Introduction
Glioblastoma is the most common primary tumor of the
central nervous system in adults. Despite treatment including
maximal surgical resection, radiotherapy and temozolomide
chemotherapy, overall survival (OS) of patients remains at 14.6
months (1,2). Although several clinical trials have
emerged as potential novel treatments, there have been few advances
in treatment options during the last 3 decades.
Recently, drug repurposing has been successful in
various different therapeutic fields, including those involving
aspirin, sildenafil, erythromycin, minoxidil and thalidomide.
Thalidomide was originally used for the treatment of morning
sickness in the 1950s. However, in 1961, it was banned because of
potential teratogenicity such as phocomelia. In 1999, thalidomide
was found to exert an antitumor effect on multiple myeloma
(3). Subsequently, it was again
used and the side effects are now considered acceptable if the
thalidomide is administered to patients with a life-threatening
condition, such as multiple myeloma (4–7).
On the other hand, lenalidomide is a thalidomide
derivative and was developed in order to reduce the side effects
and enhance the antitumor effect of thalidomide. Clinically, the
antitumor effect of lenalidomide was shown to be more effective
than that of thalidomide in hematopoietic malignancies (8–12).
Recently, lenalidomide has been widely used clinically as an
antitumor drug for hematopoietic malignancies including multiple
myeloma and myelodysplastic syndrome (13–15).
Lenalidomide has been reported to display a multiple antitumor
effect including immunological activity, an inhibitory effect on
angiogenesis, a growth inhibitory effect and induction of apoptosis
(16). The main antitumor mechanism
of lenalidomide has been proposed to involve induction of growth
arrest due to an increased expression of p21 (16). It was reported recently that cell
cycle arrest induced by p21 is related to cereblon protein as a
primary target of lenalidomide (17), and cereblon expression is required
for the antitumor activity of lenalidomide (18).
Thalidomide has been previously reported to exert an
antitumor effect on glioma cells. Several clinical trials since
then have revealed an effect of thalidomide on malignant glioma
patients (19–21). However, these trials failed to
demonstrate an extended median survival time. An antitumor effect
of lenalidomide in glioblastoma can also be expected, as
lenalidomide is superior to thalidomide in cases of hematopoietic
malignancies clinically. Furthermore, lenalidomide has a low
molecular weight of 259.26 g/mol and can therefore pass through the
blood-brain barrier. Lenalidomide may display an antitumor effect
by prolonging progression-free survival and overall survival of
patients with glioblastoma, and Phase I trials using lenalidomide
for central nervous system tumors were reported in 2007 and 2011
(22,23). In these studies, lenalidomide was
well tolerated in patients including children with recurrent
central nervous system tumors, although the toxicity due to
lenalidomide mainly involved myelosuppression and thromboembolism.
Preliminary data indicated that the effect of lenalidomide was
favorable. However, its effect on malignant glioma cells has
remained controversial. In the present study, we aimed to elucidate
the antitumor effect of lenalidomide on malignant glioblastoma
cells.
Materials and methods
Materials
Lenalidomide (Celgene Corp.) was employed for the
experiments. Lenalidomide was dissolved in DMSO, which was used as
a control at corresponding concentrations.
Cell lines and culture
Human malignant glioma cell lines A-172 (cell no.
JCRB0228; lot no. 021999), AM-38 (cell no. IFO05492; lot no.
12082003), T98G (cell no. IFO50303; lot no. 1007), U-251MG (cell
no. IFO 50288; lot no. 12132002), and YH-13 (cell no. IFO50493; lot
no. 1164) (purchased from Health Science Research Resources Bank),
and U-138MG (cat. no. HTB-16; lot no. 1104428; purchased from
American Type Culture Collection) were employed in the present
study. These cells were regularly cultured in Dulbecco's modified
Eagle's medium (DMEM) (Nissui Pharmaceutical) supplemented with 5%
fetal bovine serum (FBS) (Thermo Fisher Scientific, Inc.) using
plastic flasks (Corning) in a humidified incubator at 37°C with an
atmosphere containing 5% CO2, as described previously
(24).
Coulter counter assays
The growth inhibition of malignant glioma cells by
lenalidomide was evaluated by counting the numbers of cells using a
Z1 Coulter Counter® (Beckman Coulter). Each well was
seeded at 1×104 cells in 24 plates and cultured for 24
h. The cells were incubated in the medium with 0.1, 1, 10 and 100
µM of lenalidomide, respectively. After 72 h of exposure to the
various concentrations of lenalidomide, the cells were trypsinized
and counted with the cell Coulter counter. The experiments were
repeated 4 times at each concentration.
Furthermore, to evaluate the underlying mechanisms
of the antitumor effect of lenalidomide, we undertook further
experiments employing A-172 and AM-38 human glioma cell lines. The
cells were incubated for 24 h to achieve attachment and exposed to
lenalidomide at a concentration of 10 µM in the new medium.
Cell cycle distribution analysis
A-172 and AM-38 cells were harvested using 0.25%
trypsin-EDTA solution (Invitrogen; Thermo Fisher Scientific, Inc.)
after 0, 8, 24, 48 and 72 h of lenalidomide treatment, fixed in
ice-cold 70% of ethanol, and treated with 0.5% of RNase A (Roche
Diagnostics). The treated cells were stained with 1 µg/ml propidium
iodide (PI) (Miltenyi Biotec) for 30 min. The fluorescence was
measured with a FACS-Calibur flow cytometer (BD Biosciences) at a
wavelength of 610 nm (FL3). The DNA histograms were analyzed
employing FlowJo software (v.9. 9. 6; BioLegend, Inc.).
Determination of apoptotic cells
The apoptosis induced by lenalidomide was evaluated
by flow cytometry. The cells were incubated in the medium with 0.1,
1, 10 and 100 µM of lenalidomide, respectively. Following
lenalidomide treatment, the cells were harvested using 0.25%
trypsin-EDTA solution. After rinsing twice with phosphate-buffered
saline (PBS) (−) (Nissui Pharmaceutical), the fixed cells were
treated with RNase A for 1 h, and propidium iodide (PI) and Annexin
V Alexa Flour 488 conjugate (Thermo Fisher Scientific, Inc.) were
added. Flow cytometry was performed with a FACS-Calibur flow
cytometer (BD Biosciences). The fluorescence was detected with FL1
(510 nm) and FL3 (610 nm). The apoptosis was analyzed employing
FlowJo software (BioLegend, Inc.).
Western blot analysis
Soluble protein lysates of sub-confluent glioma
cells were obtained using radioimmunoprecipitation assay (RIPA)
buffer (Wako Pure Chemical) containing protease inhibitors (Complet
Mini, EDTA-free, Roche Diagnostics) for 20 min on ice. After
centrifugation at 20,400 × g for 1 h at 4°C, the protein content of
the separated supernatants was determined by BCA assay employing a
Pierce BCA protein assay kit (Thermo Scientific, Inc.). The protein
(100 µg proteins) was loaded and separated by 12% polyacrylamide
gel electrophoresis and then transferred onto nitrocellulose
membranes (Bio-Rad Laboratories) for 1 h at 15 V with a Bio-Rad
transblot (Bio-Rad Laboratories). The membrane was treated with
blocking buffer (PBS/0.05% containing 1% skimmed milk) for 1 h at
room temperature, and subsequently treated with primary antibodies
(Table I) in new blocking buffer
for 24 h at 4°C. After washing the membrane with washing buffer
(PBS/0.05% Tween 20), treatment with the secondary antibody
(Table I) conjugated with
horseradish peroxidase was undertaken for 1 h in room air. The
transferred membrane was washed 3 times with washing buffer and
visualized using LAS-4000 (GE Healthcare) after treatment with ECL
Prime Western Blotting detection reagent (dilution 1:1,000) (GE
Healthcare). The proteins (Bax and cleaved PARP in A-172 cells,
caspase-9 and caspase-8 in A-172 cells, and cleaved PARP and
caspase-8 in AM-38 cells) were probed from the same membranes.
 | Table I.Primary and secondary antibodies for
the western blot analysis. |
Table I.
Primary and secondary antibodies for
the western blot analysis.
| Antibody name | Species | Dilution | Supplier |
|---|
| Primary
antibody |
|
|
|
|
Anti-cereblon antibody | Rabbit
polyclonal | 1:1,000 | Proteintech |
|
Anti-p53 antibody | Mouse
monoclonal | 1:500 | Santa Cruz
Biotechnology |
|
Anti-p-p53 antibody | Mouse
monoclonal | 1:500 | Santa Cruz
Biotechnology |
|
Anti-p21 antibody | Mouse
monoclonal | 1:500 | Santa Cruz
Biotechnology |
|
Anti-Bcl-2-associated ×
protein (Bax) antibody | Mouse
monoclonal | 1:500 | Santa Cruz
Biotechnology |
|
Anti-caspase-9 antibody | Mouse
monoclonal | 1:1,000 | Cell Signaling
Technology |
|
Anti-caspase-3 antibody | Mouse
monoclonal | 1:500 | Santa Cruz
Biotechnology |
|
Anti-caspase-8 antibody | Mouse
monoclonal | 1:1,000 | Cell Signaling
Technology |
|
-β-actin antibody | Mouse
monoclonal | 1:2,000 | Wako Pure Chemical
Industries, Ltd. |
|
Anti-cleaved PARP
antibody | Rabbit
polyclonal | 1:500 | Cell Signaling
Technology |
| Secondary
antibodies |
|
|
|
|
Conjugated anti-mouse IgG
peroxidase |
|
| Sigma-Aldrich;
Merck KGaA |
|
Conjugated anti-rabbit IgG
peroxidase |
|
| Cell Signaling
Technology |
Quantification of mRNA by real-time
quantitative reverse transcriptional-PCR (RT-qPCR)
Cells were cultured in 25 cm2
collagen-coated flasks. After treatment with 10 µM lenalidomide,
the total RNA was extracted from the cells by employing a RNeasy
Mini kit (Qiagen, Inc.). RT-qPCR was performed on a Step-One
Real-time PCR system (Applied Biosystems) with SYBR-Green PCR Mix
(Toyobo). The PCR conditions were as follows: 1st stage, 90°C for
30 sec, 61°C for 20 min, and 95°C for 1 min; 2nd stage, 45 cycles
at 95°C for 15 sec, 55°C for 15 sec and 74°C for 45 sec; and 3rd
stage, 95°C for 15 sec, 60°C for 1 min, and 95°C for 15 sec.
Glyceraldehyde-3-phosphatase dehydrogenase (GADPH) mRNA
expression levels were employed as the quantitative internal
control. The expression levels were calculated employing the
following equations by comparing the threshold cycle (Cq): ∆Cq=Cq
of p21 or p53-Cq of GAPDH, ∆∆Cq (target cell line)-∆Cq (reference
cell line), and ratio=2−∆∆Cq (25). Fold changes of the related mRNA
expression were calculated by using the comparative
2−ΔΔCq method to normalize the mRNA level. The sequences
of primers that were utilized for the PCR are shown in Table II.
| Table II.Primer sets. |
Table II.
Primer sets.
| Gene | Primer |
|---|
| GAPDH | Forward;
5′-CAGAACATCATCCCTGCCTCT-3′ |
|
| Reverse;
5′-GCTTGACAAAGTGGTCGTTGAG-3′ |
| p53 | Forward;
5′-GGCCCACTTCACCGTACTAA-3′ |
|
| Reverse;
5′-GTGGTTTCAAGGCCAGATGT-3′ |
| p21 | Forward;
5′-TGGAGACTCTCAGGGTCGAAA-3′ |
|
| Reverse;
5′-GGCGTTTGGAGTGGTAGAAATC-3′ |
| Bax | Forward;
5′-TTTGCTTCAGGGTTTCATCC-3′ |
|
| Reverse;
5′-CAGTTGAAGTTGCCGTCAGA-3′ |
|
Caspase-9 | Forward;
5′-TTCCCAGGTTTTGTTTCCTG-3′ |
|
| Reverse;
5′-CCTTTCACCGAAACAGCATT-3′ |
|
Caspase-3 | Forward;
5′-AGAACTGGACTGTGGCATTGAG-3′ |
|
| Reverse;
5′-GCTTGTCGGCATACTGTTTCAG-3′ |
Statistical analysis
All studies were carried out more than 3 times. The
resultant data are expressed as the means ± SE (standard error) and
were considered significant at P<0.05 or P<0.01. The
statistical significance was evaluated by the Student's t-test and
one-way analysis of variance (ANOVA) with Tukey's post hoc test.
Data analyses were performed employing the statistical software IBM
SPSS statistics v.21.0 (International Business Machines
Corporation).
Results
Coulter counter assay of glioma cells
exposed to lenalidomide
To evaluate the antitumor effect of lenalidomide,
the six malignant glioma cell lines (A-172, AM-38, T98 G, U-138MG,
U-251MG, and YH-13) were incubated in medium with 0.1, 1.0, 10.0
and 100.0 µM of lenalidomide, respectively. Lenalidomide inhibited
the cell counts of all malignant glioma cells in a
concentration-dependent manner (Fig.
1). The cell growth inhibition effect tended to be confirmed at
10 and 100 µM exposure to lenalidomide.
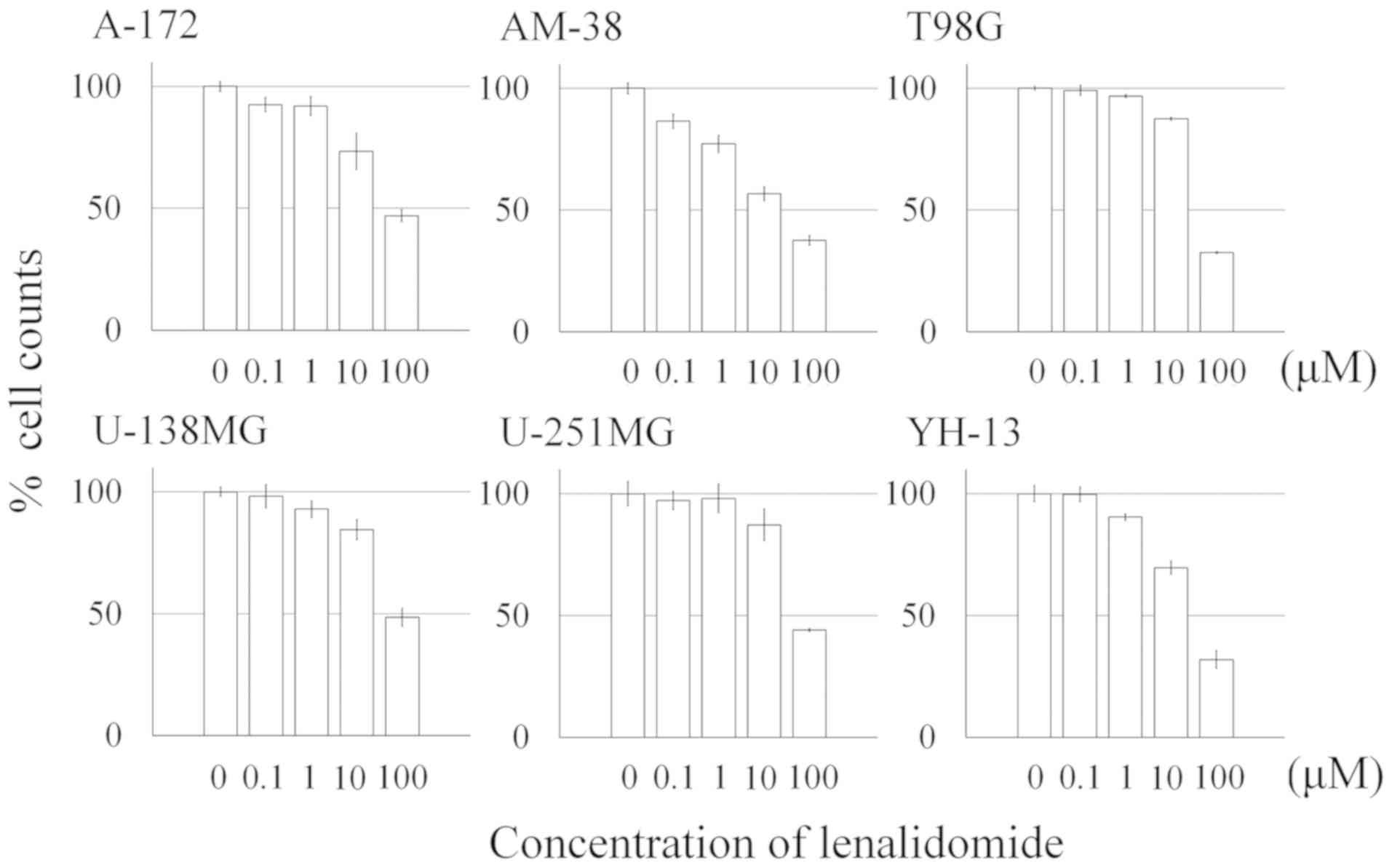 | Figure 1.Percentage of the cell counts of
glioma cells following treatment with lenalidomide. Six malignant
glioma cell lines (A-172, AM-38, T98G, U138MG, U-251MG and YH-13)
were incubated in medium with 0.1, 1.0, 10.0 and 100.0 µM of
lenalidomide, respectively. After 72 h of exposure to the various
concentrations of lenalidomide, the cells were trypsinized and
counted using a Z1 Coulter Counter. The number of all malignant
glioma cells was inhibited in a concentration-dependent manner.
Lenalidomide exerted an antitumor effect in the malignant glioma
cell lines, and in most cell lines, this antitumor effect was
confirmed at a concentration of >10 µM. The results are
expressed as the means ± SE. |
Effects of lenalidomide on the
proliferation of A-172 and AM-38 cells
Hallmarks of cell cycle arrest including p53, p-p53
and p21 were determined in the glioma cells treated with
lenalidomide using western blot analysis (Fig. 2A). The expression levels of p-p53
and p21 protein were increased at 4 h of treatment, while the
expression of p53 protein was unchanged in the A-172 cells. In
AM-38 cells, the expression of p-p53 protein was increased at 8 h
of treatment, while the expression of p53 protein was decreased at
8 h of treatment and increased at 24 h of treatment. The expression
of p21 protein was increased at 24 h after lenalidomide treatment
(Fig. 2A). The RT-qPCR data showed
that the p53 and p21 mRNA expression levels were
significantly increased in the A-172 and AM-38 cells receiving 3 h
of lenalidomide treatment (Fig.
2B). DNA flow cytometric analysis demonstrated a significantly
increased population of cells in the G0/G1
phase at 24 h after treatment with lenalidomide. These findings
indicated that alterations in the cell cycle distribution had
occurred in the A-172 and AM-38 cells (Fig. 3A and B).
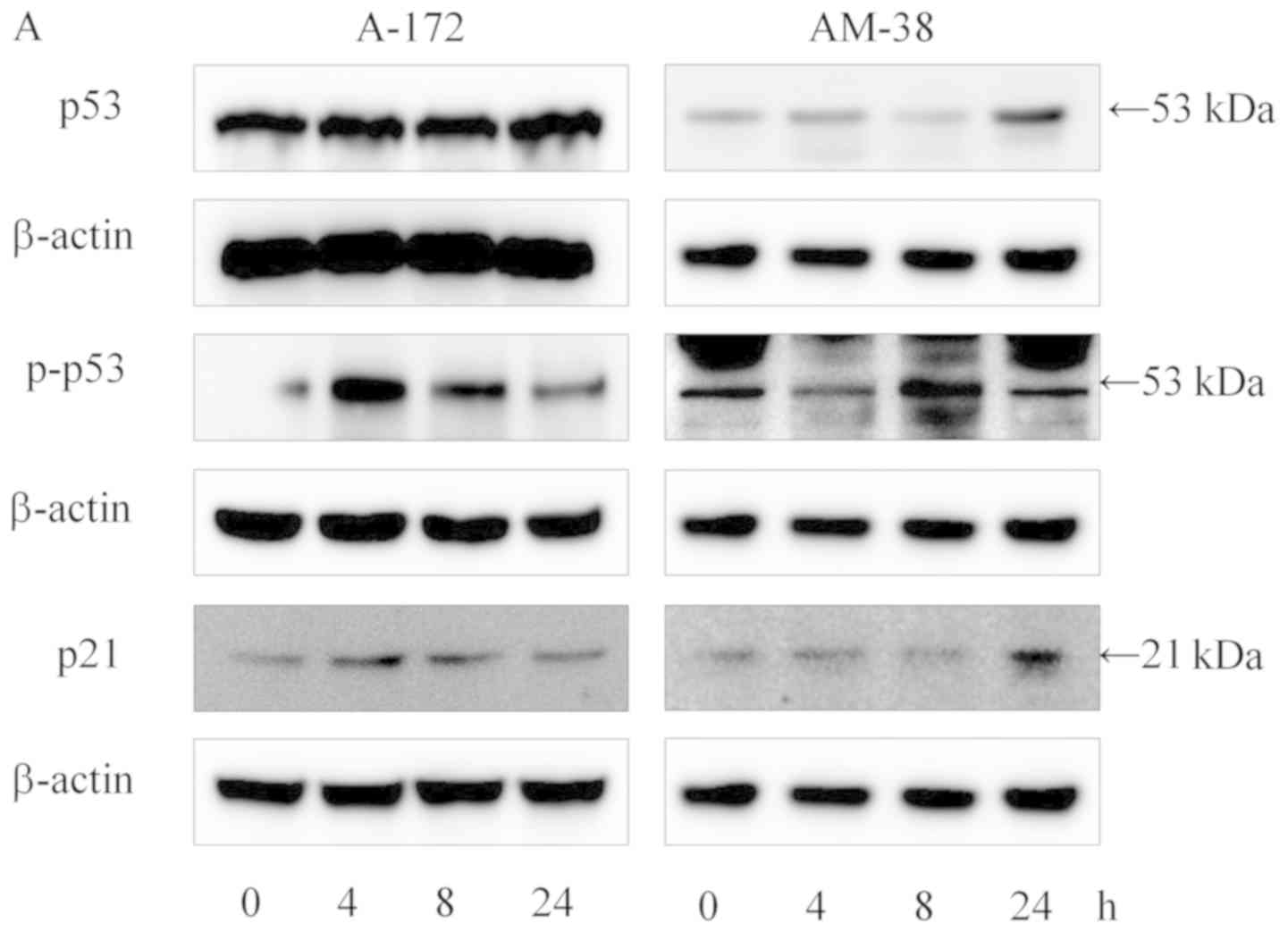 | Figure 2.Effects of lenalidomide on the
proliferation of A-172 and AM-38 glioma cell lines. (A) Proteins
related to cell cycle arrest hallmarks including p53,
phosphorylated (p)-p53 and p21 were investigated by western blot
analysis at 0, 4, 8 and 24 h after treatment with lenalidomide,
respectively. The expression levels of p-p53 and p21 proteins were
increased at 4 or 8 h of treatment, while the expression of p53
protein remained unchanged in A-172 cells. The expression of p53
protein was decreased, and the expression of p-p53 protein was
increased at 8 h of treatment in AM-38 cells, while, the expression
of p21 protein was increased at 24 h after lenalidomide treatment
in in the AM-38 cells. (B) The mRNA expression of p53 and
p21, related with cell cycle arrest, was investigated by
RT-qPCR. The data obtained showed that p53 and p21
mRNA was overexpressed at 3 h after lenalidomide treatment.
GAPDH was employed as the internal control. The relative
expression level of the genes was calculated using
2−ΔΔCq method. The results are expressed as the means ±
SE. **P<0.01 (Student's t-test). |
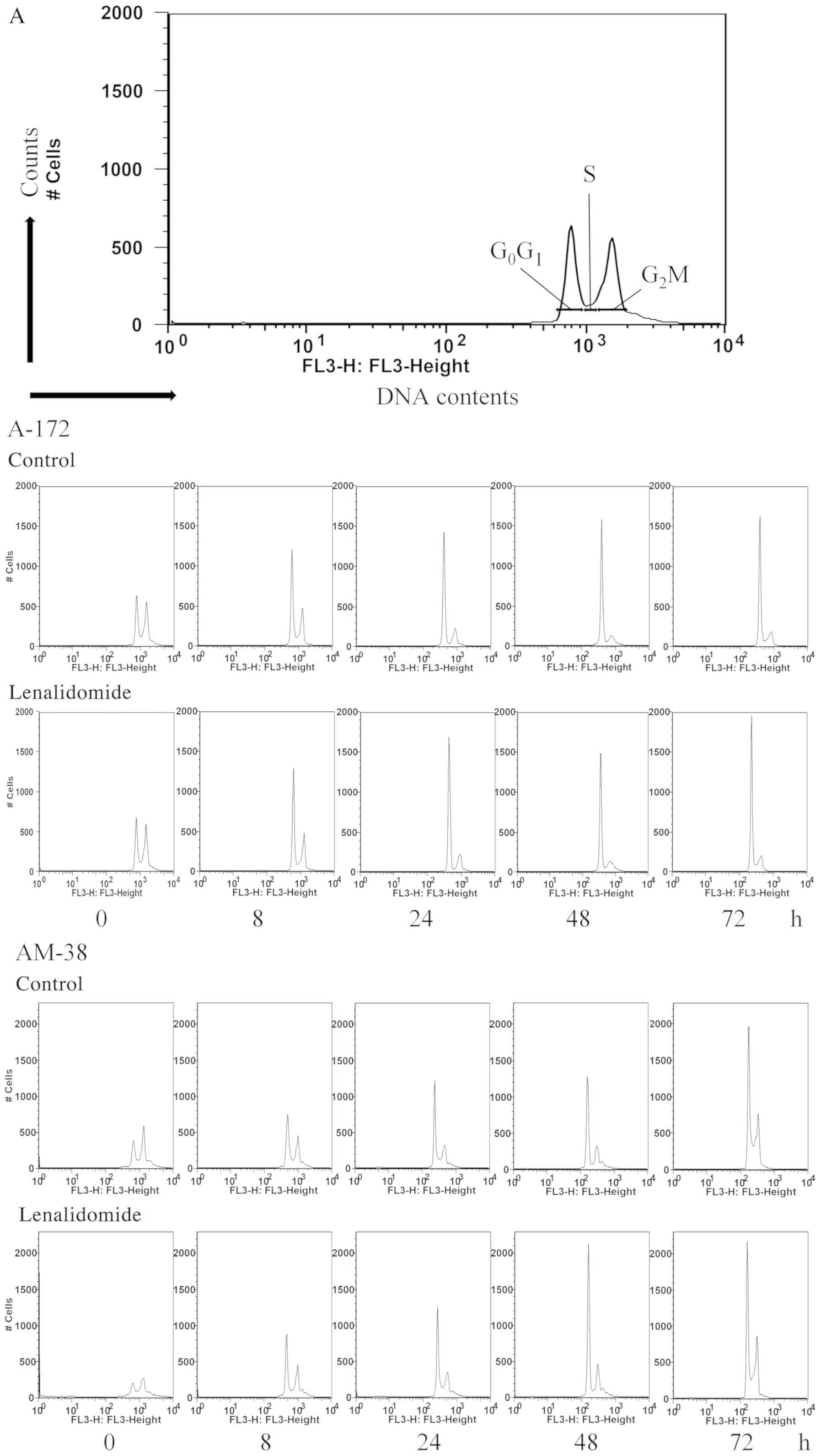 | Figure 3.(A and B) Cell cycle distribution
analysis of A-172 and AM-38 cells treated with 10 µM of
lenalidomide at 0, 8, 24, 48, and 72 h. A-172 and AM-38 were used
for this experiment. These cells were treated with 10 µM of
lenalidomide for 0, 8, 24, 48, and 72 h. A group treated with the
same amount of DMSO as that in which the lenalidomide was dissolved
was used as the control. The histogram data revealed an increase in
population of cells in the G0/G1 phase
following lenalidomide treatment (*P<0.05, **P<0.01). (A and
B) Cell cycle distribution analysis of A-172 and AM-38 cells
treated with 10 µM of lenalidomide at 0, 8, 24, 48, and 72 h. A-172
and AM-38 were used for this experiment. These cells were treated
with 10 µM of lenalidomide for 0, 8, 24, 48, and 72 h. A group
treated with the same amount of DMSO as that in which the
lenalidomide was dissolved was used as the control. The histogram
data revealed an increase in population of cells in the
G0/G1 phase following lenalidomide treatment
(*P<0.05, **P<0.01). |
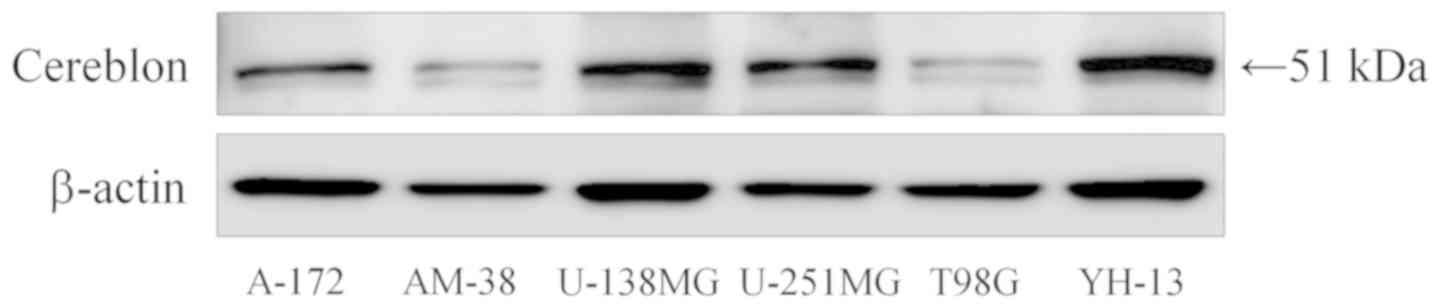
Expression of cereblon
Furthermore, to evaluate the mechanism underlying
the cell cycle arrest induced by lenalidomide, we examined the
expression of cereblon protein which is considered the primary
target of lenalidomide. Cereblon protein was expressed in all cell
lines, but the degree of expression was varied (Fig. 4). There was no obvious relationship
between the expression level of cereblon and inhibition of
malignant glioma due to lenalidomide.
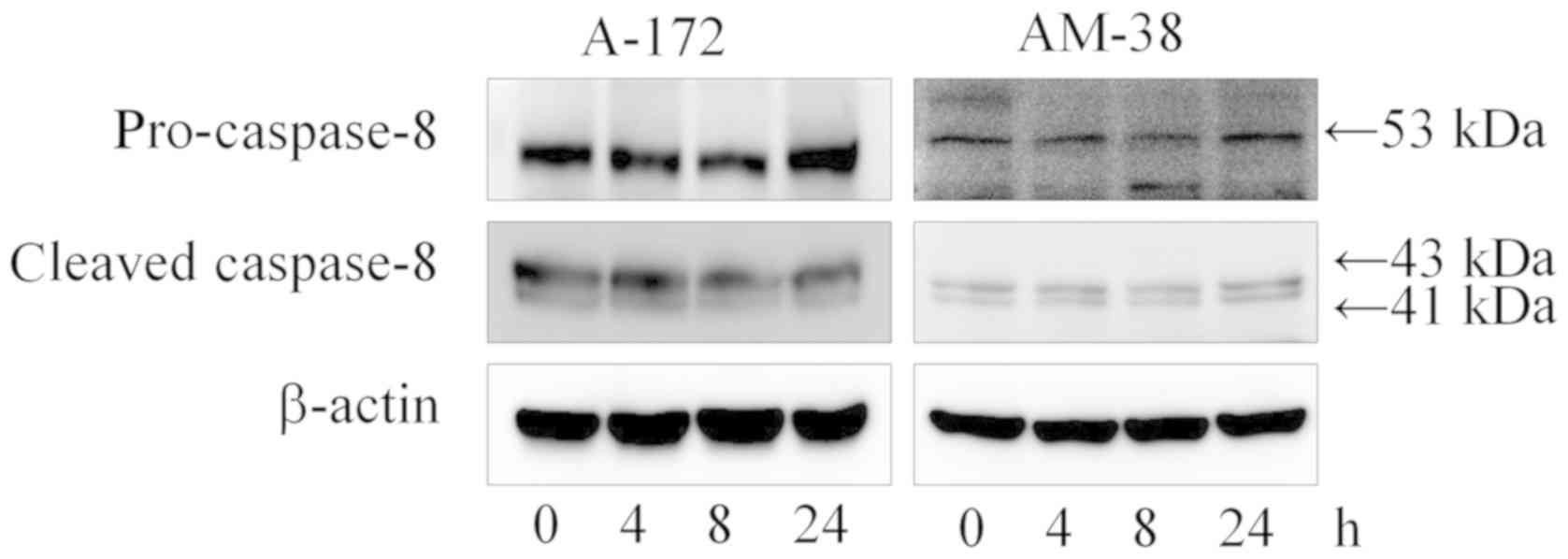
Effects of lenalidomide on the
apoptosis of the A-172 and AM-38 cells
To assess the apoptosis induced by lenalidomide, the
expression levels of apoptotic hallmark proteins including Bax,
caspase-9, caspase-3, cleaved poly-ADP-ribose polymerases (PARP)
and caspase-8 were evaluated by western blot analysis. The data
showed that there was little induction of apoptosis in the A-172
and AM-38 cells (Fig. 5). No band
of cleaved PARP was detected, although the apoptotic hallmarks of
the intrinsic pathway such as Bax and caspase-9 were accumulated
over time following treatment with 10 µM lenalidomide. Cleaved
caspase-9 and cleaved caspase-3 were accumulated during the same
time course. The apoptotic hallmarks of the extrinsic pathway such
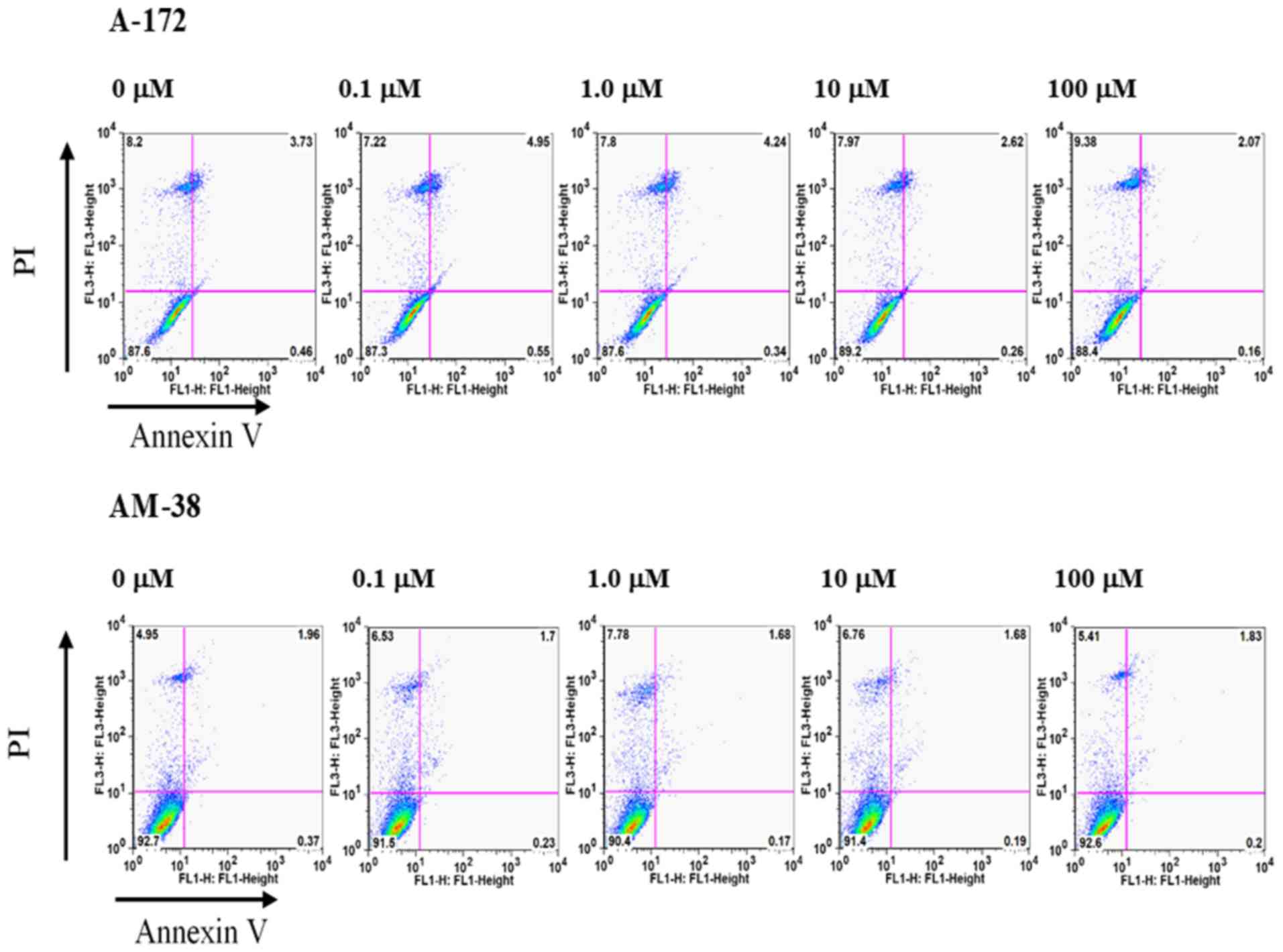
as caspase-8 and cleaved caspase-8 remained unaffected (Fig. 6). The induction of apoptosis by
lenalidomide in A-172 and AM-38 cells was evaluated by flow
cytometry using Annexin V/PI double staining. This analysis
revealed no increased population of early-stage apoptosis or
late-phase apoptosis. After 24, 48, and 72 h of treatment with 0.1,
1.0, 10.0, and 100.0 µM of lenalidomide in the medium, the FACS
data indicated that little apoptosis had occurred (Fig. 7).
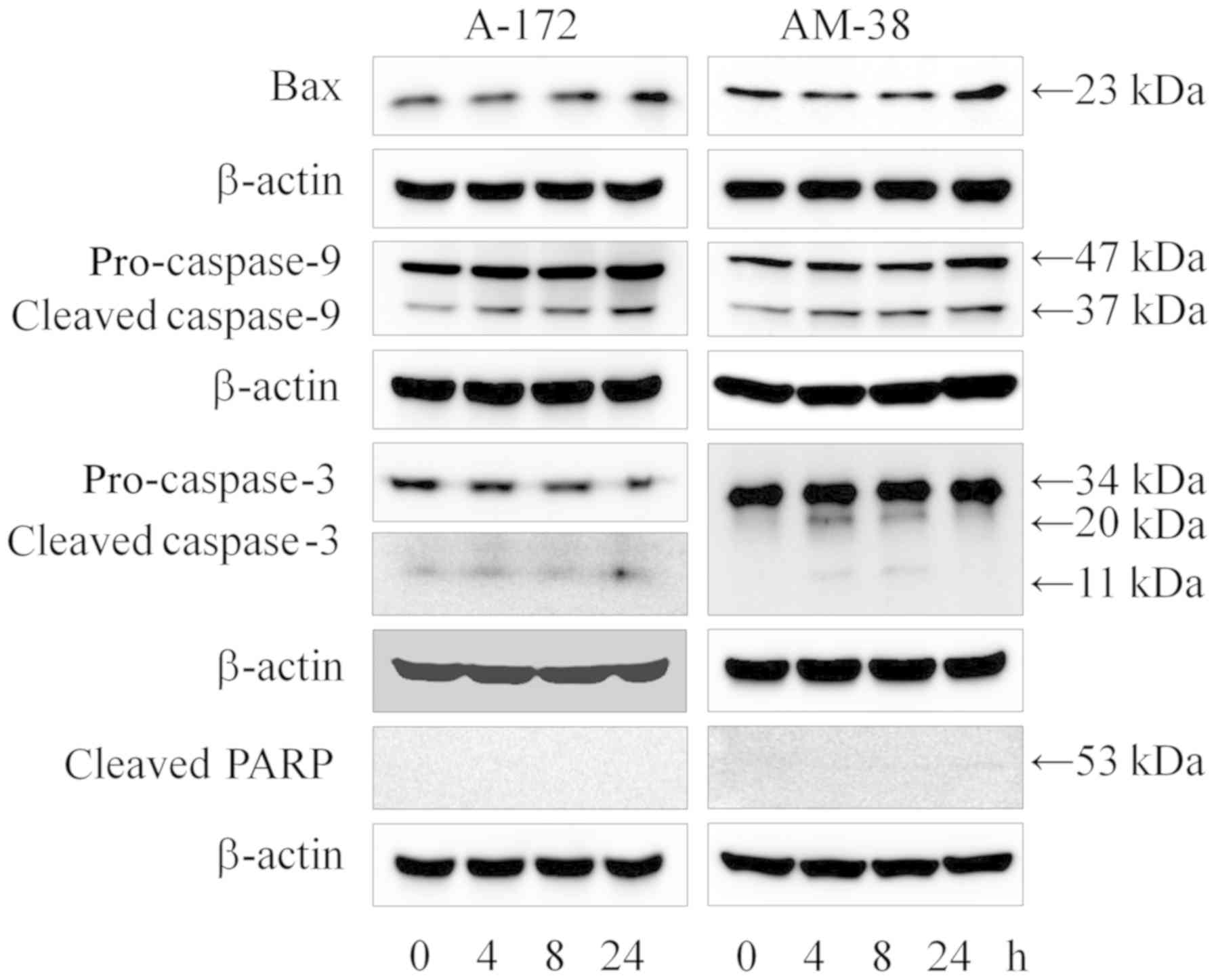 | Figure 5.Effects of lenalidomide on the
intrinsic pathway of apoptosis. Proteins related with the intrinsic
apoptotic pathway hallmarks including Bax, caspase-9 and caspase-3
were investigated by western blot analysis at 0, 4, 8, and 24 h
after lenalidomide treatment, respectively. The data obtained
revealed that the expression of Bax protein was increased at 24 h
in the A-172 and AM-38 cells. Cleavage of caspase-9 protein was
increasingly expressed in the time series following 4, 8, and 24 h
of treatment. The expression of caspase-3 protein cleavage was
increased at 24 h of treatment in the A-172 cells, and increased at
4 and 8 h of treatment in the AM-38 cells. However, cleavage of
PARP was not confirmed. PARP, poly(ADP-ribose) polymerase. |
Discussion
The antitumor activity of lenalidomide has been
previously reported in hematopoietic cells. Kuramitsu et al
demonstrated an effect of lenalidomide on the immune system in
vitro and in vivo (26).
Lenalidomide exerts an antitumor effect due to enhancement of the
cytotoxic effect in T cell interferon-γ secretion. Kuhnol et
al reported that the immune reaction was enhanced by
lenalidomide in in vitro models (27). These studies evaluated the antitumor
effect of immune cells induced by lenalidomide. However, no effect
of lenalidomide on malignant glioma cells has yet been confirmed.
We focused on the direct antitumor effects of lenalidomide, such as
its influence on the cell cycle and induction of apoptosis in
glioma cells. Much of the mechanisms underlying the antitumor
effect in glioma cells remains unknown. It was newly discovered
that lenalidomide showed a direct antitumor effect of cell cycle
arrest in malignant glioma cell lines.
In the Coulter counter assays, the number of
malignant glioma cells was decreased in a concentration-dependent
manner. In most cell lines, an antitumor effect of lenalidomide was
confirmed at a concentration of 10 µM.
We examined the mechanisms underlying the antitumor
effect of lenalidomide using A-172 and AM-38 glioma cell lines. The
glioma cells were exposed to lenalidomide, and we evaluated the
expression levels of various proteins employing western blot
analysis, flow cytometric analysis and mRNA expression analysis
using the RT-qPCR to observe alterations occurring in cell cycle
distribution and apoptosis.
The concentration utilized for our experiments was
10 µM, since our results indicated that the antitumor effect of
lenalidomide was observed at a concentration less than 10 µM. In
addition, Warren's phase I trial demonstrated that the safety dose
of lenalidomide was in the range of 2.5 to 116 mg/m2
(23). Following administration of
20 mg/m2 of lenalidomide, the blood concentration in the
body rises to approximately 2.2 µM with a peak at 1 h (28). We inferred that 10 µM of
lenalidomide was reasonable for clinical use without harmful side
effects.
In previous research concerning lenalidomide
employing hematopoietic cancer cells, various antitumor effects
including a growth inhibition effect, induction of apoptosis,
anti-angiogenic activity and immunological activity were observed
(16,29). In addition, induction of apoptosis
was found to be related to caspase-3 activation. In addition to
these direct antitumor effects, anti-angiogenesis mediated by
suppression of basic fibroblast growth factor (bFGF) and vascular
endothelial growth factor (VEGF) was described (29,30).
In glioma cells, we confirmed that phosphorylated
p53 and functional p53 were activated after 4 h of lenalidomide
treatment based on western blot analysis. In the present study, we
investigated whether the p53/p21 pathway was activated after
treatment with lenalidomide, and it was found that increased p53
and p21 protein expression induced cell cycle arrest in the glioma
cells. Fecteau et al and Escoubet-Lozach et al
reported that activation of p21 occurred after treatment with
lenalidomide; however, functional p53 was not correlated with p21
activation in multiple myeloma cells and chronic lymphocytic
leukemia cells (18). In
hematopoietic cancer cells, the expression of p21 protein was
increased even when p53 protein was knocked out and loading with
lenalidomide treatment was undertaken (31). These observations indicated that the
activation of p21 and functional p53 may be independent in
hematopoietic cancer cells. In glioma cells, it seemed difficult to
conclude that p21 and p53 were dependent; however, lenalidomide did
induce an increased expression of p21, p53, and phosphorylated p53
and cell cycle arrest occurred.
We found that cereblon protein was expressed in all
six glioma cell lines. Recently, it was reported that cereblon is a
target protein for thalidomide, lenalidomide, and its derivatives
(32–34). In hematopoietic malignancies, it was
suggested that the expression level of cereblon regulates the
antitumor effect via p21 (18). We
investigated the expression of cereblon in glioma cells employing
western blot analysis. It was difficult to establish the precise
relationship between the expression level of cereblon and the
antitumor effect. In order to investigate whether cereblon plays a
crucial role in the antitumor effect mediated by lenalidomide,
research using a glioma cell line with knockdown of cereblon will
be carried out.
However, cereblon, a key target protein for
lenalidomide, was expressed in the glioma cells. Previous studies
have shown that lenalidomide directly binds cereblon and promotes
the recruitment of Ikaros (IKZF1) and Aiolos (IKZF3) to the E3
complex containing cereblon. The complex of cereblon with IKZF1 and
IKZF4 leads to ubiquitylation and degradation (35–37).
In the future, we believe that it will also be necessary to
investigate the protein levels of IKZF1 and IKZF3.
We were unable to confirm that lenalidomide
exhibited apoptosis-inducing effects in the glioma cells.
Previously, it was demonstrated that lenalidomide induced apoptosis
in multiple myeloma cells (38). In
particular, lenalidomide activated the intrinsic pathway in mantle
cell lymphoma cells (39). We
hypothesized that lenalidomide may induce apoptosis in glioma cells
as well, and investigated the expression changes in
apoptosis-related proteins using western blot analysis. However, in
the glioma cells, apoptosis was not clearly observed. Western blot
analysis showed that apoptotic hallmarks of the intrinsic pathway
such as Bax, cleaved-caspase-9, and cleaved-caspase-3 were
increased in the A-172 and AM-38 cells. Although the proteins
related to intrinsic apoptosis were activated, cleavage of PARP was
not accumulated. In addition, there was no change in the expression
of the extrinsic pathway such as caspase-8 during the treatment
time courses. The FACS data revealed that only a few glioma cells
were induced to undergo apoptosis. Lenalidomide is known to be an
immunomodulatory agent that has been used to treat hematopoietic
malignancies (8–12). It has been frequently reported that
lenalidomide exhibits an antitumor effect in hematopoietic cells,
but in glioma cells, the situation remains more controversial. The
present study was designed to investigate the direct antitumor
effect of lenalidomide on malignant glioma cell lines including
cell cycle arrest and apoptosis induction. The findings of previous
studies indicated that lenalidomide exerted an antitumor effect on
gliomas especially enhancement of immune reactions such as
interferon secretion from T cells (26,27).
We used a Coulter counter for cell counts. However,
the Coulter counter evaluates only the number of cells present, not
the viable cells. For proliferation or cell viability assays, the
MTT and ATP assays should be employed. We plan to perform further
studies using these assays. Our findings were not sufficient to
yield a conclusive picture concerning the mechanism of action, and
in vivo experiments will also need to be undertaken in the
future. Additionally, the effective concentration for an antitumor
effect remains to be further evaluated in in vivo
experiments. In the present study, we examined the antitumor
effects of single-use lenalidomide, but not any combination use
involving chemotherapy such as with alkylating agents. We need to
confirm whether the effect of lenalidomide in combination use may
be more effective in the case of glioma cells. Lenalidomide is
becoming a key drug in the guidelines for the treatment of
hematopoietic cancer including multiple myeloma (15). Several studies have shown that the
combination use of lenalidomide with alkylating agents or
dexamethasone can be more effective in hematopoietic cancers, and
the chemotherapy regimens for hematopoietic cancers usually employ
lenalidomide combined with alkylating agents, such as melphalan and
cyclophosphamide, and dexamethasone (11,14).
The standard protocol of the chemotherapy for glioblastoma also
includes temozolomide, which is an alkylating agent. Furthermore,
dexamethasone is often used in the postoperative management of
glioblastoma (40). We speculate
that the combination use of lenalidomide with temozolomide and
dexamethasone may have a more potent effect in glioma patients, and
this must be further researched.
In conclusion, lenalidomide displays an antitumor
effect in malignant glioma cells. It induced increased expression
of p21 and p53 and cell cycle arrest did occur, while there was
little effect on apoptosis induction in the glioma cells.
Acknowledgements
Some parts of the present study have been included
in a Japanese-language thesis submitted for the Ph.D. degree of
Yuya Hanashima at Nihon University School of Medicine (Tokyo,
Japan).
Funding
This research study was supported in part by a grant
from the Health Sciences Research Institute, Inc. (Yokohama, Japan)
for the Division of Companion Diagnostics, Department of Pathology
and Microbiology, Nihon University School of Medicine (Tokyo,
Japan).
Availability of data and materials
All data and materials during the study are
available from the corresponding author on reasonable request.
Authors' contributions
YH and AY designed the study. YH, ES and KS
collected the data and drafted the manuscript. YH, ES, YO, CY, JT,
SoY and ShY performed the in vitro experiments. YH analyzed
and interpreted the data and performed the statistical analyses.
TN, TU, HH and AY reviewed the manuscript, designed the figures and
tables. All authors read and approved the manuscript and agree to
be accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stupp R, Hegi ME, Mason WP, van den Bent
MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B,
Belanger K, et al: Effects of radiotherapy with concomitant and
adjuvant temozolomide versus radiotherapy alone on survival in
glioblastoma in a randomised phase III study: 5-year analysis of
the EORTC-NCIC trial. Lancet Oncol. 10:459–466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Singhal S, Mehta J, Desikan R, Ayers D,
Roberson P, Eddlemon P, Munshi N, Anaissie E, Wilson C, Dhodapkar
M, et al: Antitumor activity of thalidomide in refractory multiple
myeloma. N Engl J Med. 341:1565–1571. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Palumbo A, Bringhen S, Caravita T, Merla
E, Capparella V, Callea V, Cangialosi C, Grasso M, Rossini F, Galli
M, et al: Oral melphalan and prednisone chemotherapy plus
thalidomide compared with melphalan and prednisone alone in elderly
patients with multiple myeloma: Randomised controlled trial.
Lancet. 367:825–831. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Facon T, Mary JY, Hulin C, Benboubker L,
Attal M, Pegourie B, Renaud M, Harousseau JL, Guillerm G, Chaleteix
C, et al: Melphalan and prednisone plus thalidomide versus
melphalan and prednisone alone or reduced-intensity autologous stem
cell transplantation in elderly patients with multiple myeloma (IFM
99-06): A randomised trial. Lancet. 370:1209–1218. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Waage A, Gimsing P, Fayers P, Abildgaard
N, Ahlberg L, Björkstrand B, Carlson K, Dahl IM, Forsberg K,
Gulbrandsen N, et al: Melphalan and prednisone plus thalidomide or
placebo in elderly patients with multiple myeloma. Blood.
116:1405–1412. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wijermans P, Schaafsma M, Termorshuizen F,
Ammerlaan R, Wittebol S, Sinnige H, Zweegman S, van Marwijk Kooy M,
van der Griend R, Lokhorst H, et al: Phase III study of the value
of thalidomide added to melphalan plus prednisone in elderly
patients with newly diagnosed multiple myeloma: The HOVON 49 study.
J Clin Oncol. 28:3160–3166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Benboubker L, Dimopoulos MA, Dispenzieri
A, Catalano J, Belch AR, Cavo M, Pinto A, Weisel K, Ludwig H,
Bahlis N, et al: Lenalidomide and dexamethasone in
transplant-ineligible patients with myeloma. N Engl J Med.
371:906–917. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fenaux P, Giagounidis A, Selleslag D,
Beyne-Rauzy O, Mufti G, Mittelman M, Muus P, Te Boekhorst P, Sanz
G, Del Cañizo C, et al: A randomized phase 3 study of lenalidomide
versus placebo in RBC transfusion-dependent patients with
Low-/Intermediate-1-risk myelodysplastic syndromes with del5q.
Blood. 118:3765–3776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gay F, Hayman SR, Lacy MQ, Buadi F, Gertz
MA, Kumar S, Dispenzieri A, Mikhael JR, Bergsagel PL, Dingli D, et
al: Lenalidomide plus dexamethasone versus thalidomide plus
dexamethasone in newly diagnosed multiple myeloma: A comparative
analysis of 411 patients. Blood. 115:1343–1350. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stadtmauer EA, Weber DM, Niesvizky R,
Belch A, Prince MH, San Miguel JF, Facon T, Olesnyckyj M, Yu Z,
Zeldis JB, et al: Lenalidomide in combination with dexamethasone at
first relapse in comparison with its use as later salvage therapy
in relapsed or refractory multiple myeloma. Eur J Haematol.
82:426–432. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zonder JA, Crowley J, Hussein MA, Bolejack
V, Moore DF Sr, Whittenberger BF, Abidi MH, Durie BG and Barlogie
B: Lenalidomide and high-dose dexamethasone compared with
dexamethasone as initial therapy for multiple myeloma: A randomized
Southwest Oncology Group trial (S0232). Blood. 116:5838–5841. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Knop S, Gerecke C, Liebisch P, Topp MS,
Platzbecker U, Sezer O, Vollmuth C, Falk K, Glasmacher A, Maeder U,
et al: Lenalidomide, adriamycin, and dexamethasone (RAD) in
patients with relapsed and refractory multiple myeloma: A report
from the German Myeloma Study Group DSMM (Deutsche Studiengruppe
Multiples Myelom). Blood. 113:4137–4143. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schey SA, Morgan GJ, Ramasamy K, Hazel B,
Ladon D, Corderoy S, Jenner M, Phekoo K, Boyd K and Davies FE: The
addition of cyclophosphamide to lenalidomide and dexamethasone in
multiply relapsed/refractory myeloma patients; a phase I/II study.
Br J Haematol. 150:326–333. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Palumbo A, Hajek R, Delforge M, Kropff M,
Petrucci MT, Catalano J, Gisslinger H, Wiktor-Jędrzejczak W,
Zodelava M, Weisel K, et al: Continuous lenalidomide treatment for
newly diagnosed multiple myeloma. N Engl J Med. 366:1759–1769.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hideshima T, Chauhan D, Shima Y, Raje N,
Davies FE, Tai YT, Treon SP, Lin B, Schlossman RL, Richardson P, et
al: Thalidomide and its analogs overcome drug resistance of human
multiple myeloma cells to conventional therapy. Blood.
96:2943–2950. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lopez-Girona A, Mendy D, Ito T, Miller K,
Gandhi AK, Kang J, Karasawa S, Carmel G, Jackson P, Abbasian M, et
al: Cereblon is a direct protein target for immunomodulatory and
antiproliferative activities of lenalidomide and pomalidomide.
Leukemia. 26:2326–2335. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fecteau JF, Corral LG, Ghia EM, Gaidarova
S, Futalan D, Bharati IS, Cathers B, Schwaederlé M, Cui B,
Lopez-Girona A, et al: Lenalidomide inhibits the proliferation of
CLL cells via a cereblon/p21(WAF1/Cip1)-dependent mechanism
independent of functional p53. Blood. 124:1637–1644. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fine HA, Figg WD, Jaeckle K, Wen PY,
Kyritsis AP, Loeffler JS, Levin VA, Black PM, Kaplan R, Pluda JM
and Yung WK: Phase II trial of the antiangiogenic agent thalidomide
in patients with recurrent high-grade gliomas. J Clin Oncol.
18:708–715. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Giglio P, Dhamne M, Hess KR, Gilbert MR,
Groves MD, Levin VA, Kang SL, Ictech SE, Liu V, Colman H, et al:
Phase 2 trial of irinotecan and thalidomide in adults with
recurrent anaplastic glioma. Cancer. 118:3599–3606. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Penas-Prado M, Hess KR, Fisch MJ, Lagrone
LW, Groves MD, Levin VA, De Groot JF, Puduvalli VK, Colman H,
Volas-Redd G, et al: MD Anderson Community Clinical Oncology
Program; Brain Tumor Trials Collaborative: Randomized phase II
adjuvant factorial study of dose-dense temozolomide alone and in
combination with isotretinoin, celecoxib, and/or thalidomide for
glioblastoma. Neuro Oncol. 17:266–273. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fine HA, Kim L, Albert PS, Duic JP, Ma H,
Zhang W, Tohnya T, Figg WD and Royce C: A phase I trial of
lenalidomide in patients with recurrent primary central nervous
system tumors. Clin Cancer Res. 13:7101–7106. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Warren KE, Goldman S, Pollack IF,
Fangusaro J, Schaiquevich P, Stewart CF, Wallace D, Blaney SM,
Packer R, Macdonald T, et al: Phase I trial of lenalidomide in
pediatric patients with recurrent, refractory, or progressive
primary CNS tumors: Pediatric Brain Tumor Consortium study
PBTC-018. J Clin Oncol. 29:324–329. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ochiai Y, Sano E, Okamoto Y, Yoshimura S,
Makita K, Yamamuro S, Ohta T, Ogino A, Tadakuma H, Ueda T, et al:
Efficacy of ribavirin against malignant glioma cell lines:
Follow-up study. Oncol Rep. 39:537–544. 2018.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kuramitsu S, Ohno M, Ohka F, Shiina S,
Yamamichi A, Kato A, Tanahashi K, Motomura K, Kondo G, Kurimoto M,
et al: Lenalidomide enhances the function of chimeric antigen
receptor T cells against the epidermal growth factor receptor
variant III by enhancing immune synapses. Cancer Gene Ther.
22:487–495. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kühnöl CD, Staege MS and Kramm CM:
Lenalidomide in an in vitro dendritic cell model for malignant
gliomas. Anticancer Agents Med Chem. 16:1468–1473. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen N, Zhou S and Palmisano M: Clinical
pharmacokinetics and pharmacodynamics of Lenalidomide. Clin
Pharmacokinet. 56:139–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
D'Amato RJ, Loughnan MS, Flynn E and
Folkman J: Thalidomide is an inhibitor of angiogenesis. Proc Natl
Acad Sci USA. 91:4082–4085. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rao KV: Lenalidomide in the treatment of
multiple myeloma. Am J Health Syst Pharm. 64:1799–1807. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Escoubet-Lozach L, Lin IL, Jensen-Pergakes
K, Brady HA, Gandhi AK, Schafer PH, Muller GW, Worland PJ, Chan KW
and Verhelle D: Pomalidomide and lenalidomide induce p21 WAF-1
expression in both lymphoma and multiple myeloma through a
LSD1-mediated epigenetic mechanism. Cancer Res. 69:7347–7356. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Holstein SA, Hillengass J and McCarthy PL:
Next-generation drugs targeting the cereblon ubiquitin ligase. J
Clin Oncol. 36:2101–2104. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao J, Yang J and Xu G: The interacting
domains in cereblon differentially modulate the immunomodulatory
drug-mediated ubiquitination and degradation of its binding
partners. Biochem Biophys Res Commun. 507:443–449. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zheng J, Sha Y, Roof L, Foreman O,
Lazarchick J, Venkta JK, Kozlowski C, Gasparetto C, Chao N, Ebens
A, et al: Pan-PIM kinase inhibitors enhance Lenalidomide's
anti-myeloma activity via cereblon-IKZF1/3 cascade. Cancer Lett.
440-441:1–10. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Krönke J, Udeshi ND, Narla A, Grauman P,
Hurst SN, McConkey M, Svinkina T, Heckl D, Comer E, Li X, et al:
Lenalidomide causes selective degradation of IKZF1 and IKZF3 in
multiple myeloma cells. Science. 343:301–305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lu G, Middleton RE, Sun H, Naniong M, Ott
CJ, Mitsiades CS, Wong KK, Bradner JE and Kaelin WG Jr: The myeloma
drug lenalidomide promotes the cereblon-dependent destruction of
Ikaros proteins. Science. 343:305–309. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mori T, Ito T, Liu S, Ando H, Sakamoto S,
Yamaguchi Y, Tokunaga E, Shibata N, Handa H and Hakoshima T:
Structural basis of thalidomide enantiomer binding to cereblon. Sci
Rep. 8:1294. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yao R, Han D, Sun X, Xie Y, Wu Q, Fu C,
Yao Y, Li H, Li Z and Xu K: Scriptaid inhibits cell survival, cell
cycle, and promotes apoptosis in multiple myeloma via epigenetic
regulation of p21. Exp Hematol. 60:63–72. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qian Z, Zhang L, Cai Z, Sun L, Wang H, Yi
Q and Wang M: Lenalidomide synergizes with dexamethasone to induce
growth arrest and apoptosis of mantle cell lymphoma cells in vitro
and in vivo. Leuk Res. 35:380–386. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pitter KL, Tamagno I, Alikhanyan K,
Hosni-Ahmed A, Pattwell SS, Donnola S, Dai C, Ozawa T, Chang M,
Chan TA, et al: Corticosteroids compromise survival in
glioblastoma. Brain. 139:1458–1471. 2016. View Article : Google Scholar : PubMed/NCBI
|