Introduction
Esophageal squamous cell carcinoma (ESCC) is one of
the most common malignant tumors in China, and its mortality rate
ranks fourth (1,2) among the top 10 cancers in China. In
particular, the Yili region of Xinjiang is a high-incidence area of
ESCC, with the highest incidence observed in the Kazakh population,
reaching 150/100,000, with a 5-year survival rate of only 10–13%
(3,4). Therefore, identifying malignant
phenotype-related targets and elucidating the molecular regulatory
mechanisms involved in the progression of ESCC is crucial for
accurate diagnosis, timely individualized treatment and improvement
of the survival rate of ESCC.
Pyruvate kinase isoenzyme M2 (PKM2), a rate-limiting
enzyme in cellular glycolysis, plays a key role in anaerobic
glycolysis (5). Generally, PKM2
exerts tumor-promoting effects via regulation of a number of
signaling pathways in tumor cells, operating as a potential protein
kinase (6). Our previous study
(7) demonstrated that PKM2 was
markedly overexpressed in ESCC tissues from Kazakh patients
compared with paired adjacent normal tissues through proteomics,
suggesting its oncological trait. Our subsequent research on PKM2
(8) and a relevant study by other
researchers (9) indicated that
upregulated PKM2 may promote the growth and motility of ESCC cells,
and that the upregulation of PKM2 in Kazakh patients with ESCC is
closely associated with shorter overall survival. However, the
molecular mechanism underlying the role of PKM2 in Kazakh patients
with ESCC remains elusive.
Heat shock proteins (HSPs) are a group of highly
conserved molecular chaperones, mainly comprising HSP27, HSP60,
HSP70 and HSP90 (10). Although the
relative molecular mass of each of the main HSP groups is distinct,
they share similar or identical conserved domains (11), and play multiple roles in cell
functions, including protein folding, assembly and degradation
(12). HSP27 has been reported to
be commonly upregulated in several different types of tumors,
including ESCC, and is important in the regulation of cancer cell
proliferation, apoptosis and metastasis (13–15).
Additionally, it also has important biological functions in the
regulation of various carcinogenic proteins (16). However, few studies have described
the regulatory association between HSP27 and PKM2 in ESCC. By
contrast, one recent study reported that HSP90 could stabilize and
regulate PKM2 abundance, thereby accelerating anaerobic glycolysis
in hepatocellular carcinoma (HCC) (7), which suggests that HSP27 may also
interact with PKM2.
Small ubiquitin-related modified proteins (SUMOs)
are a class of small protein molecules (17). There are four SUMO homologues in the
human genome, namely SUMO1, SUMO2, SUMO3 and SUMO4. SUMO2 and SUMO3
currently cannot be distinguished by antibodies, as they have ~97%
sequence homology except for a difference of two amino acids, and
they are collectively referred to as SUMO2/3 (18). An important post-translational
modification involving SUMO proteins, which has been termed
SUMOylation, is involved in the modulation of various cellular
processes, including transcriptional regulation, subcellular
localization and target protein stability (19). Extensive research has confirmed that
SUMOylation plays a crucial role in human diseases (20). It was recently reported that HSP27
and SUMO2/3 combine directly, which induces SUMOylation of HSP27 to
accelerate the invasion and metastasis of HCC cells (21). However, it remains unclear whether
there is an interaction between HSP27 and SUMO2/3 in ESCC.
In the present study, based on the findings of
previous research, it was hypothesized that HSP27 could physically
bind with SUMO2/3, thereby upregulating PKM2 in ESCC cells. To test
this hypothesis, experiments were conducted on ESCC specimens and
cell lines. Immunohistochemistry (IHC) was performed to examine
whether the expression of HSP27 was associated with local
infiltration, lymph node metastasis and prognosis. Moreover, the
association between HSP27 and PKM2 in ESCC tissues was
investigated. The aim of the present study was to elucidate the
effects of HSP27 on the growth and motility of ESCC cells, to
determine whether SUMOylation of HSP27 can promote the malignant
behavior of ESCC cells, and whether this effect is mediated by
regulating PKM2 expression.
Materials and methods
Pathological tissue specimens
The present study was approved by the Medical Ethics
Committee of the First Affiliated Hospital of Xinjiang Medical
University, and informed consent was obtained from each participant
involved. There were 180 ESCC samples on the tissue microarray chip
(HEso-Squ180Sur-04, Outdo Biotech), including 100 cancer tissue
specimens and 80 normal adjacent tissues (NATs). A chip with 165
samples of Kazakh ESCC tissues from the First Affiliated Hospital
of Xinjiang Medical University included 99 ESCC tissues and 57
NATs. None of the patients had received anticancer treatment prior
to surgical resection. All cases were diagnosed and confirmed
histopathologically. The stages and classifications were graded in
accordance with the 2017 WHO classification and grading system
(22).
IHC staining
The tissue microarray was dewaxed, hydrated, treated
with EDTA antigen repair solution (Sigma-Aldrich; Merck KGaA) and
blocked in serum (Zhongshanjinqiao) for 1 h at room temperature.
Endogenous peroxidase activity was blocked with 3%
H2O2. Subsequently, the tissues were probed
at 4°C overnight with primary antibodies targeting HSP27 (1:200,
ab12351, Abcam) and PKM2 (1:100, 15822-1-AP, ProteinTech Group,
Inc.), followed by addition of the secondary antibody (KIT-5030,
MXB Biotech) with incubation at 37°C for 1 h, and staining using a
DAB kit (ZLI-9018, OriGene Technologies, Inc.) for 2 min at room
temperature. The immunostaining was scored by a pathological image
analysis software processing system (ImageJ software, version 1.50;
National Institutes of Health). According to the comprehensive
analysis of staining intensity (0, none; 1, weak; 2, moderate; and
3, strong) and percentage of stained cells (0, <5%; 1, 5–25%; 2,
25–50%; and 3, >50%), each specimen was graded using a total
score derived from adding the staining intensity and percentage
scores. Scores <2 were defined as negative and >2 as
positive.
Cell culture
The KYSE30, KYSE150, KYSE450 and KYSE510 cells were
kind gifts from Professor Zhihua Liu (Chinese Academy of Sciences,
Beijing, China); and TE-1, Eca109 and EC9706 cells were purchased
from Wuhan University, China. All the cells were maintained in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and placed at 5% CO2 and 37°C.
Lentivirus transfection
ESCC cells were plated in 6-well plates
(2×105/well) with 20 µl lentivirus (Lv; 1×108
TU/ml; Genechem Co., Ltd.), 40 µl HiTrans transfection reagent
(Genechem Co., Ltd.) and 940 µl RPMI-1640 medium (10% FBS) at 5%
CO2 and 37°C for 18 h. The infected cells were sorted by
flow cytometry and evaluated by western blotting. HSP27 shRNA
(Lv-shRNA-HSP27-1, Lv-shRNA-HSP27-2 and Lv-shRNA-HSP27-3) and
SUMO2/3 shRNA were obtained from Shanghai GeneChem Co., Ltd. The
shRNA target sequences were as follows: shHSP27-1:
5′-GATGAGACTGCCGCCAAGT-3′; shHSP27-2: 5′-CGGACGAGCTGACGGT-CAA-3′;
shHSP27-3: 5′-CAGTGGTTAGGCGGCAGCA-3′; and scramble:
5′-TTCTCCGAACGT-GTCACGT-3′.
Western blotting
The total proteins (on ice) were extracted with RIPA
lysis buffer (Beijing Solarbio Science & Technology Co., Ltd.),
and the concentration of protein extracted was calculated by a BCA
kit. The proteins were separated via 12% SDS-PAGE and then
transblotted to PVDF membranes. Then, they were incubated with 0.5%
skimmed milk for 2 h at room temperature and the following primary
antibodies overnight at 4°C: HSP27 (1:800, cat. no. ab12351,
Abcam), PKM2 (1:1,000, cat. no. 15822-1-AP, ProteinTech Group,
Inc.), E-cadherin (1:1,000, cat. no. 20874-1-AP, ProteinTech Group,
Inc.), vimentin (1:1,000, cat. no. 10366-1-AP, ProteinTech Group,
Inc.), SUMO2/3 (1:800, cat. no. ab190757, Abcam), and GAPDH (1:500,
cat. no. 20874-1-AP, ProteinTech Group, Inc.). A western blot assay
kit (WB7105, Thermo Fisher Scientific, Inc.) provided the secondary
antibody, which was applied at room temperature for 1 h, and then
chromogen (supplied in the kit) was applied for 5 min in the
dark.
MTT assay
A total of 5,000 cells per well were seeded in a
96-well plate. Cells were cultured in RPMI-1640 medium containing
10% FBS. Detection was conducted at 0, 24, 48, 72 and 96 h. Each
group was detected in quintuplicate at each time point. MTT (5
mg/ml, Sigma-Aldrich; Merck KGaA) was added to each well at 20
µl/well for 4 h in the dark, after which time the liquid was
discarded and DMSO (135 µl/well, Sigma-Aldrich; Merck KGaA) was
added for 10 min in the dark to dissolve the crystals. The
absorbance was measured at 490 nm by an enzyme labelling instrument
(Benchmark™ Microplate Reader; Bio-Rad Laboratories, Inc.).
Transwell assay
Transwell chambers (24-well; Corning, Inc.) with a
5-µm pore filter were coated with a mixture of 60 ml 1:9 Matrix (BD
Biosciences) and serum-free RPMI-1640 medium to form a matrix
barrier. Precoating was performed at 37°C for 1 h and the Transwell
inserts were refrigerated at 4°C overnight. Before inoculation,
RPMI-1640 medium was used to hydrate the basement membrane for 30
min at 37°C. The upper chamber cells were filled with 200 µl
serum-free RPMI-1640 medium, and the lower chamber was filled with
500 µl RPMI-1640 medium supplemented with 10% FBS. Following
incubation at 37°C for 48 h, the cells were washed twice with PBS
(5 min per wash) and fixed with 600 µl polyformaldehyde for 15 min
at 4°C. The non-invading cells were wiped with a cotton swab,
washed with ddH2O three times and stained with 5%
crystal violet solution for 5 min at room temperature (23). The number of invading cells was
counted in four random fields using an inverted phase contrast
microscope (Olympus Corporation) at a ×50 magnification.
Wound healing assay
Cells were seeded in a 6-well cell culture plate
(Corning, Inc.) and cultured in serum-free medium for 24 h at room
temperature. After the cells had reached a confluence of >80%,
three parallel scratches were created with a 10-µl pipette tip in
each hole. Images were captured using an inverted phase contrast
microscope (Olympus Corporation) at 0 and 24 h after scratching.
Cell migration was quantitated by measuring the blank area between
the moving cell boundaries using Image J software, version 1.50
(National Institutes of Health).
Co-immunoprecipitation (Co-IP)
Co-IP was carried according to the instructions
included in the assay kit (Pierce 26149, Thermo Fisher Scientific,
Inc.). First, the total protein lysate was extracted, and 500 µg
total protein was mixed with 2 µg monoclonal antibody or IgG and
oscillated for 4 h at 4°C. The protein beads in the kit were added
to the mixture and oscillated overnight at 4°C. Subsequently, the
beads were washed using IP buffer three times. Sample buffer (5X)
was mixed with the beads and boiled for 5 min. The supernatant was
prepared for western blot analysis.
Immunofluorescence
Cells for immunofluorescence were fixed with 4%
paraformaldehyde at 4°C for 20 min and then subjected to treatment
with 0.5% Triton X-100 diluted with PBS at room temperature for 30
min. Cells were blocked in PBS with 2% BSA (Sigma-Aldrich; Merck
KGaA) for 30 min at 37°C and incubated with antibodies specific for
HSP27 (1:200, cat. no. 66767-1-Ig, ProteinTech Group Inc.) and
SUMO2/3 (1:200, cat. no. ab190757, Abcam) overnight at 4°C. Alexa
Fluor-conjugated secondary antibodies (Alexa Fluor 488, cat. no.
ab150077, Abcam and Alexa Fluor 594, cat. no. ab150120, Abcam) were
incubated for 2 h at room temperature in the dark. Hoechst 33342
was then used for counterstaining the nucleus for 15 min at room
temperature, and images were captured by a laser scanning confocal
microscope at a magnification of ×200 (LSM710, Carl Zeiss AG).
Statistical analysis
SPSS 19.0 (IBM Corp.) was used to analyze and
process the data. The data are expressed as the mean ± standard
deviation. Student's t-test or one-way ANOVA with Bonferroni
analysis were used to evaluate the differences among groups.
Cross-table test and Pearson's correlation analysis were adopted to
analyze the correlation between HSP27 and PKM2. The Kaplan-Meier
method was employed to analyze overall survival data. P<0.05 was
considered to indicate statistically significant differences.
Results
Overexpression of HSP27 is associated
with ESCC progression and adverse outcomes
To investigate whether HSP27 is involved in the
pathological process of ESCC, 224 ESCC tissues and 121 NATs were
collected. HSP27 expression was detected by IHC, which revealed
that HSP27 was markedly overexpressed in cancer tissues compared
with control tissues (Fig. 1A,
Table I). The association between
HSP27 level and pathological information was analyzed; Table I shows that HSP27 was significantly
correlated with T classification and lymph node metastasis.
However, there was no marked correlation with other pathological
information, including demographic characteristics, tumor size,
pathological grade and pathological classification. Kaplan-Meier
survival analysis revealed that the survival of ESCC patients with
high HSP27 expression was markedly inferior to that of those with
low HSP27 expression (Fig. 1B,
P<0.05). In addition, to further confirm the correlation between
HSP27 and PKM2 in ESCC tissues, the expression of PKM2 and HSP27
expression was next investigated in 165 Kazakh ESCC tissues by IHC
staining. The results demonstrated that both HSP27 and PKM2 were
upregulated in Kazakh ESCC tissues compared with NATs (Fig. 1A and C). The subcellular
localization of HSP27 and PKM2 was mainly cytoplasmic, with that of
PKM2 being nucleoplasmic in a few cases, in accordance with the
positive staining indicated by the primary antibodies used.
Furthermore, the χ2 test (χ2=4.797, P=0.029,
Table II) and Spearman's
correlation analysis (r=0.253, P=0.0011, Fig. 1D) revealed that HSP27 expression was
significantly correlated with PKM2.
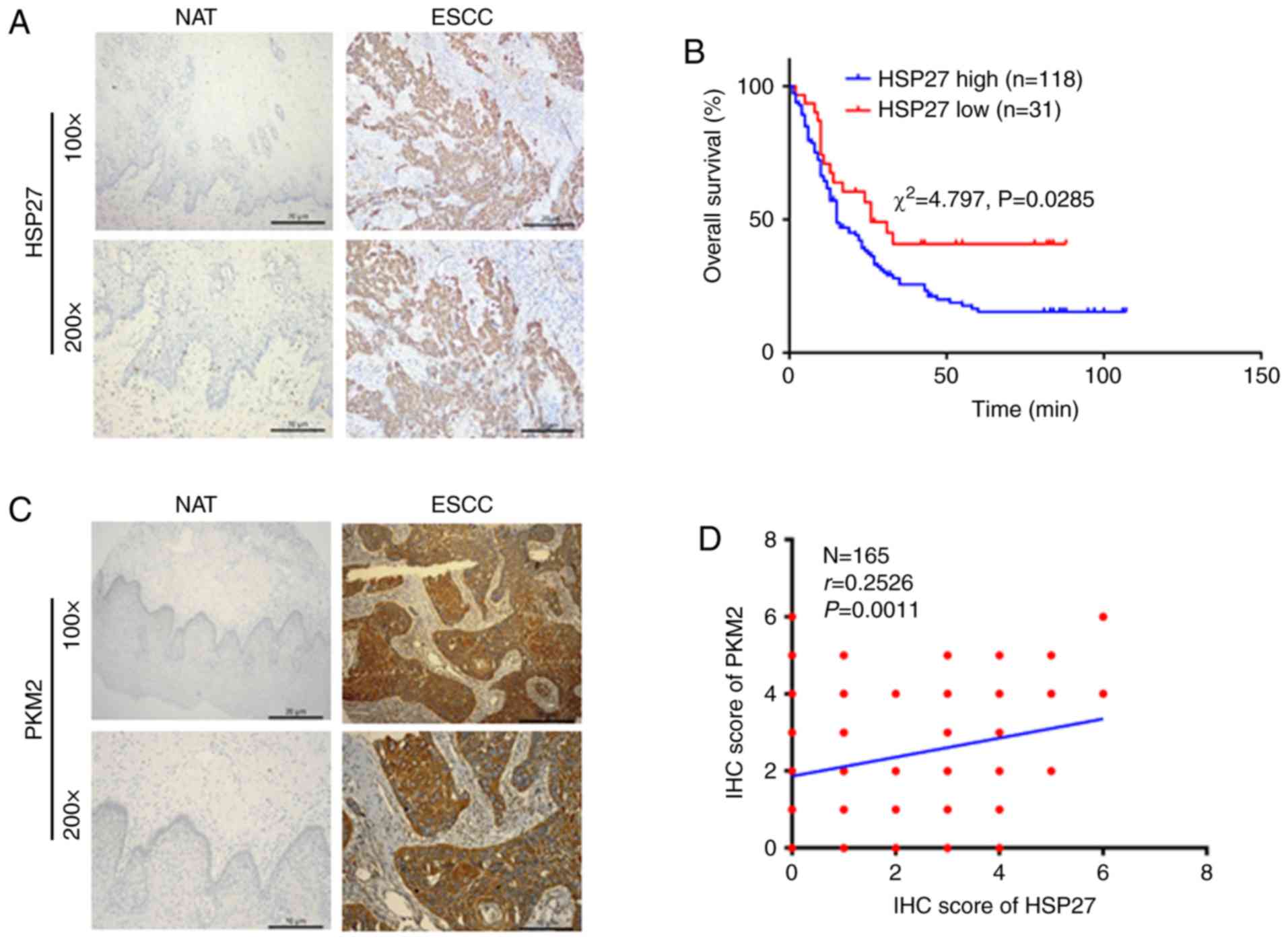 | Figure 1.HSP27 is markedly upregulated in ESCC
and is correlated with PKM2 expression and poor prognosis. (A) IHC
results of HSP27 expression in ESCC tissues and NAT (magnification,
×100 and ×200). (B) Survival curve of HSP27 expression in ESCC. (C)
IHC results of PKM2 in ESCC tissues and NAT (magnification, ×100
and ×200). (D) Spearman's correlation analysis between HSP27 and
PKM2 expression in ESCC. Representative selected images are shown.
The log-rank test was used in survival analysis
(χ2=4.797, P=0.0285), and Spearman's method was employed
to analyze the correlation (N=165, r=0.2526, P=0.0011). ESCC,
esophageal squamous cell carcinoma; HSP, heat shock protein; PKM2,
pyruvate kinase isoenzyme 2; IHC, immunohistochemistry; NAT, normal
adjacent tissue. |
 | Table I.Clinicopathological significance of
HSP27 expression in ESCC. |
Table I.
Clinicopathological significance of
HSP27 expression in ESCC.
|
|
| HSP27 expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Characteristics | N | High | Low | χ2 | P-value |
|---|
| Tissue type |
|
|
|
|
|
|
ESCC | 224 | 152 | 72 | 18.854 | 0.000 |
|
NAT | 121 | 53 | 68 |
|
|
| Sex |
|
|
| 1.817 | 0.178 |
|
Male | 239 | 147 | 92 |
|
|
|
Female | 106 | 57 | 49 |
|
|
| Age (years) |
|
|
| 0.008 | 0.928 |
|
≤60 | 139 | 83 | 56 |
|
|
|
>60 | 206 | 122 | 84 |
|
|
| Tumor diameter
(cm) |
|
|
| 1.34 | 0.247 |
|
<5 | 200 | 115 | 85 |
|
|
| ≥5 | 117 | 75 | 42 |
|
|
| Pathological
grade |
|
|
| 2.636 | 0.268 |
| I | 41 | 28 | 13 |
|
|
| II | 211 | 121 | 90 |
|
|
|
III | 91 | 59 | 32 |
|
|
| Pathological
type |
|
|
| 5.216 | 0.157 |
|
Ulcerative | 190 | 111 | 79 |
|
|
|
Medullary | 88 | 50 | 38 |
|
|
|
Protruding | 37 | 23 | 14 |
|
|
|
Stenosing | 7 | 7 | 0 |
|
|
| T
classification |
|
|
| 16.4 | 0.000 |
|
T1-2 | 86 | 34 | 52 |
|
|
|
T3-4 | 243 | 157 | 86 |
|
|
| Lymph node
metastasis |
|
|
| 3.889 | 0.049 |
| N0 | 204 | 111 | 93 |
|
|
|
N1-N3 | 135 | 88 | 47 |
|
|
| Table II.Correlation between HSP27 and PKM2
expression in ESCC. |
Table II.
Correlation between HSP27 and PKM2
expression in ESCC.
|
|
| HSP27
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| PKM2
expression | N | High | Low | χ2 | P-value |
|---|
| High | 91 | 64 | 27 | 5.406 | 0.024 |
| Low | 74 | 39 | 35 |
|
|
| Total | 165 | 103 | 62 |
|
|
HSP27 promotes the growth and motility
of ESCC cells
Next, to explore the biological roles of HSP27 in
the growth and invasion of ESCC, HSP27 expression in seven ESCC
cell lines (Eca109, EC9706, KYSE30, KYSE450, TE-1, KYSE150 and
KYSE510) was evaluated using immunoblotting; among these cell
lines, KYSE450 exhibited the highest level of HSP27 expression,
while KYSE150 had the second-highest HSP70 level (Fig. 2A). To determine the biological
function of HSP27 in ESCC, Lv-based shRNA targeting HSP27 was
transfected into KYSE450 and KYSE150 cells. Stable HSP27 knockdown
cell lines were established successfully and confirmed by western
blotting (Fig. 2B). Next, the
absorbance of the HSP27 knockdown group was reduced significantly
compared with that of the control group in KYSE450 cells with the
extension of time in the MTT assay (Fig. 2C). KYSE150 cells exhibited a similar
trend in the MTT assay: The absorbance of the HSP27 knockdown group
was significantly reduced at 72 and 96 h (Fig. 2D). MTT data supported that HSP27
expression was correlated with the proliferation of ESCC cells.
Subsequently, the Transwell assay was used to evaluate the invasion
ability of KYSE450 and KYSE150 cells with HSP27 knockdown. Compared
with the control groups, the number of cells invading through the
cell membrane in the HSP27 knockdown group significantly decreased
after 48 h (P<0.05, Fig. 2E and
F), indicating that HSP27 promoted the invasiveness of ESCC
cells. As shown by the wound healing assay, the migration of
KYSE450 and KYSE150 cells was reduced by HSP27 knockdown
(P<0.05, Fig. 2G and H).
Therefore, HSP27 can accelerate the growth and increase the
motility of ESCC cells.
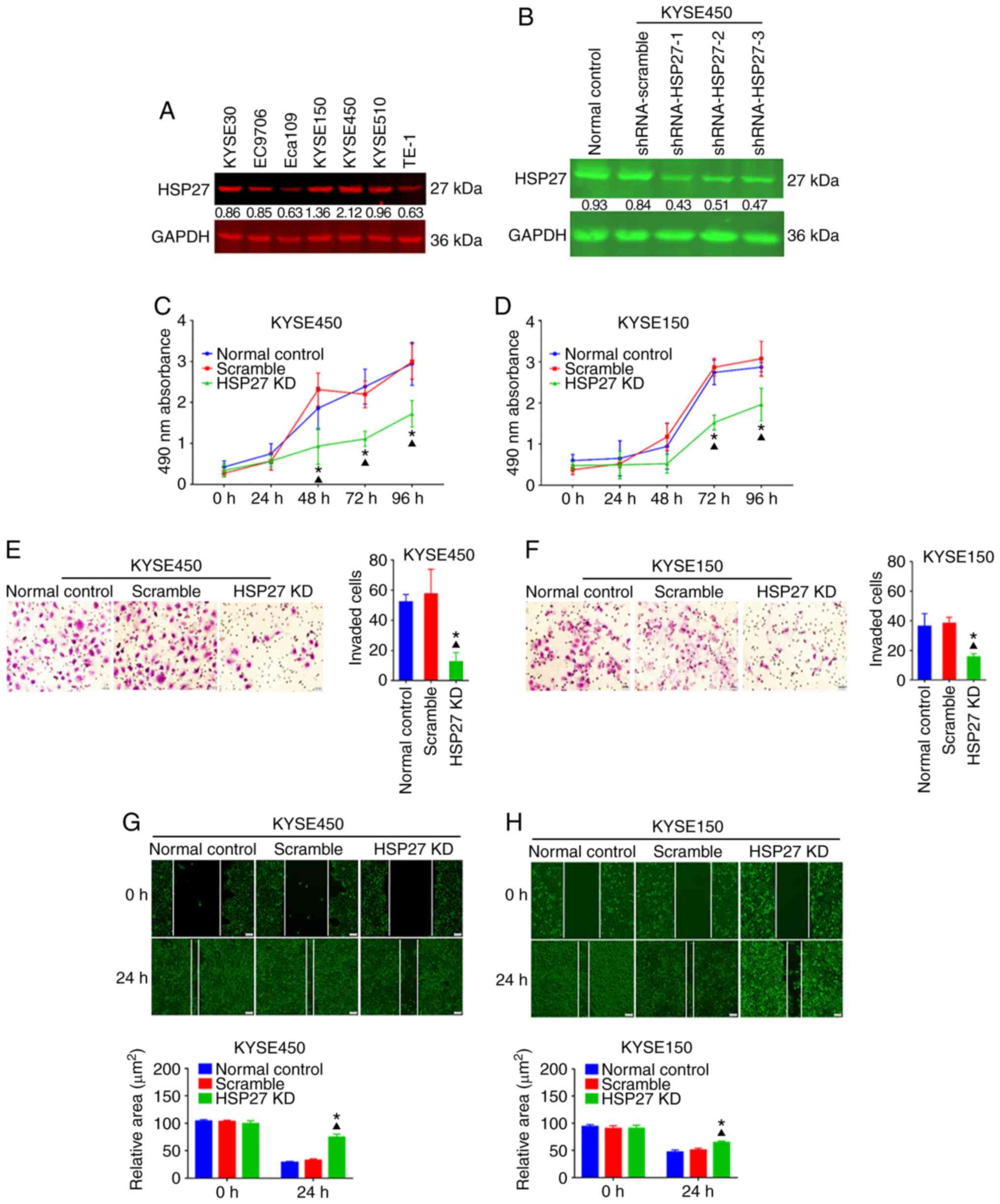 | Figure 2.HSP27 promotes the proliferation,
invasion and migration of ESCC cells. (A) Basal level of HSP27
expression in a panel of ESCC cell lines, including KYSE30,
KYSE150, KYSE450, KYSE510, TE-1, Eca109 and EC9706, as detected by
western blotting. (B) Detection of knockdown efficiency of HSP27
shRNA vectors in KYSE450 cells by western blotting. GAPDH was used
as an internal loading control. (C and D) MTT assay of
proliferation after knockdown of HSP27 in KYSE450 and KYSE150
cells. (E and F) Changes in invasion, detected by the Transwell
assay, in KYSE450 and KYSE150 cells after knockdown of HSP27
(magnification, ×100). (G and H) Variations in the migratory
activity of KYSE450 and KYSE150 cells analyzed using the wound
healing assay after knocking down HSP27 (magnification, ×100).
*P<0.05 compared with the normal control group and
▲P<0.05 compared with the shRNA-scramble control
group. ESCC, esophageal squamous cell carcinoma; HSP, heat shock
protein; KD, knockdown. |
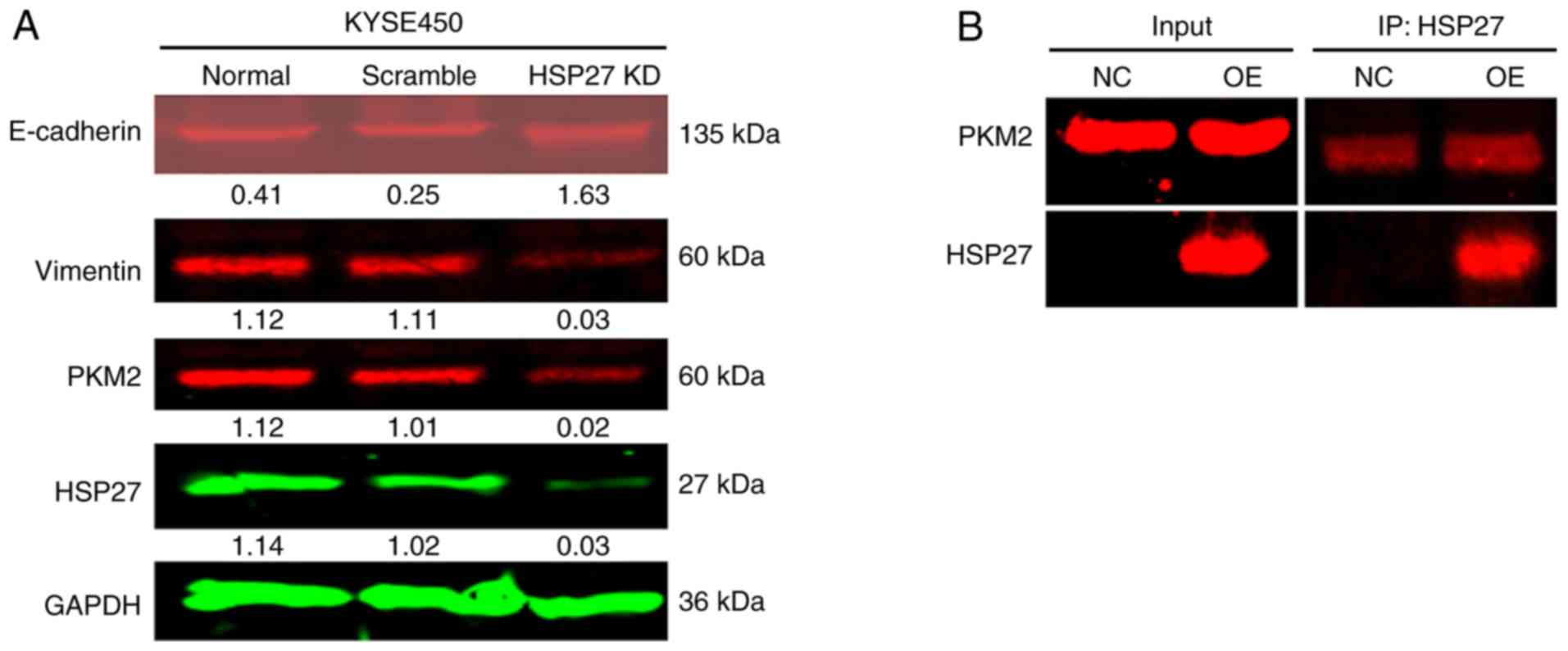
HSP27 promotes ESCC cell function by
regulating PKM2
To understand whether HSP27 enhanced the growth and
motility of ESCC cells via PKM2, the expression of PKM2 and other
biomarkers was detected by immunoblotting. The expression of PKM2
and vimentin was reduced after silencing HSP27; conversely,
E-cadherin expression was markedly increased (Fig. 3A). To determine whether there is a
direct interaction between PKM2 and HSP27, Co-IP was conducted. The
results of Co-IP revealed no direct interaction between HSP27 and
PKM2 (Fig. 3B).
HSP27 regulates the expression of PKM2
by interacting with SUMO2/3
The molecular mechanism through which HSP27
regulates PKM2 expression was further investigated. Lv-packaged
SUMO2/3 shRNAs were transfected into KYSE450 and KYSE150 cells. The
results of western blotting demonstrated that SUMO2/3 was
successfully knocked down, so the cells were used for subsequent
studies (Fig. 4A and B).
Subsequently, immunoblotting was used to detect alterations in
HSP27, PKM2 and EMT-related molecules. The expression of HSP27,
PKM2 and the EMT-related biomarker vimentin was significantly
decreased, whereas the expression of the EMT-related biomarker
E-cadherin was increased in both KYSE450 and KYSE150 knockdown
cells compared to control cells (Fig.
4C-F). Next, the Co-IP results revealed that HSP27 was capable
of directly binding with SUMO 2/3 (Fig.
4D and E). To further verify the association between SUMO2/3
and HSP27, immunofluorescence was used to detect the subcellular
localization of HSP27 and SUMO2/3. The laser confocal microscopy
images demonstrated that SUMO2/3 and HSP27 were co-localized in the
cytoplasm (Fig. 4F). Taken
together, these data demonstrated that HSP27 directly interacts
with SUMO2/3, thereby regulating PKM2 and promoting the motility of
ESCC cells through the EMT pathway.
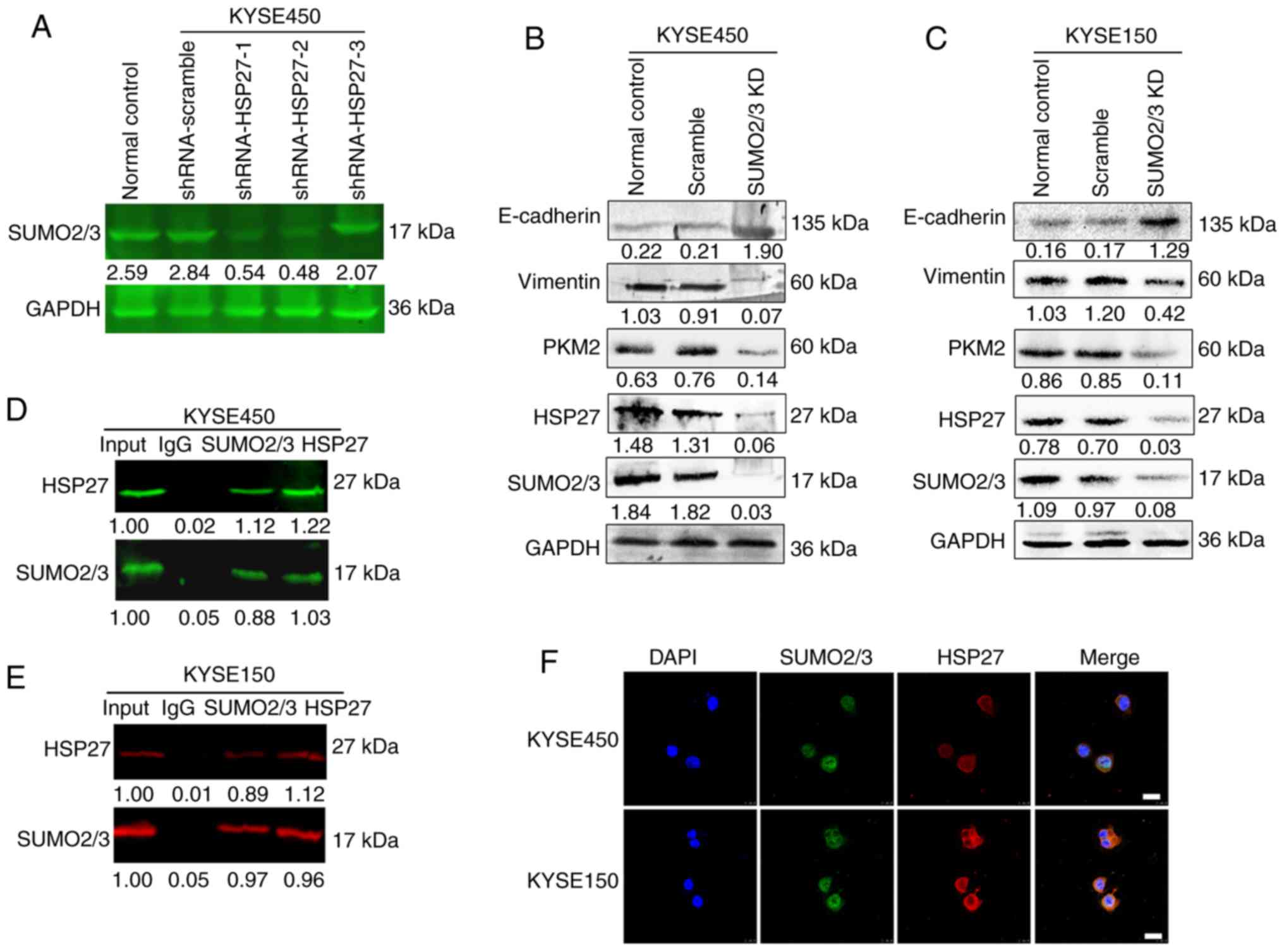 | Figure 4.HSP27 regulates PKM2 expression
through SUMO2/3 in KYSE450 and KYSE150 cells. (A) Detection of the
knockdown efficiency of SUMO2/3 shRNA vectors in KYSE450 cells by
western blotting. GAPDH, internal loading control. (B and C)
Detection of HSP27, PKM2, vimentin and E-cadherin in KYSE450 and
KYSE150 cells after SUMO2/3 silencing by western blotting. (D and
E) Interaction between HSP27 and SUMO2/3, as shown by
co-immunoprecipitation assay. (F) Immunofluorescence of nuclei
(DAPI, blue), SUMO2/3 (green) and HSP27 (red) in KYSE450 and
KYSE150 cells. HSP, heat shock protein; PKM2, pyruvate kinase
isoenzyme 2; SUMO2/3, small ubiquitin-related modified protein 2/3;
KD, knockdown. |
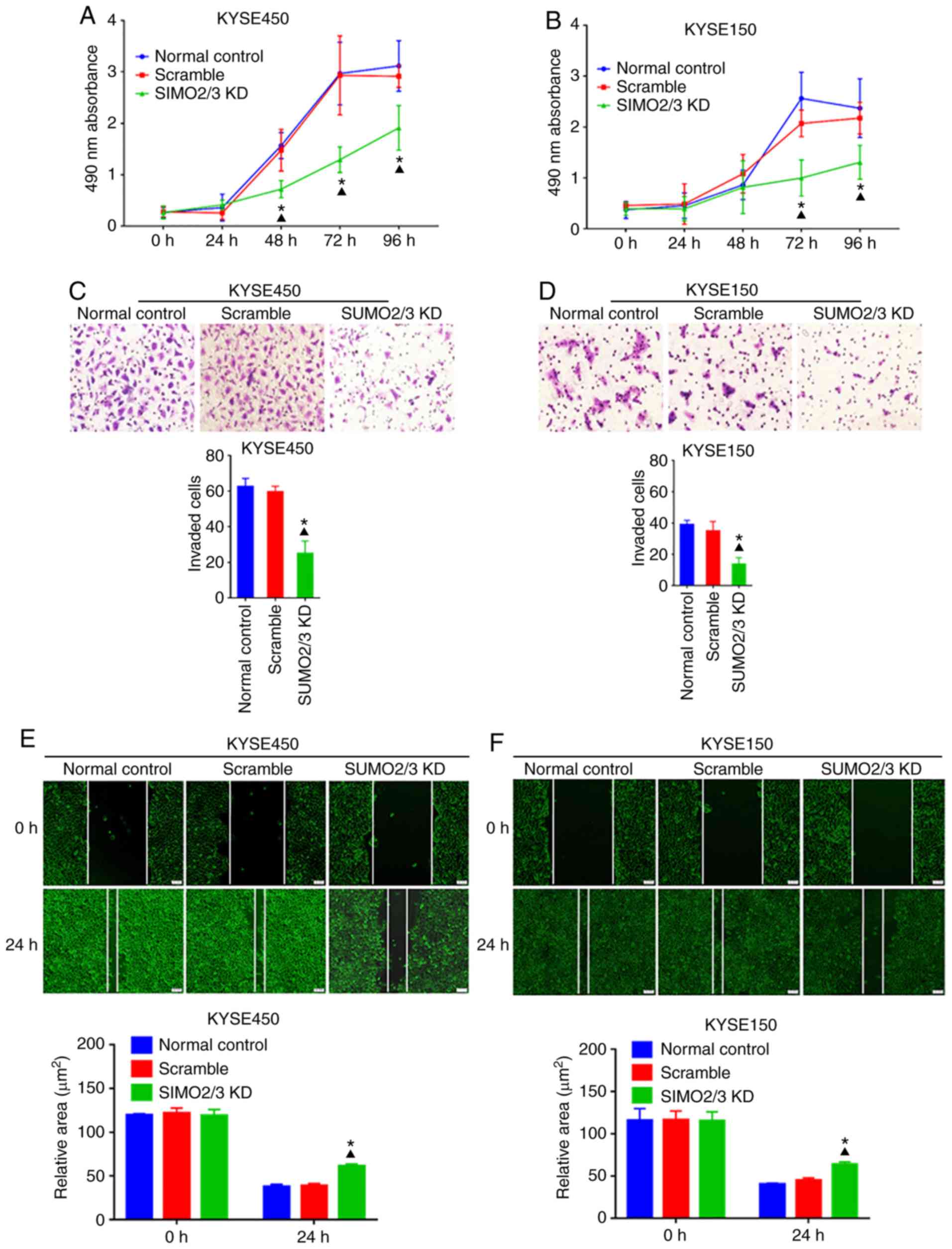
SUMO2/3 promotes the proliferation and
motility of ESCC cells
To elucidate the effects of SUMO2/3 on the malignant
behavior of ESCC cells, MTT, wound healing and Transwell assays
were performed to assess the proliferation and motility of KYSE450
and KYSE150 cells following SUMO2/3 knockdown. The data suggested
that the growth and motility of ESCC cells were significantly
suppressed by knocking down SUMO2/3 (Fig. 5), which supported our hypothesis
that SUMOylation of HSP27 regulates the expression of PKM2,
promoting the infiltration and metastasis of ESCC cells through the
EMT pathway.
Discussion
Accumulating evidence has revealed that HSPs, as
molecular chaperones, play a pivotal role in the modulation of
protein conformation and stability, and in carcinogenesis. As an
important member of the HSP family, HSP27 has been reported to be
highly expressed in different human cancers, including liver, lung
and breast cancer, and is involved in drug resistance, cell growth,
cell apoptosis, tumorigenesis and metastasis, as well as other
functions (22).
We previously demonstrated using two-dimensional gel
electrophoresis that 15 proteins, including HSP27 and PKM2, were
highly expressed in Kazakh patients with ESCC (7). In addition, our previous study
revealed that the PKM2/STAT3 pathway promoted ESCC cell
proliferation and motility through regulating EMT (24). However, the regulatory association
between HSP27 and PKM2 remains largely unknown. HSPs are important
stress response proteins that protect the stability of the
intracellular environment. Whether HSP27 participates in anaerobic
glycolysis has not been extensively investigated. High expression
of HSP90 in myocytes may increase cell uptake and utilization of
glucose, and the function of HSP90 depends on the supply of ATP
(25), strongly suggesting that
HSP90 may directly affect cell glucose metabolism. In healthy
cells, upregulation of HSP62 may induce the synthesis of pyruvate
kinase, increase the activity of pyruvate kinase, and increase the
cell content of ATP, indicating that high expression of HSP62 can
accelerate glycolysis in healthy cells (26). It was recently reported that HSP
proteins are also involved in the Warburg effect of cancer cells.
HSP90 is significantly overexpressed in HCC tissues and can
directly bind and phosphorylate threonine 328 of PKM2, stabilizing
PKM2 dimers in the cytoplasm and promoting the Warburg effect
(27).
In addition to HSP90, HSP40 has also been shown to
interact with PKM2. The binding of HSP40 and PKM2 can stabilize the
dimer of PKM2, and silencing HSP40 was able to suppress the growth
of tumor cells (29). These
findings strongly suggest a direct interaction between PKM2 and HSP
proteins. HSP27 and HSP90 share highly conserved domains.
Therefore, it was hypothesized that HSP27 may also interact with
PKM2 directly in Kazakh patients with ESCC. To test this
hypothesis, the levels of HSP27 and PKM2 were assessed by IHC in
ESCC tissue samples. The staining results demonstrated that the
expression of HSP27 was markedly associated with T stage, lymph
node metastasis and overall survival. Moreover, the expression
levels of HSP27 and PKM2 were positively correlated in Kazakh ESCC
tissues. However, the correlation between HSP27 and SUMO2/3 was not
determined in the present study, mainly due to the fact that the
expression of SUMO2/3 was not detected at the clinical tissue
level. Further verification at the ESCC cell level demonstrated
that knockdown of HSP27 inhibited the function of ESCC cells,
decreased the expression of PKM2 and vimentin, and increased the
expression of E-cadherin. These results indicate that HSP27
regulates the PKM2 pathway to promote ESCC progression. However,
the results of Co-IP revealed that HSP27 does not bind directly
with PKM2, indicating that HSP27 and PKM2 do not directly interact.
Moreover, a previous study by Xu et al (28) on breast cancer reported that
knockdown of HSP27 can suppress EMT, with decreased Snail and
vimentin and increased E-cadherin expression. These data suggested
that HSP27 can promote EMT in breast cancer. Despite the fact that
ESCC and breast cancer pathologically originate from different
tissues, it may be inferred that HSP27 acts in a similar manner in
the two cancers by regulating EMT. EMT variations following
knockdown of HSP27 in ESCC cells will be further investigated in
future studies.
SUMOylation, an important post-translational
modification, is mediated by SUMO2/3 (18). Liu et al (17) found that SUMO2/3 promotes the
transformation of chronic hepatitis B to liver cancer by
stabilizing p65 in the cytoplasm and deactivating NF-κB. Ge et
al (21) reported that SUMO2/3
in HCC cells protects HSP27 from degradation by binding to the GKHE
sequence on the HSP27 protein, thus promoting the motility of tumor
cells. However, the SUMOylation of HSP27 has rarely been reported
in ESCC. The data obtained herein demonstrated that SUMOylation of
HSP27 was able to modulate the expression PKM2 to promote the
progression of ESCC.
In the present study, HSP27 knockdown was shown to
downregulate PKM2 in ESCC cells; in addition, the proliferation and
invasion of ESCC cells were also markedly suppressed after HSP27
silencing. These data strongly suggest that HSP27 promotes ESCC
cell function by regulating PKM2. Mechanistically, the promoting
effect of HSP27 on PKM2 level may be mediated by SUMO2/3, which can
interact with HSP27. As shown in the present study, silencing of
SUMO2/3 may lead to the downregulation of HSP27, along with a
decrease in PKM2. From these data, it may be inferred that SUMO2/3
can prevent the HSP27 protein from degradation. These observations
were in agreement with a previous study reporting that SUMO2/3 was
required for the promotion of proliferation and invasion of liver
cancer cells by HSP27 (21). Of
note, the data of the present study strongly support that
SUMOylation stabilizes HSP27 and protects it from degradation,
which was also reported previously by Ge et al (21). However, as the expression of SUMO2/3
was not detected in the present study, no conclusions can be drawn
on the correlation between SUMO2/3 and HSP27, which should be
further investigated in future studies.
In conclusion, HSP27 was found to be significantly
associated with T stage, lymph node metastasis and poor prognosis
in ESCC, revealing a novel regulatory mechanism of PKM2, mediated
by SUMOylation of HSP27, which is implicated in the oncogenesis of
ESCC.
Acknowledgements
Not applicable.
Funding
The present study was funded in part by the Natural
Science Foundation of China (grant nos. 81960527, U1603284 and
81860511) and in part by Key Research and Development Project of
the Xinjiang Uygur Autonamous Region (grant no. 2020B03003-1) and
Science and Technology Projects of Xinjiang Uygur Autonomous Region
(grant no. 2018E02067) from TianshanXuesong Project of the Xinjiang
Uygur Autonomous Region (grant no. 2018XS19).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XZ and TL performed all the experiments described.
STZ drafted and revised the manuscript. QL analyzed and interpreted
the patient data obtained from immunohistochemistry. TXS and XJH
immunoscored all the sections. QQZ and LFY contributed to the
statistical analysis. XML superised the study. All the authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
The protocol of the present study was approved by
the Medical Ethics Committee of the First Affiliated Hospital of
Xinjiang Medical University, and informed consent was obtained from
each participant involved regarding the use of their tissues for
research purposes.
Patient consent or publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Liu J, Yin P, Liu S, Cai Y, You J,
Zeng X, Wang L and Zhou M: The disease burden of malignant tumor in
China, 1990 and 2010. Zhonghua Yu Fang Yi Xue Za Zhi. 49:309–314.
2015.(In Chinese). PubMed/NCBI
|
|
3
|
Zheng S, Vuitton L, Sheyhidin I, Vuitton
DA, Zhang Y and Lu X: Northwestern China: A place to learn more on
oesophageal cancer. Part one: Behavioural and environmental risk
factors. Eur J Gastroenterol Hepatol. 22:917–925. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu XM, Monnier-Benoit S, Mo LZ, Xu SY,
Prétet JL, Liu Z, Vuitton DA and Mougin C: Human papillomavirus in
esophageal squamous cell carcinoma of the high-risk Kazakh ethnic
group in Xinjiang, China. Eur J Surg Oncol. 34:765–770. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu F, Ma F, Wang Y, Hao L, Zeng H, Jia C,
Wang Y, Liu P, Ong IM, Li B, et al: PKM2 methylation by CARM1
activates aerobic glycolysis to promote tumorigenesis. Nat Cell
Biol. 19:1358–1370. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong N, Ojo D, Yan J and Tang D: PKM2
contributes to cancer metabolism. Cancer Lett. 356((2 Pt A)):
184–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu Z, Feng JG, Tuersun A, Liu T, Liu H,
Liu Q, Zheng ST, Huang CG, Lv GD, Sheyhidin I and Lu XM: Proteomic
identification of differentially-expressed proteins in esophageal
cancer in three ethnic groups in Xinjiang. Mol Biol Rep.
38:3261–3269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Q, Liang M, Liu T, Vuitton L, Zheng S,
Gao X, Lu M, Li X, Sheyhidin I and Lu X: M2 isoform of pyruvate
kinase (PKM2) is upregulated in Kazakh's ESCC and promotes
proliferation and migration of ESCC cells. Tumour Biol.
37:2665–2672. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Abdel-Haleem AM, Lewis NE, Jamshidi N,
Mineta K, Gao X and Gojobori T: The emerging facets of
non-cancerous warburg effect. Front Endocrinol (Lausanne).
8:2792017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu J, Liu T, Rios Z, Mei Q, Lin X and Cao
S: Heat shock proteins and cancer. Trends Pharmacol Sci.
38:226–256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saini J and Sharma PK: Clinical,
prognostic and therapeutic significance of heat shock proteins in
cancer. Curr Drug Targets. 19:1478–1490. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zininga T, Ramatsui L and Shonhai A: Heat
shock proteins as immunomodulants. Molecules. 23:28462018.
View Article : Google Scholar
|
|
13
|
Han L, Jiang Y, Han D and Tan W: Hsp27
regulates epithelial mesenchymal transition, metastasis and
proliferation in colorectal carcinoma. Oncol Lett. 16:5309–5316.
2018.PubMed/NCBI
|
|
14
|
Zhang Y, Feng Z, Wang W, Dong J, Gong X,
Pu H and Chen X: Expression of heat shock protein-27 (Hsp27) and
P38MAPK in esophageal squamous cell carcinoma. Med Sci Monit.
23:5246–5253. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luz CC, Noguti J, Borges de Araujo L,
Gianni MS, Simao Gomes T and Ricardo AN: Hsp27 and Hsp70 expression
in esophageal squamous. Asian Pac J Cancer Prev. 18:789–794.
2017.PubMed/NCBI
|
|
16
|
Lee MS, Lee J, Lee S, Yoo SM, Kim JH, Kim
WT, Kim WJ and Park J: Clinical, prognostic, and therapeutic
significance of heat shock protein 27 in bladder cancer.
Oncotarget. 9:7961–7974. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu J, Sha M, Wang Q, Ma Y, Geng X, Gao Y,
Feng L and Shen Y and Shen Y: Small ubiquitin-related modifier 2/3
interacts with p65 and stabilizes it in the cytoplasm in
HBV-associated hepatocellular carcinoma. BMC Cancer. 15:6752015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eifler K and Vertegaal ACO:
SUMOylation-mediated regulation of cell cycle progression and
cancer. Trends Biochem Sci. 40:779–793. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Princz A and Tavernarakis N: SUMOylation
in Neurodegenerative Diseases. Gerontology. 66:122–130. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dasso M: Biochemistry: Rear view of an
enzyme. Nature. 497:576–577. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ge H, Du J, Xu J, Meng X, Tian J, Yang J
and Liang H: SUMOylation of HSP27 by small ubiquitin-like modifier
2/3 promotes proliferation and invasion of hepatocellular carcinoma
cells. Cancer Biol Ther. 18:552–559. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pellat A and Coriat R: Well differentiated
grade 3 neuroendocrine tumors of the digestive tract: A narrative
review. J Clin Med. 9:E16772020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang N, Ren Y, Wang Y, Zhao L, Wang B, Ma
N, Gao Z and Cao B: LRG1 suppresses migration and invasion of
esophageal squamous cell carcinoma by modulating epithelial to
mesenchymal transition. J Cancer. 11:1486–1494. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma R, Liu Q, Zheng S, Liu T, Tan D and Lu
X: PKM2-regulated STAT3 promotes esophageal squamous cell carcinoma
progression via TGF-β1-induced EMT. J Cell Biochem.
12–Feb;2019.(Epub ahead of print). View Article : Google Scholar
|
|
25
|
Lang BJ, Guerrero-Gimenez ME, Prince TL,
Ackerman A, Bonorino C and Calderwood SK: Heat shock proteins are
essential components in transformation and tumor progression:
Cancer cell intrinsic pathways and beyond. Int J Mol Sci.
20:45072019. View Article : Google Scholar
|
|
26
|
Marsden M, Nickells RW, Kapoor M and
Browder LW: The induction of pyruvate kinase synthesis by heat
shock in Xenopus laevis embryos. Dev Genet. 14:51–57. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang Z, Yang C, Xiao L, Liao X, Lan A,
Wang X, Guo R, Chen P, Hu C and Feng J: Novel insights into the
role of HSP90 in cytoprotection of H2S against chemical
hypoxia-induced injury in H9c2 cardiac myocytes. Int J Mol Med.
28:397–403. 2011.PubMed/NCBI
|
|
28
|
Xu Q, Tu J, Dou C, Zhang J, Yang L, Liu X,
Lei K, Liu Z, Wang Y, Li L, et al: HSP90 promotes cell glycolysis,
proliferation and inhibits apoptosis by regulating PKM2 abundance
via Thr-328 phosphorylation in hepatocellular carcinoma. Mol
Cancer. 16:1782017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang L, Yu Z, Zhang T, Zhao X and Huang
G: HSP40 interacts with pyruvate kinase M2 and regulates glycolysis
and cell proliferation in tumor cells. PLoS One. 9:e929492014.
View Article : Google Scholar : PubMed/NCBI
|