Introduction
Renal cell carcinoma (RCC) is one of the most
prevalent malignancies of the urinary system, accounting for 2.2%
of all adult malignancies globally, with estimations of 403,262 new
cases and 175,098 deaths worldwide in 2018 (1). Clear cell (cc)RCC represents the
predominant histologic subtype of RCC and constitutes ~70% of all
cases (2). The remaining 30% of RCC
subtypes, such as chromophobe and papillary RCC, are generally
indolent (3). Surgery is the most
effective treatment for ccRCC, as chemotherapy and radiotherapy are
not as successful in controlling its progression; however, ~25% of
clinical patients with ccRCC will develop metastatic disease
despite curative surgical removal of the primary tumor (4). The overall 5-year survival rate of RCC
patient is 5–10%, with a median survival of only ~13 months
(5). Therefore, the molecular
mechanisms underlying ccRCC initiation and progression require
improved understanding, which may contribute to the development of
novel strategies for treating ccRCC.
Ubiquitin and ubiquitin-like modifications
participate in most cellular signaling pathways (6). Ubiquitination is reversed by
deubiquitinating enzymes (DUBs) that cleave ubiquitin from the
substrate protein (7). There are
~98 DUBs known to be encoded by the human genome, which are grouped
into six subfamilies, the largest one being the ubiquitin specific
peptidase (USP) family (8). The USP
family exhibits a conserved three-domain architecture, comprising
the palm, thumb and fingers of a right hand (9). USP19 belongs to the USP family and
several studies have reported that USP19 exhibits diverse roles in
biological processes. For example, USP19 is involved in the
endoplasmic reticulum-associated degradation pathway and the
unfolded protein response (10).
Additionally, USP19 has also been identified as a positive
regulator involved in regulating autophagy and innate immune
responses (11). Furthermore, USP19
also reported to have involved in the pathogenesis of certain
cancers. For example, Ewing sarcoma breakpoint region 1-Friend
leukemia integration 1 (EWS-FLI1) is a chimeric oncoprotein
uniquely expressed in tumor cells, and constant EWS-FLI1 protein
turnover is regulated by the ubiquitin proteasome system (12). USP19 as a specific modulator of
EWS-FLI1 protein stability, which mediates an increase in fusion
protein deubiquitination by binding to its N-terminal domain to
regulate Ewing sarcoma growth (12). In addition, BRCA1-associated protein
1 (BAP1) is a tumor suppressor gene important to the development
and prognosis of numerous cancers (13); Shahriyari et al (13) found that USP19 was highly positively
associated with BAP1 expression in breast and uveal melanoma. At
present, there is limited knowledge regarding the role and impact
of USP19 in other cancers, including ccRCC.
In the present study, the expression and clinical
significance of USP19 were explored in The Cancer Genome Atlas
(TCGA) and Gene Expression Omnibus (GEO) ccRCC samples, and the
biological functions of USP19 in ccRCC were investigated in
vitro and in vivo.
Materials and methods
Date sources
Level 3 mRNA sequencing (seq) data (downloaded in
November 2016) of ccRCC, also known as kidney renal clear cell
carcinoma (KIRC), including 539 tumor samples and 72 adjacent
normal samples, were downloaded from TCGA database (KIRC RNA-seq;
http://www.cbioportal.org/). Clinical
information, including age, sex, histological grade, tumor stage,
survival time and outcome, were also extracted from TCGA. The copy
numbers of 587 ccRCC tissues and 901 paired normal kidney tissues
from the TCGA database (KIRC CNV) were obtained. GSE76207 (14) and GSE102101 (15) were obtained from the GEO database
(http://www.ncbi.nlm.nih.gov/geo/), and
respectively contained 16 and 10 paired tumor and normal kidney
tissues.
Bioinformatics analysis of the USP19
transcriptional levels in ccRCC
The KIRC RNA-seq level 3 count data and GSE76207
data were normalized with the negative binomial distribution
methodology by R (version 3.4.1) (16) package DESeq2 (17). The GSE102101 sequence reads were
mapped to the human genome (hg38) using HISAT2 (version 2.1.0;
http://ccb.jhu.edu/software/hisat2/)
and transcripts were quantified using stringtie (version 1.3.4;
http://ccb.jhu.edu/software/stringtie/) with refseq
annotation, normalized and subjected to differential expression
analysis using DESeq2.
Copy number variation analysis
The KIRC copy-number variation (CNV) data were
annotated by bedtools (https://bedtools.readthedocs.io/).
Subgroup analysis based on
clinicopathologic features
To further explore the role of USP19, KIRC tumor
samples were divided into various groups based on their
clinicopathological features (age, sex, histological grade and
stage). Then, differential expression analysis of the tumor samples
in each group was conducted by t-test.
Survival analysis
Gene Expression Profiling Interactive Analysis
(http://gepia.cancer-pku.cn/) was used to
analyze the overall survival (OS) and disease-free survival (DFS)
differences between patients with high or low expression levels of
USP19. Patients above the upper quartile with respect to USP19
expression were classified as having a high expression levels,
whereas those below the lower quartile were classified as having a
low expression level. Those patients with USP19 expression values
between the lower and upper quartiles were excluded from survival
analysis. Kaplan-Meier analysis and a log-rank test was used to
calculate the significance of survival time differences between the
two classes of patients.
Plasmid construction and
transfection
Full-length human USP19 mRNA was cloned from a human
cDNA library and inserted into the mammalian expression vector
pHAGE-3×flag (Invitrogen; Thermo Fisher Scientific, Inc.) to
construct a pHAGE-3×flag-USP19 plasmid. Two short hairpin RNAs
(shRNAs) targeting different regions of USP19 mRNA (shUSP19#1 and
shUSP19#2) were inserted into the pLKO.1 vector (OligoEngine) and
confirmed by sequencing. The sequences of the shRNAs were
5′-CTCCACTGCGAGCGAAGTATT-3′ (shUSP19#1),
5′-CCGGTACTCTGTGAGTGTATT-3′ (shUSP19#2) and
5′-GUCAGCGUGCAGAUAGAGUUU-3′ [control shRNA (shctrl)]. Lentiviral
supernatants were harvested 48 h after 293T cells at 80–90%
confluency were co-transfected with packing plasmid (0.5 µg pMD2.G
and 0.75 µg psPAX2; Novagen; Merck KGaA) and 1 µg
pHAGE-3×flag-USP19, empty pHAGE-3×flag, shUSP19#1, shUSP19#2 or
shctrl plasmid using polyetherimide transfection reagent (cat. no.
GF95977287; Sigma-Aldrich; Merck KGaA). The lentivirus was
harvested 48 h after transfection, and Caki-1 cells
(5×105 cells/well) were incubated with
lentivirus-containing medium (MOI=50) supplemented with 2 µl
polybrene (Sigma-Aldrich; Merck KGaA). At 48 h later, stable Caki-1
cells were selected in the presence of 4 µg/ml puromycin
(Sigma-Aldrich; Merck KGaA) for 2 days. USP19 mRNA expression was
examined by reverse transcription-quantitative PCR (RT-qPCR) and
protein expression was evaluated using western blotting.
Cell lines and reagents
293T cells were obtained from American Type Culture
Collection. The human renal cancer cell line Caki-1 was purchased
from the Cell Bank of the Type Culture Collection of the Chinese
Academy of Sciences. 293T and Caki-1 cells were cultured in DMEM
(cat. no. C11995500BT; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (cat. no. F04-001; Biological Industries)
and 1% penicillin/streptomycin (cat. no. 15140-122; Gibco; Thermo
Fisher Scientific, Inc.), and the cells were cultured in a
humidified atmosphere containing 5% CO2 at 37°C. U0126
(cat. no. HY-12031; MedChemExpress LLC), an ERK inhibitor, was
dissolved in DMSO and used to treat Caki-1 cells at a concentration
of 20 µm.
Cell proliferation
Cell Counting Kit-8 (CCK-8) assays were performed to
evaluate the proliferation capacity of Caki-1 cells. Cells were
seeded into 96-well plates at a density of 3×103/well.
After incubation for various timepoints (0, 24, 48, 72 and 96 h),
10 µl CCK-8 reagent (Dojindo Molecular Technologies, Inc.) was
added to each well, followed by incubation for a further 4 h at
room temperature. Then, the cell proliferation ability was examined
by measuring the absorbance at 450 nm using an enzyme label
analyzer.
For soft agar assays, 3,000 cells were seeded onto
6-well plates containing 0.5% top agarose and 1% bottom agarose,
and incubated for 2 weeks in a humidified atmosphere containing 5%
CO2 at 37°C. The number of colonies in each plate was
counted using Image J software version 1.47 (National Institutes of
Health).
Transwell migration assay
Migration assays were performed using 5-mm
Transwell® inserts with 5-µm pore polycarbonate
membranes (Corning, Inc.). A 0.5-ml cell suspension
(2.5×104 cells) in serum-free culture medium was added
to the upper chambers, and 750 µl culture medium containing 10% FBS
was added to the lower chambers. After 24 h, the cells on the upper
surface of the membrane were quickly wiped away with cotton-tipped
swabs, and the cells on the lower surface were fixed with 4%
paraformaldehyde for 15 min at room temperature, following by
staining with a 0.1% crystal violet solution (Sigma-Aldrich; Merck
KGaA) for 30 min at room temperature. Cell counting was performed
by photographing the membrane through a light microscope. Four
random microscopic fields (magnification, ×100) in each chamber
were photographed.
Wound healing assay
Cells were seeded at 5×105 cells/well
into 6-well plates. After the cells reached ~90% confluence, three
separate wounds were scratched through the cells with a sterile
100-µl pipette tip. Floating cells were removed with PBS and the
medium was replaced with serum-free medium. Images were captured at
0 and 48 h following the initial scratch to evaluate cell
migration. Cell migration rate (%)=[(scratch width at 0 h-scratch
width at 48 h)/original scratch width × 100%].
RNA extraction and RT-qPCR
Total RNA from cells were extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. For qPCR assays,
total RNA was reverse transcribed into cDNA using a Transcriptor
First Strand cDNA Synthesis Kit (Roche Diagnostics) according to
the manufacturer's protocols. The expression of USP19 was measured
by qPCR using SYBR Green PCR Master Mix (Roche Diagnostics). qPCR
was performed as follows: Initial denaturation for 2 min at 55°C
and 30 sec at 95°C, followed by 35 cycles of 95°C for 5 sec and
60°C for 30 sec. 18S ribosomal RNA was used as an internal control,
and the relative mRNA expression was estimated using the
2−∆∆Cq method (18). The
primer sequences are shown in Table
I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Primer |
|---|
| p27 | F:
5′-GGTGGAGAAGGGCAGCTTG-3′ |
|
| R:
5′-GAAGAATCGTCGGTTGCAGG-3′ |
| PCNA | F:
5′-CACTCCACTCTCTTCAACGGT-3′ |
|
| R:
5′-ATCCTCGATCTTGGGAGCCA-3′ |
| Cyclin D1 | F:
5′-CAGATCATCCGCAAACACGC-3′ |
|
| R:
5′-AGGCGGTAGTAGGACAGGAA-3′ |
| MMP2 | F:
5′-CCGTCGCCCATCATCAAGTT-3′ |
|
| R:
5′-CCGCATGGTCTCGATGGTAT-3′ |
| MMP9 | F:
5′-TTTGAGTCCGGTGGACGATG-3′ |
|
| R:
5′-TTGTCGGCGATAAGGAAGGG-3′ |
| USP19 | F:
5′-TAAATCCAAGGCACGATCTGAGG-3′ |
|
| R:
5′-GCTTTGGGGTTACATGCTCCA-3′ |
| 18S rRNA | F:
5′-GTAACCCGTTGAACCCCATT-3′ |
|
| R:
5′-CCATCCAATCGGTAGTAGCG-3′ |
Protein extraction and western blot
assay
Total protein was extracted using RIPA lysis buffer
(50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% Triton X-100, 1% sodium
deoxycholate, 0.1% SDS and 1 mM EDTA) containing a protease
inhibitor cocktail (cat. no. 4963124001; Roche Diagnostics). The
protein concentrations were quantified using a bicinchoninic acid
assay kit, and 30–50 µg protein was boiled for 5 min and used for
western blot analysis. The proteins were separated by 8–12%
SDS-PAGE and transferred to PVDF membranes at 200 mA for 1 h. The
PVDF membranes were blocked with 5% nonfat milk at room temperature
for 1 h. Next, the membranes were incubated with primary antibodies
(1:1,000) at 4°C overnight and washed with TBS-0.1% Tween-20 three
times. The membranes were further incubated with horseradish
peroxidase (HRP)-conjugated secondary antibodies (1:10,000; cat.
nos. 111-035–003 and 115-035-003; Jackson ImmunoResearch
Laboratories, Inc.) at 37°C for 1 h. Finally, the membranes were
developed by Femto ECL substrates (Thermo Fisher Scientific, Inc.)
and captured by a ChemiDoc system (Bio-Rad Laboratories, Inc.)
containing a Chemi HR camera. The images were analyzed by Image Lab
3.1 software (Bio-Rad Laboratories, Inc.). Primary antibodies
targeting USP19 (cat. no. ab93159; Abcam), p27 (cat. no. 3688; Cell
Signaling Technology, Inc.), proliferating cell nuclear antigen
(PCNA; cat. no. 2586; Cell Signaling Technology, Inc.), cyclin D1
(cat. no. 2922; Cell Signaling Technology, Inc.), matrix
metalloproteinase (MMP)2 (cat. no. A11144; ABclonal Biotech Co.,
Ltd.), MMP9 (cat. no. A0289; ABclonal Biotech Co., Ltd.),
phosphorylated (p)-ERK (cat. no. 4370; Cell Signaling Technology,
Inc.), ERK (cat. no. 4695; Cell Signaling Technology, Inc.),
β-actin (cat. no. 4967; Cell Signaling Technology, Inc.) were
used.
Xenograft model
All animal protocols conducted in the present study
were approved by the Institutional Animal Care and Use Committee of
the Institute of Model Animals of Wuhan University. All animal care
outlined in our study were performed in adherence with the Guide
for the Care and Use of Laboratory Animals published by US National
Institutes of Health (19). The
following criteria were set as humane endpoints for the study:
Abnormal behavior; ±20% change in body weight; tumor diameter
>1.5 cm; or tumor ulceration. The maximum tumor diameter was not
exceeded until day 60 of the study, when animals were sacrificed;
the maximum diameter observed in any one direction was 1.95 cm.
A total of 10 BALB/c female mice (4–6 weeks; 17–18
g) were purchased from Beijing Vital River Laboratory Animal
Technology Co., Ltd. and housed under specific pathogen-free
conditions and a 12-h light/dark cycle, at 25°C with 60±10%
humidity, and free access to food and water. A total of
5×106 stably infected cells were diluted in 150 µl DMEM
and Matrigel, and inoculated subcutaneously into the left flank of
each mouse (5 mice/group). The tumor volumes (V) were measured
every 5 days, and calculated using the following formula: V=0.5×
(length × width2). At 60 days after injection, mice were
sacrificed via cervical dislocation. If no spontaneous breathing
was observed for 3 min and the blink reflex was lost, mice were
considered to be euthanized. During the study, no animals were
prematurely euthanized or found dead. The tumors were removed from
the mice, weighed and used for western blot assays.
Statistical analysis
The data were presented as the mean ± SD of ≥3
experimental repeats using GraphPad Prism 5 software (GraphPad
Software, Inc.). Comparisons between two groups were performed
using two-tailed Student's t-tests. Differences among 3 groups were
assessed using one-way ANOVA followed by Bonferroni test (equal
variances assumed) or Tamhane's T2 test (equal variances not
assumed). Differences among 4 groups were assessed using two-way
ANOVA followed by Bonferroni test. Statistical analyses were
performed using SPSS 19 Statistics software (IBM Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
USP19 is downregulated due to copy
number loss in ccRCC tissues
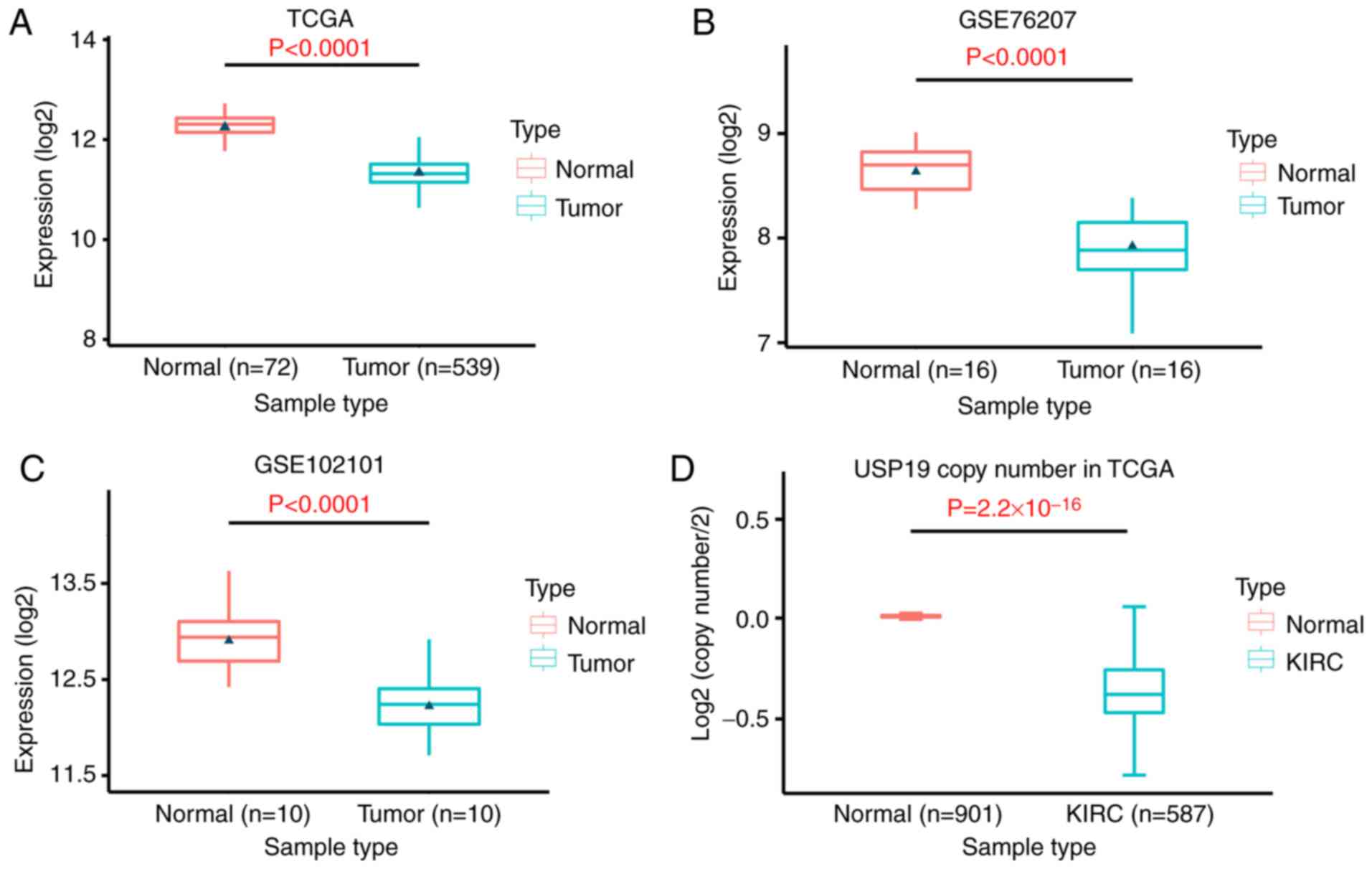
To explore whether USP19 was involved in the
pathogenesis of ccRCC, its expression was first detected in
patients with ccRCC and healthy controls. RNA-seq data from the
samples of 539 patients with ccRCC from TCGA database and 26
patients with ccRCC from GEO (GSE76207 and GSE102101) were
employed. The results indicated that USP19 expression was
significantly reduced in renal cancer tissues compared with in
normal renal tissues (P<0.0001; Fig.
1A-C). DNA CNVs are common in cancer and are responsible for
the dysregulation of gene expression (20). To explain the mechanism of USP19
downregulation in ccRCC, the CNVs of the USP19 gene in 901 normal
kidney tissue samples and 587 KIRC samples from TCGA were assessed.
The results demonstrated that the USP19 gene copy number was
significantly lower in ccRCC tissues than in normal tissues
(P=2.2×10−16; Fig. 1D).
These results indicated that USP19 expression is significantly
decreased in ccRCC tissues, which may be due to copy number
loss.
USP19 downregulation is associated
with high tumor stage and poor prognosis
The associations between USP19 expression and
clinicopathological features were further evaluated in TCGA KIRC
cohort. Data showed that USP19 expression levels were significantly
reduced in patients at a more advanced clinical stage (P<0.0001)
and higher histological grade, (P=0.0086), but was not significant
different between patients separated by age (P=0.2254) or sex
(P=0.0933; Table II).
| Table II.Relationship between the expression
of USP19 and clinicopathological characteristics. |
Table II.
Relationship between the expression
of USP19 and clinicopathological characteristics.
| Characteristic | Cases, n | USP19 mean
expression value | P-value |
|---|
| Age |
|
| 0.2254 |
|
≤60 | 249 |
2,749.977±913.719 |
|
|
>60 | 289 |
2,661.380±755.453 |
|
| Sex |
|
| 0.0933 |
|
Male | 353 |
2,658.620±827.082 |
|
|
Female | 186 |
2,785.718±837.334 |
|
| Histological
grade |
|
| 0.0086 |
|
Low | 249 |
2,775.497±890.832 |
|
|
High | 282 |
2,592.384±677.281 |
|
| Stage |
|
| <0.0001 |
|
I–II | 331 |
2,832.380±961.756 |
|
|
III–IV | 205 |
2,496.370±507.991 |
|
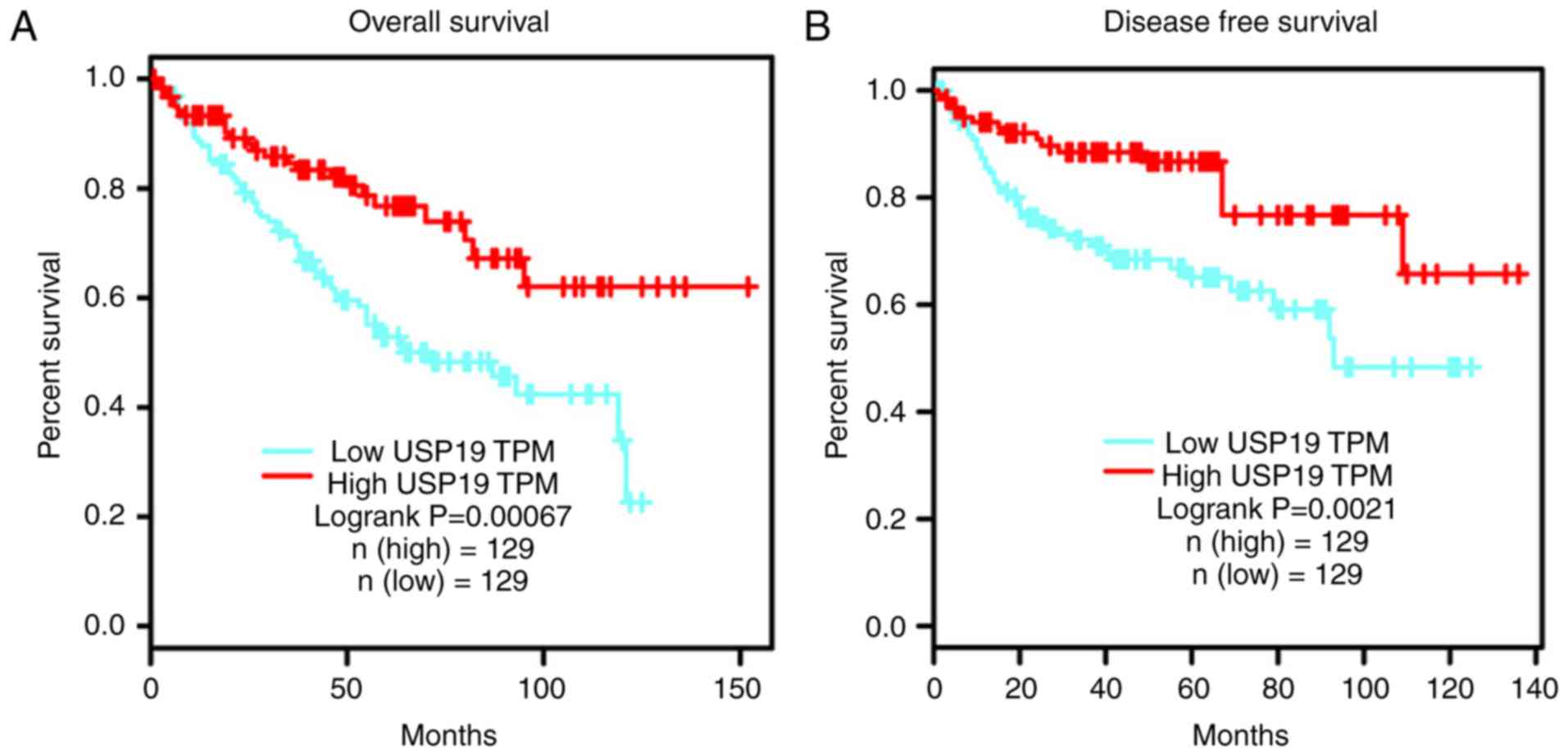
Survival analysis revealed that patients with ccRCC
with low USP19 expression exhibited significantly worse OS and DFS
compared with those with high USP19 expression (P<0.05; Fig. 2A and B). In conclusion, these
results suggested that USP19 downregulation is associated with high
tumor stage and poor prognosis.
Overexpression of USP19 inhibits ccRCC
cell proliferation in vitro
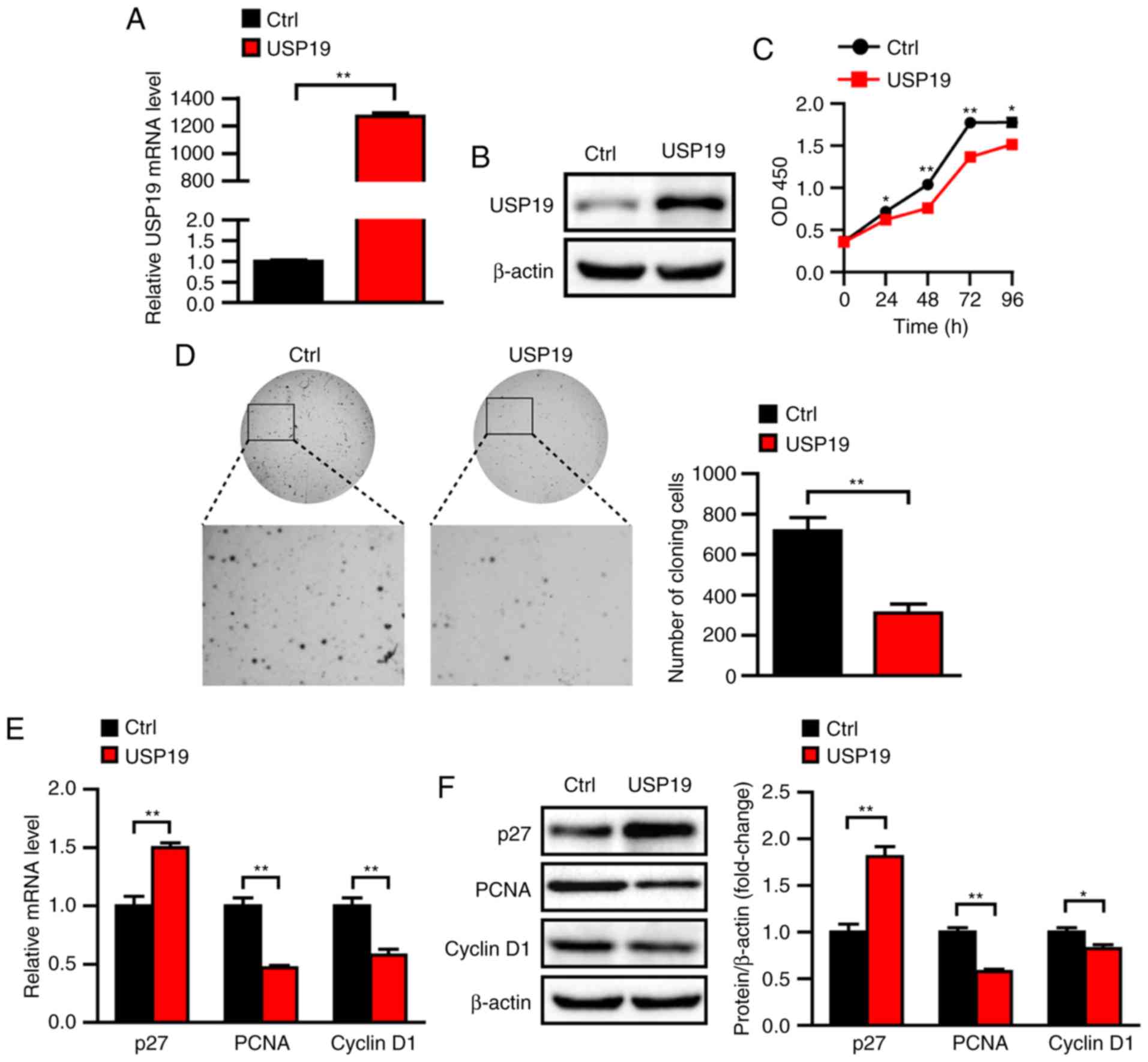
To explore the potential biological functions of
USP19 in ccRCC, USP19-overexpressing cells were constructed by
lentiviral infection of Caki-1 cells in vitro. USP19 mRNA
and protein levels were significantly increased in
USP19-overexpressing cells (P<0.05; Fig. 3A and B). Caki-1 cell proliferation
was assessed via CCK-8 assays, and the results showed that
proliferation was significantly reduced in USP19-overexpressing
Caki-1 cells compared with in control cells (all P<0.05;
Fig. 3C). Furthermore, soft agar
assays showed that the colony number was significantly decreased
for USP19-overexpressing Caki-1 cells compared with control cells
(P<0.01; Fig. 3D). Previous
reports indicated that cyclin D1 and PCNA are involved in cell
cycle control (21) and that p27 is
a cyclin-dependent kinase inhibitor (22); these genes are thus markers of cell
proliferation. Thus, mRNA levels of PCNA, cyclin D1, and p27 were
evaluated via RT-qPCR. The results showed that mRNA levels of PCNA
and cyclin D1 were downregulated when USP19 was overexpressed,
whereas p27 was upregulated (all P<0.05; Fig. 3E). Consistently, the protein levels
of PCNA and cyclin D1 were decreased, and those of p27 were
increased in USP19-ovexpressing cells compared with control cells
(all P<0.05; Fig. 3F). Taken
together, the above results indicated that USP19 inhibits ccRCC
cell proliferation.
Knockdown of USP19 expression promotes
ccRCC cells proliferation in vitro
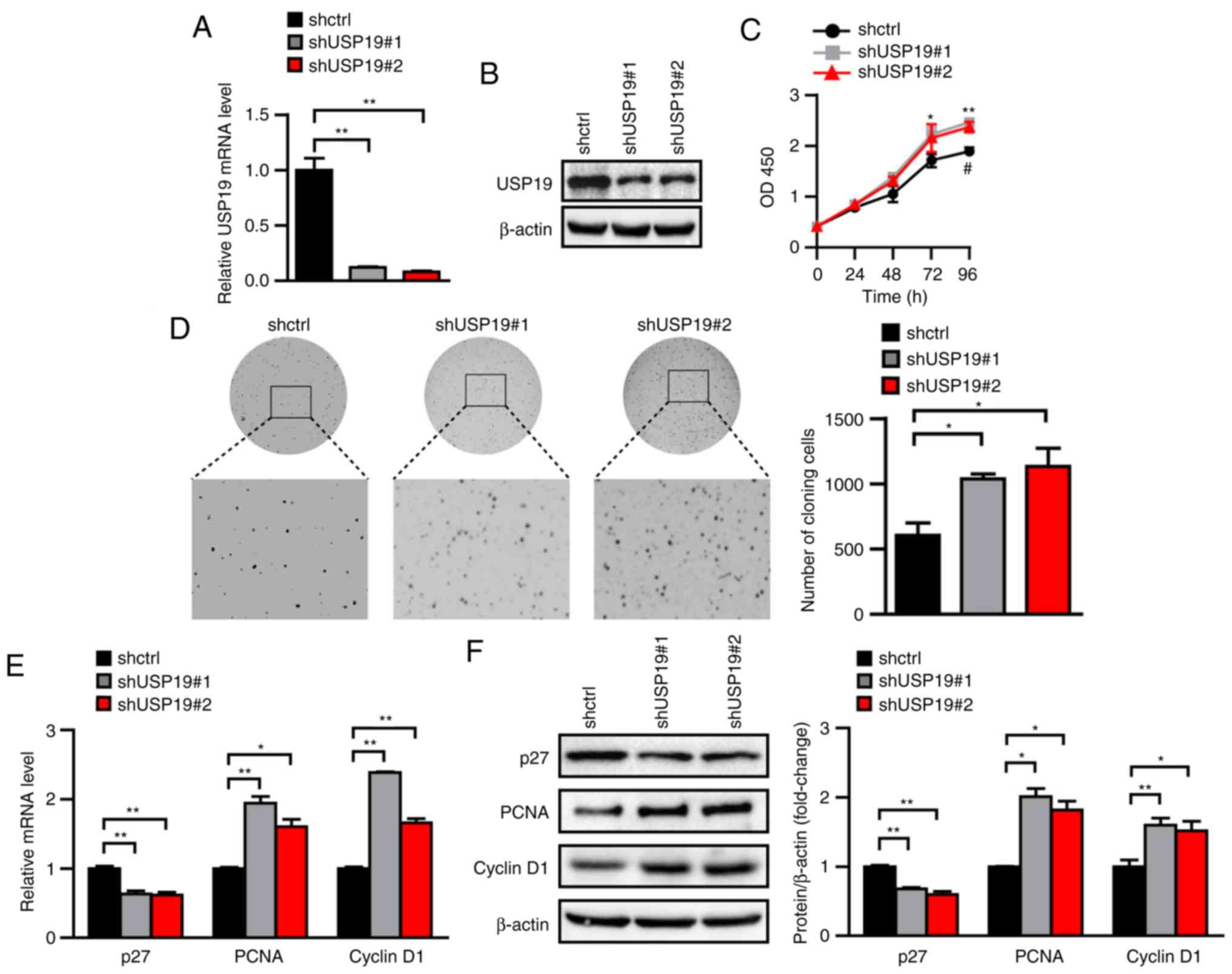
To examine whether USP19 knockdown promotes ccRCC
cell proliferation, lentivirus-mediated shRNA constructs targeting
different USP19 mRNA regions were applied to generate two stable
USP19 knockdown cell lines. USP19 mRNA and protein levels were
significantly reduced in stable USP19 knockdown Caki-1 cell lines
compared with in shctrl Caki-1 cell lines (all P<0.05; Fig. 4A and B). CCK-8 and soft agar assays
also showed that Caki-1 cell proliferation was increased following
USP19 knockdown (all P<0.05; Fig. 4C
and D). Accordingly, the mRNA levels of PCNA and cyclin D1 were
higher, and those of p27 were lower in the USP19 knockdown group
compared with in the shctrl group (all P<0.05; Fig. 4E). Similarly, the protein levels of
PCNA and cyclin D1 were increased, while those of p27 were
decreased by USP19 knockdown (all P<0.05; Fig. 4F). These results indicated that
USP19 knockdown promotes ccRCC cell proliferation.
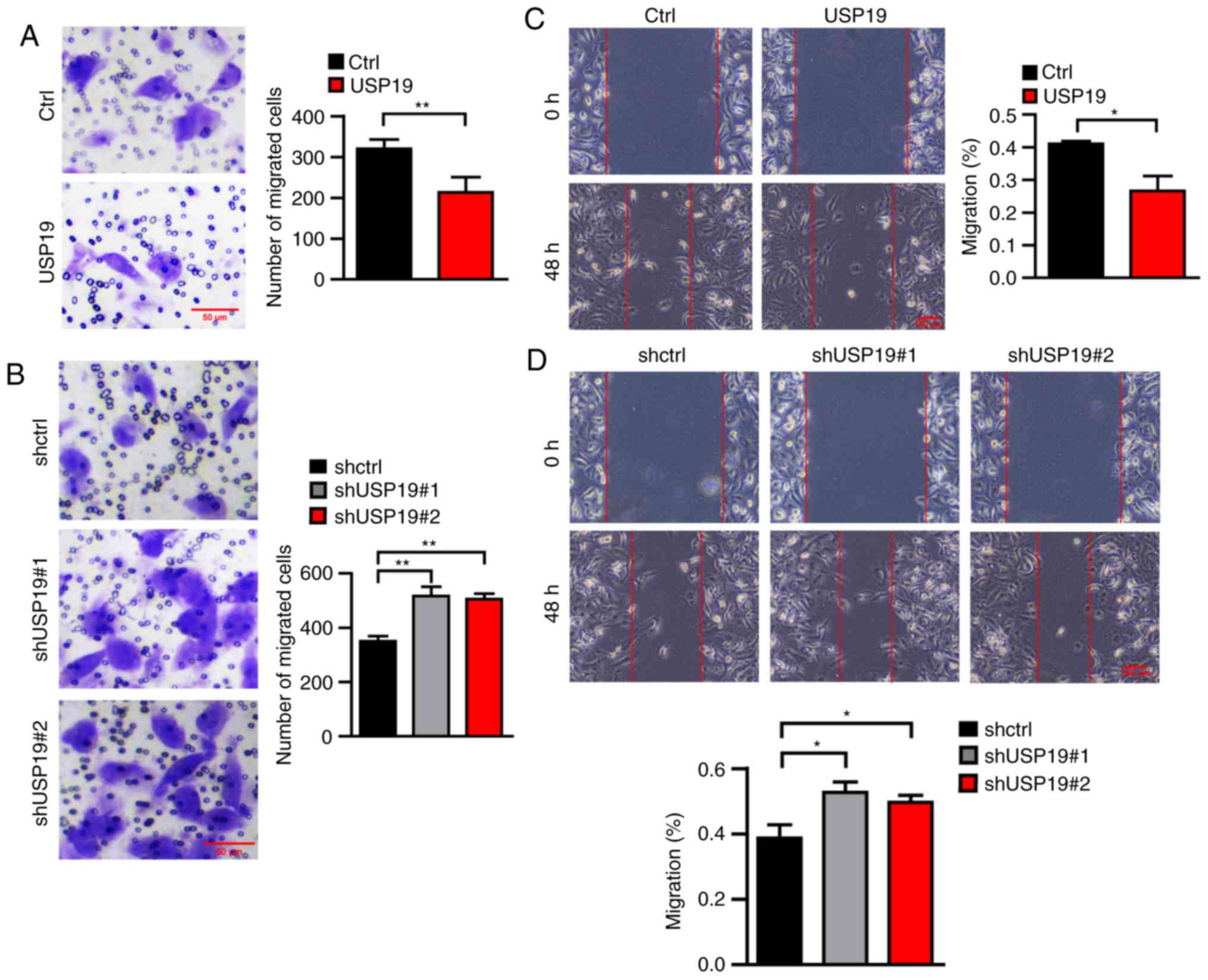
USP19 inhibits ccRCC cell migration in
vitro
Having observed a significant change in cell
proliferation following the dysregulation of USP19 expression, the
effects of USP19 on cell migration were further evaluated using
Transwell and wound healing assays. Transwell assays showed that
cell migration was impaired in USP19-overexpressing Caki-1 cells,
whereas it was promoted in USP19 knockdown Caki-1 cells compared
with the respective negative controls (all P<0.01; Fig. 5A and B). Similarly, microscopic
examination of wounds at 0 and 48 h showed that the migration rate
in the USP19 overexpression group was significantly reduced,
whereas it was increased in USP19 knockdown group compared with the
control groups (all P<0.05; Fig. 5C
and D). MMP2 and MMP9 are frequently with the malignant
phenotype of tumor cells, and are commonly used as malignant tumor
migration markers (23,24). RT-qPCR and western blot data showed
that the expression levels of MMP2 and MMP9 were lower in
USP19-overexpressing cells and higher in USP19-knockdown cells than
in control cells (all P<0.05; Fig.
5E-H). These data indicated that USP19 inhibits ccRCC cell
migration in vitro.
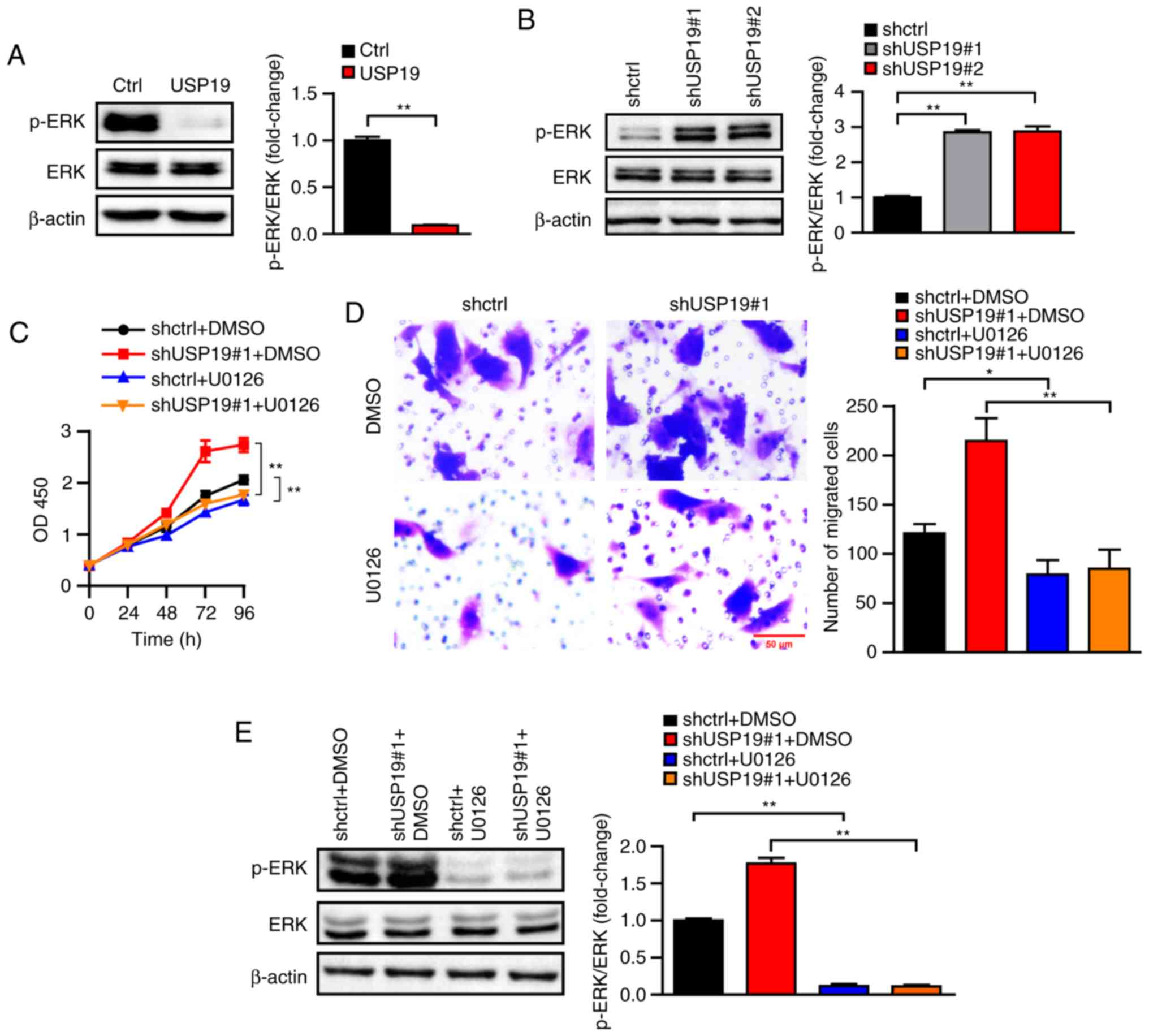
USP19 regulates ccRCC cell
proliferation and migration by regulating the ERK signaling
pathway
ERK/mitogen-activated protein kinase (MAPK)
signaling has been shown to be involved in various
cancer-associated pathological processes (25,26).
To investigate the underlying molecular signaling pathways that
participate in USP19-mediated inhibition of proliferation and
migration in ccRCC cancer cells, ERK activity was determined in
USP19-overexpressing and knockdown Caki-1 cells. Of note, it was
found that total ERK protein levels were not notably changed in the
USP19 knockdown or overexpression ccRCC cells. However, USP19
overexpression clearly reduced p-ERK levels, while USP19 knockdown
increased ERK phosphorylation (all P<0.01; Fig. 6A and B), indicating that ERK may
mediate the functions of USP19 in ccRCC cancer cells. Thus, the
effects of the ERK inhibitor U0126 on the functions of USP19 in
ccRCC cancer cells were determined. As the CCK-8 assay showed,
U0126 significantly rescued the increased Caki-1 cell proliferation
induced by USP19 knockdown (all P<0.01; Fig. 6C). Transwell assays also indicated
that the promoting effects of USP19 knockdown on Caki-1 cell
migration were reversed by U0126 (all P<0.05; Fig. 6D). Treatment of U0126 led to a
significant decrease in p-ERK levels, whilst not notably affecting
the total ERK protein levels (all P<0.01; Fig. 6E). Hence, these results demonstrated
that USP19 regulates ccRCC proliferation and migration through its
regulation of ERK signaling.
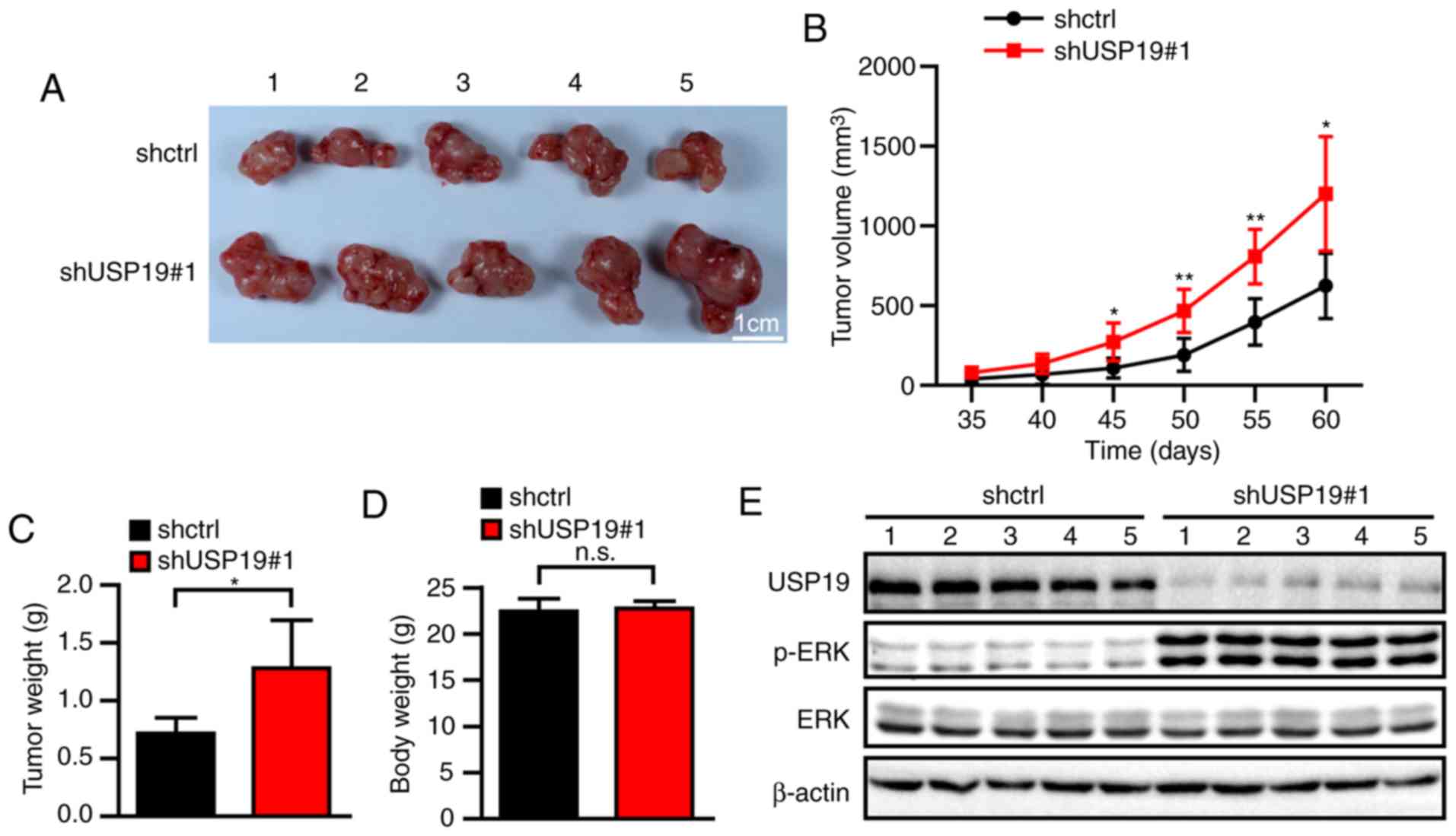
USP19 inhibits tumorigenesis in ccRCC
cells in vivo
To investigate the effect of USP19 in vivo,
nude mice were inoculated with shctrl- or shUSP19#1-infected Caki-1
cells. Compared with the tumors created by the shctrl cells, the
tumors derived from the shUSP19#1-infected Caki-1 cells were
significantly larger (all P<0.05; Fig. 7A and B). Consistently, the tumor
weights at 60 days were significantly heavier in the shUSP19#1
group than the shctrl group (P<0.05; Fig. 7C), whereas the body weights at 60
days of the mice in the two groups were not significantly different
(P>0.05; Fig. 7D) Then, western
blot experiments revealed notably increased p-ERK levels in the
tumor tissues derived from the shUSP19#1 group compared with those
derived from the shctrl group (Fig.
7E). These findings indicated that USP19 downregulation
promotes ccRCC tumor growth in vivo.
Discussion
The present study revealed that USP19 may function
as a tumor suppressor in ccRCC, and that USP19 expression is
significantly reduced in ccRCC tissues, based on multiple
downloaded datasets. Furthermore, the present study also
demonstrated that USP19 downregulation in ccRCC was associated with
more advanced tumors and predicted unfavorable prognostic outcomes
in a TCGA cohort.
CNV (20), DNA
methylation aberration (27) and
transcriptional dysregulation (28)
are all important events for dysregulation of gene expression
during various diseases. In cancer cells, in which somatic
mutations are easily induced, CNVs play an important role in the
somatic mutation-induced dysregulation of cancer genes (29–31).
In the present study, by analyzing CNV data from TCGA database, it
was revealed that the copy number of USP19 was notably reduced in
ccRCC tissues, suggesting that USP19 loss may be due to copy number
loss, which requires further investigation.
CCK-8 assays are conducted to assess cancer cell
proliferation abilities, and Transwell and wound healing assays are
used to evaluate migration abilities (32–35).
To investigate the observations from the bioinformatics analyses,
in vitro cell line experiments were conducted, indicated
that USP19 overexpression suppressed proliferation and migration,
whereas USP19 knockdown promoted proliferation and migration. At
the molecular level, USP19 overexpression decreased the expression
of the proliferation-promoting genes PCNA and cyclin D1, and
increased the expression of the proliferation-inhibiting gene p27;
conversely, USP19 overexpression decreased the expression of the
migration-associated genes MMP2 and MMP9. Conversely, depletion of
USP19 inhibited proliferation in DU145, PC-3 and 22RV1 prostate
cancer cells, indicating that USP19 has distinct functions in
different tumors (36).
Aberrant ERK-MAPK pathway signaling contributes to
cell proliferation and migration in various types of cancer
(37,38). In the present study, USP19
overexpression reduced p-ERK levels. Moreover, USP19 knockdown
promoted ERK phosphorylation, as well as promoting ccRCC
proliferation in vitro and in vivo. Therefore, these
results suggested that the molecular mechanism underlying
USP19-mediated proliferation and migration inhibition in ccRCC may
depend on inactivation of the ERK-MAPK signaling pathway. However,
one limitation of the present study is that the colocalization of
USP19 and p-ERK was not confirmed in mouse tumor tissues, due to
low quantities and poor specificity of the USP19 antibody.
Wu et al (39) previously reported that USP19
expression was lower in ccRCC tissue compared with in normal kidney
tissue by analyzing TCGA KIRC data. Liu et al (40) analyzed gene and isoform expression
signatures associated with tumor stage in KIRC and found
uc003cvz.3, the major isoform of USP19, was significantly
downregulated in patients with stage IV KIRC, whereas higher
uc003cvz.3 expression suggested improved survival rates. However,
these two studies only showed the clinical relationship of USP19
and ccRCC by analyzing databases; they did not investigate the
biological functions of USP19 in ccRCC cells. Conversely, the
present study demonstrated that USP19 overexpression inhibited
ccRCC cell proliferation and migration in vitro; conversely,
USP19 knockdown promoted proliferation and migration in
vitro, as well as promoting tumor growth in vivo. To the
best of our knowledge, this is the first study to investigate the
biological functions and molecular mechanisms of USP19 in ccRCC.
These findings expand current knowledge regarding USP19, and
suggest that USP19 may play an important role in the development
and progression of ccRCC.
Acknowledgments
Not applicable.
Funding
This work was supported by Hubei Health Research
Foundation (grant no. WJ2019F039).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZT and JZ designed the study. WH, YS, XF, XW and GZ
performed the experiments. WH, YS, CS, TD, TY and GW performed data
analysis. WH, YS, ZT and JZ contributed to manuscript revisions and
all the authors reviewed the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All protocols involving animals were approved by the
Institutional Animal Care and Use Committee of the Institute of
Model Animals of Wuhan University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar
|
|
2
|
Rini BI, Campbell SC and Escudier B: Renal
cell carcinoma. Lancet. 373:1119–1132. 2009. View Article : Google Scholar
|
|
3
|
Delahunt B, Bethwaite PB and Nacey JN:
Outcome prediction for renal cell carcinoma: Evaluation of
prognostic factors for tumors divided according to histological
subtype. Pathology. 39:459–465. 2007. View Article : Google Scholar
|
|
4
|
Lin YF, Chou JL, Chang JS, Chiu IJ, Chiu
HW and Lin YF: Dysregulation of the miR-25-IMPA2 axis promotes
metastatic progression in clear cell renal cell carcinoma.
EBioMedicine. 45:220–230. 2019. View Article : Google Scholar :
|
|
5
|
Afriansyah A, Hamid AR, Mochtar CA and
Umbas R: Targeted therapy for metastatic renal cell carcinoma. Acta
Med Indones. 48:335–347. 2016.
|
|
6
|
Husnjak K and Dikic I: Ubiquitin-binding
proteins: Decoders of ubiquitin-mediated cellular functions. Annu
Rev Biochem. 81:291–322. 2012. View Article : Google Scholar
|
|
7
|
Harada K, Kato M and Nakamura N:
USP19-mediated deubiquitination facilitates the stabilization of
HRD1 ubiquitin ligase. Int J Mol Sci. 17(pii): E18292016.
View Article : Google Scholar
|
|
8
|
Fraile JM, Quesada V, Rodríguez D, Freije
JM and López-Otín C: Deubiquitinases in cancer: New functions and
therapeutic options. Oncogene. 31:2373–2388. 2012. View Article : Google Scholar
|
|
9
|
Hu M, Li P, Li M, Li W, Yao T, Wu JW, Gu
W, Cohen RE and Shi Y: Crystal structure of a UBP-family
deubiquitinating enzyme in isolation and in complex with ubiquitin
aldehyde. Cell. 111:1041–1054. 2002. View Article : Google Scholar
|
|
10
|
Hassink GC, Zhao B, Sompallae R, Altun M,
Gastaldello S, Zinin NV, Masucci MG and Lindsten K: The ER-resident
ubiquitin-specific protease 19 participates in the UPR and rescues
ERAD substrates. EMBO Rep. 10:755–761. 2009. View Article : Google Scholar :
|
|
11
|
Jin S, Tian S, Chen Y, Zhang C, Xie W, Xia
X, Cui J and Wang RF: USP19 modulates autophagy and antiviral
immune responses by deubiquitinating Beclin-1. EMBO J. 35:866–880.
2016. View Article : Google Scholar :
|
|
12
|
Gierisch ME, Pedot G, Walser F,
Lopez-Garcia LA, Jaaks P, Niggli FK and Schäfer BW: USP19
deubiquitinates EWS-FLI1 to regulate Ewing sarcoma growth. Sci Rep.
9:9512019. View Article : Google Scholar :
|
|
13
|
Shahriyari L, Abdel-Rahman M and Cebulla
C: BAP1 expression is prognostic in breast and uveal melanoma but
not colon cancer and is highly positively correlated with RBM15B
and USP19. PLoS One. 14:e02115072019. View Article : Google Scholar :
|
|
14
|
Eikrem O, Beisland C, Hjelle K, Flatberg
A, Scherer A, Landolt L, Skogstrand T, Leh S, Beisvag V and Marti
HP: Transcriptome sequencing (RNAseq) enables utilization of
Formalin-fixed, paraffin-embedded biopsies with clear cell renal
cell carcinoma for exploration of disease biology and biomarker
development. PLoS One. 11:e01497432016. View Article : Google Scholar :
|
|
15
|
Yao X, Tan J, Lim KJ, Koh J, Ooi WF, Li Z,
Huang D, Xing M, Chan YS, Qu JZ, et al: VHL deficiency drives
enhancer activation of oncogenes in clear cell renal cell
carcinoma. Cancer Discov. 7:1284–1305. 2017. View Article : Google Scholar
|
|
16
|
R Core Team: R: A language and environment
for statistical computing. R Foundation for Statistical Computing;
Vienna: 2104
|
|
17
|
Love MI, Huber W and Anders S: Moderated
estimation of fold change and dispersion for RNA-seq data with
DESeq2. Genome Biol. 15:5502014. View Article : Google Scholar :
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals, 8th
edition. National Academies Press (US); Washington, DC: 2011
|
|
20
|
Macè A, Kutalik Z and Valsesia A: Copy
number variation. Methods Mol Biol. 1793:231–258. 2018. View Article : Google Scholar
|
|
21
|
Zińczuk J, Zaręba K, Guzińska-Ustymowicz
K1, Kędra B Kemona A and Pryczynicz A: Expression of chosen cell
cycle and proliferation markers in pancreatic intraepithelial
neoplasia. Prz Gastroenterol. 13:118–126. 2018.
|
|
22
|
Sarsik B, Doganavsargil B, Simsir A,
Yazici A, Pehlivanoglu B, Cal C and Sen S: P21 and p27
immunoexpression in upper urinary tract urothelial carcinomas.
Pathol Oncol Res. 22:839–845. 2016. View Article : Google Scholar
|
|
23
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar
|
|
24
|
Björklund M and Koivunen E:
Gelatinase-mediated migration and invasion of cancer cells. Biochim
Biophys Acta. 1755:37–69. 2005.
|
|
25
|
Wang D, Wang D, Wang N, Long Z and Ren X:
Long Non-coding RNA BANCR promotes endometrial cancer cell
proliferation and invasion by regulating MMP2 and MMP1 via ERK/MAPK
signaling pathway. Cell Physiol Biochem. 40:644–656. 2016.
View Article : Google Scholar
|
|
26
|
Tian XQ, Guo FF, Sun DF, Wang YC, Yang L,
Chen SL, Hong J and Fang JY: Downregulation of ZNF278 arrests the
cell cycle and decreases the proliferation of colorectal cancer
cells via inhibition of the ERK/MAPK pathway. Oncol Rep.
38:3685–3692. 2017.
|
|
27
|
He W, Ju D, Jie Z, Zhang A, Xing X and
Yang Q: Aberrant CpG-methylation affects genes expression
predicting survival in lung adenocarcinoma. Cancer Med.
7:5716–5726. 2018. View Article : Google Scholar :
|
|
28
|
Schuijers J, Manteiga JC, Weintraub AS,
Day DS, Zamudio AV, Hnisz D, Lee TI and Young RA: Transcriptional
dysregulation of MYC reveals common enhancer-docking mechanism.
Cell Rep. 23:349–360. 2018. View Article : Google Scholar :
|
|
29
|
Gu X, Coates PJ, Boldrup L, Wang L, Krejci
A, Hupp T, Fahraeus R, Norberg-Spaak L, Sgaramella N, Wilms T and
Nylander K: Copy number variation: A prognostic marker for young
patients with squamous cell carcinoma of the oral tongue. J Oral
Pathol Med. 48:24–30. 2019. View Article : Google Scholar
|
|
30
|
Wee Y, Wang T, Liu Y, Li X and Zhao M: A
pan-cancer study of copy number gain and up-regulation in human
oncogenes. Life Sci. 211:206–214. 2018. View Article : Google Scholar
|
|
31
|
Wang L, Zhao H, Xu Y, Li J, Deng C, Deng
Y, Bai J, Li X, Xiao Y and Zhang Y: Systematic identification of
lincRNA-based prognostic biomarkers by integrating lincRNA
expression and copy number variation in lung adenocarcinoma. Int J
Cancer. 144:1723–1734. 2019. View Article : Google Scholar
|
|
32
|
Duan X, Jiang B, Yang J, Zhou L, Tian B
and Mao X: FOXP3 inhibits MYC expression via regulating miR-198 and
influences cell viability, proliferation and cell apoptosis in
HepG2. Cancer Med. 7:6182–6192. 2018. View Article : Google Scholar :
|
|
33
|
He J, Yu L, Wang CM and Zhou XF: MiR-1275
promotes non-small cell lung cancer cell proliferation and
metastasis by regulating LZTS3 expression. Eur Rev Med Pharmacol
Sci. 22:2680–2687. 2018.
|
|
34
|
Chen Y, Zhang L, Liu L, Sun S, Zhao X,
Wang Y, Zhang Y, Du J and Gu L: Rasip1 is a RUNX1 target gene and
promotes migration of NSCLC cells. Cancer Manag Res. 10:4537–4552.
2018. View Article : Google Scholar :
|
|
35
|
Wang Y, Zheng C, Li T, Zhang R, Wang Y,
Zhang J, He Q, Sun Z and Wang X: Long noncoding RNA Z38 promotes
cell proliferation and metastasis and inhibits cell apoptosis in
human gastric cancer. Oncol Lett. 16:6051–6058. 2018.
|
|
36
|
Lu Y, Bedard N, Chevalier S and Wing SS:
Identification of distinctive patterns of USP19-mediated growth
regulation in normal and malignant cells. PLoS One. 6:e159362011.
View Article : Google Scholar :
|
|
37
|
Bao H, Guo CG, Qiu PC, Zhang XL, Dong Q
and Wang YK: Long non-coding RNA Igf2as controls hepatocellular
carcinoma progression through the ERK/MAPK signaling pathway. Oncol
Lett. 14:2831–2837. 2017. View Article : Google Scholar :
|
|
38
|
Li B, Liu YH, Sun AG, Huan LC, Li HD and
Liu DM: MiR-130b functions as a tumor promoter in glioma via
regulation of ERK/MAPK pathway. Eur Rev Med Pharmacol Sci.
21:2840–2846. 2017.
|
|
39
|
Wu M, Tu HQ, Chang Y, Tan B, Wang G, Zhou
J, Wang L, Mu R and Zhang WN: USP19 deubiquitinates HDAC1/2 to
regulate DNA damage repair and control chromosomal stability.
Oncotarget. 8:2197–2208. 2017.
|
|
40
|
Liu Q, Zhao S, Su PF and Yu S: Gene and
isoform expression signatures associated with tumor stage in kidney
renal clear cell carcinoma. BMC Syst Biol. 7 (Suppl 5):S72013.
View Article : Google Scholar
|