Introduction
Prostate cancer is the second most commonly
diagnosed type of cancer in men worldwide after lung cancer. In
2018, prostate cancer accounted for 13.5% of cancer cases in men,
and was responsible for ~360,000 deaths worldwide (1). Androgen deprivation therapy and
chemotherapy, such as docetaxel, are currently the first-line
treatment options for patients with metastatic prostate cancer
(2,3). However, patients my develop resistance
to treatment, with ensuing cancer progression (4,5).
Therefore, understanding the complex signaling network involved in
the development of prostate cancer is important for enabling the
development of novel treatments for patients with prostate
cancer.
Long non-coding RNAs (lncRNAs) are <200
nucleotides in length (6). Previous
studies have demonstrated that lncRNAs may act as regulators of a
variety of physiological processes (7,8).
Mechanistically, lncRNAs control gene expression at the epigenetic,
transcriptional and post-transcriptional levels (9,10).
Previous studies have revealed that several lncRNAs can act as
competing endogenous RNAs (ceRNAs), directly sponging microRNAs
(miRNAs) via complementary sequence binding, and upregulating their
miRNA targets (11). It has been
demonstrated that lncRNAs play a pivotal role in prostate
carcinogenesis (12,13). LncRNA-LBCS was found to be
downregulated in castration-resistant prostate cancer and it
interacts with heterogeneous nuclear ribonucleoprotein K to
suppress castration resistance of prostate cancer (14). Brain cytoplasmic RNA 1 (BCYRN1) is
the shortest lncRNA, and it is 200 nucleotides in length. BCYRN1
was originally identified as a neuron-specific transcript, and has
been shown to be overexpressed in several types of cancer (15). High expression of BCYRN1 has been
found to be associated with poor prognosis of patients with colon
cancer and esophageal squamous cell carcinoma (16,17).
In addition, the expression of BCYRN1 is a potential biomarker for
breast cancer diagnosis, as abnormal expression of BCYRN1 is
detected in serum from patients with breast cancer (18). However, the role of BCYRN1 in
prostate cancer is currently unknown.
The histone deacetylase (HDAC) family is divided
into classes I, IIa/b and IV, with 11 members, and class III, with
7 sirtuins (19). HDAC11 is a
member of class IV HDACs (20). In
contrast to class I HDACs, the role of HDAC11 in cancer remains
elusive. Furthermore, depletion of HDAC11 inhibits metabolic
activity in colorectal, prostate, ovarian and breast cancer cells
(21). However, although abnormal
expression of HDAC11 has been observed in tumors (21), the underlying molecular mechanism is
unknown.
The aim of the present study was to investigate the
role of BCYRN1 in prostate cancer progression and the involvement
of HDAC11 in this process.
Materials and methods
Clinical samples
A total of 72 pairs of prostate cancer and adjacent
healthy prostate tissues were collected from patients with prostate
cancer (aged 44–76 years), who underwent surgery at China-Japan
Union Hospital between June 2014 and July 2018. None of the
participants had received chemotherapy or radiotherapy prior to
surgery. Written informed consent was obtained from all patients.
The study was approved by and performed under the supervision of
the Institutional Ethics Review Board of China-Japan Union
Hospital.
Cell lines
The immortal prostate epithelial cell line RWPE-1,
and the prostate cancer cell lines LNCaP (androgen-dependent
prostate cancer), PC-3 (androgen-resistant prostate cancer) and
DU145 (androgen-resistant prostate cancer) were purchased from the
American Type Culture Collection. The cells were cultured in
RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and maintained in a humid incubator with 5% CO2 at
37°C.
Cell transfection
Small interfering RNAs (siRNAs) targeting BCYRN1
(5′-CGCCTGTAATCCCAGCTCTCA-3′) and control siRNA
(5′-GGATACGGAGTACTATAGC-3′) were synthesized and purchased from
GenePharma. To knock down BCYRN1, BCYRN1 siRNA was transfected into
PC-3 and DU145 cells using Lipofectamine RNAiMax (Invitrogen;
Thermo Fisher Scientific, Inc.) following the manufacturer's
protocol. The full length of HDAC11 was PCR-amplified from RWPE-1
cDNA, ligated into pcDNA3.1 and named pcDNA3.1-HDAC11. For
overexpression of HDAC11, pcDNA3.1-HDAC11 was transfected into PC-3
and DU145 cells using Lipofectamine 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
miR-939-3p mimic (5′-CCCUGGGCCUCUGCUCCCCAG-3′), miR-939-3p
inhibitor (5′-CUGGGGAGCAGAGGCCCAGGG-3′), miR-NC mimic
(5′-AGGAUGUAUUACCAGUGAUCGG-3′) and miR-NC inhibitor
(5′-UCGAGACACGUACGCAGAA-3′) were purchased from RiboBio, Inc.
miR-939-3p mimic, miR-939-3p inhibitor and their corresponding
negative controls were transfected into PC-3 and DU145 cells using
Lipofectamine 3000.
Protein extraction and western
blotting
Protein lysates were prepared with RIPA lysis buffer
(Beyotime Institute of Biotechnology). The protein concentration in
the lysates was determined with a bicinchoninic acid protein assay
kit (Pierce; Thermo Fisher Scientific, Inc.). Western blotting was
performed using a standard procedure. In total, 20 µg proteins were
loaded in each lane of an 8% SDS gel and were separated by
electrophoresis. Subsequently, the proteins were transferred to a
PVDF membrane (Merck KGaA) and blocked in 5% non-fat milk for 1 h
at room temperature. The membrane was then incubated with primary
antibody, followed by secondary antibody for 1 h at room
temperature. The blots were developed with ECL Western Blotting
Substrate (Pierce; Thermo Fisher Scientific, Inc.). For detection
of H3, H4, H3K9ac and H4K16ac, the lysates were run on a 15% gel.
Antibodies against H3 (cat. no. 61799, 1:5,000), H4 (cat. no.
61199, 1:5,000), H3K9ac (cat. no. 39917, 1:2,000) and H4K16ac (cat.
no. 61529, 1:2,000) were purchased from Active Motif. Antibodies
against HDAC11 (cat. no. 58442, 1:1,000) and β-actin (cat. no.
3700, 1:5,000) were purchased from Cell Signaling Technology, Inc.
HRP-conjugated secondary antibodies against rabbit (cat. no.
ab150077, 1:10,000) and mouse (cat. no. ab205719, 1:10,000) were
obtained from Abcam.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR) analysis
RNA was extracted from cells and specimens using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. The concentration and
quality of RNA were measured using NanoDrop 2000 (Thermo Fisher
Scientific, Inc.). RNA was reverse-transcribed into first-strand
cDNA with a PrimeScript RT reagent kit (Takara Bio, Inc.) at 37°C
for 15 min and at 85°C for 5 sec. qPCR was performed with SYBR
Premix Ex Taq (Takara Bio, Inc.) with the following conditions:
Initial denaturation at 95°C for 5 sec, followed by 35 cycles at
94°C for 15 sec and 55°C for 30 sec. β-actin and U6 were used as
internal controls for mRNA and lncRNA, and miRNA, respectively. The
relative expression level was calculated by the 2−ΔΔCq
method (22). The primer sequences
were as follows: BCYRN1, forward 5′-CTGGGCAATATAGCGAGAC-3′ and
reverse 5′-TGCTTTGAGGGAAGTTACG-3′; stem-loop primer,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCTGGGAGCUCCCCAG-3′;
miR-939-3p, forward 5′-GCCGAGCCCTGGGCCTCTGC-3′ and reverse
5′-CTCAACTGGTGTCGTGGA-3′; U6, forward 5′-ATTGGAACGATACAGAGAAGATT-3′
and reverse 5′-GGAACGCTTCACGAATTTG-3′; HDAC11, forward
5′-ACCCAGACAGGAGGAACCATA-3′ and reverse 5′-TGATGTCCGCATAGGCACAG-3′;
cyclin-dependent kinase inhibitor 1A (CDKN1A), forward
5′-TGTCCGTCAGAACCCATGC-3′ and reverse 5′-AAAGTCGAAGTTCCATCGCTC-3′;
optineurin (OPTN), forward 5′-CCAAACCTGGACACGTTTACC-3′ and reverse
5′-CCTCAAATCTCCCTTTCATGGC-3′; glucose transporter 1 (GLUT1),
forward 5′-GGCCAAGAGTGTGCTAAAGAA-3′ and reverse
5′-ACAGCGTTGATGCCAGACAG-3′; and β-actin, forward
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse
5′-CTCCTTAATGTCACGCACGAT-3′.
Cell proliferation assay
The Cell Counting Kit-8 assay (CCK-8 kit; DoJinDo
Molecular Technologies, Inc.) was used to detect cell proliferation
ability. Briefly, 2,000 cells were seeded in each well of a 96-well
plate. Subsequently, 10 µl CCK-8 solution was added into the
culture medium in the wells at 0, 1, 2, 3, 4 and 5 days after
transfection of siRNA and/or plasmid. After incubation for 2 h at
37°C, the medium containing CCK-8 solution was transferred to a new
96-well plate, and the absorbance at 450 nm was measured by a
microplate reader (Bio-Rad Laboratories, Inc.) to reflect cell
proliferation ability.
Flow cytometry analysis
Cell apoptosis was determined by flow cytometry
analysis with an Annexin V-FITC-PI Apoptosis Detection kit
(Invitrogen; Thermo Fisher Scientific, Inc.). Briefly, cells were
harvested and suspended in Annexin V Binding Buffer. Annexin V-FITC
and PI staining solution were added to the cells and incubated at
room temperature for 10 min. Subsequently, the cells were subjected
to flow cytometry analysis on a MACSQuant system (Miltenyi Biotec).
Data were analyzed using FlowJo software v10.6.1 (Becton, Dickinson
and Company).
Glucose uptake and lactate production
assay
A glucose uptake fluorometric assay kit (BioVision
Research, Inc.) was applied to detect glucose uptake in PC-3 and
DU145 cells. Cells were seeded at a density of 2,000/well in
96-well plates. Subsequently, the cells were starved overnight.
Next, 1 µM insulin was added, followed by the addition of 1 mM
2-deoxy-D-glucose (2-DG). A mixture containing Picoprobe was added
to the medium. The optical density (OD) value was then determined
by a microplate reader (Bio-Rad Laboratories, Inc.) to reflect 2-DG
uptake. Lactate production was detected using a lactic acid assay
kit (Sigma Aldrich; Merck KGaA). The OD value of treated cells was
applied to the standard curve to calculate lactate production
levels.
Determination of HDAC activity
The activity of HDACs in PC-3 and DU145 cells was
detected with a colorimetric HDAC activity assay kit (BioVision
Research, Inc.), following the manufacturer's instructions. The
relative OD value at 405 nm was analyzed with an iMark Microplate
Absorbance Reader (Bio-Rad Laboratories, Inc.) to reflect the
activity of HDACs.
Dual-luciferase reporter assay
The full length of the 3′ untranslated region (UTR)
of BCYRN1 and HDAC11 was amplified from RWPE-1 cDNA, and ligated
into pGL3-basic plasmids. Mutations were introduced into BCYRN1 and
HDAC11 3′UTR with a Quick Site-directed Mutation kit (Agilent
Technology; Thermo Fisher Scientific, Inc.). Plasmids and
miR-939-3p mimic were co-transfected into cells using Lipofectamine
3000 (Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. After 2 days, the relative luciferase
activity of each well was measured with a Dual Luciferase Reporter
Assay system (Promega Corporation). Firefly luciferase activity was
normalized to Renilla luciferase activity.
Statistical analysis
All data were analyzed using GraphPad Prism 6
(GraphPad Software, Inc.), and are presented as the mean ± standard
deviation. The correlation between BCYRN1 and HDAC11 mRNA
expression levels was examined by Pearson's correlation analysis.
Comparison of two groups was conducted with a Student's t-test,
while three or more groups were compared with one-way ANOVA
followed by Newman-Keuls analysis. The association between BCYRN1
expression and the characteristics of patients with prostate cancer
was analyzed with the χ2 test. All P-values were
two-tailed and P<0.05 was considered to indicate a statistically
significant difference.
Results
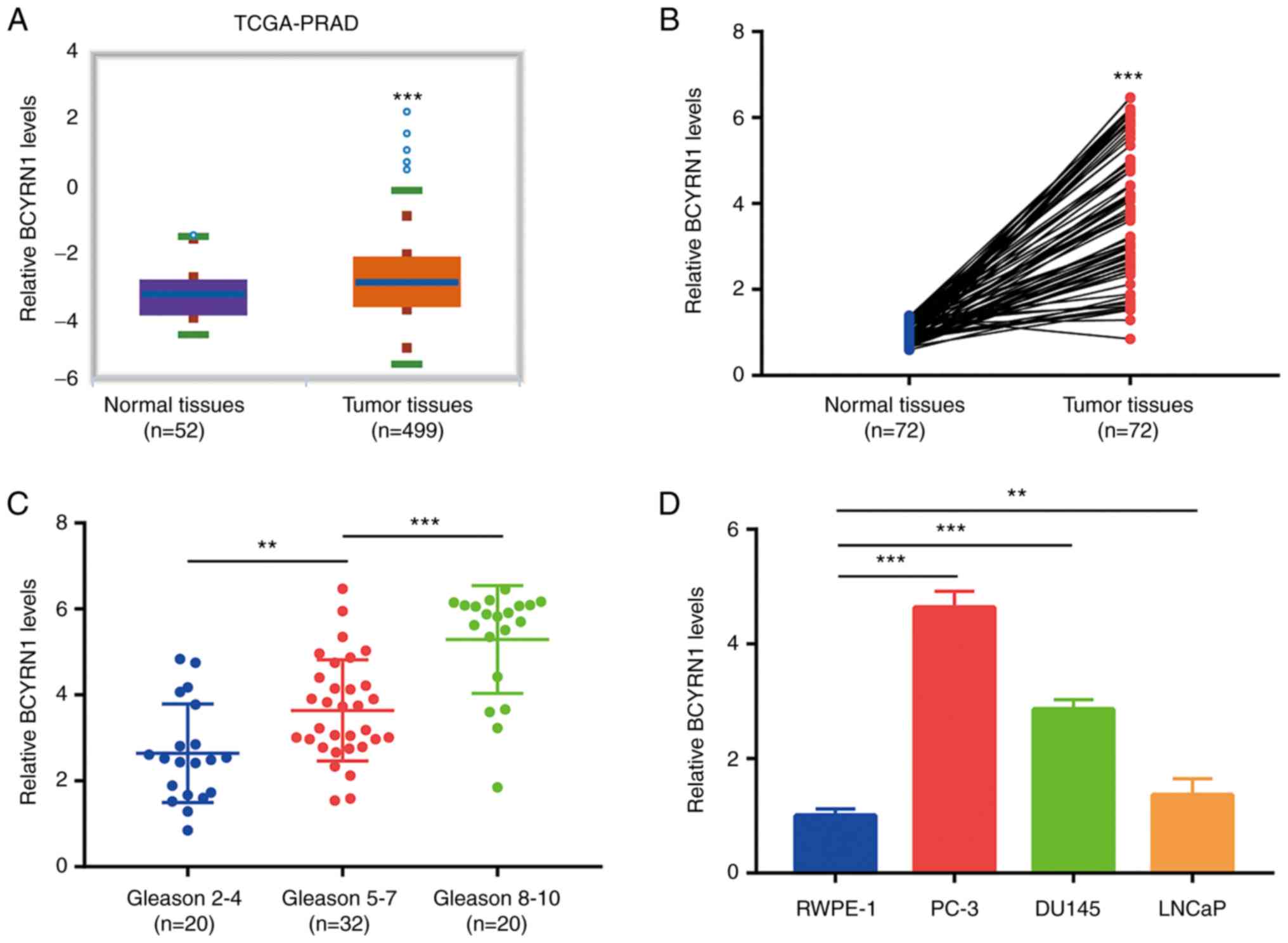
BCYRN1 is upregulated in prostate
cancer
Bioinformatics analysis of The Cancer Genome
Atlas-Prostate Adenocarcinoma (TCGA-PRAD) dataset (52 healthy
prostate tissues and 499 prostate tumors) demonstrated that the
expression of BCYRN1 was significantly upregulated in prostate
tumors (Fig. 1A). To verify this
result, BCYRN1 expression was detected in 72 pairs of prostate
tumors and matched healthy tissues. The RT-qPCR results indicated
that BCYRN1 was overexpressed in prostate tumors (Fig. 1B). Moreover, it was observed that
BCYRN1 was significantly upregulated in prostate tumors with a high
Gleason score (8–10) compared with those with an
intermediate Gleason score (5–7).
Furthermore, tumors with a low Gleason score (2–4)
exhibited a lower expression of BCYRN1 compared with those with an
intermediate Gleason score (5–7)
(Fig. 1C). The expression of BCYRN1
was not found to be associated with age, serum prostate-specific
antigen concentration or stage in patients with prostate cancer
(Table I). Therefore, these results
suggested that high expression of BCYRN1 is associated with
prostate cancer progression. The expression of BCYRN1 in
androgen-dependent prostate cancer cell lines (PC-3 and DU145), an
androgen-sensitive prostate cancer cell line (LNCaP) and an
immortal prostate epithelial cell line (RWPE-1) was also examined.
BCYRN1 was found to be overexpressed in prostate cancer cells,
particularly the androgen-resistant prostate cancer cell lines PC-3
and DU145, compared with RWPE-1 cells (Fig. 1D).
 | Table I.Association between BCYRN1 levels and
the characteristics of patients with prostate cancer. |
Table I.
Association between BCYRN1 levels and
the characteristics of patients with prostate cancer.
|
|
| BCYRN1
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Total | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.809 |
|
≤55 | 28 | 13 | 15 |
|
|
>55 | 44 | 23 | 21 |
|
| Stage |
|
|
| 0.341 |
|
I–II | 31 | 13 | 18 |
|
|
III–IV | 41 | 23 | 18 |
|
| Gleason score |
|
|
| 0.004 |
| ≤4 | 20 | 4 | 16 |
|
|
5-7 | 32 | 18 | 14 |
|
|
8-10 | 20 | 14 | 6 |
|
| Serum PSA
(ng/ml) |
|
|
| 0.406 |
| ≤4 | 15 | 6 | 9 |
|
| >4
and ≤10 | 34 | 16 | 18 |
|
|
>10 | 23 | 14 | 9 |
|
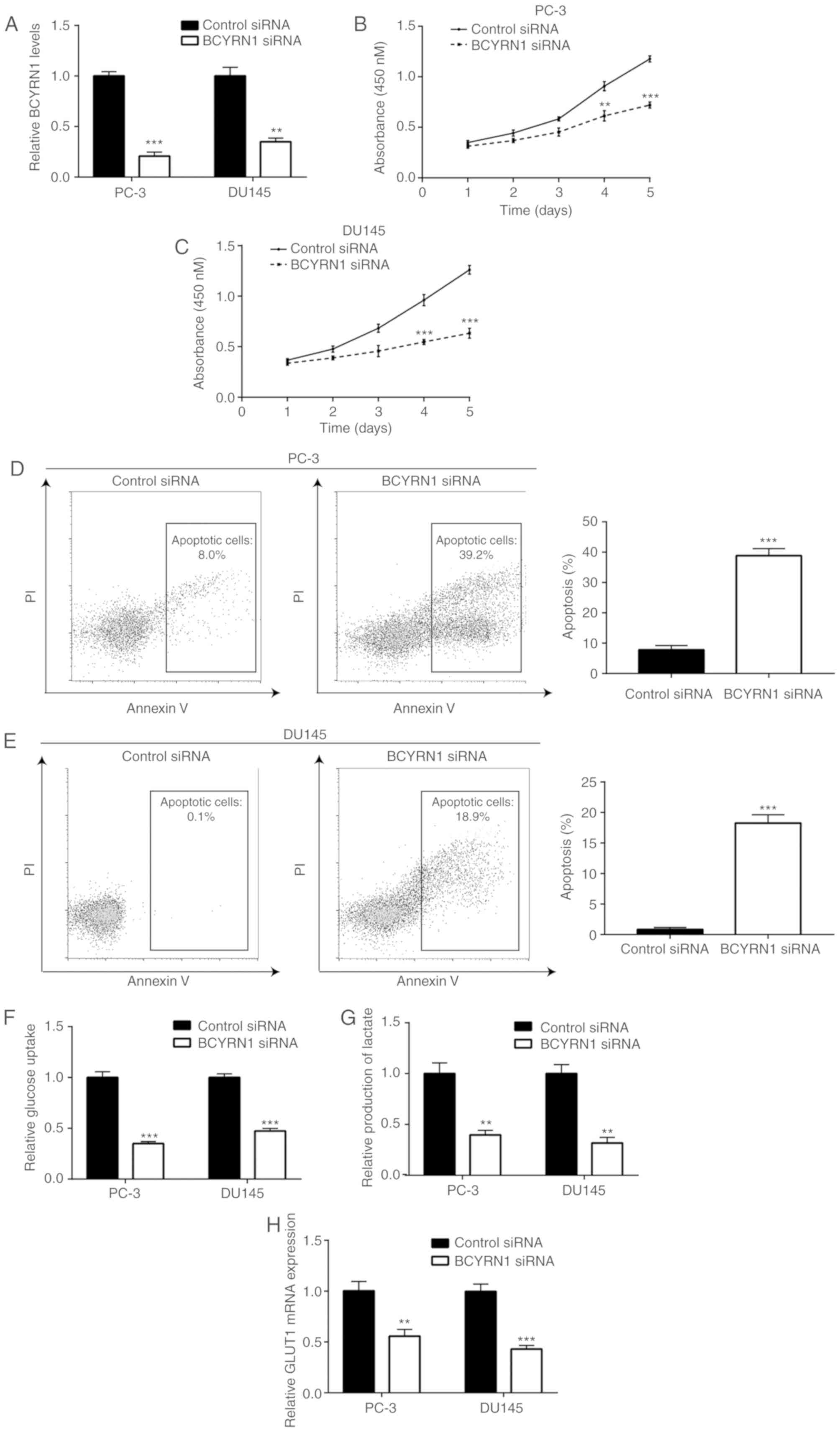
Knockdown of BCYRN1 inhibits cell
proliferation and glucose metabolism, and induces apoptosis in
prostate cancer cells
To investigate the role of BCYRN1 in prostate
cancer, BCYRN1 siRNA was transfected into prostate cancer cells. It
was observed that transfection of BCYRN1 siRNA significantly
decreased BCYRN1 expression in PC-3 and DU145 cells (Fig. 2A). Moreover, the cell proliferation
assay revealed that knockdown of BCYRN1 significantly inhibited the
proliferation of PC-3 and DU145 cells (Fig. 2B and C). Furthermore, the flow
cytometry analysis results demonstrated that BCYRN1 knockdown
induced apoptosis of PC-3 and DU145 cells (Fig. 2D and E). Abnormal glucose metabolism
is crucial for prostate cancer progression. The glucose uptake
assay results suggested that BCYRN1 silencing inhibited the glucose
uptake ability of PC-3 and DU145 cells (Fig. 2F). In addition, BCYRN1 silencing
reduced the levels of lactate produced by PC-3 and DU145 cells,
thereby indicating that glucose metabolism in PC-3 and DU145 cells
is promoted by BCYRN1 (Fig. 2G).
GLUT1 is crucial for glucose metabolism in prostate cancer cells
(23). RT-qPCR analysis revealed
that BCYRN1 silencing decreased GLUT1 mRNA levels in PC-3 and DU145
cells (Fig. 2H).
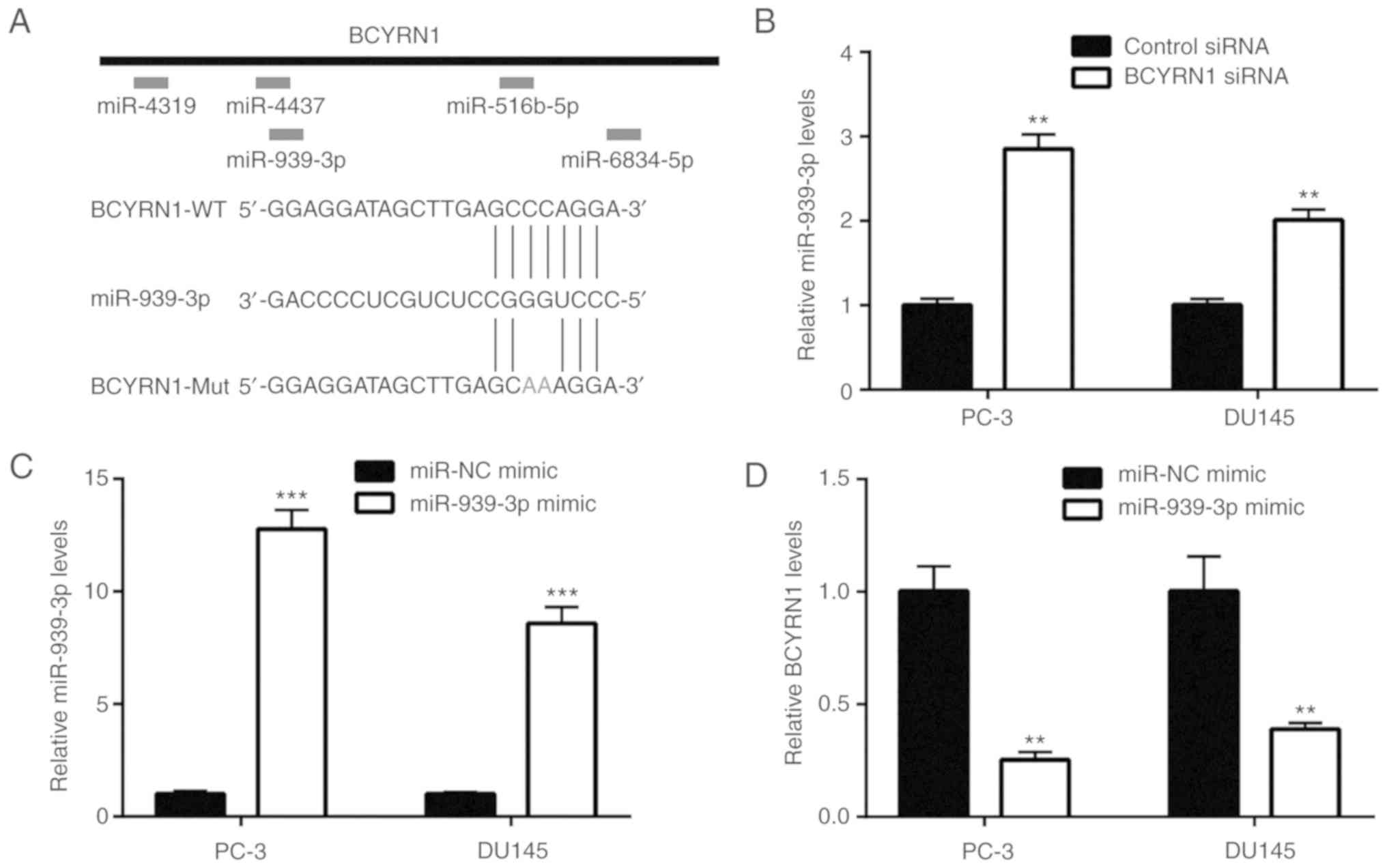
BCYRN1 binds to miR-939-3p and
represses its expression in prostate cancer cells
According to the prediction results of miRDB, five
miRNAs harbored putative binding sites for BCYRN1 (Fig. 3A). Among those, bioinformatics
analysis of prostate cancer datasets (GSE65061 and GSE46738)
revealed that miR-939-3p was downregulated in prostate cancer
(24). Furthermore, the RT-qPCR
results indicated that silencing of BCYRN1 upregulated miR-939-3p
in PC-3 and DU145 cells (Fig. 3B).
In the present study, miR-939-3p was overexpressed in prostate
cancer cells via transfection of miR-939-3p mimic (Fig. 3C). It was observed that elevation of
miR-939-3p decreased BCYRN1 expression in PC-3 and DU145 cells
(Fig. 3D). However, transfection
with a miR-939-3p inhibitor decreased miR-939-3p expression, which
was accompanied by an elevation of BCYRN1 expression in prostate
cancer cells (Fig. 3E and F). It
was also observed that overexpression of miR-939-3p decreased the
relative luciferase activity of BCYRN1-wild-type (WT), but not
BCYRN1-mutant (Mut), which contained mutations in the putative
binding site. Therefore, these results suggested that miR-939-3p
may directly bind to BCYRN1 in PC-3 cells (Fig. 3G). Moreover, miR-939-3p
overexpression reduced the relative luciferase activity of
BCYRN1-WT in DU145 cells (Fig. 3H).
Collectively, these results indicate that BCYRN1 binds to
miR-939-3p and represses its expression in prostate cancer
cells.
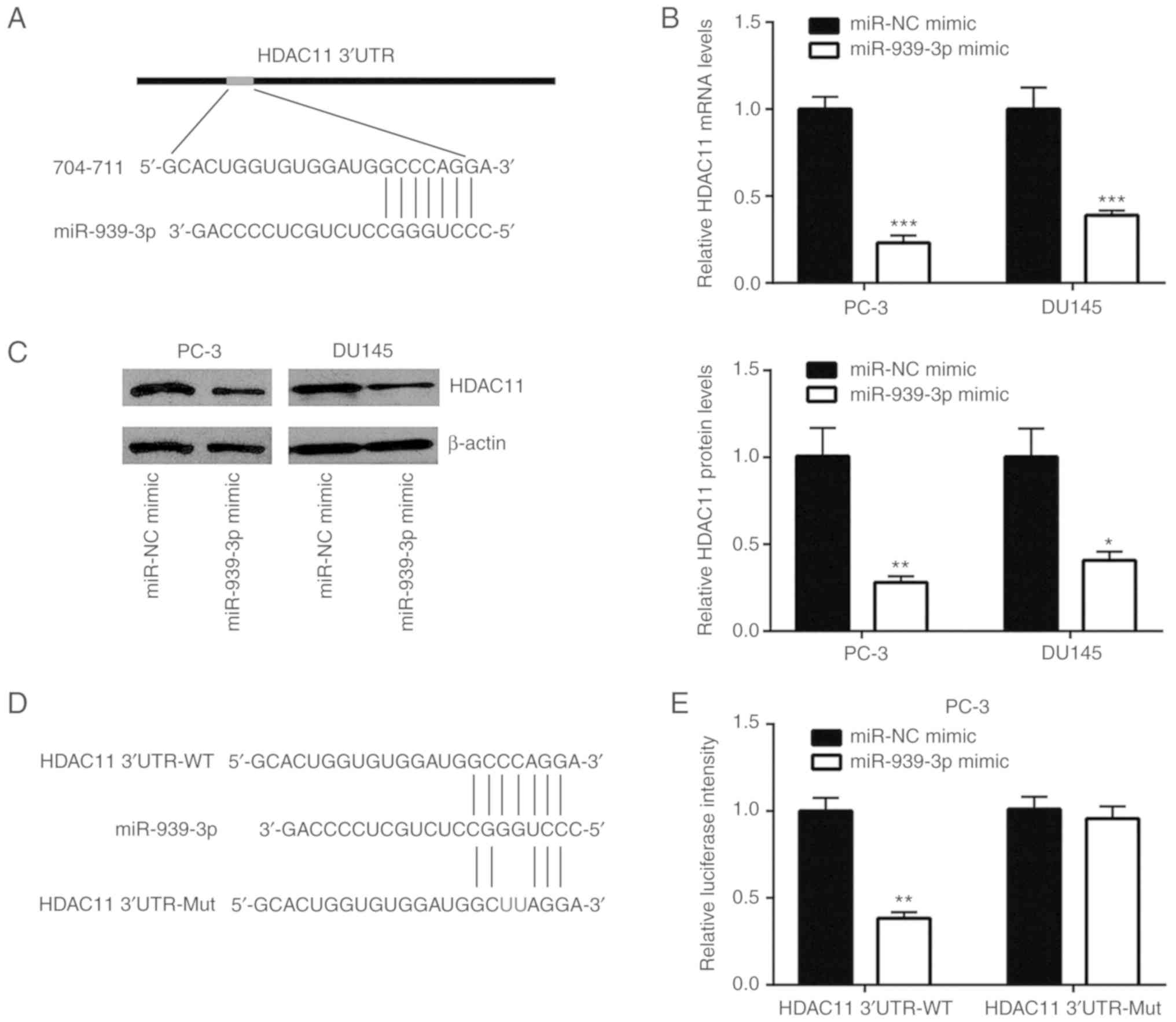
HDAC11 is directly repressed by
miR-939-3p
According to the ceRNA hypothesis, miRNA regulates
gene expression via binding to target gene mRNA (25). In the present study, TargetScan
(http://www.targetscan.org/vert_72/)
predicted HDAC11 as a potential target gene of miR-939-3p (Fig. 4A). Furthermore, overexpression of
miR-939-3p decreased HDAC11 mRNA expression in PC-3 and DU145 cells
(Fig. 4B). Western blotting
demonstrated that miR-939-3p overexpression reduced HDAC11 protein
expression in prostate cancer cells (Fig. 4C). Luciferase plasmids containing
HDAC11 3′UTR-WT and -Mut were used to study the association between
HDAC11 and miR-939-3p (Fig. 4D).
Overexpression of miR-939-3p reduced the relative luciferase
activity of HDAC11 3′UTR-WT, but not HDAC11 3′UTR-Mut in PC-3 cells
(Fig. 4E). Moreover, a similar
result was observed in DU145 cells (Fig. 4F). In addition, silencing of BCYRN1
decreased HDAC11 mRNA expression, which was reversed by miR-939-3p
inhibitor in PC-3 and DU145 cells (Fig.
4G). At the protein expression level, BCYRN1 knockdown reduced
HDAC11 expression, which was reversed by miR-939-3p inhibitor
(Fig. 4H).
BCYRN1 represses tumor suppressor
expression via upregulation of HDAC11 and deacetylation of H3K9 and
H4K16 in prostate cancer
To study the association between BCYRN1 and HDAC11
in prostate cancer, recombinant HDAC11 was transfected into PC-3
cells to upregulate HDAC11 expression (Fig. 5A). It was observed that HDAC11
promoted the progression of prostate cancer mainly via its
deacetylation activity. Overexpression of HDAC11 also increased
HDAC activity, while silencing of BCYRN1 reduced HDAC activity in
PC-3 and DU145 cells (Fig. 5B and
C). Due to abnormal HDAC activity, H3K9 and H4K16 were
hypoacetylated, and the transcription of tumor suppressor genes was
repressed in cancer (26). It was
demonstrated that the global acetylation levels of H3K9 were
elevated following BCYRN1 silencing, and this effect was reversed
after HDAC11 overexpression in PC-3 and DU145 cells (Fig. 5D). Moreover, the global acetylation
levels of H4K16 were elevated following BCYRN1 silencing, and this
effect was reversed after HDAC11 overexpression in PC-3 and DU145
cells (Fig. 5E). Furthermore, the
mRNA expression levels of CDKN1A and OPTN, two tumor suppressor
genes tightly controlled by histone acetylation in prostate cancer
(26), were increased after BCYRN1
silencing and decreased after HDAC11 overexpression in PC-3 and
DU145 cells (Fig. 5F and G).
Therefore, these results indicate that BCYRN1 regulates global
histone acetylation via HDAC11 in prostate cancer.
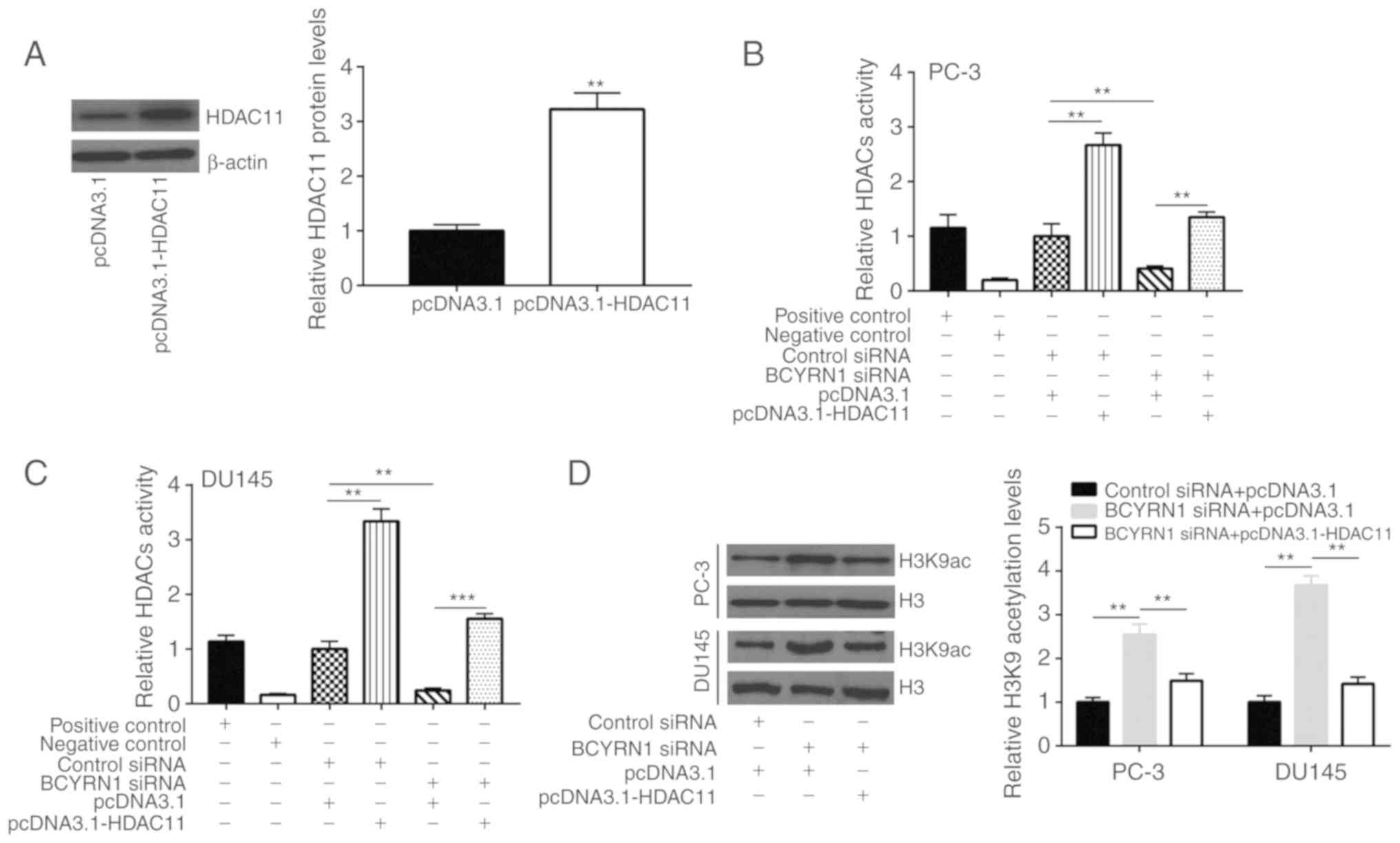 | Figure 5.BCYRN1 decreased global acetylation
of H3K9 and H4K16 via regulating HDAC11 expression. (A)
Transfection of recombinant HDAC11 increased HDAC11 expression in
PC-3 cells. (B) Overexpression of HDAC11 increased HDAC activity,
while silencing of BCYRN1 decreased HDAC activity in PC-3 cells.
(C) Overexpression of HDAC11 increased HDAC activity, while
silencing of BCYRN1 decreased HDAC activity in DU145 cells. (D)
Silencing of BCYRN1 increased global acetylation levels of H3K9,
and HDAC11 overexpression reversed the elevation of H3K9
acetylation in PC-3 and DU145 cells. (E) Silencing of BCYRN1
increased global acetylation levels of H4K16, and HDAC11
overexpression reversed the elevation of H4K16 acetylation in PC-3
and DU145 cells. (F) Silencing of BCYRN1 increased CDKN1A and OPTN
mRNA expression, and HDAC11 overexpression reversed the elevation
of CDKN1A and OPTN mRNA expression in PC-3 cells. (G) Silencing of
BCYRN1 increased CDKN1A and OPTN mRNA expression, and HDAC11
overexpression reversed the elevation of CDKN1A and OPTN mRNA
expression in DU145 cells. **P<0.01; ***P<0.001. BCYRN1,
brain cytoplasmic RNA 1; HDAC11, histone deacetylase 11; CDKN1A,
cyclin-dependent kinase inhibitor 1A; OPTN, optineurin. |
BCYRN1 promotes cell proliferation,
glucose metabolism and cell survival via upregulation of
HDAC11
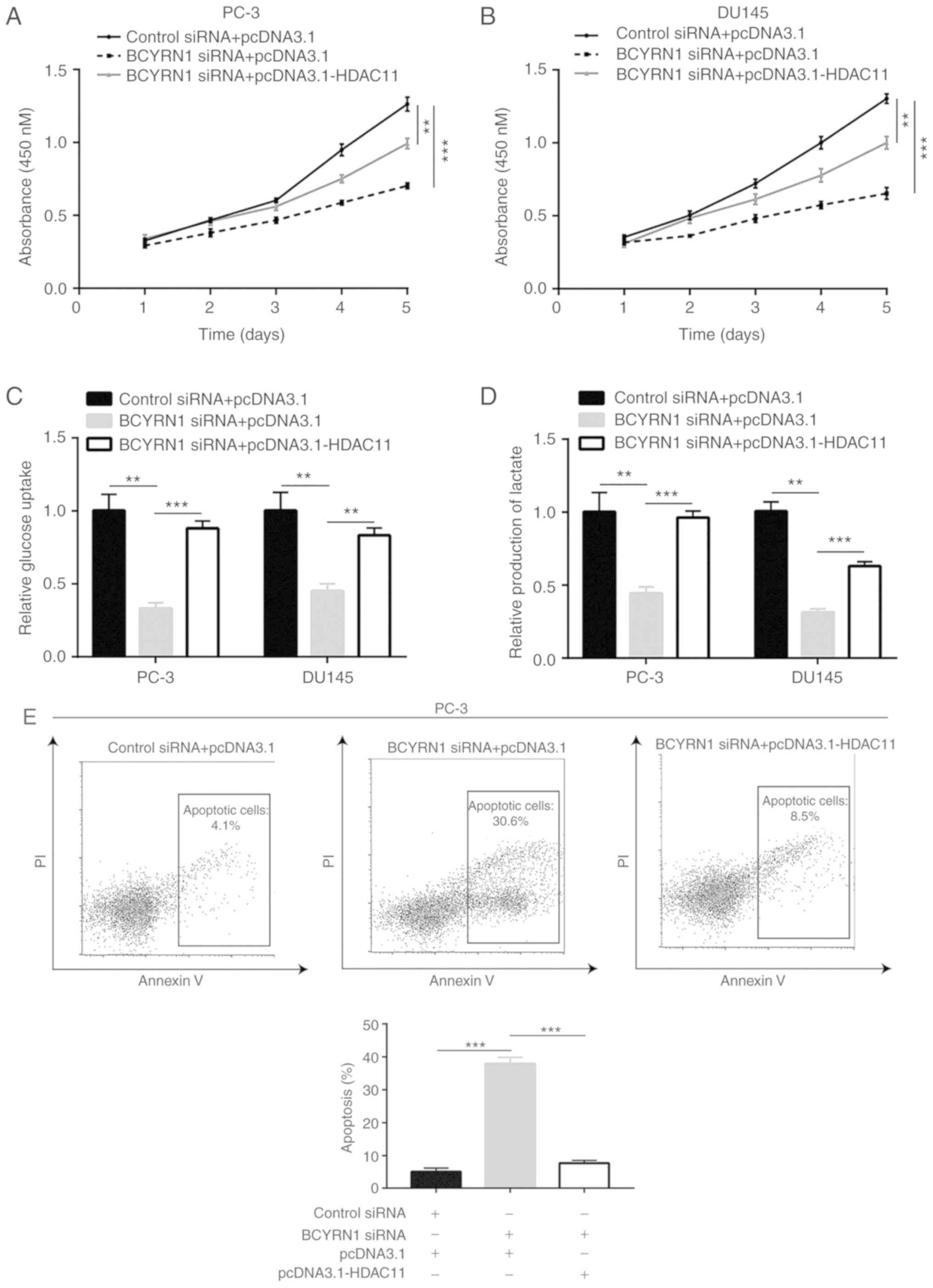
Cell proliferation assay results revealed that
overexpression of HDAC11 reversed si-BCYRN1-induced proliferation
inhibition in PC-3 and DU145 cells (Fig. 6A and B). In addition, HDAC11
upregulation reversed BCYRN1 silencing-induced inhibition of
glucose uptake and lactate production in PC-3 and DU145 cells
(Fig. 6C and D). Thus, BCYRN1 may
promote glucose metabolism via the regulation of HDAC11 expression.
Moreover, HDAC11 overexpression further inhibited apoptosis induced
by BCYRN1 silencing in PC-3 and DU145 cells (Fig. 6E and F). Collectively, the results
mentioned above indicate that BCYRN1 promotes prostate cancer
progression via upregulation of HDAC11.
BCYRN1 is positively associated with
HDAC11 in prostate tumor and healthy prostate tissues
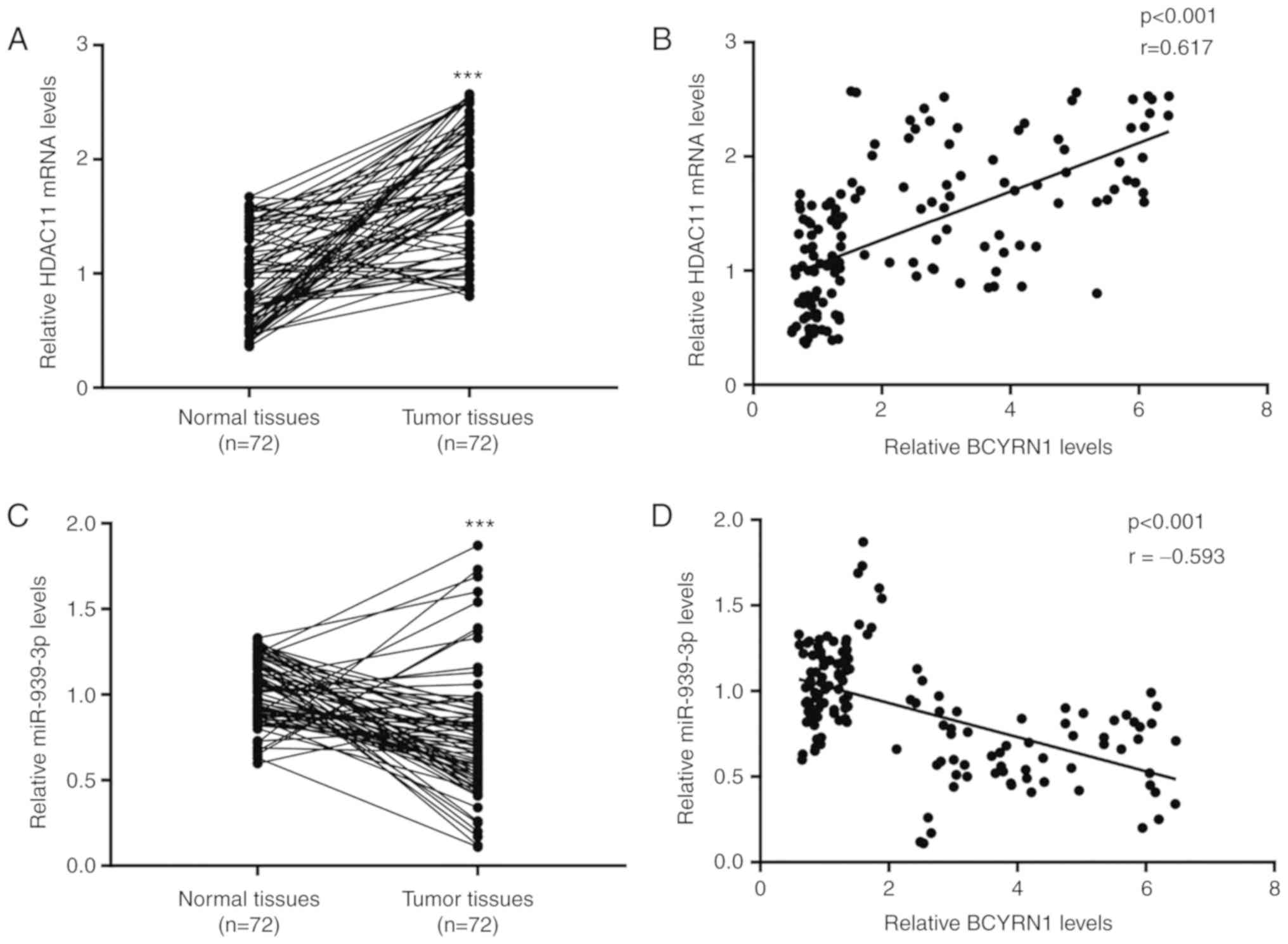
To study the association between BCYRN1 and HDAC11
in clinical samples, HDAC11 mRNA expression was detected in 72
pairs of prostate tumors and healthy prostate tissues. The RT-qPCR
analysis revealed an increase in HDAC11 levels in prostate tumors
(Fig. 7A). Moreover, Pearson's
correlation analysis revealed a strong positive correlation between
BCYRN1 and HDAC11 mRNA expression levels in prostate tumors and
healthy prostate tissues (Fig. 7B).
Furthermore, RT-qPCR also demonstrated that miR-939-3p was
decreased in prostate tumors, and a negative correlation was
observed between miR-939-3p and BCYRN1 levels (Fig. 7C and D).
Discussion
Aberrant expression of BCYRN1 has been reported in
several types of cancer, such as breast, bladder and colon cancer
(27). However, the role of BCYRN1
in prostate cancer has not fully elucidated. Using bioinformatics
analysis of TCGA-PRAD dataset, BCYRN1 was found to be one of the
most significantly upregulated lncRNAs in prostate cancer. In
addition, increased expression of BCYRN1 was detected in prostate
cancers with a high Gleason score compared with those with a low
Gleason score. Reprogrammed energy metabolism is a hallmark of
cancer (28). Furthermore, abnormal
glucose metabolism facilitates prostate cancer cell proliferation
and survival (29). It was
previously demonstrated that lncRNAs play pivotal roles in
mediating glucose metabolism of cancer cells (30). The lncRNA urothelial cancer
associated 1 promotes glucose metabolism via the
mTOR-STAT3/miR-143-HK2 cascade in bladder cancer (31). In the present study, it was observed
that BCYRN1 promoted glycolysis in prostate cancer cells. GLUT1, a
crucial mediator of glucose metabolism in prostate cancer, was
downregulated upon BCYRN1 silencing. Furthermore, BCYRN1
accelerated proliferation and facilitated survival of prostate
cancer cells. It is possible that BCYRN1 sustains glycolysis to
promote prostate cancer cell proliferation and survival.
Collectively, the results of the present study suggested an
oncogenic potential of BCYRN1 in prostate cancer. The biological
function of BCYRN1 was investigated using a single siRNA-mediated
gene knockdown. The role of BCYRN1 will be further explored by
knockdown of BCYRN1 with two independent siRNAs in the future.
HDAC11 is a newly identified oncogene in several
types of cancer (32). It was
previously reported that HDAC11 is a crucial metabolic regulator
(33). Furthermore, depletion of
HDAC11 inhibits metabolism and promotes apoptosis in a variety of
cancer cells, including prostate cancer cells (25). Moreover, the expression of HDAC11 is
regulated by miRNAs (34).
According to a meta-analysis based on 6 datasets, miR-939-3p is one
of 22 commonly downregulated miRNAs in recurrent compared with
non-recurrent prostate cancer samples (28). It has been demonstrated that
lncRNA-HEIH promotes colorectal cancer progression via sponging
miR-939 and upregulation of Bcl-xL (35). In ovarian cancer, however, miR-939
promoted cancer cell proliferation via APC2 targeting and
activation of Wnt signaling (36).
The results of the present study indicated that HDAC11 is a target
gene of miR-939-3p in prostate cancer. Moreover, BCYRN1 sponged
miR-939-3p to elevate HDAC11 levels in prostate cancer cells. As a
deacetylase, HDAC11 controls gene expression via deacetylation of
histone H3 and H4 (37). The
results of the present study suggested that BCYRN1 deacetylated
H3K9 and H4K16 via regulation of HDAC11 in prostate cancer cells.
It has been demonstrated that acetylation of H3K9 and H4K16
regulates the transcription of tumor suppressors and oncogenes in
cancer (26). Furthermore,
hypoacetylation of H4K16 decreases the expression levels of the
tumor suppressor genes CDKN1A and OPTN in prostate cancer (26). The expression of GLUT1 was also
regulated by histone modification in prostate cancer. The RT-qPCR
results demonstrated that BCYRN1 silencing decreased CDKN1A and
OPTN mRNA expression levels, and these effects were reversed
following HDAC11 overexpression in prostate cancer cells. Thus,
BCYRN1 may inhibit tumor suppressor expression via histone
modification. It was observed that HDAC11 overexpression reversed
si-BCYRN1-induced inhibition of glucose metabolism, cell
proliferation and apoptosis in prostate cancer cells. Therefore, it
appears that the BCYRN1/miR-939-3p/HDAC11 axis may be involved in
the regulation of key gene expression levels in prostate cancer
cells, via controlling global histone acetylation. However, the
biological role of BCYRN1 in the initiation, metastasis and
development of prostate cancer in vivo was not investigated
in the present study. BCYRN1 stable knockout prostate cancer cells
will be established in the future to further study the function of
BCYRN1 in vivo.
In conclusion, the results of the present study
suggested that BCYRN1 is upregulated in prostate cancer, and that
it promotes proliferation, glucose metabolism and survival of
prostate cancer cells via sponging miR-939-3p and increasing the
levels of HDAC11. Therefore, the present study may provide novel
evidence that BCYRN1 may be of value as a biomarker and therapeutic
target in prostate cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All the datasets generated and/or analyzed during
the present study are available from the corresponding author on
reasonable request.
Authors' contributions
WH, FQ and KW participated in the experimental
design and data acquisition; FQ contributed to the collection of
prostate specimens and clinical data; KW supervised the whole study
and wrote the manuscript. All the authors have read and approved
the final version of the manuscript for publication.
Ethics approval and consent to
participate
All procedures performed in the present study
involving human participants were approved by the Institutional
Ethics Review Board of China-Japan Union Hospital.
Patient consent for publication
Written informed consent for the publication of all
data was provided by all patients prior to surgery.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kaarbø M, Mikkelsen OL, Malerød L, Qu S,
Lobert VH, Akgul G, Halvorsen T, Maelandsmo GM and Saatcioglu F:
PI3K-AKT-mTOR pathway is dominant over androgen receptor signaling
in prostate cancer cells. Cell Oncol. 32:11–27. 2010.PubMed/NCBI
|
|
3
|
Bhardwaj S and Varma S: Rare incidence of
tumor lysis syndrome in metastatic prostate cancer following
treatment with docetaxel. J Oncol Pharm Pract. 24:153–155. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cai Z, Wu Y, Li Y, Ren J and Wang L: BCAR4
activates GLI2 signaling in prostate cancer to contribute to
castration resistance. Aging (Albany NY). 10:3702–3712. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang XZ, Beebe JR, Pwiti L, Bielawska A
and Smyth MJ: Aberrant sphingolipid signaling is involved in the
resistance of prostate cancer cell lines to chemotherapy. Cancer
Res. 59:5842–5848. 1999.PubMed/NCBI
|
|
6
|
Chen X and Yan GY: Novel human
lncRNA-disease association inference based on lncRNA expression
profiles. Bioinformatics. 29:2617–2624. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McHugh CA, Chen CK, Chow A, Surka CF, Tran
C, McDonel P, Pandya-Jones A, Blanco M, Burghard C, Moradian A, et
al: The Xist lncRNA interacts directly with SHARP to silence
transcription through HDAC3. Nature. 521:232–236. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang Q, Wan Q, Zhang L, Li Y, Zhang P, Li
D, Feng C, Yi F, Zhang L, Ding X, et al: Analysis of LncRNA
expression in cell differentiation. RNA Biol. 15:413–422. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qiu M, Xu Y, Wang J, Zhang E, Sun M, Zheng
Y, Li M, Xia W, Feng D, Yin R and Xu L: A novel lncRNA, LUADT1,
promotes lung adenocarcinoma proliferation via the epigenetic
suppression of p27. Cell Death Dis. 6:e18582015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Su M, Xiao Y, Tang J, Wu J, Ma J, Tian B,
Zhou Y, Wang H, Yang D, Liao QJ and Wang W: Role of lncRNA and EZH2
interaction/regulatory network in lung cancer. J Cancer.
9:4156–4165. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zan XY and Li L: Construction of
lncRNA-mediated ceRNA network to reveal clinically relevant lncRNA
biomarkers in glioblastomas. Oncol Lett. 17:4369–4374.
2019.PubMed/NCBI
|
|
12
|
Du Y, Weng XD, Wang L, Liu XH, Zhu HC, Guo
J, Ning JZ and Xiao CC: LncRNA XIST acts as a tumor suppressor in
prostate cancer through sponging miR-23a to modulate RKIP
expression. Oncotarget. 8:94358–94370. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lingadahalli S, Jadhao S, Sung YY, Chen M,
Hu L, Chen X and Cheung E: Novel lncRNA LINC00844 regulates
prostate cancer cell migration and invasion through AR signaling.
Mol Cancer Res. 15:1865–1878. 2018. View Article : Google Scholar
|
|
14
|
Gu P, Chen X, Xie R, Xie W, Huang L, Dong
W, Han J, Liu X, Shen J, Huang J and Lin T: A novel AR
translational regulator lncRNA LBCS inhibits castration resistance
of prostate cancer. Mol Cancer. 18:1092019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shin H, Kim Y, Kim M and Lee Y: BC200 RNA:
An emerging therapeutic target and diagnostic marker for human
cancer. Mol Cells. 41:993–999. 2018.PubMed/NCBI
|
|
16
|
Wu K, Xu K, Liu K, Huang J, Chen J, Zhang
J and Zhang N: Long noncoding RNA BC200 regulates cell growth and
invasion in colon cancer. Int J Biochem Cell Biol. 99:219–225.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao RH, Zhu CH, Li XK, Cao W, Zong H, Cao
XG and Hu HY: BC200 LncRNA a potential predictive marker of poor
prognosis in esophageal squamous cell carcinoma patients. Onco
Targets Ther. 9:2221–2226. 2016.PubMed/NCBI
|
|
18
|
Iacoangeli A, Adzovic L, Chen EQ, Latif
Cattie R, Soff GA and Tiedge H: Regulatory BC200 RNA in peripheral
blood of patients with invasive breast cancer. J Investig Med.
66:1055–1063. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thole TM, Lodrini M, Fabian J, Wuenschel
J, Pfeil S, Hielscher T, Kopp-Schneider A, Heinicke U, Fulda S,
Witt Om, et al: Neuroblastoma cells depend on HDAC11 for mitotic
cell cycle progression and survival. Cell Death Dis. 8:e26352017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Byun SK, An TH, Son MJ, Lee DS, Kang HS,
Lee EW, Han BS, Kim WK, Bae KH, Oh KJ and Lee SC: HDAC11 inhibits
myoblast differentiation through repression of MyoD-dependent
transcription. Mol Cells. 40:667–676. 2017.PubMed/NCBI
|
|
21
|
Deubzer HE, Schier MC, Oehme I, Lodrini M,
Haendler B, Sommer A and Witt O: HDAC11 is a novel drug target in
carcinomas. Int J Cancer. 132:2200–2208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vaz CV, Alves MG, Marques R, Moreira PI,
Oliveira PF, Maia CJ and Socorro S: Androgen-responsive and
nonresponsive prostate cancer cells present a distinct glycolytic
metabolism profile. Int J Biochem Cell Biol. 44:2077–2084. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pashaei E, Pashaei E, Ahmady M, Ozen M and
Aydin N: Meta-analysis of miRNA expression profiles for prostate
cancer recurrence following radical prostatectomy. PLoS One.
12:e01795432017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo LL, Song CH, Wang P, Dai LP, Zhang JY
and Wang KJ: Competing endogenous RNA networks and gastric cancer.
World J Gastroenterol. 21:11680–11687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Katoh H, Qin ZS, Liu R, Wang L, Li W, Li
X, Wu L, Du Z, Lyons R, Liu CG, et al: FOXP3 orchestrates H4K16
acetylation and H3K4 trimethylation for activation of multiple
genes by recruiting MOF and causing displacement of PLU-1. Mol
Cell. 44:770–784. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Samson J, Cronin S and Dean K: BC200
(BCYRN1)-The shortest, long, non-coding RNA associated with cancer.
Noncoding RNA Res. 3:131–143. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang B, Zhang L, Cao Y, Chen S, Cao J, Wu
D, Chen J, Xiong H, Pan Z, Qiu F, et al: Overexpression of lncRNA
IGFBP4-1 reprograms energy metabolism to promote lung cancer
progression. Mol Cancer. 16:1542017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen S, Xu X, Lu S and Hu B: Long
non-coding RNA HAND2-AS1 targets glucose metabolism and inhibits
cancer cell proliferation in osteosarcoma. Oncol Lett.
18:1323–1329. 2019.PubMed/NCBI
|
|
31
|
Li Z, Li X, Wu S, Xue M and Chen W: Long
non-coding RNA UCA1 promotes glycolysis by upregulating hexokinase
2 through the mTOR-STAT3/microRNA143 pathway. Cancer Sci.
105:951–955. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Leslie PL, Chao YL, Tsai YH, Ghosh SK,
Porrello A, Van Swearingen AED, Harrison EB, Cooley BC, Parker JS,
Carey LA and Pecot CV: Histone deacetylase 11 inhibition promotes
breast cancer metastasis from lymph nodes. Nat Commun. 10:41922019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bhaskara S: Histone deacetylase 11 as a
key regulator of metabolism and obesity. EBioMedicine. 35:27–28.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin L, Hou J, Ma F, Wang P, Liu X, Li N,
Wang J, Wang Q and Cao X: Type I IFN inhibits innate IL-10
production in macrophages through histone deacetylase 11 by
downregulating microRNA-145. J Immunol. 191:3896–3904. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cui C, Zhai D, Cai L, Duan Q, Xie L and Yu
J: Long noncoding RNA HEIH promotes colorectal cancer tumorigenesis
via counteracting miR-939 mediated transcriptional repression of
Bcl-xL. Cancer Res Treat. 50:992–1008. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ying X, Liya Q, Feng Z, Yin W and Ji-hong
L: MiR-939 promotes the proliferation of human ovarian cancer cells
by repressing APC2 expression. Biomed Pharmacother. 71:64–69. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu H, Hu Q, D'Ercole AJ and Ye P: Histone
deacetylase 11 regulates oligodendrocyte-specific gene expression
and cell development in OL-1 oligodendroglia cells. Glia. 57:1–12.
2009. View Article : Google Scholar : PubMed/NCBI
|