Introduction
Lung cancer is the most common malignant tumor
worldwide, and its morbidity and mortality rank first among all
cancers (1). The most common
histological subtype is non-small cell lung cancer (NSCLC),
accounting for more than 80% of all lung cancer cases (2). Unfortunately, the majority of patients
with NSCLC are at an advanced stage at the time of initial
diagnosis, and have missed the opportunity to undergo surgery,
which results in a poor prognosis (3). Although platinum-based chemotherapy is
currently the standard first-line treatment for these patients,
some NSCLC patients are not sensitive to platinum-based
chemotherapy, with an overall response rate of only 30% (4,5). In
addition, most patients with NSCLC eventually develop resistance to
the chemotherapy, thus greatly limiting the clinical application of
chemotherapeutic drugs and leading to a worse prognosis. Therefore,
it is crucial to explore the molecular mechanism underlying the low
response rate of NSCLC to platinum-based chemotherapy.
Exosomes are nanoscale membrane vesicles secreted by
cells into the extracellular space to mediate intercellular
communication (6). Therefore,
exosomes are not only important carriers of bioactive substances,
but also crucial mediators for information exchange between cells
(7,8). It has been demonstrated that exosomes
can transmit biological information by transporting coated
contents, such as microRNAs (miRNAs), long non-coding RNAs
(lncRNAs) and proteins, and participate in the malignant processes
of NSCLC through various pathways (9,10).
miRNAs are a family of small non-coding RNAs that can alter gene
expression post-transcriptionally, thereby regulating tumor cell
proliferation, migration, invasion and metastasis, angiogenesis and
apoptosis as well as genomic instability (11,12).
Since miRNAs in exosomes have the ability to regulate gene
expression, are stable in the extracellular environment and display
strong resistance to degradation, they may be widely used as
diagnostic biomarkers of cancers and other diseases (13). More importantly, secreted exosomal
miRNAs may be taken up by recipient cells, and then regulate a
variety of biological processes such as proliferation,
differentiation and apoptosis by inhibiting the expression of
target genes (14–16).
In recent years, miRNAs in exosomes have attracted
considerable attention as the changes in the type and amount of
miRNAs in exosomes are associated with the resistance of NSCLC
cells to chemotherapeutic drugs (17). A recent study demonstrated that
miRNAs were differentially expressed in the exosomes of cisplatin
(CDDP)-resistant and CDDP-sensitive NSCLC cells. In CDDP-resistant
NSCLC cell lines, the amount of miR-100-5p was significantly
decreased in exosomes and was functionally involved in CDDP
resistance of NSCLC (18). It
should be noted that platinum stimulation may also change the
expression profiles of miRNAs, many of which are considered to be
responsible for drug resistance (19). However, few studies have focused on
the expression of miRNAs in exosomes stimulated by platinum drugs
and the effect of these miRNAs on the sensitivity of NSCLC to
platinum drugs. In the present study, microarray analysis was
performed to examine the differential expression profiles of
exosomal miRNAs in the NSCLC cell line A549 with or without CDDP
treatment, and miR-1273a was identified as a candidate miRNA
involved in the sensitivity of A549 cells to CDDP. In addition, the
expression of miR-1273a in plasma exosomes of patients with
advanced NSCLC before and after CDDP chemotherapy was further
evaluated to confirm the clinical significance.
Materials and methods
Cell culture and human specimens
A549 cells were purchased from the American Type
Culture Collection (ATCC) and grown in RPMI-1640 medium
supplemented with 10% FBS (Biological Industries) and 100 U/ml
penicillin-streptomycin at 37°C in an atmosphere of 95% air and 5%
CO2. Plasma samples were collected from 49 patients with
advanced NSCLC at Daping Hospital, Chongqing, China from January to
December 2018 and stored at −80°C until use. The average age of the
patients was 57.0±9.8, 32 (65.3%) were male and 17 (34.7%) were
female patients. All patients received standard first-line
platinum-based chemotherapy (Table
S1). To evaluate therapeutic outcomes, the patients were
divided into responder [complete response (CR) + partial response
(PR)] and non-responder [stable disease (SD) + progressive disease
(PD)] groups according to the Response Evaluation Criteria In Solid
Tumors (version 1.1) (20). The
present study was conducted in accordance with the principles
outlined in the Declaration of Helsinki, and was approved by the
Ethics Committee of Daping Hospital (ratification no 2018113).
Isolation of exosomes
When the cells had grown to reach a confluence of
~70%, the culture medium was replaced with fresh RPMI-1640
containing 10% Exosome-Depleted FBS (VivaCell). After 48 h of
incubation, the supernatant was collected and the exosomes were
extracted according to the following steps. The supernatant was
centrifuged at 10,000 × g for 30 min at 4°C and filtered by a
0.22-mm filter. Subsequently, the exosomes were precipitated twice
by ultracentrifugation at 100,000 × g for 70 min at 4°C. The
isolated exosomes were added into the culture medium of the cells
for further analysis. To isolate exosomes from patient plasma,
plasma samples were centrifugated at 10,000 × g for 20 min at 4°C,
and the plasma exosomes were isolated by the Total Exosome
Isolation Kit (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The collected
exosomes were resuspended in sterile PBS and stored at −80°C until
use.
Transmission electron microscopy (TEM)
and nanoparticle-tracking analysis (NTA)
Exosome sample were added to copper grids at room
temperature for 5 min, stained with 2% uranyl acetate solution for
1 min, dried for 20 min at room temperature, and then observed
using TEM (JEOL, Ltd.). NTA was performed by a nanoparticle tracing
assay (ZetaVIEW S/N 17–310; Particle Metrix GmbH) and analyzed
using ZetaVIEW 8.04.02 software (Particle Merix GmbH).
Western blotting
Cells and exosomes were lysed in RIPA Lysis Buffer
(Beyotime Institute of Biotechnology). A total of 20 µg protein was
then separated by 10% SDS-PAGE and then transferred onto a PVDF
membrane according to methods described previously (21). Antibodies against CD63 (dilution
1:500; cat. no. ab59478), CD9 (dilution 1:500; cat. no. ab58989),
Alix (dilution 1:500; cat. no. ab117600), Calnexin (dilution 1:500;
cat. no. ab92573) and β-actin (dilution 1:500; cat. no. ab8226)
were purchased from Abcam. Antibody against syndecan binding
protein (SDCBP; dilution 1:1,000; cat. no. 22399-1-AP) was
purchased from Proteintech (Sanying Biotechnology, Inc.). The band
intensity of the western blot images was quantified using Image Lab
software (version 1.2.0.12, Bio-Rad Laboratories, Inc.).
Cellular uptake of exosomes
Exosomes were stained with PKH26 and incubated with
A549 cells at 37°C for 4 h, and then the cells were fixed with 4%
paraformaldehyde for 15 min at room temperature. The nuclei were
stained with DAPI. For the control group, PKH26 was added to an
equal amount of PBS and then used to treat A549 cells. The
fluorescence-labelled exosomes were observed under a confocal
microscope (PerkinElmer, Inc.) at ×400 magnification.
Analysis of cell viability and
apoptosis
For cell viability analysis, the A549 cells were
seeded onto 96-well plates at a density of 5,000 cells per well and
incubated for 12 h. The cells were then treated with the indicated
concentration of CDDP for an additional 48 h, and the cell
viability was determined using Cell Counting Kit-8 (CCK-8) assay
(Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. For apoptosis analysis, the cells were
seeded onto 6-well plates at a density of 2×105 cells
per well and cultured for 24 h. Subsequently, the cells were seeded
into 6-well plates overnight and then treated with 2 µg/ml CDDP.
After 36 h of CDDP treatment, the apoptotic cells were detected
with the Annexin V-FITC kit (CalbioChem) and quantified using flow
cytometry (Gallios, Beckman Coulter).
Microarray analysis of exosomal
RNA
Microarray analysis was performed to detect
differentially expressed miRNAs using Affymetrix GeneChip miRNA 4.0
array (Affymetrix, Thermo Fisher Scientific, Inc.).
GeneChip® Scanner 3000 7G (Affymetrix, Thermo Fisher
Scientific, Inc.) was used to scan the arrays. CEL files obtained
from the Affymetrix GeneChip were analyzed by the Gminix-Cloud
Biotechnology Information (GCBI) platform (http://www.gcbi.com.cn) using Affymetrix default
analysis settings and global scaling as the normalization method.
SAM method was used to analyze the difference (22). According to the filter condition
|Fold Change|>2, the final difference results were obtained.
Transfection experiment
When the cells reached 50% confluence, 20 nM
miR-1273a mimic or inhibitors or negative control (Guangzhou
RiboBio Co., Ltd.) were mixed with 10 µl of HiPerfect transfection
reagent (Qiagen GmbH), and then transfected into A549 cells
according to the HiPerfect transfection reagent operating
instructions. The sequence of miR-1273a mimic sense was
GGGCGACAAAGCAAGACUCUUUCUU and antisense was
AAGAAAGAGUUUUGCUUUGTCGCCC. The sequence of miR-1273a inhibitor was
CAGUACUUUUGUGUAGUACAA. The sequence of mimics-control was as
follows: Sense, UUUGUACUACACAAAAGUACUG and antisense,
CAGUACUUUUGUGUAGUACAAA. The sequence of inhibitor-control was
CAGUACUUUUGUGUAGUACAA. After 48 h of transfection, the cells were
used for the subsequent experiment.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from exosomes or cultured
cells using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). cDNA was synthesized from 200 ng total RNA and
amplified by RT-qPCR using TB Green PremixExTaq II (Takara
Biotechnology Co., Ltd.). The thermocycling condition for miRNA
consisted of 95°C for 20 sec followed by 40 cycles of 10 sec at
95°C, 20 sec at 60°C and 10 sec at 70°C; for mRNA: 95°C for 5 min
followed by 39 cycles of 10 sec at 95°C, 15 sec at 60°C and 20 sec
at 72°C. Cel-miR-39 was used as an external control in exosomes,
and U6 was used as an internal reference in cells. β-actin was used
as the endogenous control to normalize the expression of SDCBP. All
the primers were synthesized by Guangzhou RiboBio Co., Ltd. The
relative expression level of miRNAs and mRNAs were calculated by
the 2−ΔΔCt method (23).
The expression level of miRNAs in plasma exosomes was calculated
using the log2 (2−ΔCtx1010)
formula, as previously described (24).
Quantitation of SDCBP by ELISA
Plasma was collected from NSCLC patients and
assessed by ELISA using the SDCBP ELISA kit (catalog no. MK4079A,
Meike Biotechnology Co., Ltd.), according to the manufacturer's
instructions.
Statistical analysis
Statistical analyses were performed by GraphPad
Prism software 7.0 (GraphPad Software, Inc.) or statistical package
SPSS (IBM SPSS Statistics for Windows, version 19.0). Parametric
data are expressed as the means ± standard deviation (SD).
Difference analyses between different cell groups were compared
with Student's t-test. Difference analyses between different
patient groups were compared with Mann-Whitney U test. The
expression levels of miR-1273a before and after chemotherapy were
compared with Wilcoxon paired non-parametric test. Correlation was
analyzed using Pearson correlation. P<0.05 was considered to
indicate statistically significant differences.
Results
Identification of isolated exosomes
from CDDP-treated A549 cells
A549 cells were treated with 2 µg/ml CDDP for 48 h
based on dose-response evaluation (Appendix S1 and Fig. S1), and the exosomes were then
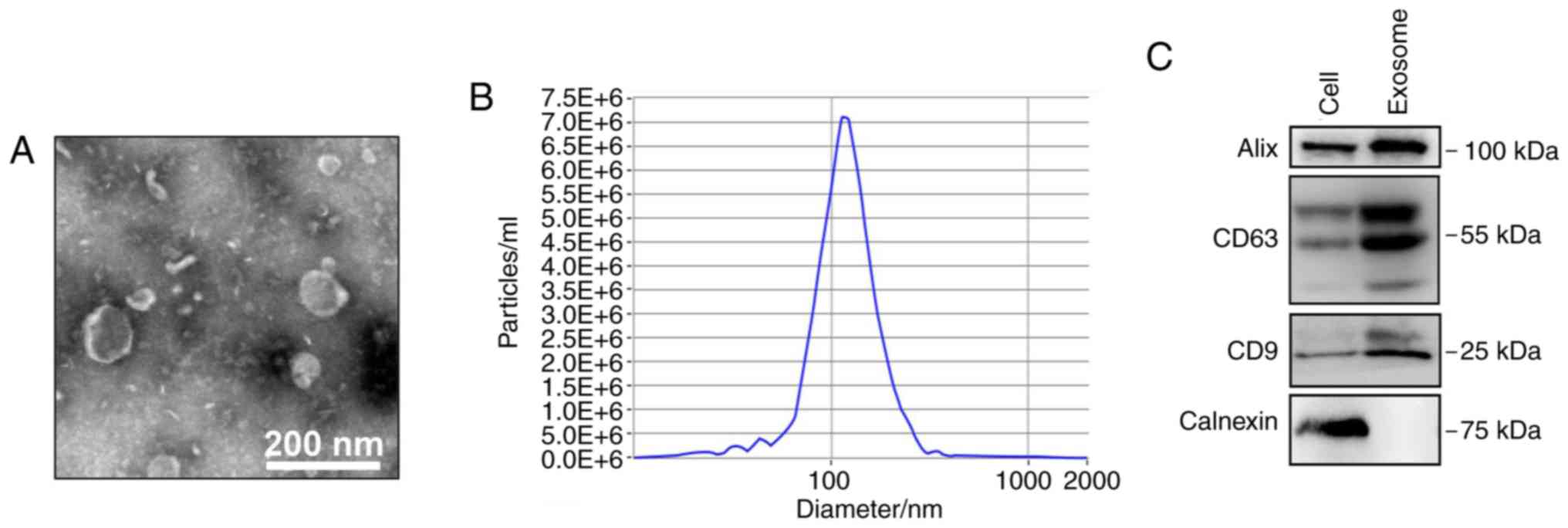
isolated from the cell culture medium. To ensure successful
isolation of exosomes from A549 cells, the collected exosomes were
observed by TEM and analyzed by nanoparticle tracking and western
blotting. The expected size range of exosomes and bilayer
membrane-shaped morphology were readily observed using TEM
(Fig. 1A). NTA demonstrated that
the mean particle diameter was 30–150 nm (Fig. 1B). Western blotting demonstrated
that the exosomes were positive for the exosomal markers CD9, CD63
and ALIX, but negative for calnexin (an endoplasmic reticulum
protein) (Fig. 1C).
Exosomes secreted from CDDP-treated
cells reduce the sensitivity of A549 cells to CDDP
Before examining the effects of exosomes isolated
from CDDP-treated NSCLC cells on NSCLC sensitivity to CDDP
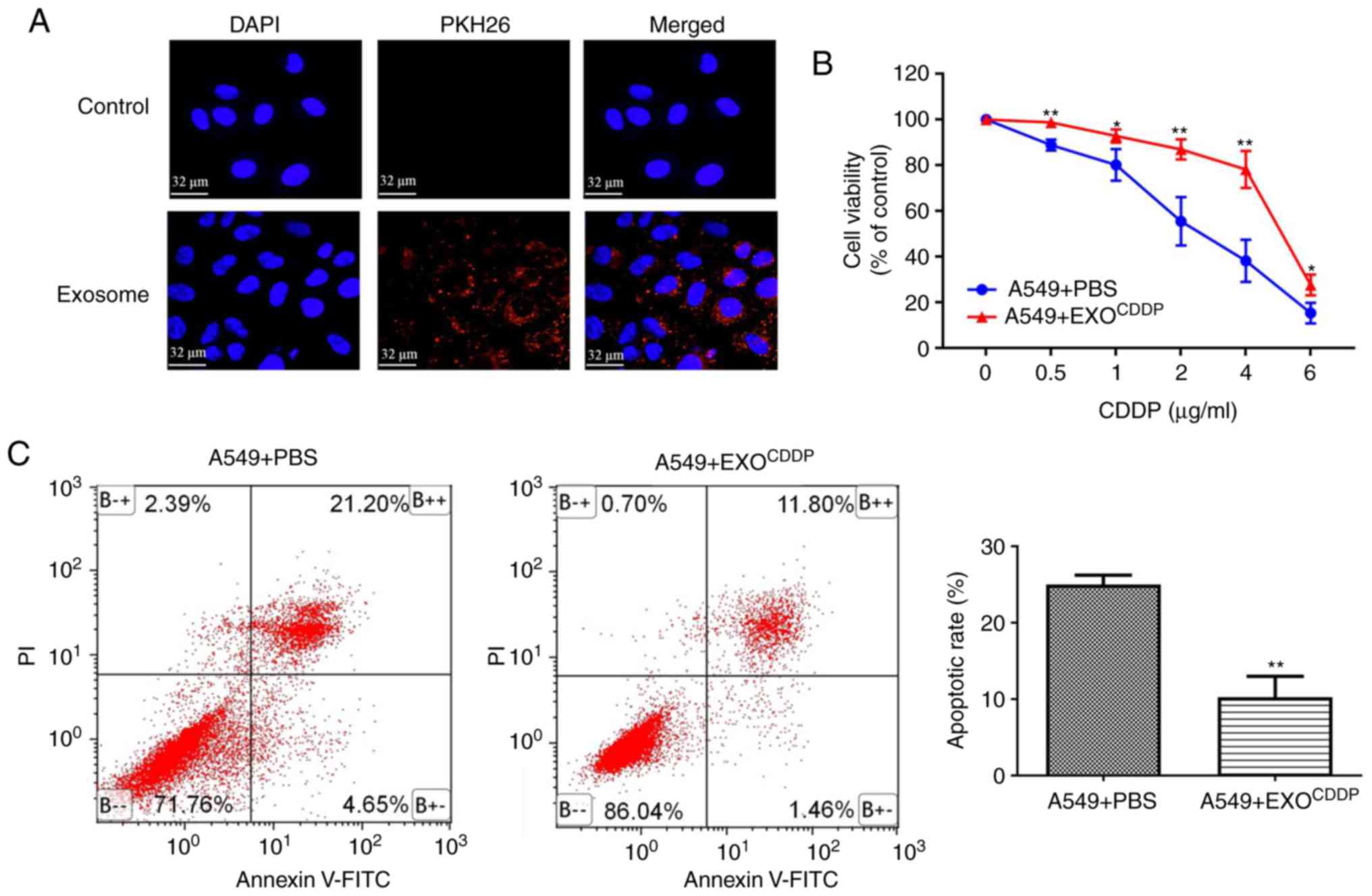
treatment, it was investigated whether exosomes can be taken up by
A549 cells. Compared with control cells, following incubation with
PKH26-stained exosomes for 4 h, a large number of PKH26
fluorescence-labeled exosomes appeared inside the A549 cells
(Fig. 2A), suggesting that the
cells can take up exosomes. In order to investigate the effects of
exosomes secreted by CDDP-treated cells on the sensitivity of NSCLC
cells to cisplatin treatment, A549 cells were co-cultured with PBS
(used as control), EXOCDDP (exosomes isolated from the
culture medium of CDDP-treated cells) for 48 h. Compared with the
control group, the cell viability of the EXOCDDP group
was significantly enhanced following treatment with various
concentrations of CDDP (Fig. 2B).
Consistently with these findings, the apoptotic rate of the
EXOCDDP group was significantly reduced compared with
that of the control group (25.03±1.24 vs. 10.23±2.76%,
respectively; P<0.01; Fig. 2C).
These results suggest that the transmission of exosomes derived
from CDDP-stimulated cells may reduce the sensitivity of A549 cells
to CDDP.
Changes in microRNA expression
profiles in exosomes derived from CDDP-treated A549 cells
Numerous studies have demonstrated that miRNAs play
an important role in exosome-mediated chemoresistance. Therefore,
miRNAs may be involved in EXOCDDP-mediated CDDP
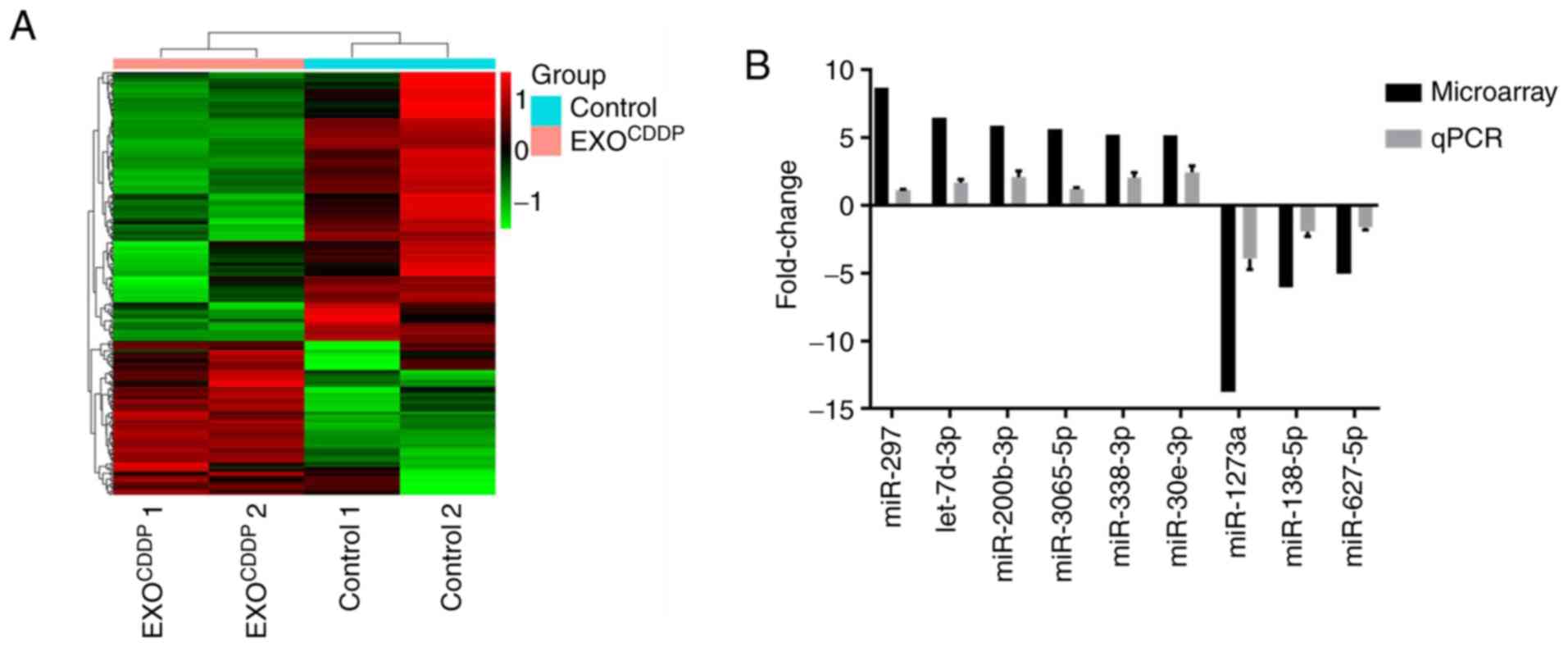
resistance. Microarray analysis was performed to examine the
expression profiles of miRNAs in EXOCDDP. Exosomes
derived from A549 cells (EXO) were used as control. Compared with
the control group, a total of 276 miRNAs in EXOCDDP were
altered >2-fold (Fig. 3A). In
particular, there was a >5-fold difference in the expression of
9 miRNAs between the two groups. Among those, the expression of
miR-297, let-7d-3p, miR-200b-3p, miR-3065-5p, miR-338-3p and
miR-30e-3p was upregulated, whereas the expression of miR-1273a,
miR-138-5p, and miR-627-5p was downregulated (Table I). Next, the expression levels of
the above mentioned 9 miRNAs were further confirmed by RT-qPCR
analysis. As shown in Fig. 3B,
compared with control exosomes, the RT-qPCR results on the changes
in the 9 miRNAs in EXOCDDP were consistent with the
microarray data. Since the expression difference of miR-1273a was
the most prominent among these miRNAs, miR-1273a was selected for
further experiments.
 | Table I.Top 9 miRNAs with differential
expression (fold change >5) between the EXO and
EXOCDDP groups. |
Table I.
Top 9 miRNAs with differential
expression (fold change >5) between the EXO and
EXOCDDP groups.
| Upregulation | Fold | Rank | Downregulation | Fold | Rank |
|---|
| hsa-miR-297 | 8.69 | 1 | hsa-miR-1273a | 13.76 | 1 |
| hsa-let-7d-3p | 6.49 | 2 | hsa-miR-138-5p | 6.05 | 2 |
|
hsa-miR-200b-3p | 5.90 | 3 | hsa-miR-627-5p | 5.03 | 3 |
|
hsa-miR-3065-5p | 5.65 | 4 |
|
|
|
| hsa-miR-338-3p | 5.21 | 5 |
|
|
|
| hsa-miR-30e-3p | 5.20 | 6 |
|
|
|
Overexpression of miR-1273a enhances
the sensitivity of A549 cells to CDDP
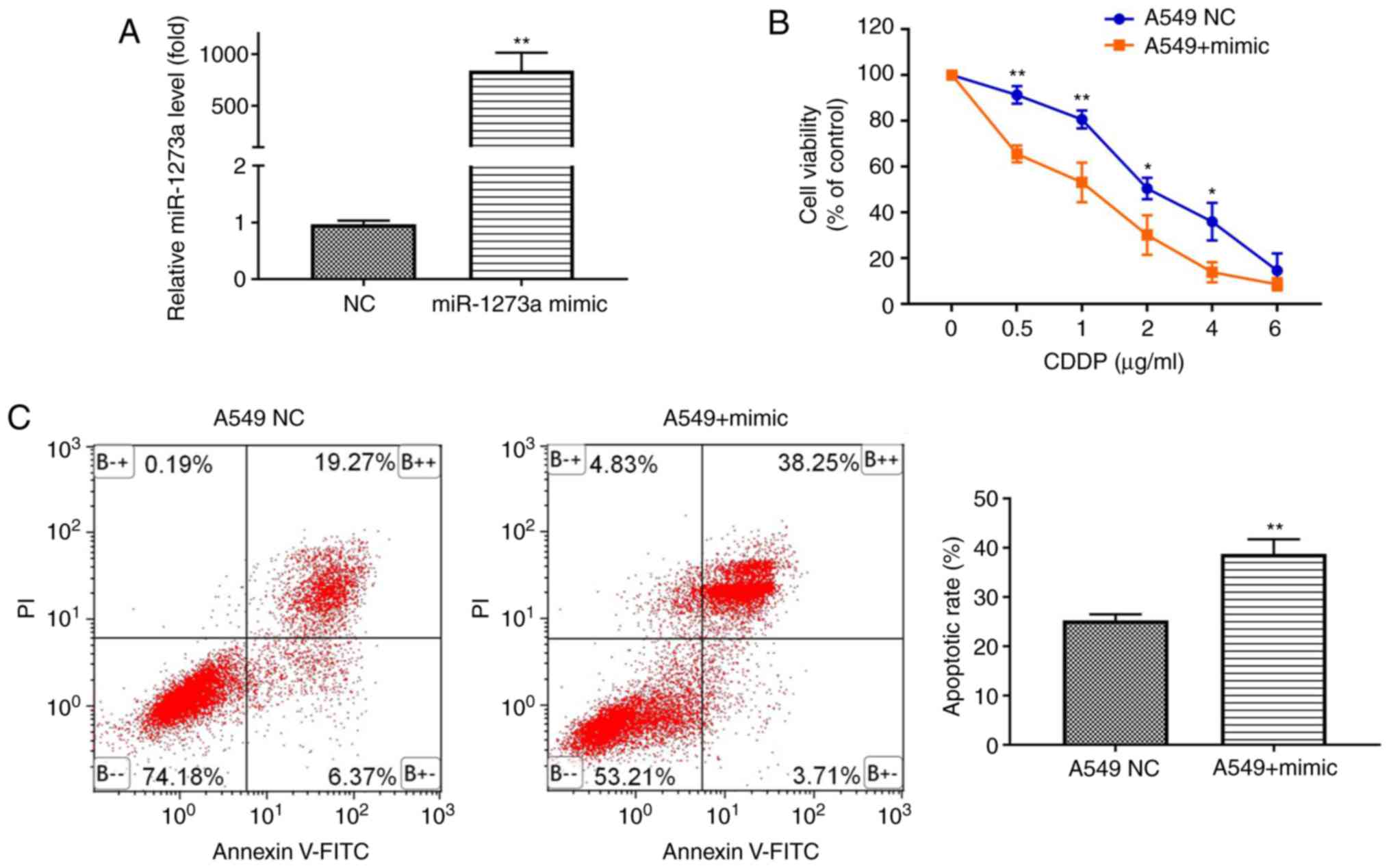
Given the decreased expression of miR-1273a in
EXOCDDP, it was hypothesized that miR-1273a acts as a
tumor suppressor. To explore whether miR-1273a affects CDDP
sensitivity, A549 cells were transfected with miR-1273a mimic or
mimic negative control (NC) (Fig.
4A), followed by treatment with different concentrations of
CDDP. The CCK-8 assay demonstrated that overexpression of miR-1273a
significantly increased the toxicity of CDDP at different
concentrations (Fig. 4B).
Consistently, the apoptosis analysis revealed that overexpression
of miR-1273a significantly enhanced CDDP-induced apoptosis compared
with the control group (25.28±1.23 vs. 38.75±3.01%, respectively;
P<0.01; Fig. 4C). Furthermore,
it was also observed that miR-1273a in exosomes can enhance the
sensitivity of A549 cells to cisplatin by exosomal transmission
(Appendix S1 and Fig. S2). These
results indicate that miR-1273a may enhance CDDP sensitivity in
A549 cells.
SDCBP may be one of the downstream
targets of miR-1273a
To explore the mechanisms underlying the regulatory
role of miR-1273a in cell sensitivity to CDDP, it was next
attempted to identify the target gene of miR-1273a. TargetScan
(www.targetscan.org) and microRNA.org (www.microrna.org) were first used to identified the
3′-untranslated region (UTR) of SDCBP which was identified as the
potential target binding region of miR-1273a (Fig. 5A). The expression of SDCBP was
detected by both RT-qPCR and western blot analyses after A549 cells
were transfected with miR-1273a mimic or mimic negative control. As
shown in Fig. 5B and C, RT-qPCR and
western blotting analyses revealed that overexpression of miR-1273a
significantly decreased the expression of SDCBP at both the mRNA
and protein levels. Furthermore, according to our findings
(Fig. S2B), miR-1273a was
significantly downregulated in EXOCDDP and upregulated
in EXOmimic (exosomes from miR-1273a mimic transfected
cells). To verify whether miR-1273a could regulate SDCBP expression
through exosome-mediated delivery, A549 cells were treated with
EXO, EXOCDDP or EXOmimic for 48 h, and the
expression of SDCBP in cells was detected by RT-qPCR and western
blotting analyses. Compared with EXO-treated cells, SDCBP
expression was significantly increased in the
EXOCDDP-treated cells but decreased in
EXOmimic-treated cells (Fig.
5D and E). These results indicated that SDCBP may be a
promising downstream target of miR-1273a.
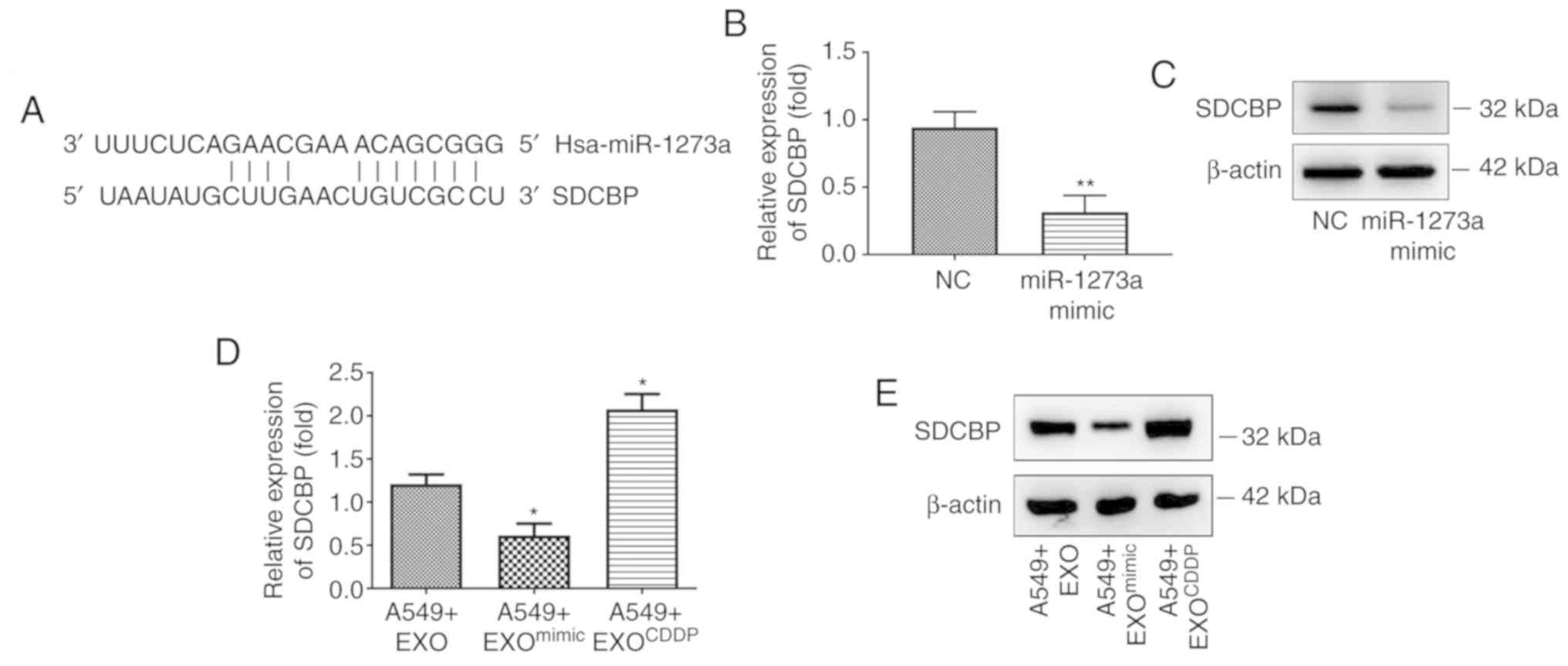 | Figure 5.SDCBP may be a downstream target of
miR-1273a. (A) Prediction of miR-1273a binding site in the
3′-untranslated region of SDCBP by TargetScan. (B and C) After
transfecting cells with miR-1273a mimics, the expression of SDCBP
was detected by reverse transcription-quantitative PCR and western
blot analyses, respectively (**P<0.01 vs. the NC group). (D and
E) After treating cells with EXO, EXOmimic, or
EXOCDDP, the expression of SDCBP was detected by reverse
transcription-quantitative PCR and western blot analyses,
respectively (*P<0.05 vs. the A549+EXO group). SDCBP, syndecan
binding protein; EXO, exosomes derived from A549 cells;
EXOCDDP, exosomes isolated from the culture medium of
CDDP-treated cells; EXOmimic, exosomes from miR-1273a
mimic-transfected cells; CDDP, cisplatin. |
Exosomal miR-1273a in plasma is
associated with the therapeutic effect of platinum-based
chemotherapy in NSCLC
Exosomes are known to contain a variety of proteins
and RNAs from tumor cells (25).
Several studies have reported that exosomal miRNAs are secreted
into the plasma, and these miRNAs may thus be used as potential
biomarkers for the diagnosis of different types of cancer (26). To further confirm whether exosomal
miR-1273a in the plasma reflects the therapeutic effect of
platinum-based chemotherapy in NSCLC, exosomal miR-1273a was
measured in the plasma in 49 patients with advanced NSCLC receiving
platinum-based chemotherapy. Exosomes isolated from the patient
plasma samples displayed typical oval-shaped vesicles on TEM
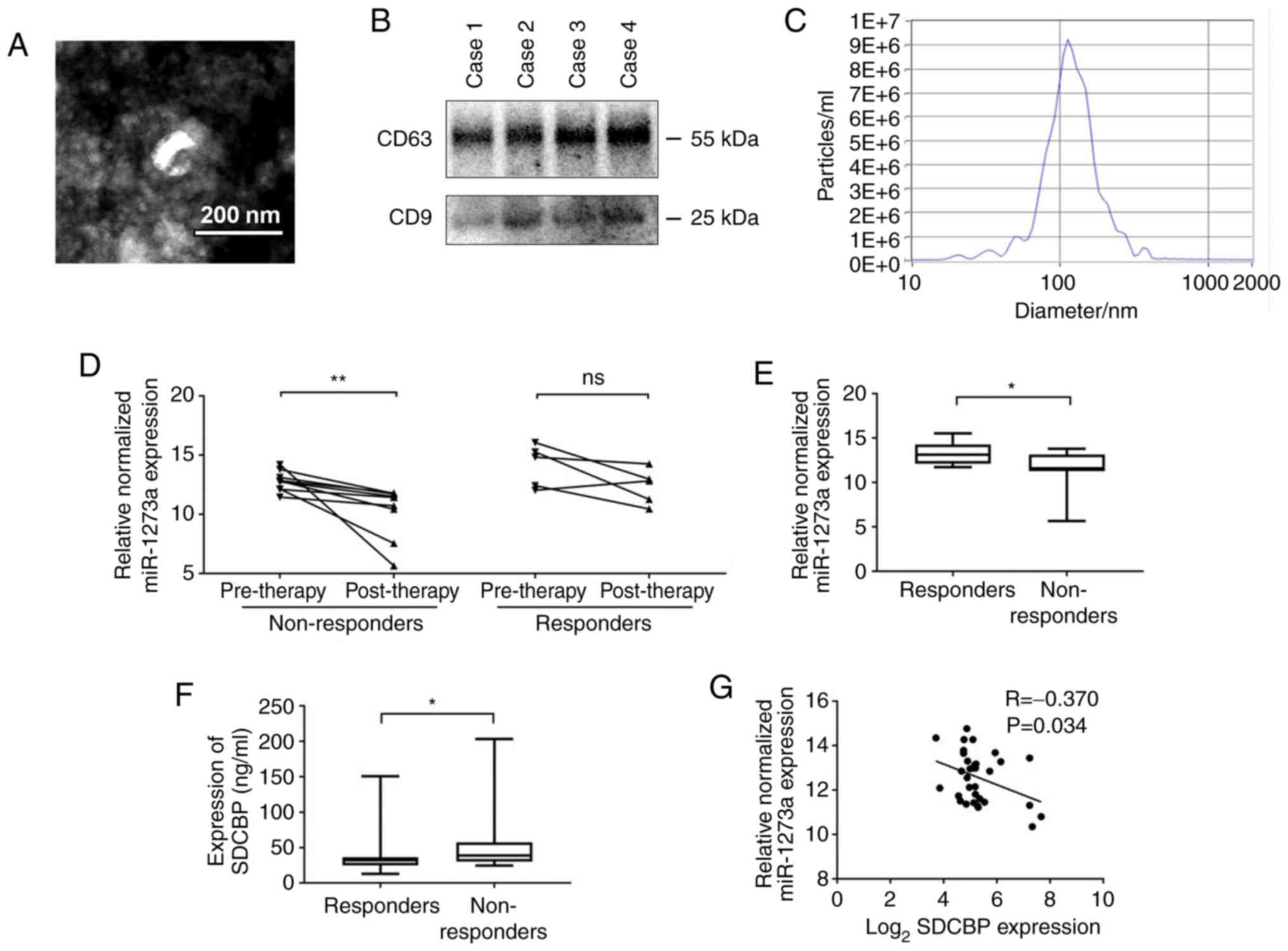
examination, with a diameter of 30–150 nm (Fig. 6A). Furthermore, the exosomal markers
CD9 and CD63 in isolated exosomes from 4 random plasma samples were
detected by western blotting (Fig.
6B), and the appropriate particle size distribution of isolated
exosomes was confirmed by NTA (Fig.
6C).
Next, we examined the miR-1273a and SDCBP expression
levels in NSCLC patients based on the therapeutic outcomes. As
shown in Fig. 6D, the level of
plasma exosomal miR-1273a in the non-responder group was
significantly reduced after chemotherapy, whereas the level of
plasma exosomal miR-1273a in the responder groups did not differ
significantly before and after chemotherapy. These results indicate
that decreased exosomal miR-1273a during chemotherapy may be
associated with poor outcome of cisplatin therapy in patients with
NSCLC. The expression of miR-1273a and SDCBP in NSCLC patients
after chemotherapy was next examined. The level of exosomal
miR-1273a in the non-responder group was significantly lower
compared with that in the responder group, whereas the expression
of SDCBP was significantly higher in the responder group (Fig. 6E and F). Furthermore, a negative
correlation between plasma exosomal miR-1273a and plasma SDCBP
expression was identified (Fig.
6G). Based on the results mentioned above, it may be inferred
that decreased exosomal miR-1273a may reduce the therapeutic effect
of platinum-based agents by regulating SDCBP expression.
Discussion
Platinum-based chemotherapy has been the major
treatment method for patients with advanced NSCLC for more than 30
years (27). However, resistance to
platinum during the course of therapy greatly limits the
therapeutic efficacy (28).
Previous research has demonstrated that exosomes derived from
CDDP-resistant NSCLC cells exhibit a differential miRNA expression
profile compared with CDDP-sensitive cells. Futhermore, exosomal
miR-100-5p and miR-425-3p were found to be involved in the
development of cisplatin resistance in NSCLC (18,29).
However, cisplatin resistance may occur rapidly in the early stages
of chemotherapy in several NSCLC cases (30). Therefore, unlike previous studies,
the present study mainly focused on the role of exosomal miRNAs in
CDDP resistance under condition of short-term CDDP stimulation.
The present study demonstrated that exosomes
secreted by CDDP-stimulated cells can reduce cell sensitivity to
CDDP, which was consistent with the findings of Xiao et al
(31). Next, our microarray
analysis revealed that the level of exosomal miR-1273a exhibited
the most notable change under CDDP treatment; therefore, it was
hypothesized that the change in exosomal miR-1273a level is
associated with CDDP resistance of A549 cells under CDDP
stimulation. Of note, although most studies have shown that
increased miRNA levels in exosomes promote the malignant behavior
of tumors, several studies have found that decreased exosomal
miRNAs may also mediate tumor progression (32). Therefore, the decreased miR-1273a
levels in exosomes may still make sense under CDDP treatment. A
recent study demonstrated that miR-1273a overexpression abrogated
the oncogenic function by rescuing the expression of activator
protein-1, interferon regulatory factor-4, CDX-2 and Zic-1 in colon
cancer cells (33), highlighting
its potential role as a tumor suppressor miRNA. Consistently, the
present study revealed that miR-1273a increased the sensitivity of
NSCLC cells to CDDP and an increase in the amount of miR-1273a in
exosomes promoted cell apoptosis. However, drug resistance is
likely the combined result of several factors. As our microarray
data also revealed other miRNA changes, additional miRNAs that may
affect the sensitivity of NSCLC cells to CDDP must be further
investigated.
A large number of studies have demonstrated that
microRNAs delivered by exosomes maintain their biological activity
after reaching the recepient cells, and then play a role by
inhibiting their target genes (34). To this end, exosomes with high
expression of miR-1273a were constructed, and it was observed that
the drug sensitivity of cells was also increased after
co-incubation with these miR-1273a-rich exosomes (Appendix S1 and
Fig. S2). Furthermore, it was
hypothesized that SDCBP may be a downstream target of miR-1273a,
based on the bioinformatics prediction of its binding site and its
oncogenic role in cell survival, stemness and chemoresistance, in
various types of human cancer (35,36).
It was next confirmed that miR-1273a overexpression inhibited the
expression of SDCBP at both the mRNA and protein levels.
Furthermore, co-incubation of cells with exosomes containing
miR-1273a at different expression levels may also result in changes
in the SDCBP expression in receptor cells. However, the effect of
SDCBP on cisplatin sensitivity in NSCLC must be thoroughly explored
in future studies.
Numerous studies have confirmed that exosomal miRNAs
may be used as biomarkers for early cancer detection and monitoring
compared with traditional blood-based cancer markers (37–39).
The present study also demonstrated a significant reduction in
exosomal miR-1273a in the plasma of patients who were not sensitive
to platinum-based chemotherapy. In agreement with our in
vitro results, there was a corresponding increase in SDCBP
levels in the plasma of non-responders. Currently, the detection of
exosomal miR-1273a in the plasma appears to be an effective and
convenient method for evaluating the efficacy of chemotherapy. Due
to the unique biological characteristics of exosomes, such as
stability, biocompatibility, permeability, low toxicity and low
immunogenicity, they may be used as a chemotherapeutic drug
delivery system (40,41). Thus, plasma exosomal miR-1273a may
also be used in the future as a promising therapeutic molecule for
NSCLC patients who are not sensitive to chemotherapy.
In conclusion, it was herein demonstrated that
decreased exosomal miR-1273a reduced the sensitivity of NSCLC to
CDDP. In addition to cell-based evidence, it was further observed
that the expression of miRNA-1273a in plasma exosomes was
significantly decreased in patients who were not sensitive to
chemotherapy. These effects may be associated with the regulatory
effect of miR-1273a on SDCBP. Therefore, for NSCLC patients who are
not sensitive to chemotherapy, exosomal miR-1273a may prove to be a
useful biomarker and feasible treatment strategy.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (no. 81672312).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
Conception and design of the study were carried out
by DW, ML and ND. Administrative and experimental support were
carried out by YY and XD. Provision of study materials and
recruitment of patients and samples were achieved by YY and YP.
Collection and assembly of data were carried out by XZ and XD. Data
analysis and interpretation were performed by XZ and CX. Writing of
the manuscript was conducted by XZ. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All patients signed informed consent forms, and this
study was authorized by the Ethics Committees of Daping Hospital
(Chongqing, China) (ratification no. 2018113). This study was
conducted in accordance with the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lam KC and Mok TS: Targeted therapy: An
evolving world of lung cancer. Respirology. 16:13–21. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mack PC, Redman MW, Chansky K, Williamson
SK, Farneth NC, Lara PN Jr, Franklin WA, Le QT, Crowley JJ and
Gandara DR; SWOG, : Lower osteopontin plasma levels are associated
with superior outcomes in advanced non-small-cell lung cancer
patients receiving platinum-based chemotherapy: SWOG study S0003. J
Clin Oncol. 26:4771–4776. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rizvi NA, Hellmann MD, Brahmer JR,
Juergens RA, Borghaei H, Gettinger S, Chow LQ, Gerber DE, Laurie
SA, Goldman JW, et al: Nivolumab in combination with platinum-based
doublet chemotherapy for first-line treatment of advanced
non-small-cell lung cancer. J Clin Oncol. 34:2969–2979. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Q, Chen Y, Feng H, Zhang B and Wang
H: Prognostic and predictive value of HURP in nonsmall cell lung
cancer. Oncol Rep. 39:1682–1692. 2018.PubMed/NCBI
|
|
6
|
Mashouri L, Yousefi H, Aref AR, Ahadi AM,
Molaei F and Alahari SK: Exosomes: Composition, biogenesis, and
mechanisms in cancer metastasis and drug resistance. Mol Cancer.
18:752019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
H Rashed M, Bayraktar E, K Helal G,
Abd-Ellah MF, Amero P, Chavez-Reyes A and Rodriguez-Aguayo C:
Exosomes: From garbage bins to promising therapeutic targets. Int J
Mol Sci. 18:5382017. View Article : Google Scholar
|
|
8
|
Ludwig AK and Giebel B: Exosomes: Small
vesicles participating in intercellular communication. Int J
Biochem Cell Biol. 44:11–15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim DH, Park S, Kim H, Choi YJ, Kim SY,
Sung KJ, Sung YH, Choi CM, Yun M, Yi YS, et al: Tumor-derived
exosomal miR-619-5p promotes tumor angiogenesis and metastasis
through the inhibition of RCAN1.4. Cancer Lett. 475:2–13. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sandfeld-Paulsen B, Jakobsen KR, Bæk R,
Folkersen BH, Rasmussen TR, Meldgaard P, Varming K, Jørgensen MM
and Sorensen BS: Exosomal proteins as diagnostic biomarkers in lung
cancer. J Thorac Oncol. 11:1701–1710. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Del Vescovo V and Denti MA: microRNA and
lung cancer. Adv Exp Med Biol. 889:153–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Czarnecka KH, Szmyd B, Baranska M,
Kaszkowiak M, Kordiak J, Antczak A, Pastuszak-Lewandoska D and
Brzeziańska-Lasota E: A strong decrease in TIMP3 expression
mediated by the presence of miR-17 and 20a enables extracellular
matrix remodeling in the NSCLC lesion surroundings. Front Oncol.
9:13722019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu G, Drescher KM and Chen XM: Exosomal
miRNAs: Biological properties and therapeutic potential. Front
Genet. 3:562012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoshii S, Hayashi Y, Iijima H, Inoue T,
Kimura K, Sakatani A, Nagai K, Fujinaga T, Hiyama S, Kodama T, et
al: Exosomal microRNAs derived from colon cancer cells promote
tumor progression by suppressing fibroblast TP53 expression. Cancer
Sci. 110:2396–2407. 2019.PubMed/NCBI
|
|
15
|
Berrout J, Kyriakopoulou E, Moparthi L,
Hogea AS, Berrout L, Ivan C, Lorger M, Boyle J, Peers C, Muench S,
et al: TRPA1-FGFR2 binding event is a regulatory oncogenic driver
modulated by miRNA-142-3p. Nat Commun. 8:9472017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu K, Liu S, Zhang W, Jia B, Tan L, Jin Z
and Liu Y: miR-494 promotes cell proliferation, migration and
invasion, and increased sorafenib resistance in hepatocellular
carcinoma by targeting PTEN. Oncol Rep. 34:1003–1010. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao L, Liu W, Xiao J and Cao B: The role
of exosomes and ‘exosomal shuttle microRNA’ in tumorigenesis and
drug resistance. Cancer Lett. 356:339–346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qin X, Yu S, Zhou L, Shi M, Hu Y, Xu X,
Shen B, Liu S, Yan D and Feng J: Cisplatin-resistant lung cancer
cell-derived exosomes increase cisplatin resistance of recipient
cells in exosomal miR-100-5p-dependent manner. Int J Nanomedicine.
12:3721–3733. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Drayton RM: The role of microRNA in the
response to cisplatin treatment. Biochem Soc Trans. 40:821–825.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eisenhauer EA, Therasse P, Bogaerts J,
Schwartz LH, Sargent D, Ford R, Dancey J, Arbuck S, Gwyther S,
Mooney M, et al: New response evaluation criteria in solid tumours:
Revised RECIST guideline (version 1.1). Eur J Cancer. 45:228–247.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang L, Pan D, Chen Q, Zhu LJ, Ou J,
Wabitsch M and Wang YX: Transcription factor Hlx controls a
systematic switch from white to brown fat through Prdm16-mediated
co-activation. Nat Commun. 8:682017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tusher VG, Tibshirani R and Chu G:
Significance analysis of microarrays applied to the ionizing
radiation response. Proc Natl Acad Sci USA. 98:5116–5121. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang YH, Liang KH, Chien RN, Hu TH, Lin
KH, Hsu CW, Lin CL, Pan TL, Ke PY and Yeh CT: A Circulating
MicroRNA signature capable of assessing the risk of hepatocellular
carcinoma in cirrhotic patients. Sci Rep. 7:5232017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang HG and Grizzle WE: Exosomes and
cancer: A newly described pathway of immune suppression. Clin
Cancer Res. 17:959–964. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fan Z, Yu J, Lin J, Liu Y and Liao Y:
Exosome-specific tumor diagnosis via biomedical analysis of
exosome-containing microRNA biomarkers. Analyst. 144:5856–5865.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zarogoulidis K, Zarogoulidis P, Darwiche
K, Boutsikou E, Machairiotis N, Tsakiridis K, Katsikogiannis N,
Kougioumtzi I, Karapantzos I, Huang H and Spyratos D: Treatment of
non-small cell lung cancer (NSCLC). J Thorac Dis. 5 (Suppl
4):S389–S396. 2013.PubMed/NCBI
|
|
28
|
Barr MP, Gray SG, Hoffmann AC, Hilger RA,
Thomale J, O'Flaherty JD, Fennell DA, Richard D, O'Leary JJ and
O'Byrne KJ: Generation and characterisation of cisplatin-resistant
non-small cell lung cancer cell lines displaying a stem-like
signature. PLoS One. 8:e541932013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma Y, Yuwen D, Chen J, Zheng B, Gao J, Fan
M, Xue W, Wang Y, Li W, Shu Y, et al: Exosomal transfer of
cisplatin-induced miR-425-3p confers cisplatin resistance In NSCLC
through activating autophagy. Int J Nanomedicine. 14:8121–8132.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lara PN Jr, Gandara DR, Longmate J,
Gumerlock PH, Lau DH, Edelman MJ, Gandour-Edwards R, Mack PC,
Israel V, Raschko J, et al: Activity of high-dose toremifene plus
cisplatin in platinum-treated non-small-cell lung cancer: A phase
II California cancer consortium trial. Cancer Chemother Pharmacol.
48:22–28. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xiao X, Yu S, Li S, Wu J, Ma R, Cao H, Zhu
Y and Feng J: Exosomes: Decreased sensitivity of lung cancer A549
cells to cisplatin. PLoS One. 9:e895342014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Z, Li X, Sun W, Yue S, Yang J, Li J,
Ma B, Wang J, Yang X, Pu M, et al: Loss of exosomal miR-320a from
cancer-associated fibroblasts contributes to HCC proliferation and
metastasis. Cancer Lett. 397:33–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Q, Zhang C, Ma JX, Ren H, Sun Y and
Xu JZ: Circular RNA PIP5K1A promotes colon cancer development
through inhibiting miR-1273a. World J Gastroenterol. 25:5300–5309.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kooijmans SA, Schiffelers RM, Zarovni N
and Vago R: Modulation of tissue tropism and biological activity of
exosomes and other extracellular vesicles: New nanotools for cancer
treatment. Pharmacol Res. 111:487–500. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jana S, Sengupta S, Biswas S, Chatterjee
A, Roy H and Bhattacharyya A: miR-216b suppresses breast cancer
growth and metastasis by targeting SDCBP. Biochem Biophys Res
Commun. 482:126–133. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Talukdar S, Das SK, Pradhan AK, Emdad L,
Windle JJ, Sarkar D and Fisher PB: MDA-9/syntenin (SDCBP) is a
critical regulator of chemoresistance, survival and stemness in
prostate cancer stem cells. Cancers (Basel). 12:532019. View Article : Google Scholar
|
|
37
|
Cui M, Wang H, Yao X, Zhang D, Xie Y, Cui
R and Zhang X: Circulating MicroRNAs in cancer: Potential and
challenge. Front Genet. 10:6262019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang C, Hopfner F, Katsikoudi A, Hein R,
Catli C, Evetts S, Huang Y, Wang H, Ryder JW, Kuhlenbaeumer G, et
al: Serum neuronal exosomes predict and differentiate Parkinson's
disease from atypical parkinsonism. J Neurol Neurosurg Psychiatry.
91:720–729. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhao X, Dou J, Cao J, Wang Y, Gao Q, Zeng
Q, Liu W, Liu B, Cui Z, Teng L, et al: Uncovering the potential
differentially expressed miRNAs as diagnostic biomarkers for
hepatocellular carcinoma based on machine learning in the cancer
genome atlas database. Oncol Rep. 43:1771–1784. 2020.PubMed/NCBI
|
|
40
|
Pullan JE, Confeld MI, Osborn JK, Kim J,
Sarkar K and Mallik S: Exosomes as drug carriers for cancer
therapy. Mol Pharm. 16:1789–1798. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Fan Z, Xiao K, Lin J, Liao Y and Huang X:
Functionalized DNA enables programming exosomes/vesicles for tumor
imaging and therapy. Small. 15:e19037612019. View Article : Google Scholar : PubMed/NCBI
|