Introduction
Triple-negative breast cancer (TNBC) is a subtype of
breast cancer in which estrogen receptor (ER) and the
progesterone receptor (PgR), as well as the epidermal growth
factor receptor 2 (HER2) are all negative (1). As one of the most aggressive breast
cancer subtypes, TNBC accounts for 10–20% of all malignant breast
tumors, and frequently has a worse prognosis and greater risks for
recurrence and metastasis than other types of breast cancer
(2).
While the number of potential therapeutic agents
being tested in clinical trials with metastatic breast cancer
patients continues to increase, currently, chemotherapy is the
standard therapeutic strategy for TNBC (3,4).
Paclitaxel (PTX), a natural taxane diterpene, was initially
isolated from the bark of the Pacific yew (Taxus
brevifolia), and is currently widely used in chemotherapy for
TNBC (5,6). PTX specifically binds to and
stabilizes the β-subunit of tubulin, thereby inhibiting the
disassembly of microtubules in dividing cells, resulting in mitotic
arrest and subsequent cell death (7,8).
However, while PTX is effective for treating several types of
cancer, >50% of patients with TNBC become resistant to
chemotherapy, typically within 6 to 10 months (9,10). The
frequent development of PTX drug resistance in TNBC patients
underscores the importance of exploring the underlying mechanisms
of PTX resistance, and identifying the critical molecules involved
in this process. A better understanding of the mechanism for PTX
resistance may improve our ability to treat BC.
In the present study, the related signaling pathways
and genes in the process of PTX resistance in BC cell lines were
investigated. Notably, a serine protease inhibitor, clade E member
1 (SERPINE1) was identified as a critical factor that
mediated PTX drug resistance in BC cells. SERPINE1, also
known as plasminogen activator inhibitor, type 1 (PAI-1), acts as a
vital inhibitor of serine proteases that play important roles in
signal transduction, cell adhesion, and cell migration (11–13).
SERPINE1 also regulates urokinase and plasminogen activators
that transform the pro-enzyme plasminogen to plasmin, which
subsequently promotes cellular invasion via activation of matrix
metalloproteinases and degradation of the extracellular matrix
(12). A high level of
SERPINE1 has been revealed to be associated with a poor
prognosis of breast cancer (11).
However, whether SERPINE1 plays a role in the development of
drug resistance in BC is unknown.
Materials and methods
Bioinformatics analysis
Gene expression datasets used for bioinformatics
analyses were downloaded from the Gene Expression Omnibus (GEO;
http://www.ncbi.nlm.nih.gov/geo/)
(14) by importing the accession
numbers GSE28784 and GSE90564. GSE28784 contains the gene
expression data of sensitive, docetaxel-resistant, and
PTX-resistant MDA-MB-231 cells. GSE90564 is a gene expression
profiling dataset consisting of 5 BC cell lines (BT20, SUM149,
MDA-MB-231, MDA-MB-436, and MDA-MB-468), which are resistant to PTX
after being exposed to increased concentrations of PTX, and their
gene expression patterns were compared to those of parental
PTX-sensitive cells. We used the 151 genes that were shared by both
datasets to perform Gene Ontology (GO) (15) and Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway enrichment analyses (16) for the purpose of identifying gene
functions and pathways related to PTX resistance. Gene interaction
analysis and Search Tool for the Retrieval of Interacting Genes
(STRING; http://string-db.org) were used to
analyze the functional associations between genes that may be
responsible for PTX resistance, and then to identify the core
regulatory genes on the list.
K-means clustering method
Based on the previous studies (17,18),
K-means clustering method was applied to calculate the different
clusters of the gene interaction network.
Cell line generation and cell
culture
MDA-MB-231 and MCF-7 cells were obtained from the
ATCC. Paclitaxel-resistant cells were created as previously
described (19,20). The parent PTX-sensitive cells were
continuously maintained in a paclitaxel containing medium in which
the paclitaxel concentration gradient was between 2 and 30 nM. Both
cell lines were grown in RPMI-1640 medium (Thermo Fisher
Scientific, Inc.) containing 1% penicillin/streptomycin and 10%
fetal bovine serum (Thermo Fisher, Scientific, Inc.). Cells were
incubated at 37°C and 5% CO2. For PTX cells, the culture
medium contained 30 nM paclitaxel.
MTT assay
Cells (1×104 cells/well) were plated into
96-well plates. After culture for 24 h at 37°C, the original medium
was replaced with fresh medium containing 0, 0.1, 0.5, 1, 5, and 10
µM paclitaxel. After 48 h, the cells in each well were treated with
20 µl of MTT (0.5 mg/ml), and incubation was continued for an
additional 4 h at 37°C, and then 100 µl of DMSO was applied to
dissolve the formazan in each well for 15 min at 37°C.
Subsequently, the optical density (OD) was examined at a wavelength
of 492 nm using a plate reader (Thermo Fisher Scientific,
Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The total RNAs were isolated from the treated cells
or clinical tissues using the TRIzol reagent (cat no. 15596-026;
Invitrogen; Thermo Fisher Scentific, Inc.). The purified RNA was
used as a template to carry out reverse transcription using the
Prime Script RT Master Mix Perfect Real Time kit (Takara
Biotechnology Co., Ltd.). Quantitative analysis of target genes was
performed using SYBR® GreenER™ qPCR SuperMix
(Invitrogen; Thermo Fisher Scientific, Inc.) on an ABI 7700 system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. The thermocycling procedures were
as follows: Incubation at 95°C for pre-denaturation for 2 min,
followed by 40 cycles with denaturation at 95°C for 22 sec, and
annealing at 59°C for 20 sec. The primer sequences used were as
follows: SERPINE1 forward, 5′-GCAAGGCACCTCTGAGAACT-3′ and
reverse, 5′-GGGTGAGAAAACCACGTTGC-3′. β-actin was utilized
for the standardization, and its primer sequences were: Forward,
5′-AGAGCTACGAGCTGCCTGAC-3′ and reverse,
5′-AGCACTGTGTTGGCGTACAG-3′.
Western blot analysis
The antibodies in this experiment were obtained from
Proteintech. Treated cells were lysed in cell lysis buffer
containing 1 mM PMSF (Beyotime Institute of Biotechnology) on ice
for 15 min and centrifuged at 16,100 × g for 5 min at 4°C. Next,
50-µg samples of total protein were dispersed on 10% SDS-PAGE gels
by electrophoresis and transferred onto cut-out nitrocellulose
membranes (Beyotime Institute of Biotechnology). The membranes were
blocked for 1 h with 5% skimmed milk at room temperature and
incubated with anti-SERPINE1 antibody (cat. no. A00637-1; 1:1,000
dilution; Boster Biological Technology), VEGFA (cat. no. BA0407;
1:500 dilution; Wuhan Boster Biological Technology Co., Ltd.),
cleaved caspase-3 (product code ab32042; 1:500 dilution; Abcam),
Bax (cat. no. A00183; 1:1,000 dilution; Wuhan Boster Biological
Technology Co., Ltd.) and β-actin (product code ab8224; 1:1,000
dilution; Abcam) overnight at 4°C. Subsequently, they were
incubated with horseradish peroxidase (HRP)-labeled secondary
antibodies (1:1,000 dilution; product code ab7090; Abcam) for 1 h
at room temperature. After incubation, the membranes were developed
using chemiluminescent substrates (Beyotime Institute of
Biotechnology). The densitometric quantification of the bands were
calculated using the ImageJ software (version 1.50b; National
Institutes of Health; http://imagej.nih.gov/ij/).
Gene knockdown with short hairpin RNAs
(shRNAs)
shRNA
(5′-CCGGCCTGAAGGTGAAGAACATCATCTCGAGATGATGTTCTTCACCTTCAGGTTTTTG-3′)
that specifically targeted SERPINE1 and control shRNA
plasmids
(5′-CCGGCAACAAGATGAAGAGCACCAACTCGAGTTGGTGCTCTTCATCTTGTTGTTTTTG-3′)
were acquired from Sigma-Aldrich; Merck KGaA. For transient
transfection, the treated MDA-MB-231/PTX and MCF-7/PTX cells
(6×105 cells/well) were plated into 6-well plates and
cultured overnight at 37°C. The adherent cells were then
transfected with the shRNAs using Lipofectamine 2000™ reagent
(Thermo Fisher Scientific, Inc.) according to the instructions
supplied by the manufacturer. Briefly, MDA-MB-231/PTX and MCF-7/PTX
cells were transfected with control and SERPINE1 shRNA at
concentration of 100 ng/µl for 5 min at room temperature. After
being cultured for 5 h at 37°C, the cells were washed with PBS and
cultured with fresh culture medium. Finally, the transfected cells
were cultured for an additional 48 h at 37°C prior to being treated
or harvested for further evaluation.
Flow cytometric assay
The treated MDA-MB-231/PTX and MCF-7/PTX cells
(1×106 cells/well) were double-stained with an Annexin
V-FITC/PI apoptosis detection kit (Hangzhou Multi Sciences (Lianke)
Biotech Co., Ltd.) according to the manufacturer's instructions.
The results were monitored by flow cytometry (FACScan; BD
Biosciences). The data was analyzed using the CellQuest™ Pro
software (version 5.1; BD Biosciences). Early-stage apoptotic cells
contained Annexin V-positive and PI-negative cells, while
late-stage apoptotic cells included both Annexin V-positive and
PI-positive cells. All experiments were performed independently in
triplicate.
In vivo tumorigenic assay
A total of 30 male nude mice (6 weeks old, 20.6±2.3
g) were provided from Department of Laboratory Animals of Central
South University, and the housing conditions of the nude mice were
as follows: 22±1°C temperature, 50–60% humidity, 12-h light/dark
cycle, and ad libitum access to food and water. To evaluate
the tumorigenic capacity of BC cells and MDA-MB-231 cells, they
were first transfected with either the control shRNA construct or
shSERPINE1 and then cultured to achieve a sufficient
population of SERPINE1-knockdown cells. Next,
5×106 cells were harvested, re-suspended, and
subcutaneously injected into the 30 nude mice (5 mice in each
group). The tumor growth was periodically monitored by examining
the tumor size. At the end of the study, the mice were sacrificed
after being anesthetized with 1% pentobarbital sodium (i.p.) at a
dose of 50 mg/kg and then sacrificed by decapitation. Successful
anesthesia was considered mice breathing and with a heartbeat, and
mice that did not have a heartbeat or breath were presumed
euthanized. Then tumors were removed for further evaluation. Animal
experiments were approved by the Animal Ethics Committee of
Xiangnan University Affiliated Hospital (approval no.
2019sydw0821).
Statistical analysis
All results were expressed as a mean value ±
standard deviation (SD). Statistical analyses were conducted using
GraphPad Prism (version 6.0; GraphPad Software, Inc.) or SPSS 14.0
(SPSS, Inc.). Unpaired Student's t-test was used to analyze the
differences between two groups. One-way analysis of variance
(ANOVA) with Tukey's post hoc test was used for comparisons among
three or more groups. A P-value <0.05 was considered to indicate
a statistically significant difference. All experiments were
repeated at least three times.
Results
Genes and signaling pathways involved
in PTX resistance in BC cell lines
In order to identify common mechanisms for PTX
resistance among BC cells, two datasets that were generated by
comparing the gene expression patterns of PTX-resistant cells with
those of PTX-sensitive cells were examined. A list of
differentially expressed genes that may be related to PTX
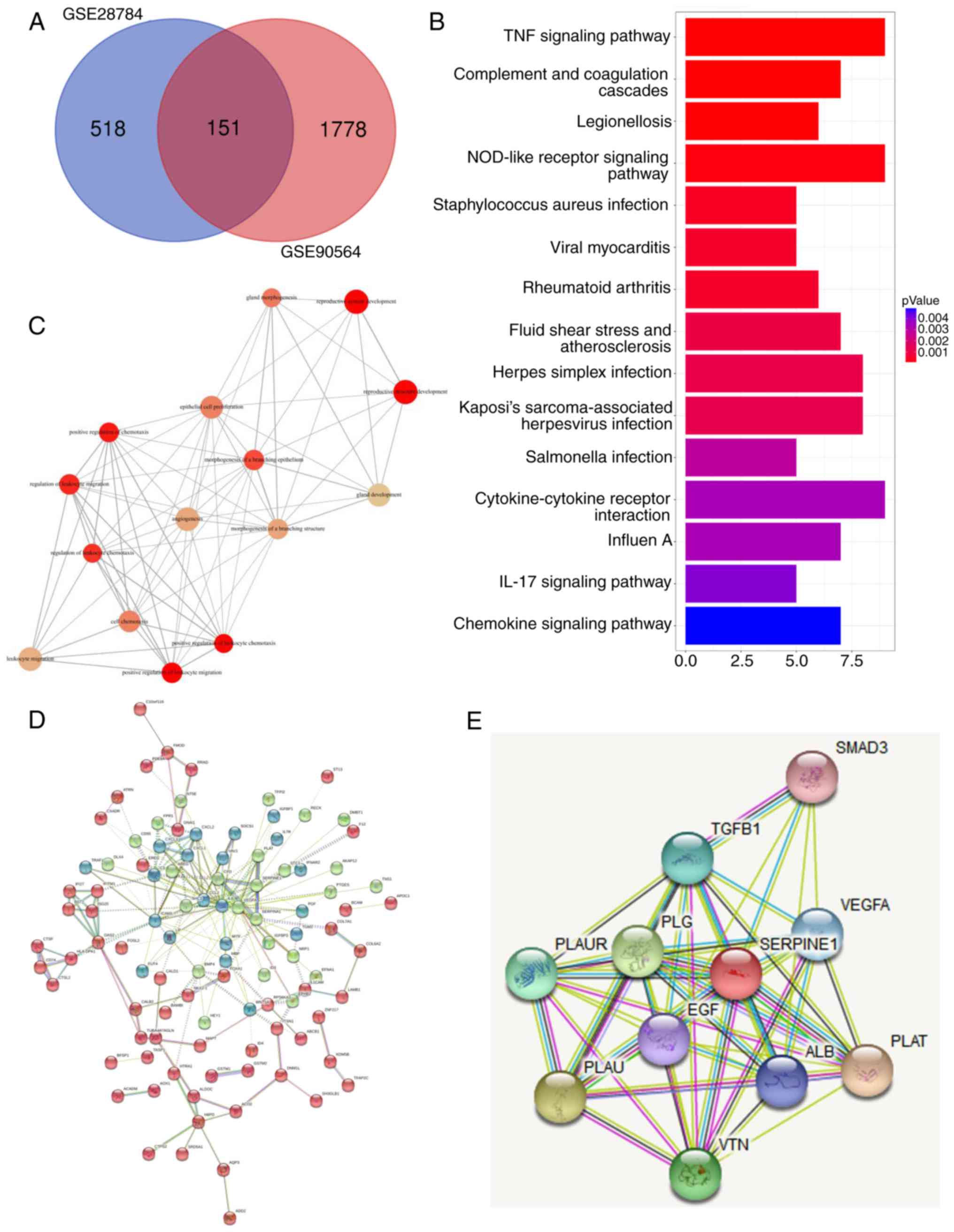
resistance was generated for each dataset. A Venn diagram revealed
that 151 overlapping genes were differentially regulated in both
datasets (Fig. 1A). Pathway
analysis indicated that the differentially expressed molecules were
highly involved in TNF and NOD signaling (Fig. 1B), which are pathways known to
regulate cancer metastasis (21,22).
In addition, a subsequent annotation analysis also revealed that
these differentially expressed genes mainly participated in
multiple biological processes including cell migration and
chemotaxis (Fig. 1C). Next, based
on the differentially expressed genes, various genes with high core
connectivity were identified through the gene interaction network
(Fig. 1D). In addition, STRING was
applied to reveal the interaction of SERPINE1-generated proteins,
among which, it was observed that SERPINE1 exhibited close
associations with several genes (SMAD3, TGF-β1, VEGFA, PLAUR, PLG,
PLAU, VTN, ALB, PLAT and EGF) (Fig.
1E). To further examine the role of SERPINE1 in the
development of PTX resistance in TNBC cells, SERPINE1
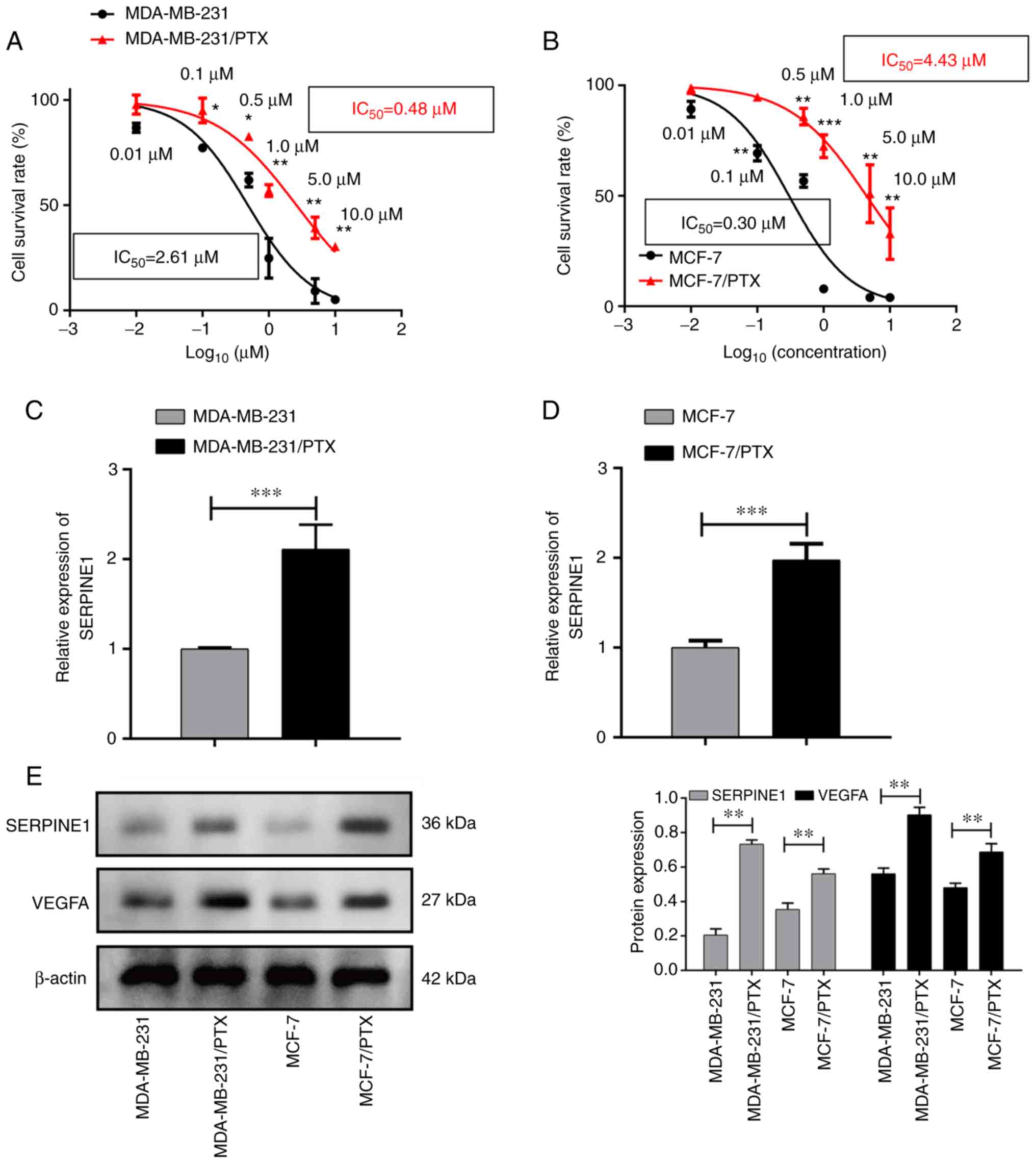
expression in PTX-resistant and -sensitive cells was evaluated. As
revealed in Fig. 2A and B, two
breast cancer cell lines were revealed to develop PTX resistance,
characterized by increased IC50 to PTX, as demonstrated
by their higher survival rates when compared to their PTX-sensitive
parental cells after treatment with specific doses of PTX. qPCR
results revealed that SERPINE1 was highly expressed in
PTX-resistant cells (Fig. 2C and
D), and was overexpressed in the PTX-resistant cells when
compared to the parental cells. Notably, a concomitant increase in
VEGFA expression in the PTX-resistant cells was also
detected (Fig. 2E).
Suppression of SERPINE1 abolishes PTX
resistance and promotes BC cell apoptosis
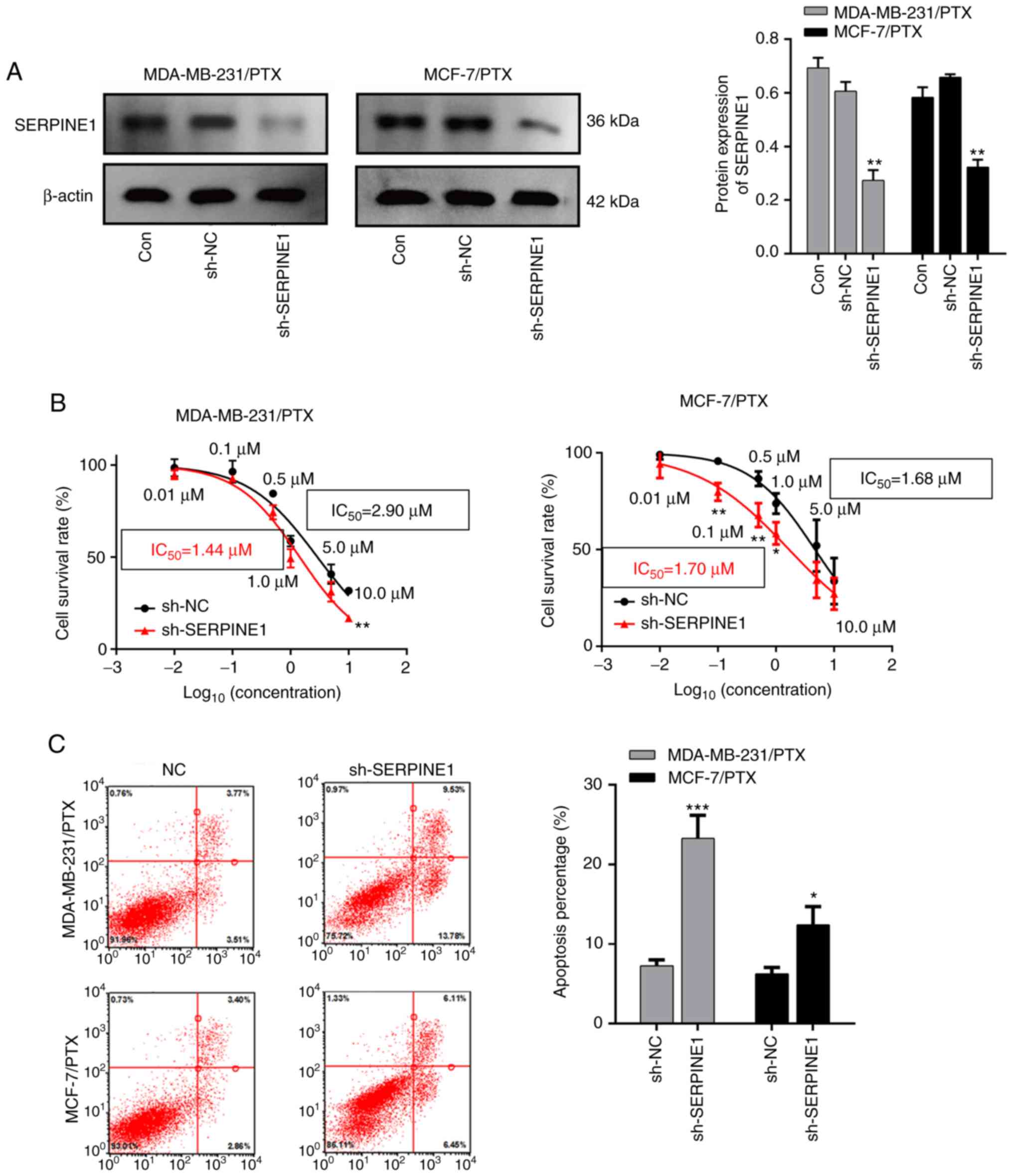
Given that SERPINE1 levels were significantly
increased in PTX resistant cells, it was investigated whether
knockdown of SERPINE1 impaired resistance of BC to PTX. A
shRNA that specifically targeted SERPINE1 RNA was used to
efficiently suppress SERPINE1 expression in MDA-MB-231 and
MCF-7 PTX-resistant cells (Fig.
3A), and it was determined that knockdown of SERPINE1
significantly inhibited the survival of PTX-resistant BC cells
(Fig. 3B) with attenuated the
IC50 to PTX. Furthermore, a flow cytometric analysis
indicated that SERPINE1 suppression significantly induced
apoptosis in large populations of both PTX-resistant cell lines
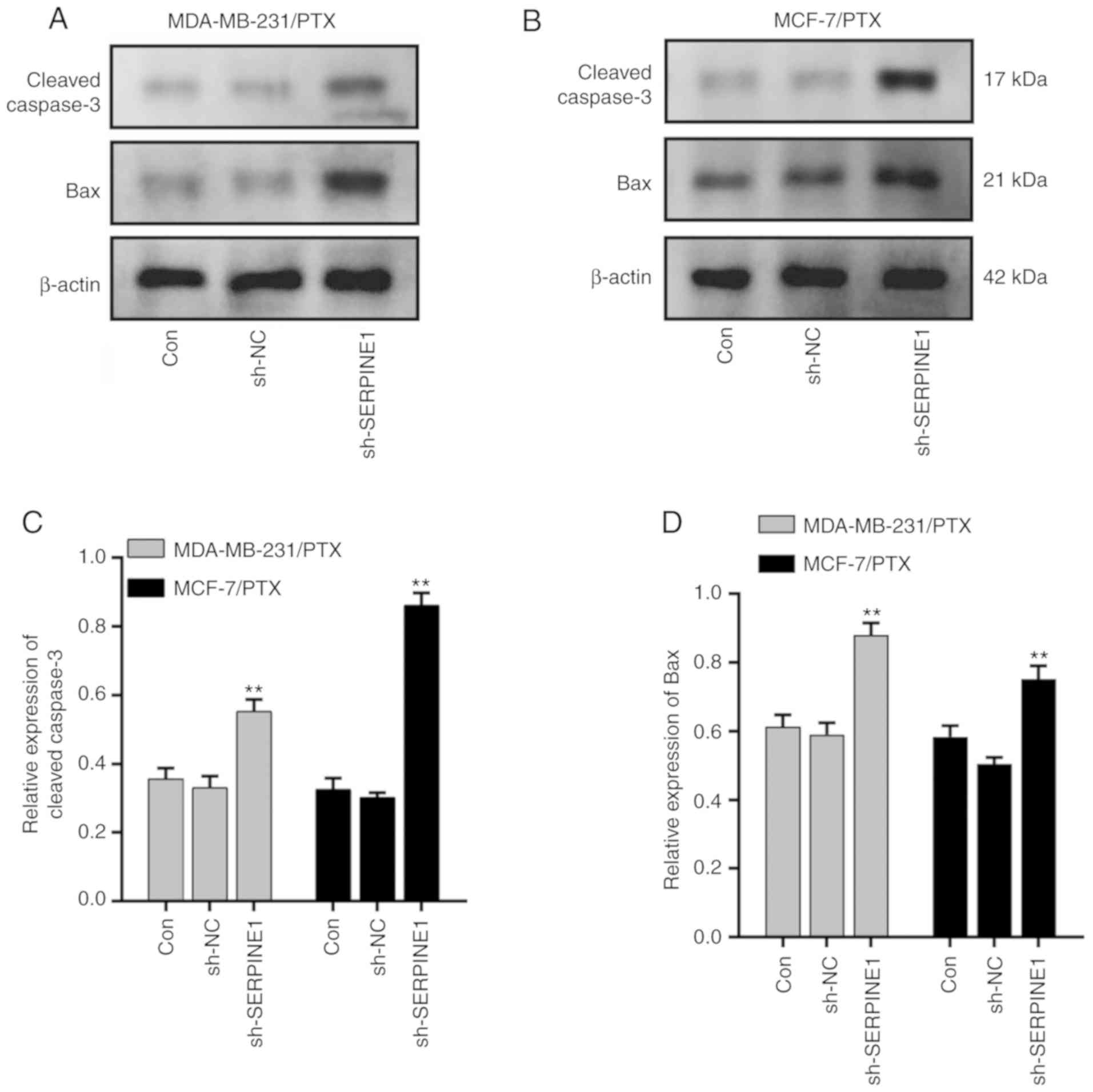
(Fig. 3C). Moreover, cleaved
caspase-3 and Bax, which are important markers of cell apoptosis
activation (23), were both
significantly upregulated following SERPINE1 suppression
(Fig. 4) in MDA-MB-231/PTX
(Fig. 4A and C) and MCF-7/PTX
(Fig. 4B and D) cells.
Knockdown of SERPINE1 suppresses the
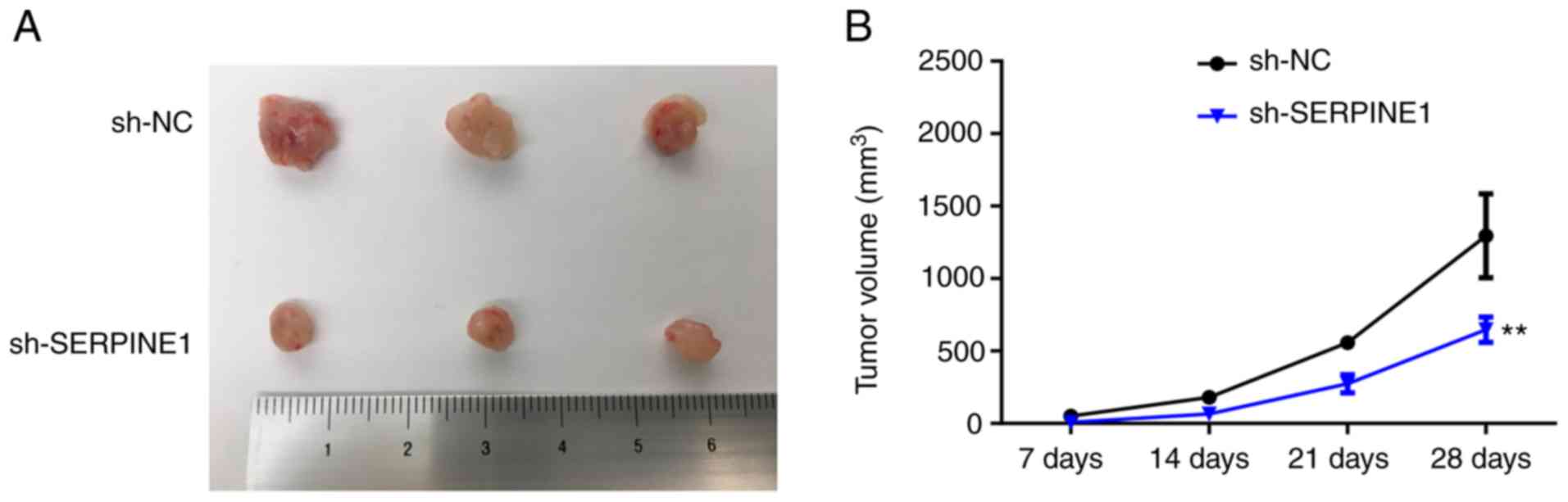
tumorigenic capacity of PTX-resistant TNBC cells in vivo
To explore how SERPINE1 may regulate the
survival of PTX-resistant cells in vivo, PTX-resistant BC
cells with or without SERPINE1 knockdown were inoculated
into mice. Consistent with the in vitro observations, the
tumors generated by SERPINE1-knockdown cells were
significantly smaller than those generated by the control cells
(Fig. 5), indicating that
suppression of SERPINE1 may be an effective strategy for
counteracting PTX-resistant BC cell proliferation in
vivo.
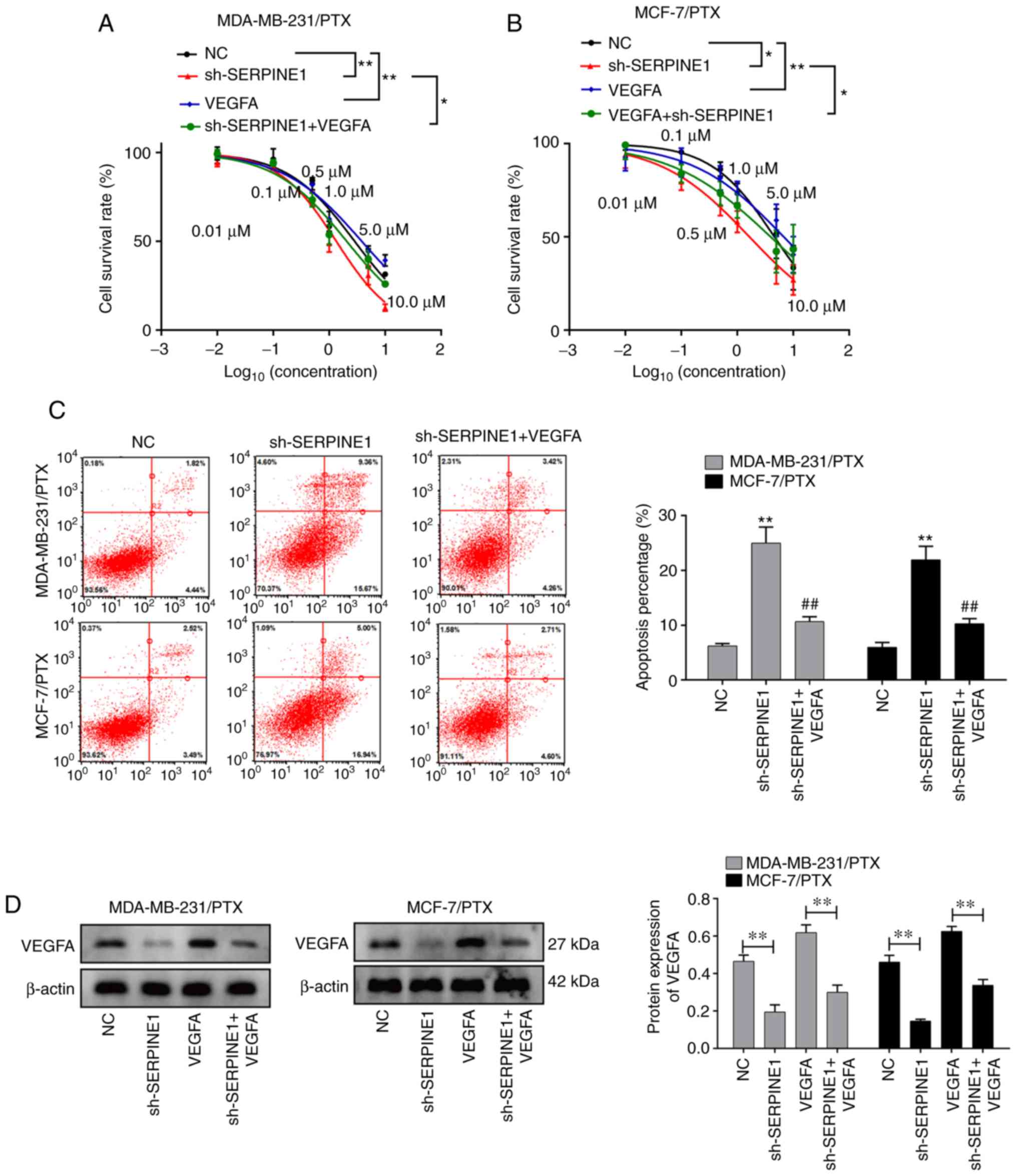
VEGFA may mediate SERPINE1 in
promoting PTX resistance
Since concomitant increases in VEGFA and
SERPINE1 were detected in PTX-resistant BC cell lines, and
given the central role of VEGFA in promoting tumor
angiogenesis and growth (23–25),
it was hypothesized that SERPINE1 may promote PTX resistance
through induction of VEGFA. To test this hypothesis,
PTX-resistant cells with or without SERPINE1 knockdown were
further treated with VEGFA. Notably, it was revealed that
VEGFA treatment abolished the effect of SERPINE1
suppression in regulating cell survival (Fig. 6A and B), suggesting VEGFA
induction as a potential mechanism for SERPINE1-regulated
PTX resistance. Consistent with this observation, the addition of
VEGFA also significantly prevented cells from entering
apoptosis (Fig. 6C). The ability of
SERPINE1 to regulate VEGFA in TNBC cell lines was
also confirmed. As revealed in Fig.
6D, knockdown of SERPINE1 significantly decreased
VEGFA expression in both BC cell lines, indicating that
SERPINE1 acts to upregulate VEGFA in BC cells.
Discussion
Breast cancer, as a heterogeneous disease, possesses
multifarious molecular subtypes. The most well recognized BC
markers are hormonal receptors (ER, PGR, and HER2), and breast
cancers that express specific receptors can be treated with drugs
that specifically target those receptor molecules (26–29).
Conversely, TNBC, which lacks specific receptors, has emerged as
the most aggressive BC subtype and is challenging to treat
(30–35). To date, no single treatment has
proven to be effective in all BC subtypes. Chemotherapeutic drugs
are commonly used to treat receptor-negative subtypes, and taxanes
are considered to be the first-line treatment for TNBC (36–38).
However, treatment with taxanes (such as PTX) usually leads to a
short-lasting benefit due to the development of PTX resistance
(36,39). Therefore, gaining a better
understanding of the underlying mechanisms for BC PTX resistance is
critical for improving the efficacy of chemotherapy and developing
new strategies for the treatment of BC.
In the present study, SERPINE1 was identified
as an important factor that mediates development of PTX resistance
in TNBC cells. SERPINE1 expression was significantly
increased in PTX-resistant cells when compared with PTX-sensitive
cells. Overexpression of SERPINE1 has been reported to
enhance the migration of cancer cells (40). Although the genetic and
environmental determinants of SERPINE1 expression are not
fully understood, several studies have suggested that
SERPINE1 levels can be regulated by cytokines, growth
factors, and hormones (41,42). Higgins (43) reported that SERPINE1 was
localized at the tumor invasive front, and its expression could be
induced by TGF-β1 during the early progression stage of squamous
cell carcinoma. Other findings suggested involvement of the
EGFR/MEK/Rho-ROCK signaling pathway in SERPINE1 induction
(44).
In PTX-resistant TNBC cells, it was revealed that
knockdown of SERPINE1 by shRNA significantly decreased cell
survival and promoted cell apoptosis. Previous study indicated that
an increase of SERPINE1 expression protects against
programmed cell death (45).
Conversely, studies revealed that SERPINE1 deficiency
promotes apoptosis in multiple cancer types, which is consistent
with our observation (45–47). The underlying mechanism for
SERPINE1-mediated cell survival may involve activation of
the Akt and ERK signaling pathways and suppression of
Fas/FasL-dependent apoptosis (48).
The present study demonstrated that knockdown of SERPINE1
disrupted tumorigenesis, which suggests the value of targeting
SERPINE1 to treat BC and prevent PTX drug resistance in
vivo.
Several mechanisms may contribute to
SERPINE1-mediated cancer progression and PTX resistance.
According to previous studies, SERPINE1 functions to prevent
excessive degradation of the extracellular matrix, modulates cell
adhesion, and stimulates cell proliferation and angiogenesis
(49,50). Tumor growth and metastasis are
heavily dependent on angiogenesis (51). Notably, PTX displays antiangiogenic
activity via its antiproliferative effect on activated endothelial
cells, and that effect is achieved by downregulating the levels of
VEGF and Ang-1 in tumor cells (52). It was hypothesized that PTX
resistance mediated by SERPINE1 may involve a VEGF factor.
When SERPINE1 expression was knocked down in BC cells,
VEGFA expression was significantly decreased, suggesting
that SERPINE1 may function as an inducer of VEGFA in
order to promote PTX resistance in cancer cells. VEGFs are known to
play important roles in angiogenic processes that are critical for
tumor cell survival, proliferation, and migration (24). Loss of VEGFA expression was
reported to increase the sensitivity of colorectal cancer cells to
5-fluorouracil by inducing apoptosis (53,54).
Humanized monoclonal antibodies directed against VEGFA were
used for antiangiogenic therapy in the clinical treatment of cancer
(55). Moreover, accumulating
evidence indicates that VEGFA plays an important role in
enhancing cancer cell resistance to chemotherapeutic drugs by
protecting cancer cells from the cytotoxic effects of those agents
(56). For example, the efficacy of
nanoparticle albumin-bound PTX (nab-PTX) in treating human breast
cancer was significantly enhanced by the concurrent administration
of anti-VEGFA (57).
Although potential mechanisms must be further explored, studies
have suggested the involvement of apoptosis regulatory molecules
and the PI3K/AKT signaling pathway (58), which are associated with
SERPINE1 function.
In conclusion, the present data demonstrated that
increased levels of SERPINE1 expression contributed to the
resistance of BC to treatment with PTX. In addition,
SERPINE1-mediated drug resistance is mediated by an
upregulation of VEGFA and subsequent suppression of cell
apoptosis. The present findings also suggest SERPINE1 as a
potential target for eliminating PTX resistance during cancer
treatment. However, certain limitations still remain in the present
study, such as the specific mechanism of SERPINE1 in BC with
PTX resistance, the influences of SERPINE1 on the migration
and invasion abilities of BC with PTX resistance.
Acknowledgements
We would like to thank Dr Edward C. Mignot, Shandong
University, for linguistic advice.
Funding
The present study was supported by Hunan Provincial
Natural Science Foundation of China (grant no. 2018JJ6123), the
Planned Science and Technology Project of Hunan Province (grant no.
2017SK4010), the Scientific Research Fund of Hunan Provincial
Health and Family Planning Commission (grant no. C2017014),
Chenzhou Science and Technology Project, and the Natural Science
Foundation of Xiangnan University (grant no. 2015XB12).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
QZ and DJ designed the experiments. QZ performed
most of the experiments with the assistance of LL. QZ and LL
collected and analyzed the data. DJ validated the data analysis. QZ
drafted the manuscript and LL and DJ revised the draft. All authors
approved the final manuscript before submission.
Ethics approval and consent to
participate
Animal experiments were approved by the Animal
Ethics Committee of Xiangnan University Affiliated Hospital
(approval no. 2019sydw0821).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gong Z, Wang J, Wang D, Buas MF, Ren X,
Freudenheim JL, Belinsky SA, Liu S, Ambrosone CB and Higgins MJ:
Differences in microRNA expression in breast cancer between women
of African and European ancestry. Carcinogenesis. 40:61–69. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tomao F, Papa A, Zaccarelli E, Rossi L,
Caruso D, Minozzi M, Vici P, Frati L and Tomao S: Triple-negative
breast cancer: New perspectives for targeted therapies. Onco
Targets Ther. 8:177–193. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cardoso F, Costa A, Norton L, Senkus E,
Aapro M, André F, Barrios CH, Bergh J, Biganzoli L, Blackwell KL,
et al: ESO-ESMO 2nd international consensus guidelines for advanced
breast cancer (ABC2). Breast. 23:489–502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Senkus E, Kyriakides S, Ohno S,
Penault-Llorca F, Poortmans P, Rutgers E, Zackrisson S and Cardoso
F; ESMO Guidelines Committee, : Primary breast cancer: ESMO
clinical practice guidelines for diagnosis, treatment and
follow-up. Ann Oncol. 26 (Suppl 5):v8–v30. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li J, Wang F, Sun D and Wang R: A review
of the ligands and related targeting strategies for active
targeting of paclitaxel to tumours. J Drug Target. 24:590–602.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paridaens R, Biganzoli L, Bruning P, Klijn
JG, Gamucci T, Houston S, Coleman R, Schachter J, Van Vreckem A,
Sylvester R, et al: Paclitaxel versus doxorubicin as first-line
single-agent chemotherapy for metastatic breast cancer: A European
organization for research and treatment of cancer randomized study
with cross-over. J Clin Oncol. 18:724–733. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gascoigne KE and Taylor SS: How do
anti-mitotic drugs kill cancer cells? J Cell Sci. 122:2579–2585.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kavallaris M: Microtubules and resistance
to tubulin-binding agents. Nat Rev Cancer. 10:194–204. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Koudelka S and Turánek J: Liposomal
paclitaxel formulations. J Control Release. 163:322–334. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan P, Shentu J, Xu J, Burke W, Hsu K,
Learoyd M, Zhu M and Xu B: Pharmacokinetics and safety of olaparib
tablets as monotherapy and in combination with paclitaxel: Results
of a Phase I study in Chinese patients with advanced solid tumours.
Cancer Chemother Pharmacol. 83:963–974. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Azimi I, Petersen RM, Thompson EW,
Roberts-Thomson SJ and Monteith GR: Hypoxia-induced reactive oxygen
species mediate N-cadherin and SERPINE1 expression, EGFR signalling
and motility in MDA-MB-468 breast cancer cells. Sci Rep.
7:151402017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Freeberg MAT, Farhat YM, Easa A,
Kallenbach JG, Malcolm DW, Buckley MR, Benoit DSW and Awad HA:
Serpine1 knockdown enhances MMP activity after flexor tendon injury
in mice: Implications for adhesions therapy. Sci Rep. 8:58102018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang K, Zhang S, Zhang D, Tao Q, Zhang T,
Liu G, Liu X and Zhao T: Identification of SERPINE1, PLAU and ACTA1
as biomarkers of head and neck squamous cell carcinoma based on
integrated bioinformatics analysis. Int J Clin Oncol. 24:1030–1041.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41((Database Issue)):
D991–D995. 2013.PubMed/NCBI
|
|
15
|
The Gene Ontology Consortium, . The gene
ontology resource: 20 Years and still GOing strong. Nucleic Acids
Res. 47(D1): D330–D338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45(D1): D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lei Y, Yu D, Bin Z and Yang Y: Interactive
K-means clustering method based on user behavior for different
analysis target in medicine. Comput Math Methods Med.
2017:49158282017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang TN, Li TJ, Shao GF and Wu SX: An
improved K-means clustering method for cDNA microarray image
segmentation. Genet Mol Res. 14:7771–7781. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen S, Dong Q, Hu S, Cai J, Zhang W, Sun
J, Wang T, Xie J, He H, Xing J, et al: Proteomic analysis of the
proteins that are associated with the resistance to paclitaxel in
human breast cancer cells. Mol Biosyst. 10:294–303. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Panayotopoulou EG, Müller AK, Börries M,
Busch H, Hu G and Lev S: Targeting of apoptotic pathways by SMAC or
BH3 mimetics distinctly sensitizes paclitaxel-resistant triple
negative breast cancer cells. Oncotarget. 8:45088–45104. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Edilova MI, Abdul-Sater AA and Watts TH:
TRAF1 signaling in human health and disease. Front Immunol.
9:29692018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu S, Jin J, Gokhale S, Lu AM, Shan H,
Feng J and Xie P: Genetic alterations of TRAF proteins in human
cancers. Front Immunol. 9:21112018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dewangan J, Srivastava S, Mishra S,
Divakar A, Kumar S and Rath SK: Salinomycin inhibits breast cancer
progression via targeting HIF-1α/VEGF mediated tumor angiogenesis
in vitro and in vivo. Biochem Pharmacol. 164:326–335. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jia J, Zhang H, Zhang H, Du H, Liu W and
Shu M: Activated androgen receptor accelerates angiogenesis in
cutaneous neurofibroma by regulating VEGFA transcription. Int J
Oncol. 55:157–166. 2019.PubMed/NCBI
|
|
26
|
Alluri P and Newman LA: Basal-like and
triple-negative breast cancers: Searching for positives among many
negatives. Surg Oncol Clin N Am. 23:567–577. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carotenuto P, Roma C, Rachiglio AM, Botti
G, D'Alessio A and Normanno N: Triple negative breast cancer: From
molecular portrait to therapeutic intervention. Crit Rev Eukaryot
Gene Expr. 20:17–34. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Foulkes WD, Smith IE and Reis-Filho JS:
Triple-negative breast cancer. N Engl J Med. 363:1938–1948. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kalimutho M, Parsons K, Mittal D, López
JA, Srihari S and Khanna KK: Targeted therapies for triple-negative
breast cancer: Combating a stubborn disease. Trends Pharmacol Sci.
36:822–846. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Burstein MD, Tsimelzon A, Poage GM,
Covington KR, Contreras A, Fuqua SA, Savage MI, Osborne CK,
Hilsenbeck SG, Chang JC, et al: Comprehensive genomic analysis
identifies novel subtypes and targets of triple-negative breast
cancer. Clin Cancer Res. 21:1688–1698. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jhan JR and Andrechek ER: Triple-negative
breast cancer and the potential for targeted therapy.
Pharmacogenomics. 18:1595–1609. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lehmann BD, Bauer JA, Chen X, Sanders ME,
Chakravarthy AB, Shyr Y and Pietenpol JA: Identification of human
triple-negative breast cancer subtypes and preclinical models for
selection of targeted therapies. J Clin Invest. 121:2750–2767.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ng CK, Schultheis AM, Bidard FC, Weigelt B
and Reis-Filho JS: Breast cancer genomics from microarrays to
massively parallel sequencing: Paradigms and new insights. J Natl
Cancer Inst. 107:djv0152015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Podo F, Buydens LM, Degani H, Hilhorst R,
Klipp E, Gribbestad IS, Van Huffel S, van Laarhoven HW, Luts J,
Monleon D, et al: Triple-negative breast cancer: Present challenges
and new perspectives. Mol Oncol. 4:209–229. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shah SP, Roth A, Goya R, Oloumi A, Ha G,
Zhao Y, Turashvili G, Ding J, Tse K, Haffari G, et al: The clonal
and mutational evolution spectrum of primary triple-negative breast
cancers. Nature. 486:395–399. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gardner ER, Dahut WL, Scripture CD, Jones
J, Aragon-Ching JB, Desai N, Hawkins MJ, Sparreboom A and Figg WD:
Randomized crossover pharmacokinetic study of solvent-based
paclitaxel and nab-paclitaxel. Clin Cancer Res. 14:4200–4205. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schiff PB and Horwitz SB: Taxol stabilizes
microtubules in mouse fibroblast cells. Proc Natl Acad Sci USA.
77:1561–1565. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
ten Tije AJ, Verweij J, Loos WJ and
Sparreboom A: Pharmacological effects of formulation vehicles:
Implications for cancer chemotherapy. Clin Pharmacokinet.
42:665–685. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Menderes G, Lopez S, Han C, Altwerger G,
Gysler S, Varughese J, Schwartz PE and Santin AD: Mechanisms of
resistance to HER2-targeted therapies in HER2-amplified uterine
serous carcinoma, and strategies to overcome it. Discov Med.
26:39–50. 2018.PubMed/NCBI
|
|
40
|
Leik CE, Su EJ, Nambi P, Crandall DL and
Lawrence DA: Effect of pharmacologic plasminogen activator
inhibitor-1 inhibition on cell motility and tumor angiogenesis. J
Thromb Haemost. 4:2710–2715. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Erickson LA, Ginsberg MH and Loskutoff DJ:
Detection and partial characterization of an inhibitor of
plasminogen activator in human platelets. J Clin Invest.
74:1465–1472. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ghosh AK, Murphy SB, Kishore R and Vaughan
DE: Global gene expression profiling in PAI-1 knockout murine heart
and kidney: Molecular basis of cardiac-selective fibrosis. PLoS
One. 8:e638252013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Higgins PJ: Balancing AhR-dependent
pro-oxidant and Nrf2-responsive anti-oxidant pathways in
age-related retinopathy: Is SERPINE1 expression a therapeutic
target in disease onset and progression? J Mol Genet Med.
8:1012014.PubMed/NCBI
|
|
44
|
Freytag J, Wilkins-Port CE, Higgins CE,
Higgins SP, Samarakoon R and Higgins PJ: PAI-1 mediates the
TGF-beta1+EGF-induced ‘scatter’ response in transformed human
keratinocytes. J Invest Dermatol. 130:2179–2190. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pavón MA, Arroyo-Solera I, Téllez-Gabriel
M, León X, Virós D, López M, Gallardo A, Céspedes MV, Casanova I,
López-Pousa A, et al: Enhanced cell migration and apoptosis
resistance may underlie the association between high SERPINE1
expression and poor outcome in head and neck carcinoma patients.
Oncotarget. 6:29016–29033. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pavón MA, Arroyo-Solera I, Céspedes MV,
Casanova I, León X and Mangues R: uPA/uPAR and SERPINE1 in head and
neck cancer: Role in tumor resistance, metastasis, prognosis and
therapy. Oncotarget. 7:57351–57366. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wu DM, Wang S, Wen X, Han XR, Wang YJ, Fan
SH, Zhang ZF, Shan Q, Lu J and Zheng YL: MircoRNA-1275 promotes
proliferation, invasion and migration of glioma cells via SERPINE1.
J Cell Mol Med. 22:4963–4974. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yao H, He G, Chen C, Yan S, Lu L, Song L,
Vijayan KV, Li Q, Xiong L, Miao X and Deng X: PAI1: A novel
PP1-interacting protein that mediates human plasma's anti-apoptotic
effect in endothelial cells. J Cell Mol Med. 21:2068–2076. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
McCann JV, Xiao L, Kim DJ, Khan OF,
Kowalski PS, Anderson DG, Pecot CV, Azam SH, Parker JS, Tsai YS, et
al: Endothelial miR-30c suppresses tumor growth via inhibition of
TGF-β-induced Serpine1. J Clin Invest. 130:1654–1670. 2019.
View Article : Google Scholar
|
|
50
|
Takayama Y, Hattori N, Hamada H, Masuda T,
Omori K, Akita S, Iwamoto H, Fujitaka K and Kohno N: Inhibition of
PAI-1 limits tumor angiogenesis regardless of angiogenic stimuli in
malignant pleural mesothelioma. Cancer Res. 76:3285–3294. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Viallard C and Larrivée B: Tumor
angiogenesis and vascular normalization: Alternative therapeutic
targets. Angiogenesis. 20:409–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Luan X, Guan YY, Lovell JF, Zhao M, Lu Q,
Liu YR, Liu HJ, Gao YG, Dong X, Yang SC, et al: Tumor priming using
metronomic chemotherapy with neovasculature-targeted,
nanoparticulate paclitaxel. Biomaterials. 95:60–73. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Samuel S, Fan F, Dang LH, Xia L, Gaur P
and Ellis LM: Intracrine vascular endothelial growth factor
signaling in survival and chemoresistance of human colorectal
cancer cells. Oncogene. 30:1205–1212. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Iqbal S and Lenz HJ: Integration of novel
agents in the treatment of colorectal cancer. Cancer Chemother
Pharmacol. 54 (Suppl 1):S32–S39. 2004.PubMed/NCBI
|
|
55
|
Fallah A, Sadeghinia A, Kahroba H, Samadi
A, Heidari HR, Bradaran B, Zeinali S and Molavi O: Therapeutic
targeting of angiogenesis molecular pathways in
angiogenesis-dependent diseases. Biomed Pharmacother. 110:775–785.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tsai JL, Lee YM, Pan CY and Lee AY: The
novel VEGF121- VEGF165 fusion attenuates angiogenesis and drug
resistance via targeting VEGFR2-HIF-1α-VEGF165/Lon signaling
through PI3K-AKT-mTOR pathway. Curr Cancer Drug Targets.
16:275–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Tonissi F, Lattanzio L, Merlano MC,
Infante L, Lo Nigro C and Garrone O: The effect of paclitaxel and
nab-paclitaxel in combination with anti-angiogenic therapy in
breast cancer cell lines. Invest New Drugs. 33:801–809. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|