Introduction
Esophageal cancer is the seventh most common type of
cancer and the sixth leading cause of cancer-associated death
worldwide (1). In Asia, ~90% of
patients with esophageal cancer have esophageal squamous cell
carcinoma (ESCC) (2). The majority
of patients with ESCC are diagnosed with an advanced stage cancer
in the first instance, and therefore not eligible for surgery
(2). Chemotherapy and radiotherapy
are well-established treatments for patients with locally advanced
esophageal cancer. However, severe adverse events and complications
caused by chemotherapy and radiotherapy pose additional clinical
challenges (3,4). Brachytherapy via radioactive
iodine-125 (125I) seeds, a method of continuous
low-dose-rate irradiation, has been used to treat various types of
unresectable or locally recurrent cancer (5–7).
Compared with external beam radiotherapy (EBRT) using
single/fractionated high-dose-rate irradiation, 125I
seed brachytherapy has the advantages of easy preparation, direct
contact with tumor tissue, less radiation to adjacent normal
tissues and a higher relative biological effect (RBE) on tumor
cells (8,9). A series of studies have reported that
combination therapy of 125I seed brachytherapy and stent
placement shows good tumor control and survival benefits in
patients with advanced esophageal cancer (10–12).
However, to the best of our knowledge, there are no studies
examining the mechanisms underlying the anti-cancer effects of
125I seed radiation in ESCC.
Apoptosis is the classic type of programmed cell
death (PCD) and operates in a caspase-dependent manner. Following
DNA damage and G2/M cell cycle arrest, apoptosis is considered the
major mode of cell death induced by 125I seed radiation
as reported in studies on various types of cancer, such as
colorectal cancer (8), lung cancer
(13) and pancreatic cancer
(14). However, cancer cells
inherently have several anti-apoptotic mechanisms, resulting in
resistance to radiotherapy, and thus, cell survival (15). Other types of caspase-independent
PCD include autophagy and paraptosis, and these may also
participate in the anti-cancer mechanisms of 125I seed
radiation, but relevant studies are rare (16). Autophagy is an evolutionarily
conserved process, in which cells degrade damaged cytoplasmic
materials to facilitate survival, but excessive autophagy may also
lead to autophagic cell death (ACD) (17,18).
Previous studies have suggested that EBRT-induced autophagy serves
a protective role in ESCC (19,20);
however, the association between autophagy and 125I seed
radiation in ESCC has not yet been studied, to the best of our
knowledge. Paraptosis, or paraptosis-like cell death, is
characterized by cytoplasmic vacuolation resulting from
mitochondria and/or endoplasmic reticulum (ER) swelling, and it
lacks typical apoptotic morphological characteristics, including
chromatin condensation, nuclear fragmentation and apoptotic body
formation (21). Paraptosis differs
from necrosis in that, paraptosis does not result in early plasma
membrane rupture and the release of cellular contents (21,22).
Recent studies have shown that paraptosis-associated cell death is
an important anti-cancer mechanism of various natural products, and
is a novel area of interest in cancer research (23–26).
It has been reported that photodynamic therapy may also induce
paraptosis (27); however, aside
from one report on colorectal cancer (16), radiation has not hitherto been
associated with paraptosis.
In the present study, the modes and mechanisms of
cell death induced by 125I seed radiation in two human
ESCC cell lines, Eca-109 and KYSE-150, were systematically
investigated. 125I seed radiation kills KYSE-150 cells
primarily by inducing paraptosis, rather than apoptosis. Oxidative
stress, caused by an imbalance between the generation of reactive
oxygen species (ROS) and the ability of the cell to clear oxidants,
is critical for cellular proliferation and death (28). Herein, the roles of ROS in
apoptosis, autophagy and paraptosis induced by 125I seed
radiation in Eca-109 and KYSE-150 cells were further
investigated.
Materials and methods
Reagents and antibodies
Annexin V-enhanced green fluorescent protein
(EGFP)/propidium iodide (PI) apoptosis detection kit, cell cycle
detection kit, DAPI, bicinchoninic acid assay kit, and cell lysis
assay kit containing protease and phosphatase inhibitors were
purchased from Nanjing Keygen Biotech, Co., Ltd.
2,7-dichlorodihydrofluorescein diacetate (DCFH-DA) and trypan blue
dye were purchased from Beyotime Institute of Biotechnology. In
situ cell death detection TUNEL kit was purchased from Roche
Diagnostics GmbH. 3-Methyladenine (3-MA) and rapamycin were
purchased from Selleck Chemicals. N-Acetyl-L-cysteine (NAC) was
purchased from Sigma-Aldrich (Merck KGaA). Cycloheximide (CHX) was
purchased from MedChem Express. Rabbit monoclonal antibodies
against β-actin (cat. no. 4970), γ-H2AX (cat. no. 9718), caspase-3
(cat. no. 9662), cleaved caspase-3 (cat. no. 9664), LC3 (cat. no.
3868), CHOP (cat. no. 5554) and Ki-67 (cat. no. 9027) were obtained
from Cell Signaling Technology, Inc. Rabbit polyclonal antibodies
against p62 (cat. no. 18420) and Grp78/Bip (cat. no. 11587) were
obtained from ProteinTech Group, Inc. Horseradish peroxidase
(HRP)-conjugated goat anti-rabbit secondary antibody (cat. no.
G-21234) and Alexa Fluor 488-conjugated goat anti-rabbit secondary
antibody (cat. no. A-11008) were obtained from Invitrogen (Thermo
Fisher Scientific, Inc.).
125I seed irradiation
125I radioactive seeds (0.8 mCi, model
6711) were kindly provided by Shanghai Xinke Pharmaceutical, Co.,
Ltd. The 125I seed irradiation model used in the present
study was designed according to previous studies (29,30),
and was designed to provide a relatively homogeneous dose
distribution in vitro. This model consisted of a lower
irradiation plane and an upper treatment plane (the bottom of a
35-mm polystyrene dish). The height between the irradiation plane
and treatment plane was 6 mm. In the irradiation plane, 8 seeds
with the same activity were equidistantly spaced around a 35-mm
diameter circumference, and a ninth seed was confined to the
center. The initial dose rate in the treatment plane was 2.7 cGy/h,
and the exposure time for delivering cumulative radiation doses of
2, 4, 6 or 8 Gy were 74, 151, 230 and 312 h, respectively. For the
cells in the control group, sham seeds (without radioactivity) were
used.
Cell lines and culture
The human ESCC cell lines Eca-109 and KYSE-150 were
obtained from The Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences. The cells were maintained in RPMI 1640
medium (Hyclone; GE Healthcare), containing 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) and 1% penicillin/streptomycin (Hyclone;
GE Healthcare), in a 5% CO2 atmosphere at 37°C. In
certain experiments, the cells were incubated with 0.5 mM 3-MA, 2
µM CHX or 5 mM NAC.
Colony formation assay
After 2, 4, 6 or 8 Gy irradiation, cells were
cultured without radiation for 8–10 days to form colonies.
Subsequently, cells were fixed with 4% paraformaldehyde for 10 min
at room temperature and then stained with 0.1% crystal violet
solution for 20 min at room temperature. The planting efficiency
(PE) was defined as the ratio of colony number to inoculating cell
number, and the survival fraction (SF) was calculated as follows:
SF = PE (irradiated group)/PE (control group) ×100%. Cell survival
curves were fitted using a multi-target single-hit model:
SF=1-(1-eD/D0)N.
Cell viability assay
After 48 h of the indicated treatments, the cells
were suspended and stained with trypan blue dye for 3 min at room
temperature. Cell viability was determined using a trypan blue
exclusion assay and observed under a light microscope
(magnification, ×100; Eclipse TS100; Nikon Corporation).
Cell cycle and apoptosis assay
Cells were collected 48 h after irradiation. For
cell cycle analysis, cells were fixed with 70% ethanol overnight at
4°C and then stained with PI/ribonuclease A for 30 min at room
temperature in the dark. For apoptosis analysis, the cells were
stained with Annexin V-EGFP and PI for 20 min at room temperature
in the dark. Samples were analyzed by flow cytometry (NovoCyte
2060R, ACEA Biosciences, Inc.) using NovoExpress software version
1.3.0 (ACEA Biosciences, Inc.).
Intracellular ROS generation
levels
After 48 h of the indicated treatments, the cells
were incubated with 10 µM DCFH-DA for 30 min at 37°C in the dark.
Subsequently, the cells were washed with PBS, and then collected
for flow cytometry analysis.
Transmission electron microscopy
(TEM)
A total of 48 h after irradiation, the cells were
collected and fixed in 2.5% glutaraldehyde phosphate buffer
overnight at 4°C, washed with PBS, and then fixed in 1% osmium
tetroxide for 2 h at 4°C. Subsequently, the fixed cells were washed
with PBS, dehydrated in 50–100% ethanol, and then embedded in epoxy
resin. After polymerization for 48 h at 60°C, the ultrathin
sections were stained with uranyl for 15 min and lead acetate for
10 min both at room temperature, and examined under a transmission
electron microscope (H-7650; Hitachi, Ltd.).
Immunofluorescence microscopy
Cells were cultured on coverslips in 35-mm dishes.
Following irradiation and culturing for 48 h, cells were fixed with
ice-cold methanol for 5 min at −20°C and subsequently blocked in 5%
BSA for 1 h at room temperature. Fixed cells were incubated with
primary antibodies (LC3, 1:200; γ-H2AX, 1:200) overnight at 4°C,
washed with PBS, and then incubated with Alexa Fluor 488-conjugated
secondary antibodies (1:500) for 1 h at room temperature. Nuclei
were stained with DAPI for 10 min at room temperature and the cells
were examined under a fluorescence microscope (magnification, ×200;
BX53, Olympus Corporation).
Western blot analysis
Cells were lysed in the ice-cold lysis buffer,
containing freshly added protease and phosphatase inhibitors. Cell
lysates were centrifuged for 15 min at 16,000 × g at 4°C, and the
concentration of protein supernatants was determined using a
bicinchoninic acid assay. The protein supernatants were boiled in
loading buffer for 5 min, loaded on an SDS-gel. resolved by
SDS-PAGE, and transferred to PVDF membranes. The membranes were
blocked in 5% BSA in Tris-buffered saline solution containing 0.05%
Tween-20 (TBST) for 1 h at room temperature, incubated with the
indicated primary antibodies (β-actin, 1:1,000; γ-H2AX, 1:1,000;
caspase-3, 1:1,000; cleaved caspase-3, 1:1,000; LC3, 11,:000; p62,
1:2,000; Grp78/Bip, 1:4,000; CHOP, 1:1,000) overnight at 4°C,
washed with TBST, and then incubated with the HRP-conjugated
secondary antibody (1:3,000) for 1 h at room temperature.
Antibody-antigen complexes were detected using an enhanced
chemiluminescence kit (Tiangen Biotech, Co., Ltd.) and a light
capture system (Clinx Science Instruments, Co., Ltd.). Densitometry
analysis was performed using ImageJ version 1.51 (National
Institutes of Health). β-actin was used as a loading control.
Animal xenograft analysis
Eca-109 or KYSE-150 cells (5×106 in 0.1
ml PBS) were injected subcutaneously into the left axilla of female
BALB/c nude mice. Tumor volumes were measured every three days
using the following formula: Volume = length × width2 ×
0.5. When the average tumor volume reached 300 mm3
(after ~12 days for Eca-109 cells or 18 days for KYSE-150 cells),
the mice were randomized into two groups (n=6 per group): Control
group and the 125I seed group. For mice in the
125I seed group, an 18-gauge needle was punctured into
the center of the tumor mass, and then the 125I seed
(0.8 mCi) was implanted using a Mick applicator. For the control
group, only tumor puncture was performed. All mice were euthanized
15 days after treatment, and the cumulative dose in the
125I seed group was ~20 Gy. Tumor specimens were
harvested and weighed, and then processed for histological
analysis.
Tumor specimens were fixed in 4% paraformaldehyde
for 48 h at room temperature and embedded in paraffin. Tumor
sections (4 µm thickness) were subsequently used for hematoxylin
and eosin (H&E) staining, TUNEL assay, and immunohistochemistry
staining. TUNEL assay was performed according to the manufacturers
protocol. Immunohistochemistry was performed as described
previously (23) using primary
antibodies against Ki-67 (1:400; overnight at 4°C). HRP-conjugated
secondary antibody (1:1,000; 30 min at room temperature) and
diaminobenzidine were used for detection. Frozen tumor sections
were used for detection of ROS using DCFH-DA staining, as described
above. After incubation with DCFH-DA, tumor sections were counter
stained with DAPI for 10 min at room temperature.
All experiments involving mice were performed in
accordance with the guidelines on animal care and experiments on
laboratory animals of the Center of Experimental Animals, Southeast
University (Nanjing, China), and were approved by the Animal
Experimental Ethics Committee of Southeast University (approval no.
20190225004). Humane endpoints included mice exhibiting poor
physical conditions, confined activities affected by the tumor,
tumor sizes >2.0 cm in diameter (4.0 cm3), and severe
tumor ulceration, necrosis or infection.
Statistical analysis
All experiments were performed in triplicate. Data
are expressed as the mean ± standard deviation. A Students t-test
or one-way ANOVA followed by a post-hoc Tukeys test was used to
detect differences. P<0.05 was considered to indicate a
statistically significant difference. All analyses were performed
using SPSS version 23.0 (IBM, Corp.).
Results
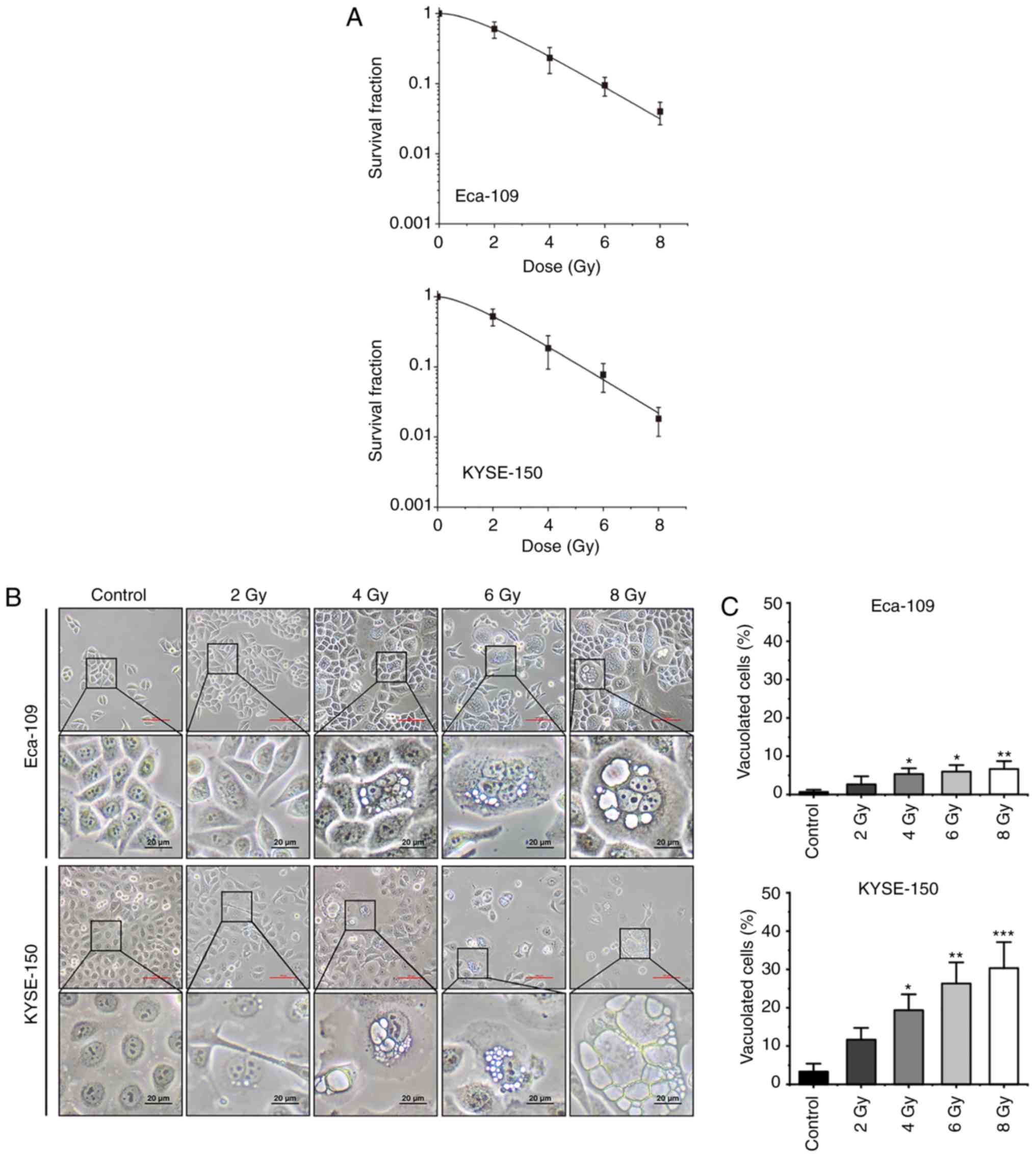
125I seed radiation reduces
proliferation and induces cytoplasmic vacuolation
To investigate the anti-cancer effects of
125I seed radiation, Eca-109 and KYSE-150 cells were
irradiated at different radiation doses (up to 8 Gy). As shown in
Fig. 1A, the effects on cell
proliferation were increased as the cumulative dose administered
was increased. The SF values of 8 Gy were 0.04±0.01 for Eca-109
cells and 0.02±0.01 for KYSE-150 cells (P=0.099). In Eca-109 cells,
the number of multinucleated giant cells (MNGCs) increased
following irradiation, and cytoplasmic vacuolation was observed.
The cytoplasmic vacuoles were primarily observed in MNGCs, and the
number as well as the size of the vacuoles increased over time
(Fig. 1B). In KYSE-150 cells,
extensive cytoplasmic vacuolation was observed following
irradiation, and the number of vacuolated cells increased in a
dose-dependent manner (Fig.
1C).
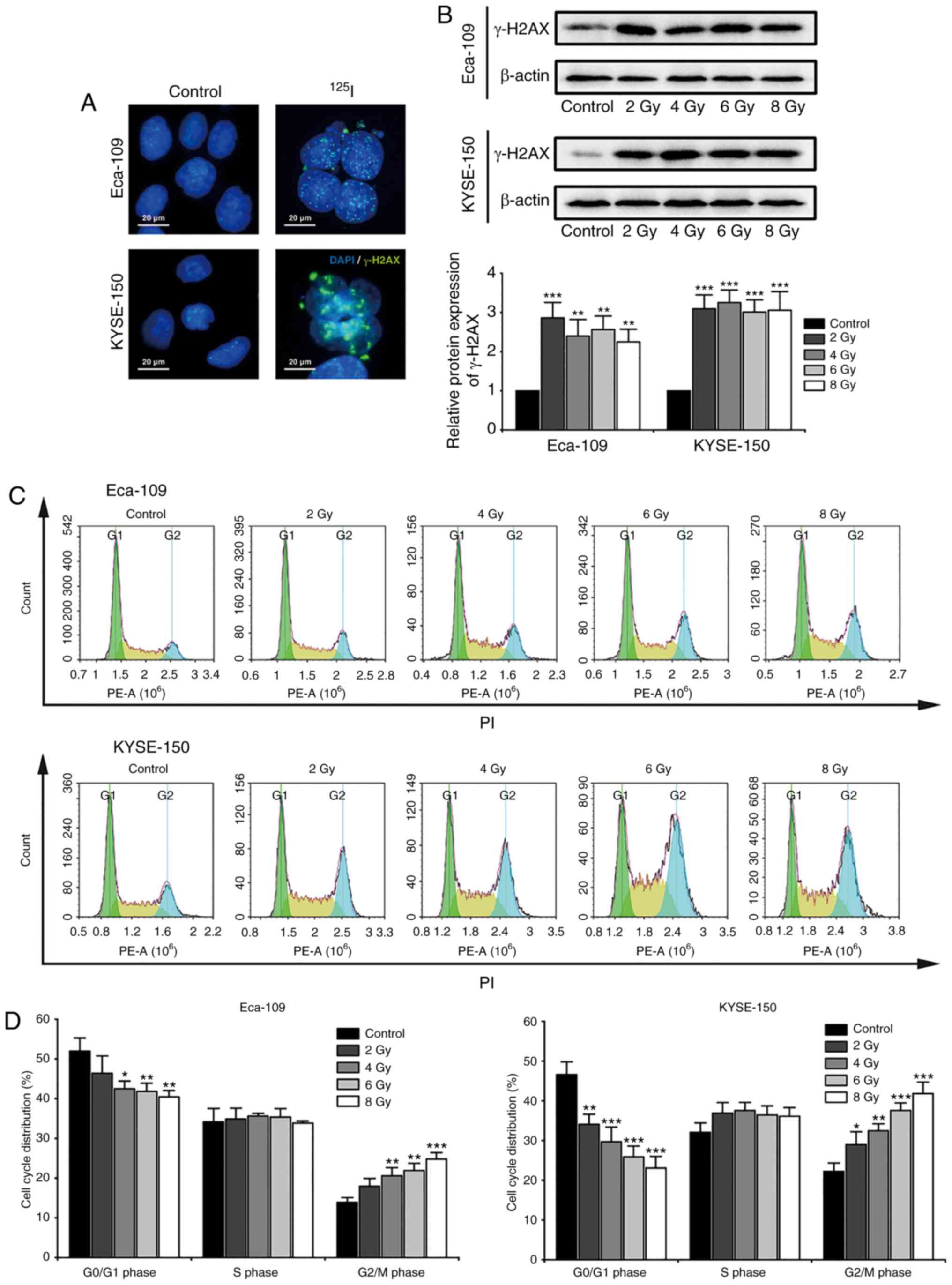
125I seed radiation induces
DNA damage and G2/M cell cycle arrest
Radiation-induced DNA damage serves an important
role in the anti-cancer effects of 125I seed radiation.
Histone H2AX localizes to the site of DNA double stranded breaks,
where it is phosphorylated to γ-H2AX. The phosphorylated γ-H2AX
further recruits DNA damage repair factors to repair the damaged
DNA double strands (31). For the
purpose of investigating cellular DNA damage and repair, nuclear
γ-H2AX foci and the protein expression levels of γ-H2AX were
observed 48 h after irradiation (Fig.
2A and B). The results suggested that 125I seed
radiation significantly induced DNA damage and repair in both
Eca-109 and KYSE-150 cells, even at a low cumulative dose of 2 Gy.
Progression of the cell cycle is delayed until the damaged DNA is
repaired (31), thus cell cycle
distribution was assessed (Fig.
2C). As shown in Fig. 2D,
125I seed radiation significantly increased the
proportion of cells in the G2/M phase and decreased the proportion
of cells in the G0/G1 phase, in a dose-dependent manner.
Furthermore, the degree of G2/M cell cycle arrest following
irradiation was higher in KYSE-150 cells compared with the Eca-109
cells.
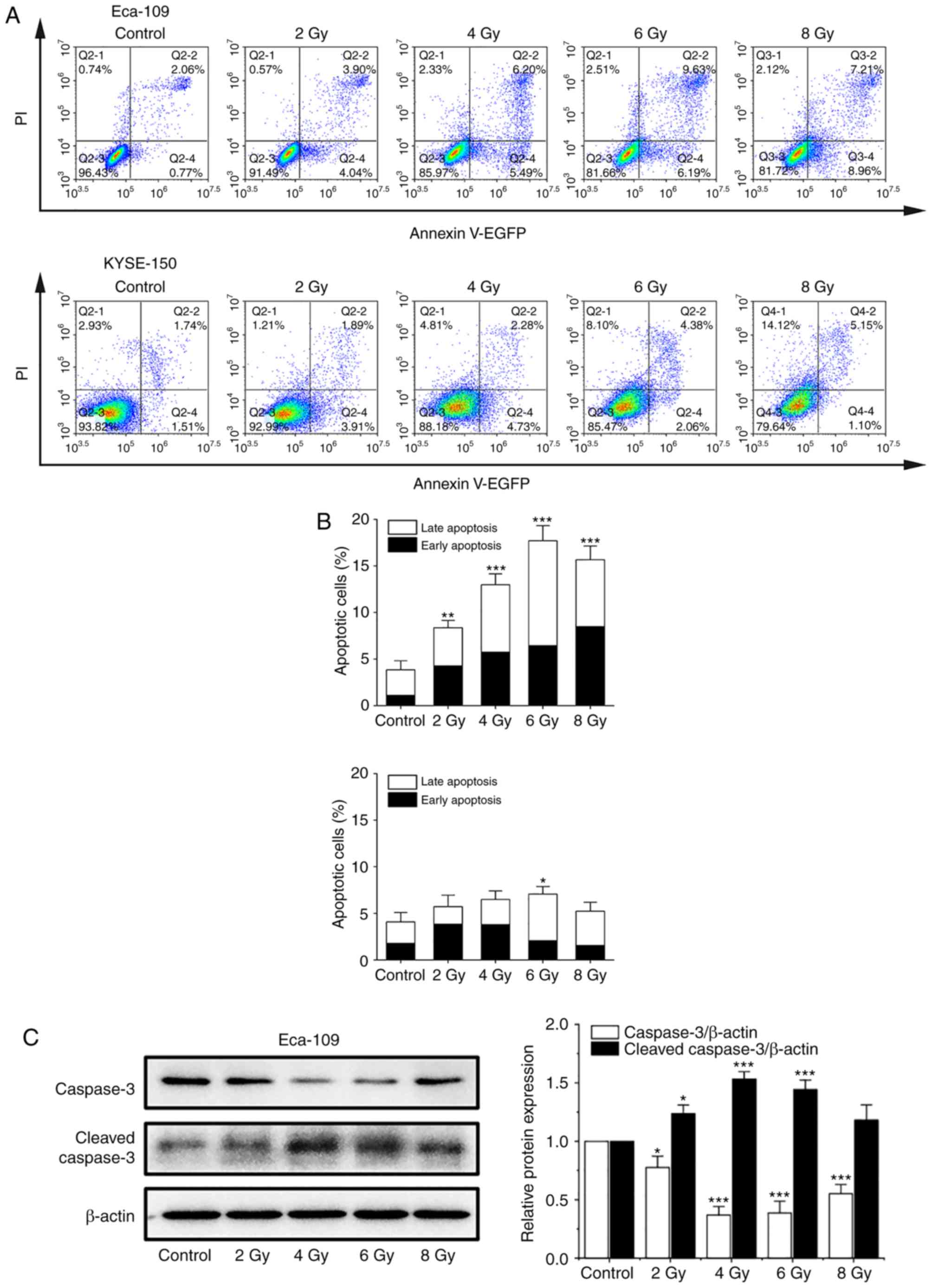
125I seed radiation induces
apoptosis and non-apoptotic cell death in KYSE-150 cells
In previous studies, apoptosis was considered the
major mode of cell death induced by 125I seed radiation
in various types of cancer cells (8,13,14).
An annexin V/PI assay was used to investigate apoptosis induced by
125I seed radiation in both cell lines. As shown in
Fig. 3A and B, the apoptotic rates
of Eca-109 and KYSE-150 cells increased as the cumulative dose
increased, peaking at 6 Gy and declining at 8 Gy. The apoptotic
rates of KYSE-150 cells were relatively low compared with the
Eca-109 cells. The maximum apoptotic rate was 7.07±0.81% for
KYSE-150 cells and 17.70±1.61% for Eca-109 cells (P=0.001).
However, the proportion of necrotic cells (EGFP-/PI+) in KESY-150
cells increased along with the increase in cumulative dose. These
results suggest that apoptosis is not the major mode of cell death
induced by 125I seed radiation in KESY-150 cells. To
assess caspase activation, the levels of caspase-3 and cleaved
caspase-3 were determined by western blot analysis. In Eca-109
cells, increase in cleavage of caspase-3 was consistent with the
results obtained from the Annexin V/PI assay (Fig. 3C). In the KYSE-150 cells, caspase-3
cleavage was observed with 2 Gy radiation, and the levels decreased
at higher doses (Fig. 3D). These
results suggest that cell death induced by 125I seed
radiation in Eca-109 cells was primarily through caspase-dependent
apoptosis, whereas in KYSE-150 cells, both apoptosis and
non-apoptotic (primary) modes were responsible.
125I seed radiation induces
protective autophagy in Eca-109 and KYSE-150 cells
Autophagy is characterized by the formation of
autophagosomes and autolysosomes, which manifest as cytoplasmic
vacuoles (27). In the present
study, extensive cytoplasmic vacuoles were observed following
irradiation, particularly in KYSE-150 cells (Fig. 1B and C). Therefore, whether
125I seed radiation induced autophagy in Eca-109 and
KYSE-150 cells was further investigated. During autophagy, LC3-I is
converted to LC3-II through lipidation, thus the levels of LC3, and
the ratio of LC3-II to LC3-I are frequently used as indicators of
autophagy (20,24,32). A
total of 48 h after 6 Gy irradiation, the cells were stained with
an anti-LC3 antibody and observed under a fluorescent microscope.
As shown in Fig. 4A, the number of
LC3 punctuations per cell increased notably following irradiation.
Subsequently, western blot analysis was performed to determine the
ratio of LC3-II to LC3-I in cells following exposure to 2, 4, 6 or
8 Gy radiation. The ratio of LC3-II to LC3-I in cells increased
significantly at 4 Gy in both cell lines, and then is decreased
slightly in Eca-109 cells at higher does, but maintained in
KYSE-150 cells (Fig. 4B).
Theoretically, the increased levels of LC3-II may have been the
result of augmented formation of autophagosomes or impaired
maturation of autophagosomes, which can block the degradation of
LC3-II (24). To determine
autophagic degradation activity, the protein levels of p62, a
ubiquitin-binding protein which is selectively degraded in
autolysosomes (18), was analyzed
by western blot analysis. The levels of p62 decreased in both cell
lines following irradiation (Fig.
4B), suggesting that 125I seed radiation induced
autophagic flux in Eca-109 and KYSE-150 cells.
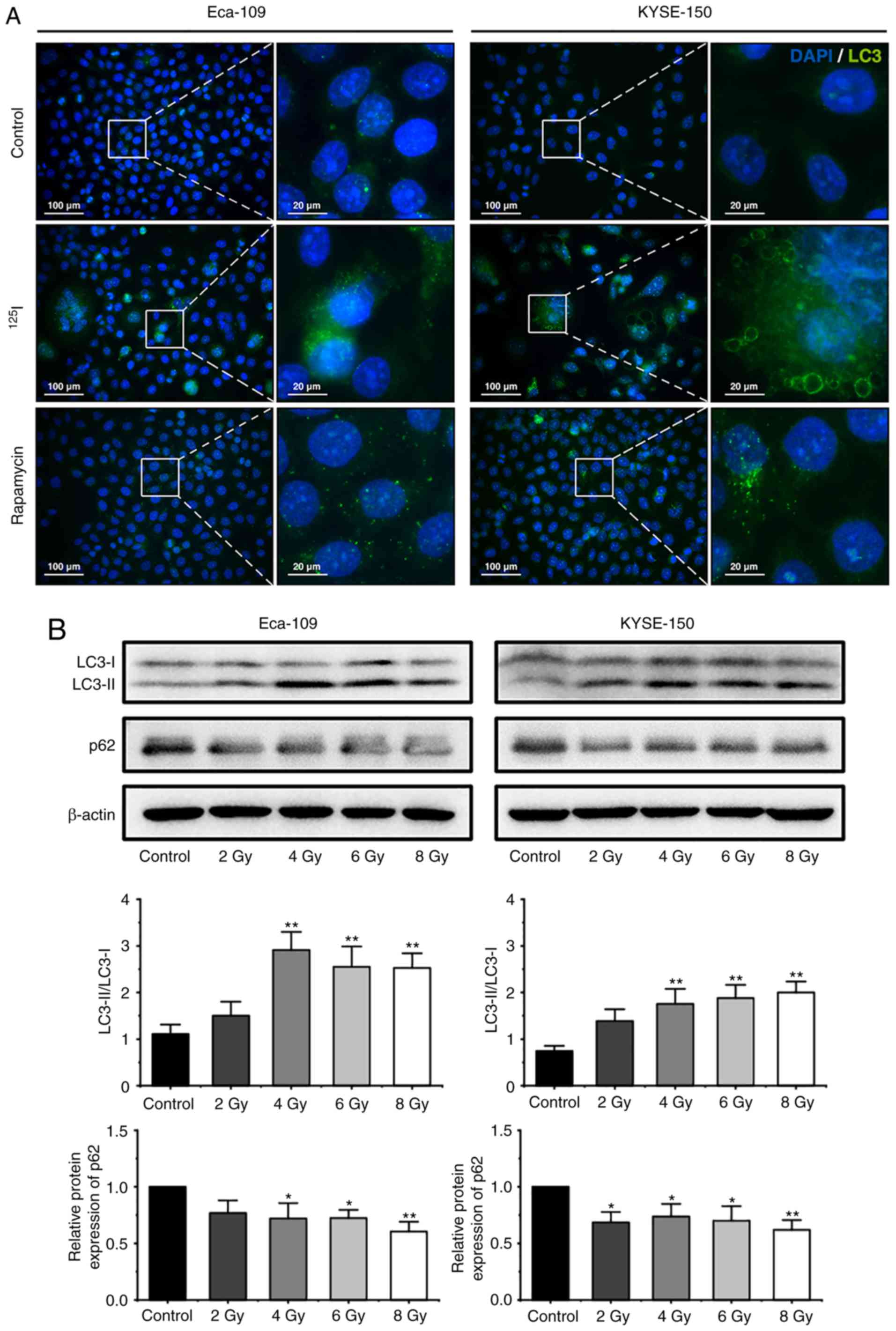 | Figure 4.125I seed radiation
induces protective autophagy in Eca-109 and KYSE-150 cells. (A)
Following 6 Gy irradiation, and 48 h of culture, LC3 punctuation
(green) was detected using a fluorescent microscope. Nuclei were
stained with DAPI (blue). Cells treated with 200 nM rapamycin
overnight were used as the positive control. (B) Cells were exposed
to 2, 4, 6 or 8 Gy radiation. The ratios of LC3-II to LC3-I
expression, and the relative protein expression of p62 were
analyzed by western blot. β-actin was used as a loading control.
Cells were pretreated with or without 3-MA, 2 h prior to 6 Gy
irradiation. After 48 h of the indicated treatment, (C) the ratios
of LC3-II to LC3-I were determined by western blot, and (D)
apoptosis was analyzed by Annexin V-EGFP/PI assay using flow
cytometry. (E) The percentage of cells with trypan blue exclusion,
and (F) the percentage of vacuolated cells were measured under a
light microscope. *P<0.05, **P<0.01, ***P<0.001 vs.
control group. 125I, Iodine-125; 3-MA, 3-methyladenine;
EGFP, enhanced green fluorescent protein; PI, propidium iodide. |
To further investigate the roles of autophagy in
cell death induced by 125I seed radiation, the effect of
3-MA, an autophagy inhibitor, were assessed. Given that the
apoptotic rates of both cell lines peaked at 6 Gy, the cells were
pretreated with 0.5 mM 3-MA for 2 h and then exposed to 6 Gy
radiation. Western blot analysis demonstrated that the ratios of
LC3-II to LC3-I decreased following 3-MA treatment in both cell
lines (Fig. 4C), suggesting that
3-MA attenuated 125I seed radiation-induced autophagy.
Annexin V/PI assay demonstrated that 3-MA improved 125I
seed radiation-induced apoptosis in both cell lines (Fig. 4D). Given the higher degree of
non-apoptotic cell death in KYSE-150 cells, cell viability was
assessed in these cells. As shown in Fig. 4E, 3-MA further decreased cell
viability of irradiated Eca-109 and KYSE-150 cells, but did not
affect the cell viability of unirradiated cells. These results
suggest that in Eca-109 and KYSE-150 cells, autophagy serves a
protective role in 125I seed radiation-induced cell
death.
3-MA did not reduce the percentage of vacuolated
cells in both irradiated and unirradiated cells (Fig. 4F). Additionally, immunofluorescence
images stained with an anti-LC3 antibody showed that certain
vacuoles did not co-localize with the LC3 punctuations (Fig. 5A), suggesting that these vacuoles
did not originate from autophagosome/autolysosomes. These findings
demonstrate that autophagy is not wholly responsible for
cytoplasmic vacuolation in either cell line following
125I seed irradiation.
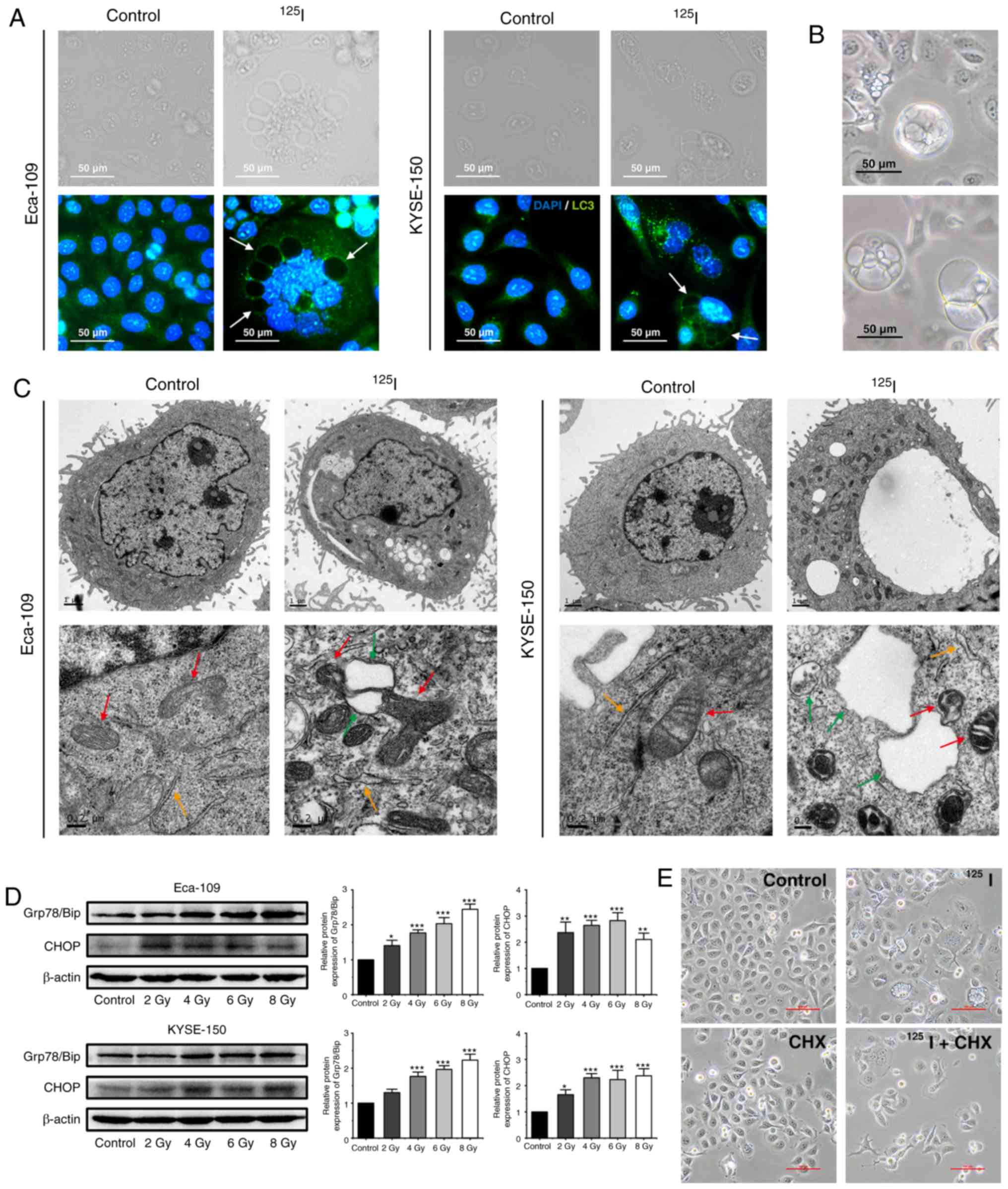 | Figure 5.125I seed radiation
induces paraptosis in Eca-109 and KYSE-150 cells. (A) Following
irradiation, LC3 punctuation (green) was detected under a
fluorescence microscope; the nuclei were stained with DAPI (blue).
Certain vacuolar membranes appeared negative for anti-LC3 antibody
(white arrow). (B) Cell morphology of detached KYSE-150 cells was
observed under the light microscope following 4 Gy irradiation. (C)
Ultrastructure of Eca-109 and KYSE-150 cells was observed using
transmission electron microscopy. Swollen mitochondria (red arrow),
endoplasmic reticulum (orange arrow) and vacuoles (green arrow)
were observed. (D) Levels of ER stress markers, Grp78/Bip and CHOP,
were determined by western blotting. β-actin was used as a loading
control. (E) KYSE-150 cells were pretreated with or without CHX, 2
h before 4 Gy irradiation. Morphological changes of KYSE-150 cells
were examined under a light microscope. Scale bar, 100 µm.
*P<0.05, **P<0.01, ***P<0.001 vs. control group.
125I, Iodine-125; CHX, cycloheximide. |
125I seed radiation induces
paraptosis and ER stress in KYSE-150 and Eca-109 cells
Paraptosis results in cytoplasmic vacuolation, and
the number and size of vacuoles increase over time during
paraptosis (21,26). In the irradiated KYSE-150 cells, the
cytoplasmic vacuoles enlarged in size gradually, the nuclei were
located peripherally, and subsequently, death-associated cellular
detachment occurred (Fig. 5B). To a
lesser degree, similar phenomena were also observed in irradiated
Eca-109 cells, particularly in MNGCs (Fig. 1B). Therefore, the ultrastructure of
mitochondria and ER by TEM were observed. Compared with the
unirradiated controls, irradiated cells exhibited swollen
mitochondria and ER, and also consequent vacuolation in both cell
lines (Fig. 5C). Given that ER
vacuolization is typically associated with persistent ER stress
(33), ER stress was assessed by
measuring the protein expression levels of the ER stress markers,
Grp78/Bip and CHOP. Elevated levels of Grp78/Bip and CHOP were
detected in both Eca-109 and KYSE-150 cells following irradiation,
and persistent elevation of CHOP was observed in KYSE-150 cells
(Fig. 5D). Studies have shown that
de novo protein synthesis is required for cytoplasmic
vacuolation in paraptosis, and CHX, a protein synthesis inhibitor,
inhibits paraptosis (21).
Therefore, KYSE-150 cells were pre-treated with CHX (2 µM) for 2 h
prior to 4 Gy irradiation. The results showed that CHX effectively
attenuated cytoplasmic vacuolation in irradiated cells (Fig. 5E). Taken together, these results
suggest that paraptosis is a key mechanism of cell death induced by
125I seed radiation in KYSE-150 cells, and paraptosis is
partially responsible for 125I seed radiation induced
cell death in Eca-109 cells.
125I seed radiation-induced
increases in ROS levels serve an important role in apoptosis,
autophagy and paraptosis
It has been reported that oxidative stress induced
by single high-dose radiation results in apoptosis and autophagy
(17). Thus, the effects of ROS on
cell death induced by 125I seed radiation were assessed.
Firstly, 48 h after 4 Gy irradiation, the cells were labeled with
the intracellular ROS probe, DCFH-DA, and analyzed by flow
cytometry. The results showed that 125I seed radiation
increased the levels of intracellular ROS in both Eca-109 and
KYSE-150 cells. KYSE-150 cells had higher basal levels of ROS
compared with Eca-109 cells (P<0.001). The increase in ROS
levels were more prominent in KYSE-150 cells compared with Eca-109
cells (fold change, 3.13±0.34 vs. 2.00±0.39, respectively, P=0.020;
Fig. 6A). Subsequently, cells were
pretreated with 5 mM NAC, an ROS scavenger, 4 h prior to 4 Gy
irradiation. The results showed that NAC reduced the accumulation
of intracellular ROS induced by 125I seed radiation in
both cell lines (Fig. 6B). Western
blot analysis demonstrated that NAC decreased the levels of the
autophagy indicator, the ratio of LC3-II to LC3-I, and ER stress
markers, Grp78/Bip and CHOP, in irradiated Eca-109 and KYSE-150
cells (Fig. 6C). Furthermore, as
shown in Fig. 6D, NAC attenuated
125I seed radiation-induced apoptosis in Eca-109 cells
(P=0.002), but did not significantly attenuate apoptosis in
KYSE-150 cells (P=0.695). As 125I seed radiation killed
KYSE-150 cells primarily through paraptosis, the changes in cell
viability and cytoplasmic vacuolation were assessed. The results
showed that NAC attenuated 125I seed radiation-induced
decreases in cell viability in both cell lines (Fig. 6E). Furthermore, for both irradiated
cell lines, the percentage of vacuolated cells decreased
significantly following NAC treatment (Fig. 6F). Taken together, these results
suggest that 125I seed radiation-induced increases in
ROS levels are critical for autophagy, apoptosis and paraptosis in
Eca-109 and KYSE-150 cells.
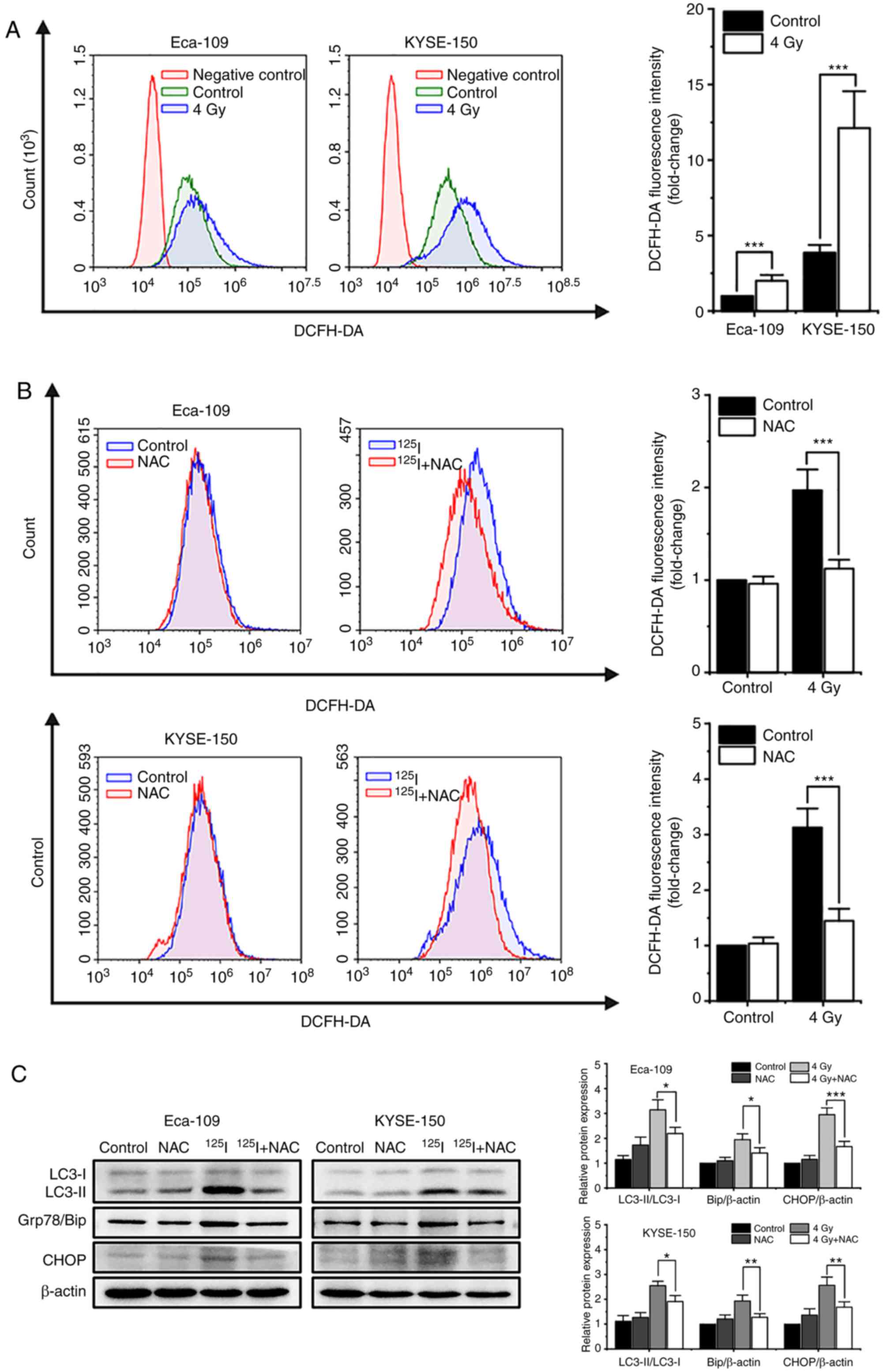 | Figure 6.125I seed
radiation-induced production of ROS is critical for apoptosis,
autophagy and paraptosis in Eca-109 and KYSE-150 cells. Cells were
pretreated with or without NAC 4 h prior to 4 Gy irradiation. (A
and B) Cells were labeled with DCFH-DA probe, the intracellular ROS
levels were quantitatively analyzed calculating the mean
fluorescence intensity using flow cytometry. Unlabeled cells were
used as the negative control. (C) Protein expression levels of
LC3-I, LC3-II, Grp78/Bip and CHOP, were determined by western blot.
β-actin was used as the loading control. (D) Apoptosis was detected
by Annexin V-EGFP/PI assay and quantitatively analyzed using flow
cytometry. (E) Percentage of cells with trypan blue excluded was
measured under a light microscope. (F) Morphological changes of
cells were observed, and the percentage of vacuolated cells was
measured under a light microscope. *P<0.05, **P<0.01,
***P<0.001 vs. control group. 125I, Iodine-125; ROS,
reactive oxygen species; NAC, N-acetyl-L-cysteine; DCFH-DA,
2,7-dichlorodihydrofluorescein diacetate; EGFP, enhanced green
fluorescent protein; PI, propidium iodide. |
125I seed radiation
inhibits ESCC xenograft growth in vivo
The in vivo effects of 125I seed
radiation using both Eca-109 and KYSE-150 human ESCC xenograft
mouse models were assessed. The 125I seed was implanted
into the tumor mass of each mouse, which received continuous
low-dose-rate irradiation for 15 days with a cumulative dose of ~20
Gy. Tumor growth was significantly reduced after 9 days of
125I seed irradiation (Fig.
7A), and the weights and volumes of harvested tumors were
significantly lower in the 125I seed group compared with
the control group (Fig. 7B and C).
To evaluate oxidative stress in vivo, the levels of ROS were
measured by staining the tissue with DCFH-DA. Increased levels of
ROS were observed in the 125I seed group (Fig. 7D). H&E staining showed cellular
vacuolation was present in tissue sections obtained from the
irradiated tumor mass (Fig. 7E).
Furthermore, 125I seed radiation resulted in apoptosis
(TUNEL assay, Fig. 7F) and reduced
proliferation (immunohistochemistry for Ki-67, Fig. 7G) in irradiated tumor tissues. The
results show that 125I seed radiation induced ROS
generation and DNA damage, initiated cell apoptosis and
vacuolation-related cell death (potentially via paraptosis), and
reduced cell proliferation and tumor growth in vivo.
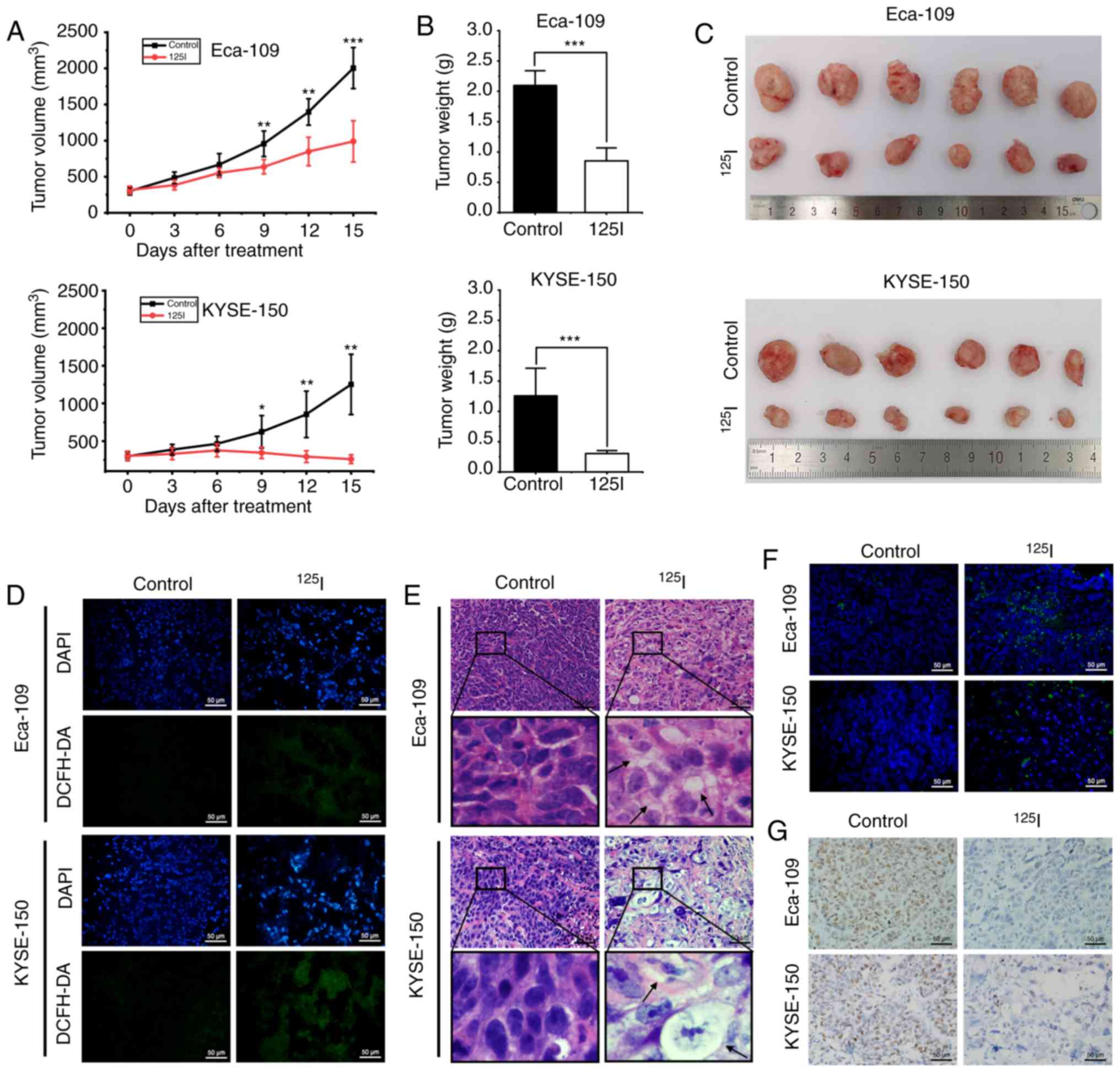 | Figure 7.125I seed radiation
inhibits growth of Eca-109 and KYSE-150 esophageal cancer
xenografts in mice. (A) Mice received tumor puncture (control
group) or 125I seed implantation (125I seed
group) when the average tumor volume reached 300 mm3.
Tumor volumes were measured every three days for 15 days after
treatment. (B) Weight of harvested tumors was measured 15 days
after treatment. (C) Representative images of the harvested tumors.
(D) Immunofluorescence detection of ROS using DCFH-DA staining
(green). Tissues were counterstained with DAPI (blue). Scale bar,
50 µm. (E) Representative hematoxylin and eosin staining of
harvested tumor specimens. Cytoplasmic vacuolation (black arrows)
was observed after 125I seed implantation. Scale bar, 50
µm. (F) Representative TUNEL staining of harvested tumor specimens.
Scale bar, 50 µm. (G) Representative immunohistochemistry staining
of the proliferation marker, Ki-67 in harvested tumor specimen.
Scale bar, 50 µm. *P<0.05, **P<0.01, ***P<0.001 vs.
control group. 125I, Iodine-125; ROS, reactive oxygen
species; DCFH-DA, 2,7-dichlorodihydrofluorescein diacetate. |
Discussion
125I seed brachytherapy was first used
for the treatment of prostate cancer in the 1970s (34). Because of its high efficacy and low
complication rate, 125I seed radiation was eventually
used for treatment of various unresectable and/or locally recurrent
types of cancer, including malignant glioma (5), non-small cell lung cancer (35) and esophageal cancer (11). Multiple studies demonstrated that,
compared with EBRT, 125I seed brachytherapy has greater
RBE values, suggesting that cancer cells are more sensitive to
125I seed radiation (8,9,36).
However, the mechanisms underlying the effects of 125I
seed radiation are not completely understood. In the present study,
the anti-cancer mechanisms of 125I seed radiation in
ESCC cells (Eca-109 and KYSE-150) were assessed. To the best of our
knowledge, this is the first report describing the roles of
apoptosis, autophagy and paraptosis in ESCC cells following
125I seed irradiation.
Previous studies have attributed the anti-cancer
effects of 125I seed radiation to DNA damage, G2/M cell
cycle arrest and apoptosis (8,14,37,38).
DNA damage can be induced directly by radiation or indirectly by
radiation-induced ROS (22). Cell
cycle checkpoints function to protect cells by delaying the
progression of the cell cycle until damaged DNA is repaired
(31). For cells with irreparable
DNA damage, the cell death pathway, primarily intrinsic
caspase-dependent apoptosis, is activated (22). In the present study, to reasonably
compare our data with previous studies, cells were cultured for 48
h following treatment (8,13,14).
125I seed radiation induced significant DNA damage and
cell cycle arrest at the G2/M phase in both cell lines; however,
the mode of cell death and the prominence of each mode differed in
each cell line. In Eca-109 cells, 125I seed radiation
resulted in noticeable caspase-dependent apoptosis, and the changes
in the tendency of apoptosis was almost consistent with the data
reported in studies on other types of cancer cells (8,14). In
KYSE-150 cells, 125I seed radiation initiated both
apoptotic and non-apoptotic cell death. As the level of apoptosis
was too low to exert a noticeable effect on cell death, it was
hypothesized that non-apoptotic modes were the primary mode of cell
death in KYSE-150 cells following 125I seed
irradiation.
Instead of the typical apoptotic morphology,
extensive cytoplasmic vacuolation was observed in KYSE-150 cells,
particularly at a cumulative dose of ≥4 Gy. Cytoplasmic vacuolation
is usually accompanied by autophagy and/or paraptosis. The vacuoles
associated with autophagy are derived from double-membraned
autophagosome and autolysosomes (27). The autophagosome sequesters damaged
cytoplasmic material, and then fuses with the lysosome to form an
autolysosome, where the cytoplasmic material is degraded. This
dynamic process is termed autophagic flux (18). In the present study, autophagic flux
was assessed by measuring the expression levels of the
autophagy-related proteins, LC3 and p62. The results supported the
notion 125I seed radiation initiated autophagic flux in
both Eca-109 and KYSE-150 cells. As excessive autophagy results in
ACD, it was determined whether the non-apoptotic cell death mode
observed in KYSE-150 cells was ACD. However, our data show that
autophagy inhibition increased cell apoptosis and reduced cell
viability in irradiated cells, indicating that 125I seed
radiation induces pro-survival autophagy in both cell lines. These
results are consistent with previous studies showing that EBRT
induces protective autophagy, and inhibition of autophagy by 3-MA
enhances the radiosensitivity of Eca-109 cells (19,32).
These data also suggest that autophagy inhibition may be a
potentially effective anti-cancer therapeutic strategy in
125I seed brachytherapy.
Autophagy inhibition by 3-MA did not reduce the
percentage of vacuolated cells, suggesting that paraptosis occurred
in the majority of the vacuolated cells. As a type of PCD,
paraptosis appears to differ from that of apoptosis, autophagy and
necrosis. Also, paraptosis is insensitive to apoptotic and
autophagic inhibitors (24).
Furthermore, paraptosis may be an effective cell-killing mode for
cells with impaired apoptotic mechanisms (39,40).
The molecular mechanism of paraptosis is not yet fully understood,
and there is hitherto no specific biochemical assay for paraptosis
(27,41). In the present study, paraptosis was
confirmed primarily through its characteristics (42): Extensive and intense cytoplasmic
vacuolation, fused from the swollen ER and/or mitochondria;
correlation with ER stress; and de novo protein synthesis.
The results of the present study suggest that 125I seed
radiation prominently initiates paraptosis in KYSE-150 cells, which
is the primary mode of cell death induced by 125I seed
radiation. 125I seed radiation mediated cell death is
different from that triggered by EBRT, which exhibits typical
apoptosis in KYSE-150 cells, and the cells are prone to form
apoptosis resistance following fractionated EBRT (43,44).
Characteristic morphological changes associated with paraptosis
were also observed in Eca-109 cells in vitro and in
vivo. In Eca-109 cells, paraptosis was primarily observed in
MNGCs, which is a morphological characteristic of mitotic
catastrophe (45). Although mitotic
catastrophe is regarded as a step preceding cell death (46), a large proportion of MNGCs still
remain viable and exhibit DNA synthesis following EBRT (45,47).
The results of the present study showed that, in Eca-109 cells,
paraptosis was at least partially responsible for cell death
induced by 125I seed radiation, and may be important for
cell death following mitotic catastrophe. In line with previous
studies (8,14), the results of the present study also
showed that caspase-dependent apoptosis decreased at a cumulative
dose of >6 Gy, suggesting that the remaining cancer cells had
improved apoptosis resistance, and for these cells, paraptosis may
have been the primary cell-killing mechanism. Hu et al
(16) previously demonstrated that
125I seed radiation triggers paraptosis in HCT116
colorectal cancer cells at a dose of 2 Gy. In the present study,
the characteristics of paraptosis were not notably present at 2 Gy,
and were more prominent at higher doses (≥4 Gy). To the best of our
knowledge, paraptosis has not been reported in studies on EBRT.
Based on the results of the present study, it may be hypothesized
that paraptosis may be associated with the mode of irradiation used
as well as the cell type, and this warrants further investigation.
The decrease in apoptosis observed at a dose of >6 Gy also
suggest that combination therapy of 125I seed
brachytherapy and other treatments such as chemotherapy may be
promising, but relevant basic and clinic studies are required.
It is well established that ionizing radiation can
induce ROS overproduction in cancer cells (17,48,49).
Compared with DNA damage directly induced by ionizing radiation,
ROS-induced damage of nucleic acids, proteins and lipids serve more
important roles in cell death (50). The majority of studies on
125I seed radiation have only focused on the
ROS-mediated DNA damage and consequent apoptosis (8,14,37),
whereas over-accumulation of ROS also interferes with various
organelles, including the mitochondria and ER, resulting in
mitochondrial dysfunction and ER stress response (50). Therefore, it was hypothesized that
125I seed radiation-induced autophagy and paraptosis are
downstream of 125I seed radiation-mediated ROS
generation. The results of the present study demonstrated that
125I seed radiation results in a significant increase in
ROS levels in both cell lines, and this was significantly reduced
by treatment with NAC, a ROS scavenger. Furthermore, NAC attenuated
125I seed radiation-induced ER stress, autophagy and
reduced cell viability in both cell lines. As the cell death modes
were different in the two cell lines, NAC noticeably attenuated
125I seed radiation-induced apoptosis in Eca-109 cells
and paraptotic changes in KYSE-150 cells. These results suggest
that 125I seed radiation-induced apoptosis, autophagy
and paraptosis are predominantly mediated by ROS in Eca-109 and
KYSE-150 cells.
Apoptosis resistance is an obstacle to successful
radiotherapy in patients with advanced ESCC, thus, it would be more
effective to treat cancer by initiating cell death through
different cell death modes. The results of the present study
demonstrated that, in the human ESCC cell lines Eca-109 and
KYSE-150, in addition to DNA damage, G2/M cell cycle arrest and
apoptosis, 125I seed radiation initiated paraptosis, a
relatively novel type of PCD. Additionally, 125I seed
radiation also resulted in the initiation of protective autophagy
to attenuate cell death, and 125I seed radiation-induced
apoptosis, paraptosis and autophagy were predominantly mediated by
ROS production in these two ESCC cell lines.
Acknowledgements
We would like to thank Dr Jian Yang, Dr Yue Chen
and Dr Ting-Ting Xu (Jiangsu Key Laboratory of Molecular and
Functional Imaging, Southeast University, Nanjing, China) for
providing technical assistance.
Funding
The present study was funded by the National
Natural Science Foundation of China (grant no. 81971716) and
Jiangsu Provincial Special Program of Social Development (grant no.
BE2016783).
Availability of data and material
The datasets used and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors contributions
JHG, CW and TKL designed the study CW, TKL, CHZ,
RF, YW performed the experiments. YW and GYZ analyzed the data.
JHG, CW and CHZ drafted the manuscript. All authors contributed to
reviewing and revising the manuscript and approved the final
version of the manuscript.
Ethics approval and consent to
participate
All experiments involving mice were performed in
accordance with the guidelines on animal care and experiments on
laboratory animals of the Center of Experimental Animals, Southeast
University (Nanjing, China), and were approved by the Animal
Experimental Ethics Committee of Southeast University (approval no.
20190225004).
Patient consent for publication
Not applicable
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar
|
|
2
|
Lagergren J, Smyth E, Cunningham D and
Lagergren P: Oesophageal cancer. Lancet. 390:2383–2396. 2017.
View Article : Google Scholar
|
|
3
|
Jatoi A, Martenson JA, Foster NR, McLeod
HL, Lair BS, Nichols F, Tschetter LK, Moore DF Jr, Fitch TR and
Alberts SR; North Central Cancer Treatment Group (N0044), :
Paclitaxel, carboplatin, 5-fluorouracil, and radiation for locally
advanced esophageal cancer: Phase II results of preliminary
pharmacologic and molecular efforts to mitigate toxicity and
predict outcomes: North central cancer treatment group (N0044). Am
J Clin Oncol. 30:507–513. 2007. View Article : Google Scholar
|
|
4
|
Tsushima T, Mizusawa J, Sudo K, Honma Y,
Kato K, Igaki H, Tsubosa Y, Shinoda M, Nakamura K, Fukuda H, et al:
Risk factors for esophageal fistula associated with
chemoradiotherapy for locally advanced unresectable esophageal
cancer: A supplementary analysis of JCOG0303. Medicine (Baltimore).
95:e36992016. View Article : Google Scholar :
|
|
5
|
Schnell O, Scholler K, Ruge M, Siefert A,
Tonn JC and Kreth FW: Surgical resection plus stereotactic 125I
brachytherapy in adult patients with eloquently located
supratentorial WHO grade II glioma-feasibility and outcome of a
combined local treatment concept. J Neurol. 255:1495–1502. 2008.
View Article : Google Scholar
|
|
6
|
Zhang F, Wang J, Guo J, Li Y, Huang X,
Guan Z, Lei G, Wang J, Ye X, Zhao X, et al: Chinese expert
consensus workshop report: Guideline for permanent iodine-125 seed
implantation of primary and metastatic lung tumors. Thorac Cancer.
10:388–394. 2019. View Article : Google Scholar
|
|
7
|
Mo Z, Zhang T, Zhang Y, Xiang Z, Yan H,
Zhong Z, Gao F and Zhang F: Feasibility and clinical value of
CT-guided (125)I brachytherapy for metastatic soft tissue sarcoma
after first-line chemotherapy failure. Eur Radiol. 28:1194–1203.
2018. View Article : Google Scholar
|
|
8
|
Zhuang HQ, Wang JJ, Liao AY, Wang JD and
Zhao Y: The biological effect of 125I seed continuous low dose rate
irradiation in CL187 cells. J Exp Clin Cancer Res. 28:122009.
View Article : Google Scholar :
|
|
9
|
Wang H, Li J, Qu A, Liu J, Zhao Y and Wang
J: The different biological effects of single, fractionated and
continuous low dose rate irradiation on CL187 colorectal cancer
cells. Radiat Oncol. 8:1962013. View Article : Google Scholar :
|
|
10
|
Guo JH, Teng GJ, Zhu GY, He SC, Fang W,
Deng G and Li GZ: Self-expandable esophageal stent loaded with 125I
seeds: Initial experience in patients with advanced esophageal
cancer. Radiology. 247:574–581. 2008. View Article : Google Scholar
|
|
11
|
Zhu HD, Guo JH, Mao AW, Lv WF, Ji JS, Wang
WH, Lv B, Yang RM, Wu W, Ni CF, et al: Conventional stents versus
stents loaded with (125)iodine seeds for the treatment of
unresectable oesophageal cancer: A multicentre, randomised phase 3
trial. Lancet Oncol. 15:612–619. 2014. View Article : Google Scholar
|
|
12
|
Chen HL, Shen WQ and Liu K: Radioactive
self-expanding stents for palliative management of unresectable
esophageal cancer: A systematic review and meta-analysis. Dis
Esophagus. 30:1–16. 2017. View Article : Google Scholar
|
|
13
|
Qu A, Wang H, Li J, Wang J, Liu J, Hou Y,
Huang L and Zhao Y: Biological effects of (125)i seeds radiation on
A549 lung cancer cells: G2/M arrest and enhanced cell death. Cancer
Invest. 32:209–217. 2014. View Article : Google Scholar
|
|
14
|
Wang ZM, Lu J, Zhang LY, Lin XZ, Chen KM,
Chen ZJ, Liu FJ, Yan FH, Teng GJ and Mao AW: Biological effects of
low-dose-rate irradiation of pancreatic carcinoma cells in vitro
using 125I seeds. World J Gastroenterol. 21:2336–2342. 2015.
View Article : Google Scholar :
|
|
15
|
Kim BM and Hong Y, Lee S, Liu P, Lim JH,
Lee YH, Lee TH, Chang KT and Hong Y: Therapeutic implications for
overcoming radiation resistance in cancer therapy. Int J Mol Sci.
16:26880–26913. 2015. View Article : Google Scholar :
|
|
16
|
Hu L, Wang H, Zhao Y and Wang J: (125)I
seeds radiation induces paraptosis-like cell death via PI3K/AKT
signaling pathway in HCT116 cells. Biomed Res Int.
2016:81454952016. View Article : Google Scholar :
|
|
17
|
Chaurasia M, Bhatt AN, Das A, Dwarakanath
BS and Sharma K: Radiation-induced autophagy: Mechanisms and
consequences. Free Radic Res. 50:273–290. 2016. View Article : Google Scholar
|
|
18
|
Leidal AM, Levine B and Debnath J:
Autophagy and the cell biology of age-related disease. Nat Cell
Biol. 20:1338–1348. 2018. View Article : Google Scholar
|
|
19
|
Lu C and Xie C: Radiation-induced
autophagy promotes esophageal squamous cell carcinoma cell survival
via the LKB1 pathway. Oncol Rep. 35:3559–3565. 2016. View Article : Google Scholar
|
|
20
|
Tao H, Qian P, Lu J, Guo Y, Zhu H and Wang
F: Autophagy inhibition enhances radiosensitivity of Eca109 cells
via the mitochondrial apoptosis pathway. Int J Oncol. 52:1853–1862.
2018.
|
|
21
|
Lee D, Kim IY, Saha S and Choi KS:
Paraptosis in the anti-cancer arsenal of natural products.
Pharmacol Ther. 162:120–133. 2016. View Article : Google Scholar
|
|
22
|
Prokhorova EA, Egorshina AY, Zhivotovsky B
and Kopeina GS: The DNA-damage response and nuclear events as
regulators of nonapoptotic forms of cell death. Oncogene. 39:1–16.
2020. View Article : Google Scholar
|
|
23
|
Chen X, Chen X, Zhang X, Wang L, Cao P,
Rajamanickam V, Wu C, Zhou H, Cai Y, Liang G and Wang Y:
Curcuminoid B63 induces ROS-mediated paraptosis-like cell death by
targeting TrxR1 in gastric cells. Redox Biol. 21:1010612019.
View Article : Google Scholar
|
|
24
|
Fontana F, Moretti RM, Raimondi M,
Marzagalli M, Beretta G, Procacci P, Sartori P, Montagnani Marelli
M and Limonta P: delta-Tocotrienol induces apoptosis, involving
endoplasmic reticulum stress and autophagy, and paraptosis in
prostate cancer cells. Cell Prolif. 52:e125762019. View Article : Google Scholar :
|
|
25
|
Seo MJ, Lee DM, Kim IY, Lee D, Choi MK,
Lee JY, Park SS, Jeong SY, Choi EK and Choi KS: Gambogic acid
triggers vacuolization-associated cell death in cancer cells via
disruption of thiol proteostasis. Cell Death Dis. 10:1872019.
View Article : Google Scholar :
|
|
26
|
Zhao H, Xu X, Lei S, Shao D, Jiang C, Shi
J, Zhang Y, Liu L, Lei S, Sun H and Huang Q: Iturin A-like
lipopeptides from Bacillus subtilis trigger apoptosis, paraptosis,
and autophagy in Caco-2 cells. J Cell Physiol. 234:6414–6427. 2019.
View Article : Google Scholar
|
|
27
|
Kessel D: Apoptosis, paraptosis and
autophagy: Death and survival pathways associated with photodynamic
therapy. Photochem Photobiol. 95:119–125. 2019. View Article : Google Scholar
|
|
28
|
Weinberg F and Chandel NS: Reactive oxygen
species-dependent signaling regulates cancer. Cell Mol Life Sci.
66:3663–3673. 2009. View Article : Google Scholar
|
|
29
|
Aird EG, Folkard M, Mayes CR, Bownes PJ,
Lawson JM and Joiner MC: A purpose-built iodine-125 irradiation
plaque for low dose rate low energy irradiation of cell lines in
vitro. Br J Radiol. 74:56–61. 2001. View Article : Google Scholar
|
|
30
|
Gan Z, Jing J, Zhu G, Qin Y, Teng G and
Guo J: Preventive effects of (1)(2)(5)I seeds on benign restenosis
following esophageal stent implantation in a dog model. Mol Med
Rep. 11:3382–3390. 2015. View Article : Google Scholar
|
|
31
|
Fernandez-Capetillo O, Chen HT, Celeste A,
Ward I, Romanienko PJ, Morales JC, Naka K, Xia Z, Camerini-Otero
RD, Motoyama N, et al: DNA damage-induced G2-M checkpoint
activation by histone H2AX and 53BP1. Nat Cell Biol. 4:993–997.
2002. View
Article : Google Scholar
|
|
32
|
Chen YS, Song HX, Lu Y, Li X, Chen T,
Zhang Y, Xue JX, Liu H, Kan B, Yang G and Fu T: Autophagy
inhibition contributes to radiation sensitization of esophageal
squamous carcinoma cells. Dis Esophagus. 24:437–443. 2011.
View Article : Google Scholar
|
|
33
|
Shubin AV, Demidyuk IV, Komissarov AA,
Rafieva LM and Kostrov SV: Cytoplasmic vacuolization in cell death
and survival. Oncotarget. 7:55863–55889. 2016. View Article : Google Scholar :
|
|
34
|
Zaorsky NG, Davis BJ, Nguyen PL, Showalter
TN, Hoskin PJ, Yoshioka Y, Morton GC and Horwitz EM: The evolution
of brachytherapy for prostate cancer. Nat Rev Urol. 14:415–439.
2017. View Article : Google Scholar
|
|
35
|
Wu C, Li B, Sun G, Peng C and Xiang D:
Efficacy and safety of iodine-125 brachytherapy combined with
chemotherapy in the treatment of advanced NSCLC in the elderly.
Onco Targets Ther. 11:6617–6624. 2018. View Article : Google Scholar :
|
|
36
|
Lehnert S, Reniers B and Verhaegen F:
Relative biologic effectiveness in terms of tumor response of 125I
implants compared with 60Co gamma rays. Int J Radiat Oncol Biol
Phys. 63:224–229. 2005. View Article : Google Scholar
|
|
37
|
Liu C, Wang L, Qiu H, Dong Q, Feng Y, Li
D, Li C and Fan C: Combined strategy of radioactive (125)I seeds
and salinomycin for enhanced glioma chemo-radiotherapy: Evidences
for ROS-mediated apoptosis and signaling crosstalk. Neurochem Res.
43:1317–1327. 2018. View Article : Google Scholar
|
|
38
|
Zhang WF, Jin WD, Li B, Wang MC, Li XG,
Mao WY and Luo KY: Effect of brachytherapy on NF-kB and VEGF in
gastric carcinoma xenografts. Oncol Rep. 32:635–640. 2014.
View Article : Google Scholar
|
|
39
|
Lee DM, Kim IY, Seo MJ, Kwon MR and Choi
KS: Nutlin-3 enhances the bortezomib sensitivity of p53-defective
cancer cells by inducing paraptosis. Exp Mol Med. 49:e3652017.
View Article : Google Scholar :
|
|
40
|
Zhou Y, Huang F, Yang Y, Wang P, Zhang Z,
Tang Y, Shen Y and Wang K: Paraptosis-inducing nanomedicine
overcomes cancer drug resistance for a potent cancer therapy.
Small. 14:2018.
|
|
41
|
Kim JY, Lee DM, Woo HG, Kim KD, Lee HJ,
Kwon YJ and Choi KS: RNAi screening-based identification of USP10
as a novel regulator of paraptosis. Sci Rep. 9:49092019. View Article : Google Scholar :
|
|
42
|
Sperandio S, de Belle I and Bredesen DE:
An alternative, nonapoptotic form of programmed cell death. Proc
Natl Acad Sci USA. 97:14376–14381. 2000. View Article : Google Scholar
|
|
43
|
Wang G, Liu L, Sharma S, Liu H, Yang W,
Sun X and Dong Q: Bmi-1 confers adaptive radioresistance to
KYSE-150R esophageal carcinoma cells. Biochem Biophys Res Commun.
425:309–314. 2012. View Article : Google Scholar
|
|
44
|
Zhang H, Luo H, Jiang Z, Yue J, Hou Q, Xie
R and Wu S: Fractionated irradiation-induced EMT-like phenotype
conferred radioresistance in esophageal squamous cell carcinoma. J
Radiat Res. 57:370–380. 2016. View Article : Google Scholar :
|
|
45
|
Mirzayans R, Andrais B, Scott A, Wang YW,
Kumar P and Murray D: Multinucleated giant cancer cells produced in
response to ionizing radiation retain viability and replicate their
genome. Int J Mol Sci. 18:E3602017. View Article : Google Scholar
|
|
46
|
Denisenko TV, Sorokina IV, Gogvadze V and
Zhivotovsky B: Mitotic catastrophe and cancer drug resistance: A
link that must to be broken. Drug Resist Updat. 24:1–12. 2016.
View Article : Google Scholar
|
|
47
|
Weihua Z, Lin Q, Ramoth AJ, Fan D and
Fidler IJ: Formation of solid tumors by a single multinucleated
cancer cell. Cancer. 117:4092–4099. 2011. View Article : Google Scholar :
|
|
48
|
Srinivas US, Tan BWQ, Vellayappan BA and
Jeyasekharan AD: ROS and the DNA damage response in cancer. Redox
Biol. 25:1010842019. View Article : Google Scholar
|
|
49
|
Zou Z, Chang H, Li H and Wang S: Induction
of reactive oxygen species: An emerging approach for cancer
therapy. Apoptosis. 22:1321–1335. 2017. View Article : Google Scholar
|
|
50
|
Tang JY, Ou-Yang F, Hou MF, Huang HW, Wang
HR, Li KT, Fayyaz S, Shu CW and Chang HW: Oxidative
stress-modulating drugs have preferential anticancer
effects-involving the regulation of apoptosis, DNA damage,
endoplasmic reticulum stress, autophagy, metabolism, and migration.
Semin Cancer Biol. 58:109–117. 2019. View Article : Google Scholar
|