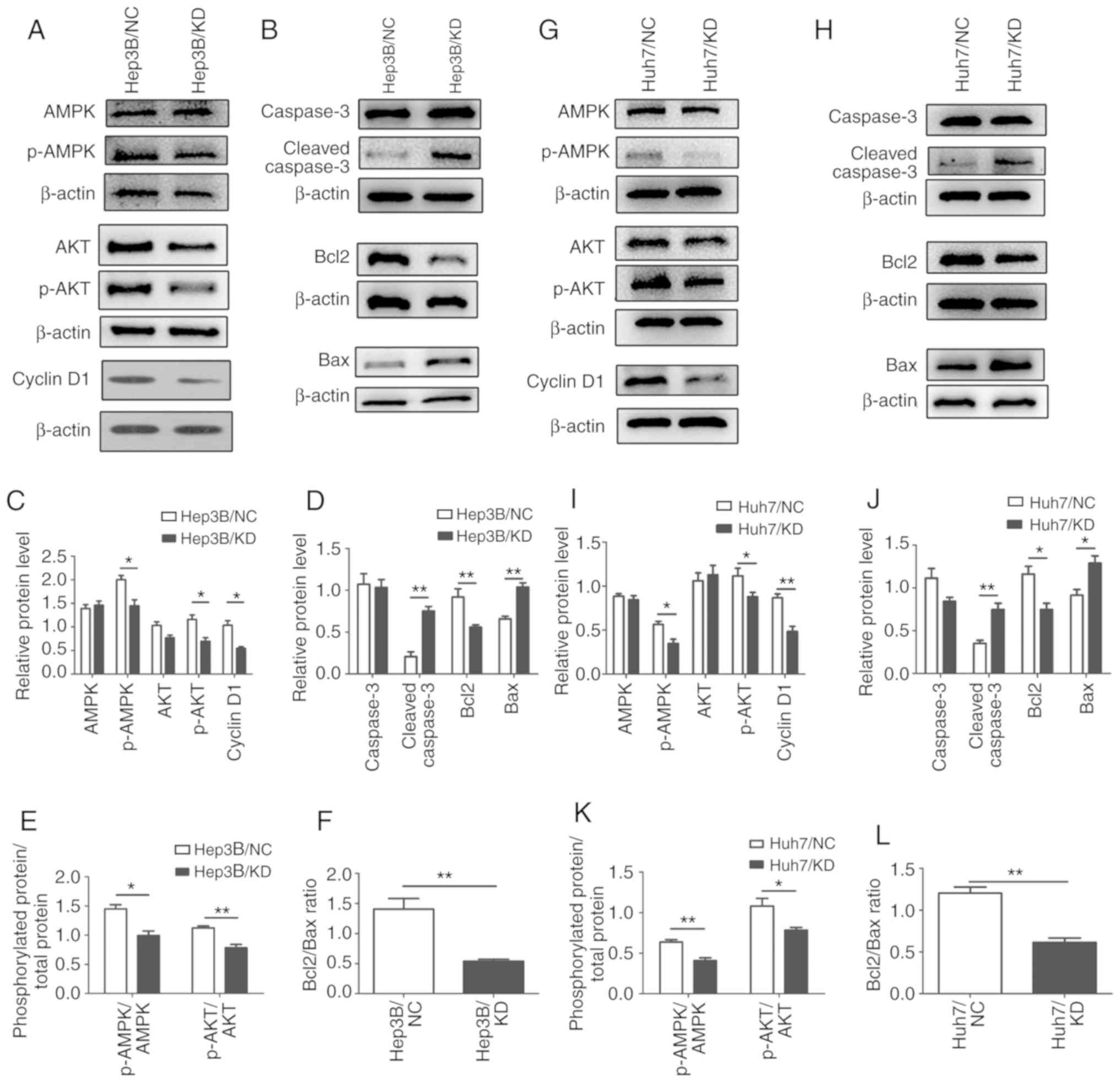 | Figure 8.Mechanisms underlying the
WIPI2-mediated regulation of the proliferation of HCC cells. (A)
Determination of various downstream factors associated with cell
proliferation of the AMPK pathway by western blot analysis in the
Hep3B cells in the WIPI2-knockdown (KD) or negative control (NC)
groups. (B) Detection of various apoptotic biomarkers by western
blot analysis in the Hep3B cells in the two groups. (C) Statistical
analysis of proteins in the Hep3B cells. *P<0.05. (D)
Statistical analysis of apoptosis biomarkers in the Hep3B cells.
**P<0.01. (E) Phosphorylated protein/total protein ratio
analysis after WIPI2 knockdown in the Hep3B cells. *P<0.05,
**P<0.01. (F) Bcl2/Bax ratio analysis after WIPI2 knockdown in
the Hep3B cells. **P<0.01. (G) Determination of various
downstream factors associated with cell proliferation of the AMPK
pathway by western blot analysis in the Huh7 cells in the
WIPI2-knockdown (KD) or negative control (NC) groups. (H) Detection
of various apoptotic biomarkers by western blot analysis in the
Huh7 cells in the two groups. (I) Statistical analysis of proteins
in the Huh7 cells. *P<0.05, **P<0.01. (J) Statistical
analysis of apoptosis biomarker in the Huh7 cells. *P<0.05,
**P<0.01. (K) Phosphorylated protein/total protein ratio
analysis after WIPI2 knockdown in the Huh7 cells, *P<0.05,
**P<0.01. (L) Bcl2/Bax ratio analysis after WIPI2 knockdown in
the Huh7 cells. **P<0.01. WIPI2, WD repeat domain
phosphoinositide-interacting protein 2; AMPK, adenosine
5′-monophosphate (AMP)-activated protein kinase; AKT, protein
kinase B; Bcl2, B-cell lymphoma 2; Bax, Bcl-2-associated X
protein. |
Discussion
In the present study, we first demonstrated the
association between WD repeat domain phosphoinositide-interacting
protein 2 (WIPI2) expression and prognosis of HCC patients. We
found that WIPI2 expression was increased in human HCC tissues when
compared with that in adjacent normal tissues. In vitro
experiments showed that knockdown of WIPI2 significantly inhibited
the proliferation and promoted the apoptosis of HCC cell lines.
In order to reveal how WIPI2 regulated the
pathogenesis of HCC, we performed RNA microarray and western blot
analysis. The bioinformatic analysis indicated that the AMPK
signaling pathway is significantly modified after WIPI2 depletion.
As a heterotrimer, AMPK consists of a catalytic subunit (α) and two
regulatory subunits (β and γ) (18). Compound C
[6-(4-(2-piperidin-1-ylethoxy)phenyl)-3-pyridin-4-ylpyrazolo(1,5-a)pyrimidine]
is a selective AMPK inhibitor (19). Moreover, inhibition of AMPK induced
by compound C can lead to cell cycle arrest and apoptosis (19). Glioma cell growth was found to be
inhibited by treatment of compound C, and the apoptotic effect was
also observed after AMPK knockdown (20). Further research revealed that human
glioma treated with inhibitor of AMPK could be suppressed by
multiple mechanisms containing inhibition of AKT/mTOR, G2-M cell
cycle arrest, and induction of apoptosis mediated with caspase 3
and Bcl2 (20). AMPK downregulation
was able to reduce the proliferation of many solid cancer cell
lines such as HeLa, PC12 and prostate cancer cells (21,22).
We hypothesize that the AMPK/AKT axis is important for cell
proliferation and apoptosis. In the present study, we further
detected the levels of AMPK, p-AMPK, AKT and p-AKT and found that
the AMPK pathway was significantly inhibited after WIPI2 ablation.
And these results were consistent with the above microarray
assay.
The AKT pathway plays a vital role in many
fundamental cellular progresses including proliferation, survival,
apoptosis and metabolism (23,24).
AKT is activated by AMPK phosphorylation (25,26).
After activation, AKT translocates to the cytoplasm or nucleus to
phosphorylate its substrates. Cyclin D1 is a key molecule that is
involved in G1-S phase transition, and its expression is associated
with cell proliferation (27).
Cyclin D1 is an important downstream factor of the AKT pathway
(28), thus we detected cyclin D1
expression. Western blot analysis indicated that cyclin D1 was
significantly decreased accompanied by a decrease in p-AMPK and
p-AKT after WIPI2 knockdown.
Apoptosis is a process of programmed cell death
mainly through the caspase cascade and the Bcl2 gene family. The
caspases consist of ‘initiator’ caspases such as caspase-6,
capase-8, caspase-9 and ‘effector’ caspases including caspase-2,
caspase-3 and caspase 7 (29). When
apoptosis starts, caspase-3 functions as an effector. The apoptosis
induced by the Bcl2 gene family is determined by the ratio of
Bcl2/Bax. A higher ratio of Bcl2/Bax reduces cell apoptosis
(30). In the present study, flow
cytometry and TUNEL assay showed that knockdown of WIPI2 induced
apoptosis, and AMPK inhibition also induced apoptosis (20). We further determined the level of
Bcl2, Bax and cleaved-caspase-3, and found that apoptosis was
increased after WIPI2 downregulation. Taken together, the study
indicated that WIPI2 regulates the growth of HCC cells, at least
partly, through the AMPK pathway.
Collectively, our present study identified that
WIPI2 expression is increased in HCC tissues when compared with
that in adjacent normal tissues and high WIPI2 expression predicts
a poor patient prognosis. Moreover, WIPI2 modulates the growth of
HCC cells mainly through the AMPK/AKT/cyclin D1 axis and induces
apoptosis via caspase-3 and Bcl2. Therefore, WIPI2 may be employed
as a potential therapeutic target for HCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (nos. 31300103 and 81670566), and the
funding body aided us in the design of the study, collection and
analysis of the data. This study was also supported by the Research
of Institute of Hospital Management, Nanjing University
(NDYG2017016), and the funding body aided in the interpretation of
the data and writing of the manuscript.
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contribution
XS and XY conceived and designed this study. CL, FL,
MC, XL, GF and XY performed the experiments. XY and XS collected
and analyzed the data. XS drafted the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The human samples used in the present study were
purchased from Shanghai Outdo Biotech (Shanghai, China). The
company is registered in the National Human Genetic Resources
Sharing Service Platform. The web site is: http://www.egene.org.cn/cms/g-index.jhtml, and the
registration number is 2005DKA21300. The samples of this array were
obtained from Taizhou Hospital of Zhejiang Province, and the use of
this array was approved by the Ethics Committee of Taizhou Hospital
of Zhejiang Province, China. Written informed consent was obtained
from the patients.
Patient consent for publication
Consent was obtained from all individual
participants included in the study.
Competing interests
The authors state that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
WIPI2
|
WD repeat domain
phosphoinositide-interacting protein 2
|
|
Huh7/KD cells
|
Huh7 cells containing WIPI2 siRNA
|
|
Huh7/NC cells
|
Huh7 cells containing non-silencing
sequence
|
|
Hep3B/KD cells
|
Hep3B cells containing WIPI2 siRNA
|
|
Hep3B/NC cells
|
Hep3B cells containing non-silencing
sequence
|
|
KD
|
knockdown
|
|
NC
|
negative control
|
|
AMPK
|
adenosine 5′-monophosphate
(AMP)-activated protein kinase
|
References
|
1
|
Baradaran Noveiry B, Hirbod-Mobarakeh A,
Khalili N, Hourshad N, Greten TF, Abou-Alfa GK and Rezaei N:
Specific immunotherapy in hepatocellular cancer: A systematic
review. J Gastroenterol Hepatol. 32:339–351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ye S, Zhao XY, Hu XG, Li T, Xu QR, Yang
HM, Huang DS and Yang L: TP53 and RET may serve as biomarkers of
prognostic evaluation and targeted therapy in hepatocellular
carcinoma. Oncol Rep. 37:2215–2226. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB: Hepatocellular carcinoma. N
Engl J Med. 365:1118–1127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rampone B, Schiavone B and Confuorto G:
Current management of hepatocellular cancer. Curr Oncol Rep.
12:186–192. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Grimmel M, Backhaus C and Proikas-Cezanne
T: WIPI-mediated autophagy and longevity. Cells. 4:202–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu TW, Mochida GH, Tischfield DJ, Sgaier
SK, Flores-Sarnat L, Sergi CM, Topçu M, McDonald MT, Barry BJ,
Felie JM, et al: Mutations in WDR62, encoding a
centrosome-associated protein, cause microcephaly with simplified
gyri and abnormal cortical architecture. Nat Genet. 42:1015–1020.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Casper AL, Baxter K and Van Doren M: No
child left behind encodes a novel chromatin factor required for
germline stem cell maintenance in males but not females.
Development. 138:3357–3366. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dooley HC, Razi M, Polson HE, Girardin SE,
Wilson MI and Tooze SA: WIPI2 links LC3 conjugation with PI3P,
autophagosome formation, and pathogen clearance by recruiting
Atg12-5-16L1. Mol Cell. 55:238–252. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Axe EL, Walker SA, Manifava M, Chandra P,
Roderick HL, Habermann A, Griffiths G and Ktistakis NT:
Autophagosome formation from membrane compartments enriched in
phosphatidylinositol 3-phosphate and dynamically connected to the
endoplasmic reticulum. J Cell Biol. 182:685–701. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Itakura E and Mizushima N:
Characterization of autophagosome formation site by a hierarchical
analysis of mammalian Atg proteins. Autophagy. 6:764–776. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wan W, You Z, Zhou L, Xu Y, Peng C, Zhou
T, Yi C, Shi Y and Liu W: mTORC1-regulated and HUWE1-mediated WIPI2
degradation controls autophagy flux. Mol Cell. 72:303–315.e6. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yuan X, Sun X, Shi X, Jiang C, Yu D, Zhang
W and Ding Y: USP39 regulates the growth of SMMC-7721 cells via
FoxM1. Exp Ther Med. 13:1506–1513. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan X, Sun X, Shi X, Jiang C, Yu D, Zhang
W, Guan W, Zhou J, Wu Y, Qiu Y and Ding Y: USP39 promotes the
growth of human hepatocellular carcinoma in vitro and in vivo.
Oncol Rep. 34:823–832. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kanehisa M, Sato Y, Furumichi M, Morishima
K and Tanabe M: New approach for understanding genome variations in
KEGG. Nucleic Acids Res. 47(D1): D590–D595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
The Gene Ontology Consortium, . The gene
ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47(D1): D330–D338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hardie DG and Alessi DR: LKB1 and AMPK and
the cancer-metabolism link-ten years after. BMC Biol. 11:362013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Akimoto T, Umemura M, Nagasako A, Ohtake
M, Fujita T, Yokoyama U, Eguchi H, Yamamoto T and Ishikawa Y:
Alternating magnetic field enhances cytotoxicity of Compound C.
Cancer Sci. 109:3483–3493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu X, Chhipa RR, Nakano I and Dasgupta B:
The AMPK inhibitor compound C is a potent AMPK-independent
antiglioma agent. Mol Cancer Ther. 13:596–605. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shaw MM, Gurr WK, McCrimmon RJ, Schorderet
DF and Sherwin RS: 5′AMP-activated protein kinase alpha deficiency
enhances stress-induced apoptosis in BHK and PC12 cells. J Cell Mol
Med. 11:286–298. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Duong HQ, Hwang JS, Kim HJ, Seong YS and
Bae I: BML-275, an AMPK inhibitor, induces DNA damage, G2/M arrest
and apoptosis in human pancreatic cancer cells. Int J Oncol.
41:2227–2236. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y, Wang H, Zhang Y, Wang Z, Liu S and
Cui L: Pretreatment of ghrelin protects H9c2 cells against
hypoxia/reoxygenation-induced cell death via PI3K/AKT and AMPK
pathways. Artif Cells Nanomed Biotechnol. 47:2179–2187. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Alam H, Weck J, Maizels E, Park Y, Lee EJ,
Ashcroft M and Hunzicker-Dunn M: Role of the
phosphatidylinositol-3-kinase and extracellular regulated kinase
pathways in the induction of hypoxia-inducible factor (HIF)-1
activity and the HIF-1 target vascular endothelial growth factor in
ovarian granulosa cells in response to follicle-stimulating
hormone. Endocrinology. 150:915–928. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang M, Zhang Z, Wang C, Li K, Li S, Boden
G, Li L and Yang G: Nesfatin-1 action in the brain increases
insulin sensitivity through Akt/AMPK/TORC2 pathway in diet-induced
insulin resistance. Diabetes. 61:1959–1968. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hur W, Lee JH, Kim SW, Kim JH, Bae SH, Kim
M, Hwang D, Kim YS, Park T, Um SJ, et al: Downregulation of
microRNA-451 in non-alcoholic steatohepatitis inhibits fatty
acid-induced proinflammatory cytokine production through the
AMPK/AKT pathway. Int J Biochem Cell Biol. 64:265–276. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei Y, Huang C, Wu H and Huang J: Estrogen
receptor Beta (ERβ) mediated-cyclin D1 degradation via autophagy
plays an anti-proliferation role in colon cells. Int J Biol Sci.
15:942–952. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He YY, Council SE, Feng L and Chignell CF:
UVA-induced cell cycle progression is mediated by a disintegrin and
metalloprotease/epidermal growth factor receptor/AKT/Cyclin D1
pathways in keratinocytes. Cancer Res. 68:3752–3758. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dubey M, Nagarkoti S, Awasthi D, Singh AK,
Chandra T, Kumaravelu J, Barthwal MK and Dikshit M: Nitric
oxide-mediated apoptosis of neutrophils through caspase-8 and
caspase-3-dependent mechanism. Cell Death Dis. 7:e23482016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Maes ME, Schlamp CL and Nickells RW: BAX
to basics: How the BCL2 gene family controls the death of retinal
ganglion cells. Prog Retin Eye Res. 57:1–25. 2017. View Article : Google Scholar : PubMed/NCBI
|